The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Stocks and Maintenance
2.2. Chemicals and Reagents
2.3. Callus Culture
2.4. Liquid Chromatography
2.5. Lifespan Assay
2.6. Startle-Induced Negative Geotaxis Assay
2.7. Body Weight Measurement
2.8. Eye Imaging with Light Microscope
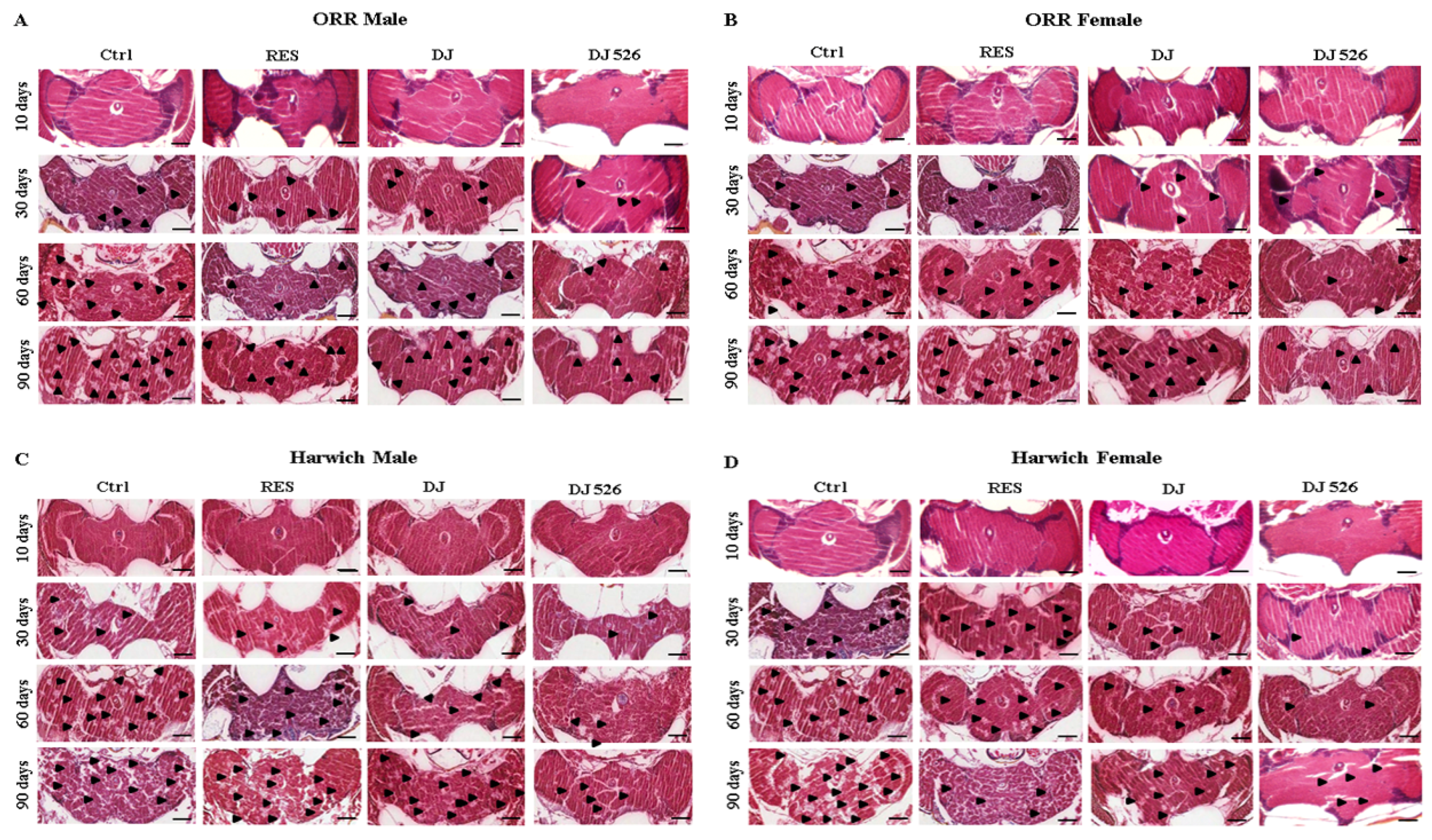
2.9. Histology
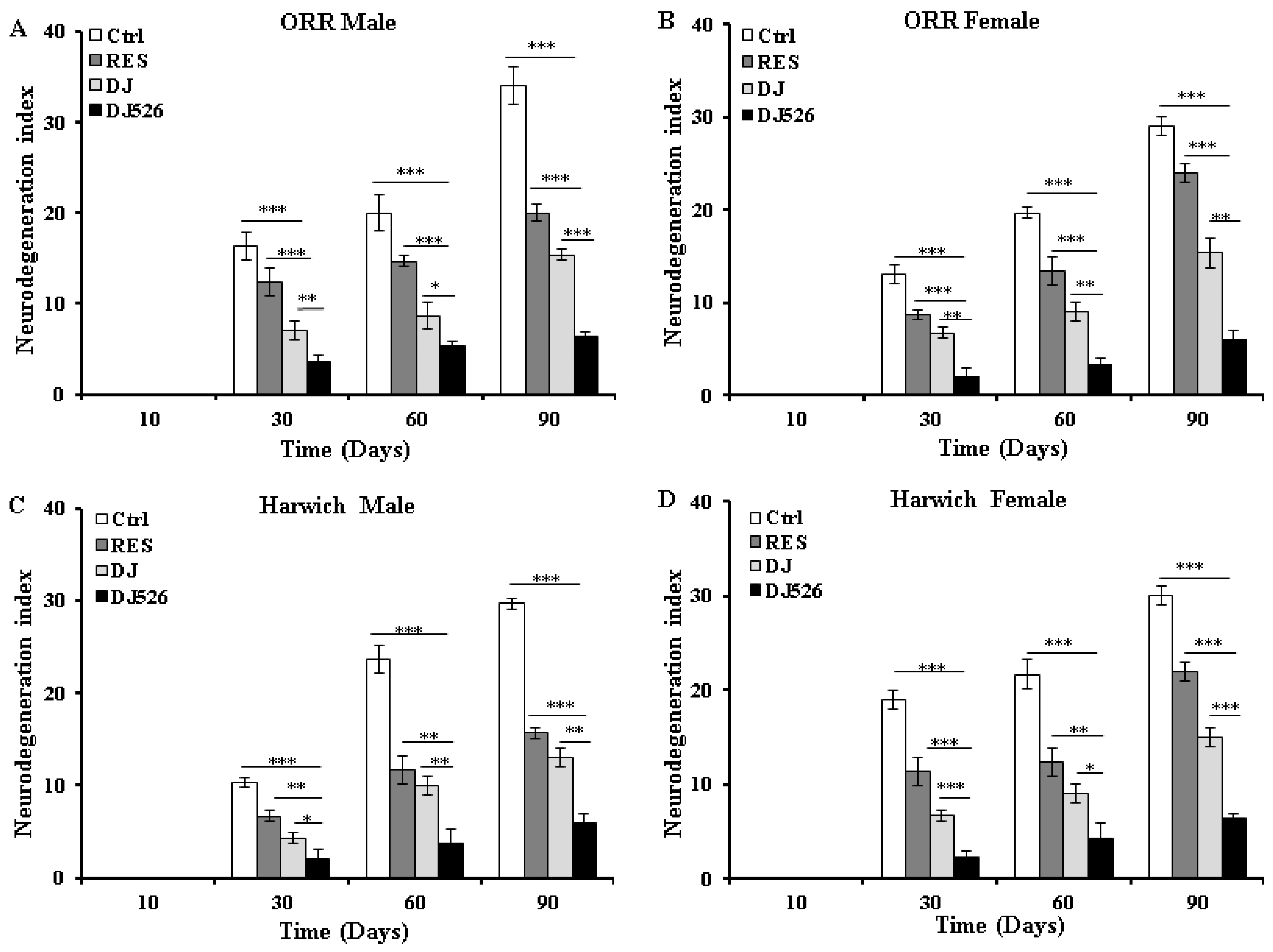
2.10. Scoring of Neurodegeneration
