Yak-Kong Soybean (Glycine max) Fermented by a Novel Pediococcus pentosaceus Inhibits the Oxidative Stress-Induced Monocyte–Endothelial Cell Adhesion

 , , ,
, , ,  and
and 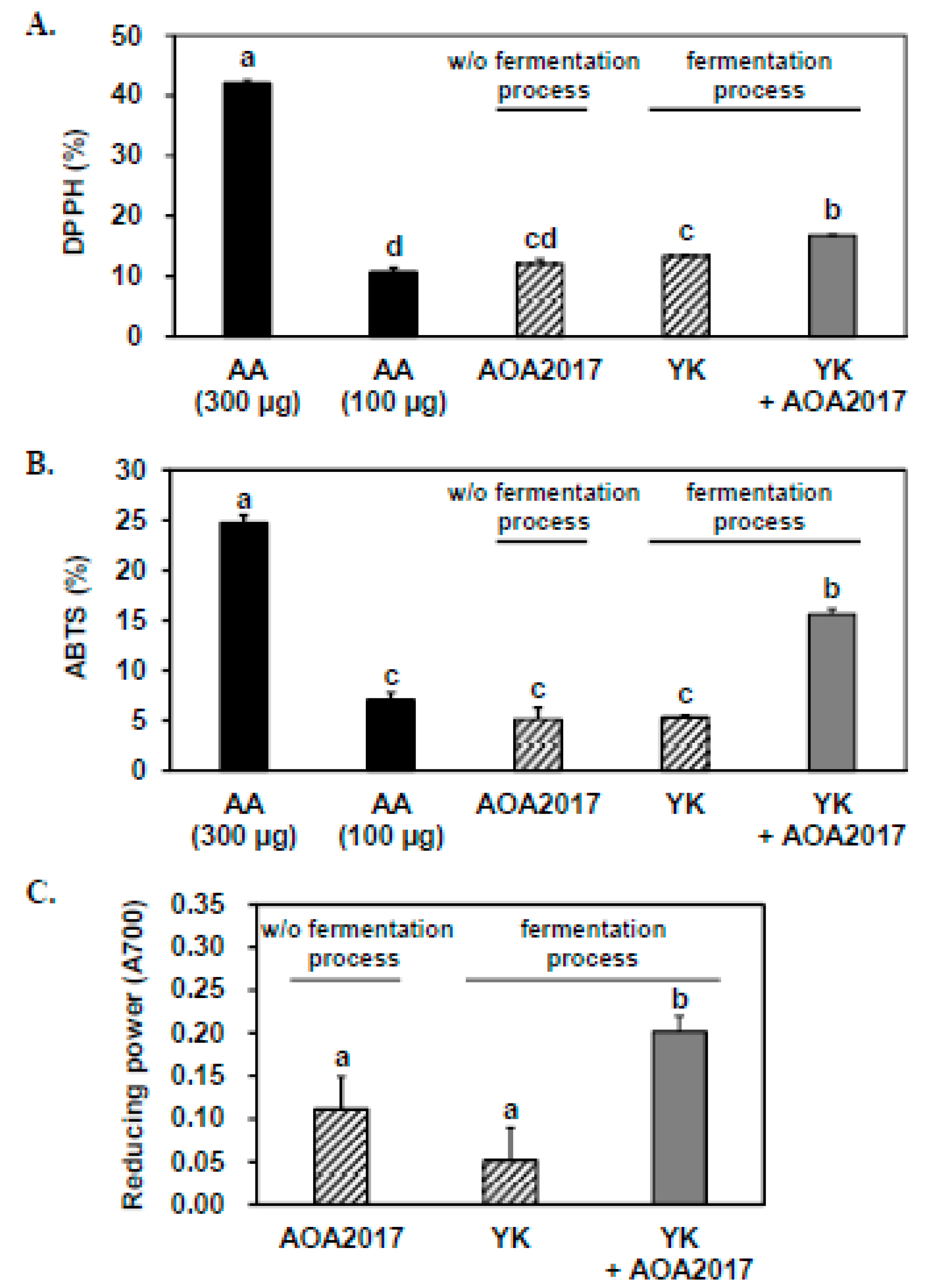{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Microorganism and Bean Extract Fermentation
2.3. Analysis of Antioxidant Activity
2.3.1. DPPH Free Radical Scavenging Activity
2.3.2. ABTS Free Radical Scavenging Activity
2.3.3. Reducing Power Assay
2.4. Analysis of Phenolic Composition
2.4.1. UHPLC–MS/MS Analysis
2.4.2. UHPLC–PDA Analysis
2.4.3. HPLC-Fluorescence Analysis
2.5. Cell Culture
2.6. THP-1 Monocyte Adhesion Assay
2.7. Western Blot Assay
2.8. Measurement of Intracellular ROS
2.9. Statistical Analysis
3. Results
3.1. Fermentation Using P. pentosaceus AOA2017 Increases the Antioxidant Activity of YK
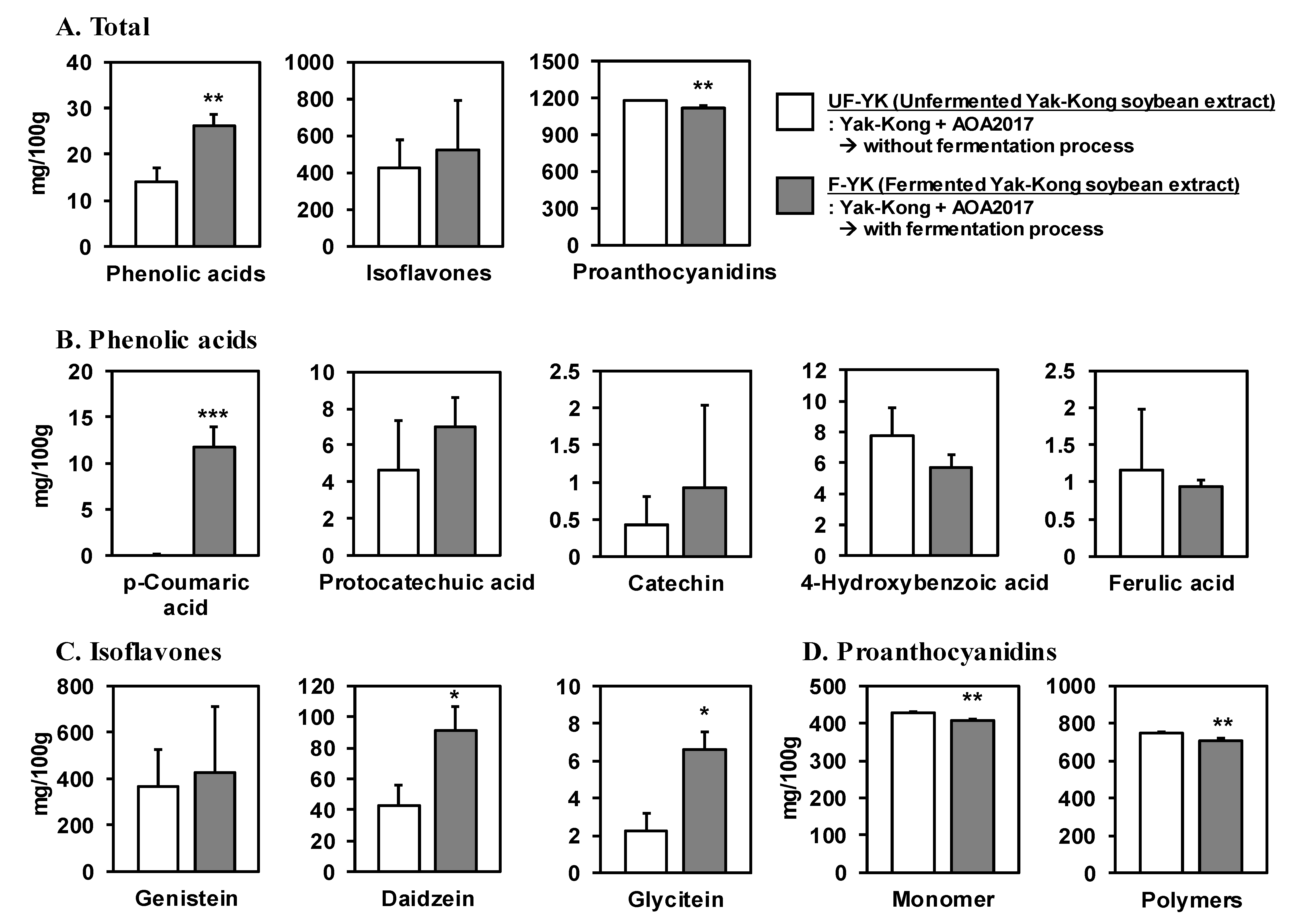
3.2. Fermentation of YK Using P. pentosaceus AOA2017 Increases the Content of Phenolic Acids and Decreases Proanthocyanidins
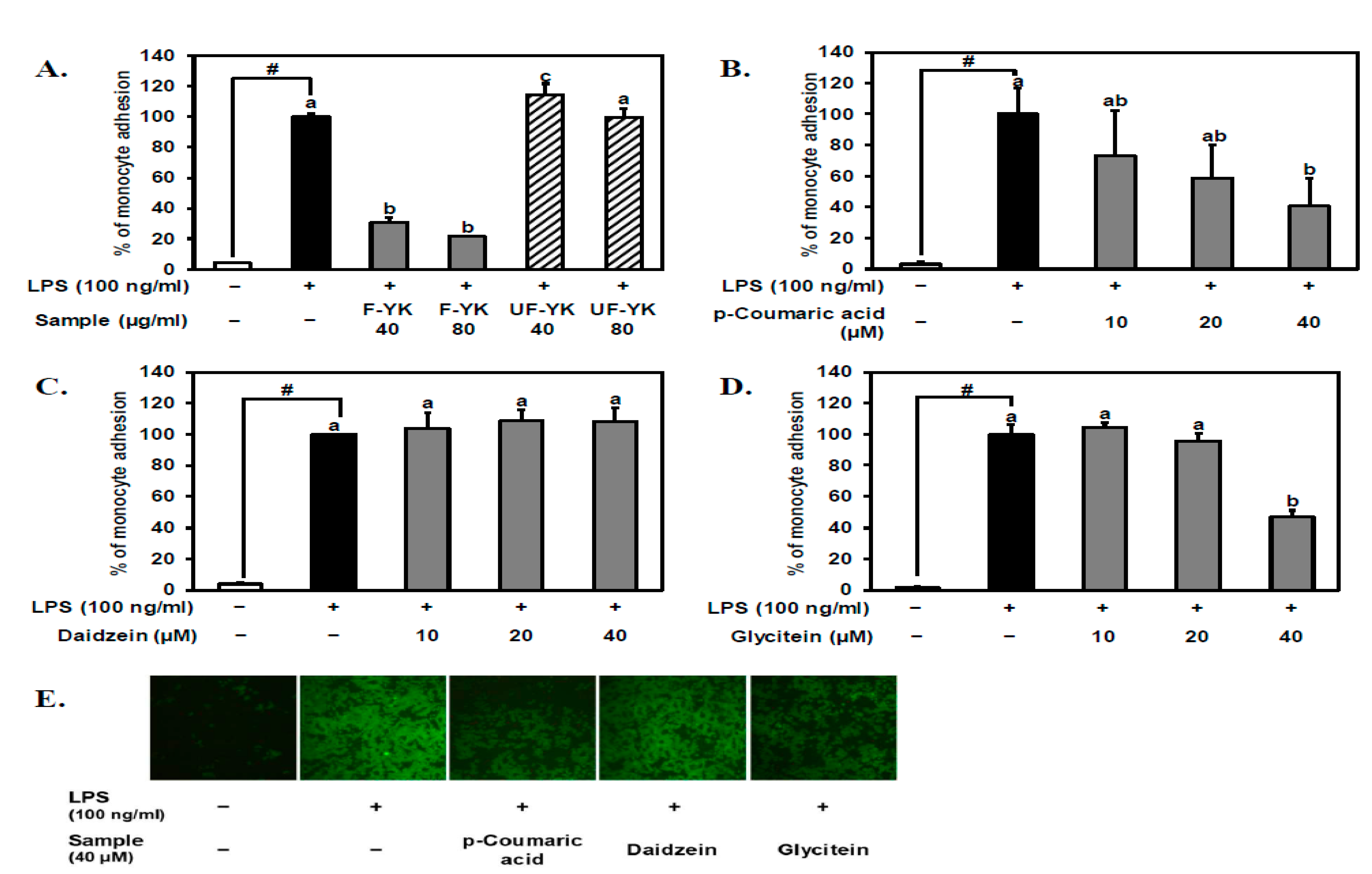
3.3. Fermented YK Extract and Its Bioactive Compound p-Coumaric Acid Have a Strong Inhibitory Effect on LPS-Stimulated THP-1 Monocyte–Endothelial Cell Adhesion
3.4. Fermented YK Extract and Its Bioactive Compound p-Coumaric Acid Decrease VCAM-1 Expression Levels in LPS-Stimulated Endothelial Cells
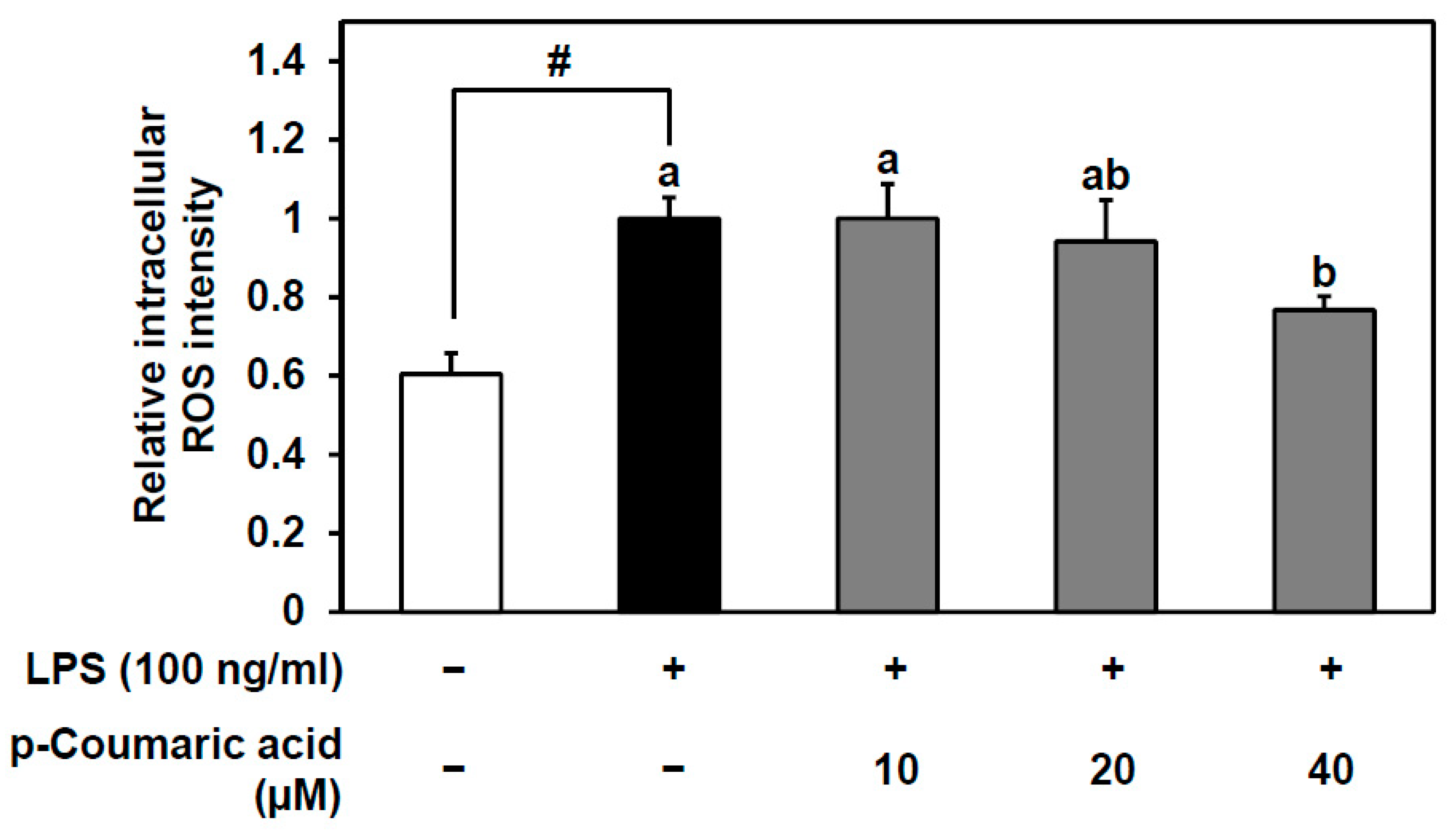
3.5. p-Coumaric Acid, the Bioactive Compound of Fermented YK Extract, Decreases the LPS-Stimulated ROS Production in Endothelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Tousoulis, D.; Psaltopoulou, T.; Androulakis, E.; Papageorgiou, N.; Papaioannou, S.; Oikonomou, E.; Synetos, A.; Stefanadis, C. Oxidative Stress and Early Atherosclerosis: Novel Antioxidant Treatment. Cardiovasc. Drugs Ther. 2015, 29, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.K.; Mehra, N.K.; Swarnakar, N.K. Role of Antioxidants for the Treatment of Cardiovascular Diseases: Challenges and Opportunities. Curr. Pharm. Des. 2015, 21, 4441–4455. [Google Scholar] [CrossRef] [PubMed]
- Marui, N.; Offermann, M.K.; Swerlick, R.; Kunsch, C.; Rosen, C.A.; Ahmad, M.; Alexander, R.W.; Medford, R.M. Vascular cell adhesion molecule-1 (VCAM-1) gene transcription and expression are regulated through an antioxidant-sensitive mechanism in human vascular endothelial cells. J. Clin. Investig. 1993, 92, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.A.; Berhow, M.; Singletary, K.W. Induction of macroautophagy in human colon cancer cells by soybean B-group triterpenoid saponins. Carcinogenesis 2005, 26, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Copeland, J.; Browning, J.D.; Sleper, D.; Rottinghaus, G.E.; Berhow, M.A.; Macdonald, R.S. Environmental Influences on Isoflavones and Saponins in Soybeans and Their Role in Colon Cancer. J. Nutr. 2005, 135, 1239–1242. [Google Scholar]
- Liao, H.-F.; Chen, Y.-J.; Yang, Y.-C. A novel polysaccharide of black soybean promotes myelopoiesis and reconstitutes bone marrow after 5-flurouracil- and irradiation-induced myelosuppression. Life Sci. 2005, 77, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Dudonné, S.; Dubé, P.; Desjardins, Y.; Kim, J.H.; Kim, J.S.; Kim, J.-E.; Park, J.H.Y.; Lee, K.W.; Lee, C.Y. Comprehensive phenolic composition analysis and evaluation of Yak-Kong soybean (Glycine max) for the prevention of atherosclerosis. Food Chem. 2017, 234, 486–493. [Google Scholar] [CrossRef]