2.11. Statistical Analysis
3. Results
3.1. Resveratrol Rice DJ526C Producing a High Level of Resveratrol Was Obtained
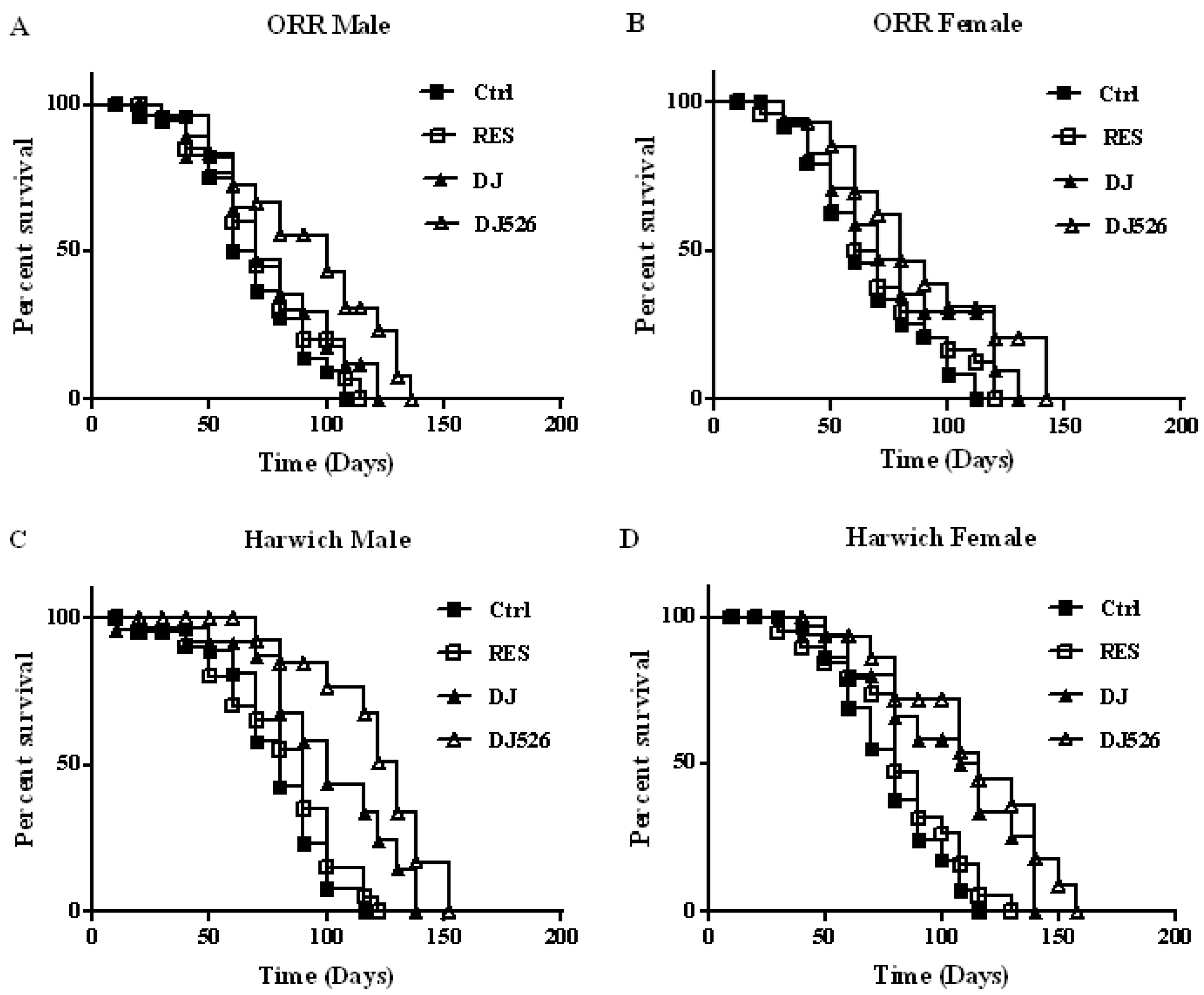
3.2. The Resveratrol Rice DJ526 Callus Significantly