- Lee, C.C.; Dudonné, S.; Kim, J.H.; Kim, J.S.; Dubé, P.; Kim, J.-E.; Desjardins, Y.; Park, J.H.Y.; Lee, K.W.; Lee, C.Y. A major daidzin metabolite 7,8,4′-trihydroxyisoflavone found in the plasma of soybean extract-fed rats attenuates monocyte-endothelial cell adhesion. Food Chem. 2018, 240, 607–614. [Google Scholar] [CrossRef]
- Bobe, G.; Barrett, K.G.; Mentor-Marcel, R.A.; Saffiotti, U.; Young, M.R.; Colburn, N.H.; Albert, P.S.; Bennink, M.R.; Lanza, E. Dietary Cooked Navy Beans and Their Fractions Attenuate Colon Carcinogenesis in Azoxymethane-InducedOb/ObMice. Nutr. Cancer 2008, 60, 373–381. [Google Scholar] [CrossRef]
- Anderson, J.W.; Bush, H.M. Soy Protein Effects on Serum Lipoproteins: A Quality Assessment and Meta-Analysis of Randomized, Controlled Studies. J. Am. Coll. Nutr. 2011, 30, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.-J. Optimization of roasting conditions through antioxidant and anti-inflammatory activities of Yak-kong (Rhynchosia nulubilis). Food Sci. Biotechnol. 2016, 25, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.H.; Lee, O.-H.; Choi, U.-K.; Kim, Y.-C. Antinociceptive and Anti-Inflammatory Effects of Ethanolic Extracts of Glycine max (L.) Merr and Rhynchosia nulubilis Seeds. Int. J. Mol. Sci. 2009, 10, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.-J.; Park, H.-S.; Kim, T.-H.; Shin, S.-R.; Hong, J.-Y.; Yang, K.-M. Analysis of Nutrition and Antioxidants of Yak-Kong Chungkukjang Powder Added Black Foods. J. Korean Soc. Food Sci. Nutr. 2013, 42, 1727–1735. [Google Scholar] [CrossRef]
- Chun, J.; Kim, J.S.; Kim, J.H. Enrichment of isoflavone aglycones in soymilk by fermentation with single and mixed cultures of Streptococcus infantarius 12 and Weissella sp. 4. Food Chem. 2008, 109, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, L.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enhancement of the antioxidant capacity of soy whey by fermentation with Lactobacillus plantarum B1–6. J. Funct. Foods 2015, 12, 33–44. [Google Scholar] [CrossRef]
- Lai, L.-R.; Hsieh, S.-C.; Huang, H.-Y.; Chou, C.-C. Effect of lactic fermentation on the total phenolic, saponin and phytic acid contents as well as anti-colon cancer cell proliferation activity of soymilk. J. Biosci. Bioeng. 2013, 115, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Rafi, M.M.; DiPaola, R.S.; Xin, J.; Chin, C.-K.; Badmaev, V.; Ghai, G.; Rosen, R.T.; Ho, C.-T. Bioactive constituents from gum guggul (Commiphora wightii). Phytochemistry 2001, 56, 723–727. [Google Scholar] [CrossRef]
- Biglari, F.; AlKarkhi, A.F.; Easa, A.M. Antioxidant activity and phenolic content of various date palm (Phoenix dactylifera) fruits from Iran. Food Chem. 2008, 107, 1636–1641. [Google Scholar] [CrossRef]
- Yen, G.C.; Der Duh, P. Scavenging Effect of Methanolic Extracts of Peanut Hulls on Free-Radical and Active-Oxygen Species. J. Agric. Food Chem. 1994, 42, 629–632. [Google Scholar] [CrossRef]
- Dudonné, S.; Dubé, P.; Pilon, G.; Marette, A.; Jacques, H.; Weisnagel, J.; Desjardins, Y. Modulation of strawberry/cranberry phenolic compounds glucuronidation by co-supplementation with onion: Characterization of phenolic metabolites in rat plasma using an optimized μSPE-UHPLC-MS/MS method. J. Agric. Food Chem. 2014, 62, 3244–3256. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Boil. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Déprez, S.; Brézillon, C.; Philippe, C.; Mila, I.; Scalbert, A.; Rabot, S.; Lapierre, C. Polymeric Proanthocyanidins Are Catabolized by Human Colonic Microflora into Low-Molecular-Weight Phenolic Acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar] [CrossRef] [PubMed]
- Bvochora, J.; Danner, H.; Miyafuji, H.; Braun, R.; Zvauya, R. Variation of sorghum phenolic compounds during the preparation of opaque beer. Process. Biochem. 2005, 40, 1207–1213. [Google Scholar] [CrossRef]
- Fuentes, E.; Palomo, I. Mechanisms of endothelial cell protection by hydroxycinnamic acids. Vasc. Pharmacol. 2014, 63, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Kildegaard, K.R.; Li, M.; Borodina, I.; Nielsen, J. Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis. Metab. Eng. 2015, 31, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Sakamula, R.; Thong-Asa, W. Neuroprotective effect of p-coumaric acid in mice with cerebral ischemia reperfusion injuries. Metab. Brain Dis. 2018, 33, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Pei, K.; Ou, J.; Huang, J. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-B.; Lee, S.; Hwang, E.-S.; Maeng, S.; Park, J.-H. p-Coumaric acid enhances long-term potentiation and recovers scopolamine-induced learning and memory impairments. Biochem. Biophys. Res. Commun. 2017, 492, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-W.; Kang, S.-I.; Shin, H.-S.; Yoon, S.-A.; Kim, J.-H.; Ko, H.-C.; Kim, S.-J. Sasa quelpaertensis Nakai extract and its constituent p-coumaric acid inhibit adipogenesis in 3T3-L1 cells through activation of the AMPK pathway. Food Chem. Toxicol. 2013, 59, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Amalan, V.; Vijayakumar, N.; Indumathi, D.; Ramakrishnan, A. Antidiabetic and antihyperlipidemic activity of p-coumaric acid in diabetic rats, role of pancreatic GLUT 2: In vivo approach. Biomed. Pharmacother. 2016, 84, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Calabriso, N.; Scoditti, E.; Massaro, M.; Pellegrino, M.; Storelli, C.; Ingrosso, I.; Giovinazzo, G.; Carluccio, M.A. Multiple anti-inflammatory and anti-atherosclerotic properties of red wine polyphenolic extracts: Differential role of hydroxycinnamic acids, flavonols and stilbenes on endothelial inflammatory gene expression. Eur. J. Nutr. 2016, 55, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S. Mechanisms of anti-atherosclerotic functions of soy-based diets. J. Nutr. Biochem. 2010, 21, 255–260. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, C.M.; de Sá, M.S.; Toloi, M.T. Effects of phytoestrogens derived from soy bean on expression of adhesion molecules on HUVEC. Climacteric 2012, 15, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial Dysfunction in Cardiovascular Diseases: The Role of Oxidant Stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef]
- Heitzer, T.; Schlinzig, T.; Krohn, K.; Meinertz, T.; Münzel, T. Endothelial Dysfunction, Oxidative Stress, and Risk of Cardiovascular Events in Patients with Coronary Artery Disease. Circulation 2001, 104, 2673–2678. [Google Scholar] [CrossRef]
- Fresquet, F.; Pourageaud, F.; Leblais, V.; Brandes, R.P.; Marthan, R.; Muller, B.; Savineau, J.-P.; Savineau, J. Role of reactive oxygen species and gp91phox in endothelial dysfunction of pulmonary arteries induced by chronic hypoxia. Br. J. Pharmacol. 2006, 148, 714–723. [Google Scholar] [CrossRef]
- Harrison, D.; Griendling, K.K.; Landmesser, U.; Hornig, B.; Drexler, H. Role of oxidative stress in atherosclerosis. Am. J. Cardiol. 2003, 91, 7–11. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Sun, W.; Li, W.; Hu, H.-Z.; Zhou, L.; Jiang, H.-H.; Xu, J.-X. Calycosin-7-O-β-d-glucoside promotes oxidative stress-induced cytoskeleton reorganization through integrin-linked kinase signaling pathway in vascular endothelial cells. BMC Complement. Altern. Med. 2015, 15, 315. [Google Scholar] [CrossRef][Green Version]
- Zhang, M.; Pan, H.; Xu, Y.; Wang, X.; Qiu, Z.; Jiang, L. Allicin Decreases Lipopolysaccharide-Induced Oxidative Stress and Inflammation in Human Umbilical Vein Endothelial Cells through Suppression of Mitochondrial Dysfunction and Activation of Nrf2. Cell. Physiol. Biochem. 2017, 41, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Mahoney, C.E.; Leopold, J.A.; Zhang, Y.-Y.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 modulates lipopolysaccharide-induced adhesion molecule expression in endothelial cells by altering CD14 expression. FASEB J. 2010, 24, 2525–2532. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J.; Ischiropoulos, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: Challenges and limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Kim, J.H.; Kim, J.S.; Oh, Y.S.; Han, S.M.; Park, J.H.Y.; Lee, K.W.; Lee, C.Y. 5-(3′,4′-Dihydroxyphenyl-γ-valerolactone), a Major Microbial Metabolite of Proanthocyanidin, Attenuates THP-1 Monocyte-Endothelial Adhesion. Int. J. Mol. Sci. 2017, 18, 1363. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.S.; Kim, J.H.; Palaniyandi, S.A.; Lee, C.C.; You, J.W.; Yang, H.; Yoon Park, J.H.; Yang, S.H.; Lee, K.W. Yak-Kong Soybean (Glycine max) Fermented by a Novel Pediococcus pentosaceus Inhibits the Oxidative Stress-Induced Monocyte–Endothelial Cell Adhesion. Nutrients 2019, 11, 1380. https://doi.org/10.3390/nu11061380
Kim JS, Kim JH, Palaniyandi SA, Lee CC, You JW, Yang H, Yoon Park JH, Yang SH, Lee KW. Yak-Kong Soybean (Glycine max) Fermented by a Novel Pediococcus pentosaceus Inhibits the Oxidative Stress-Induced Monocyte–Endothelial Cell Adhesion. Nutrients. 2019; 11(6):1380. https://doi.org/10.3390/nu11061380
Chicago/Turabian StyleKim, Ji Seung, Jong Hun Kim, Sasikumar Arunachalam Palaniyandi, Charles C. Lee, Ji Woo You, Hee Yang, Jung Han Yoon Park, Seung Hwan Yang, and Ki Won Lee. 2019. "Yak-Kong Soybean (Glycine max) Fermented by a Novel Pediococcus pentosaceus Inhibits the Oxidative Stress-Induced Monocyte–Endothelial Cell Adhesion" Nutrients 11, no. 6: 1380. https://doi.org/10.3390/nu11061380
APA StyleKim, J. S., Kim, J. H., Palaniyandi, S. A., Lee, C. C., You, J. W., Yang, H., Yoon Park, J. H., Yang, S. H., & Lee, K. W. (2019). Yak-Kong Soybean (Glycine max) Fermented by a Novel Pediococcus pentosaceus Inhibits the Oxidative Stress-Induced Monocyte–Endothelial Cell Adhesion. Nutrients, 11(6), 1380. https://doi.org/10.3390/nu11061380