Extended the Median Lifespan of D. melanogaster
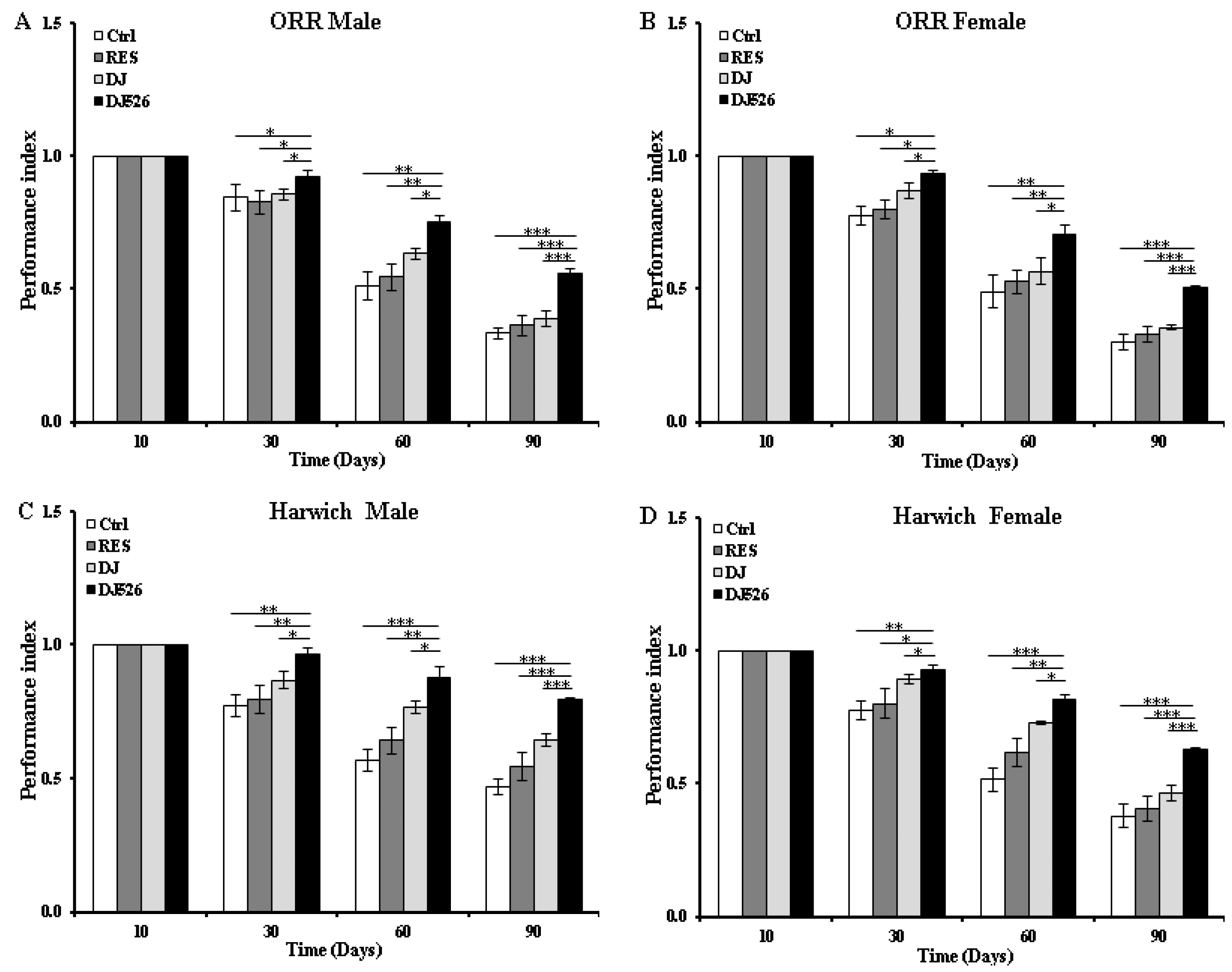
3.3. The Resveratrol Rice DJ526 Callus Ameliorated the Locomotor Deterioration of D. melanogaster during Age Progression
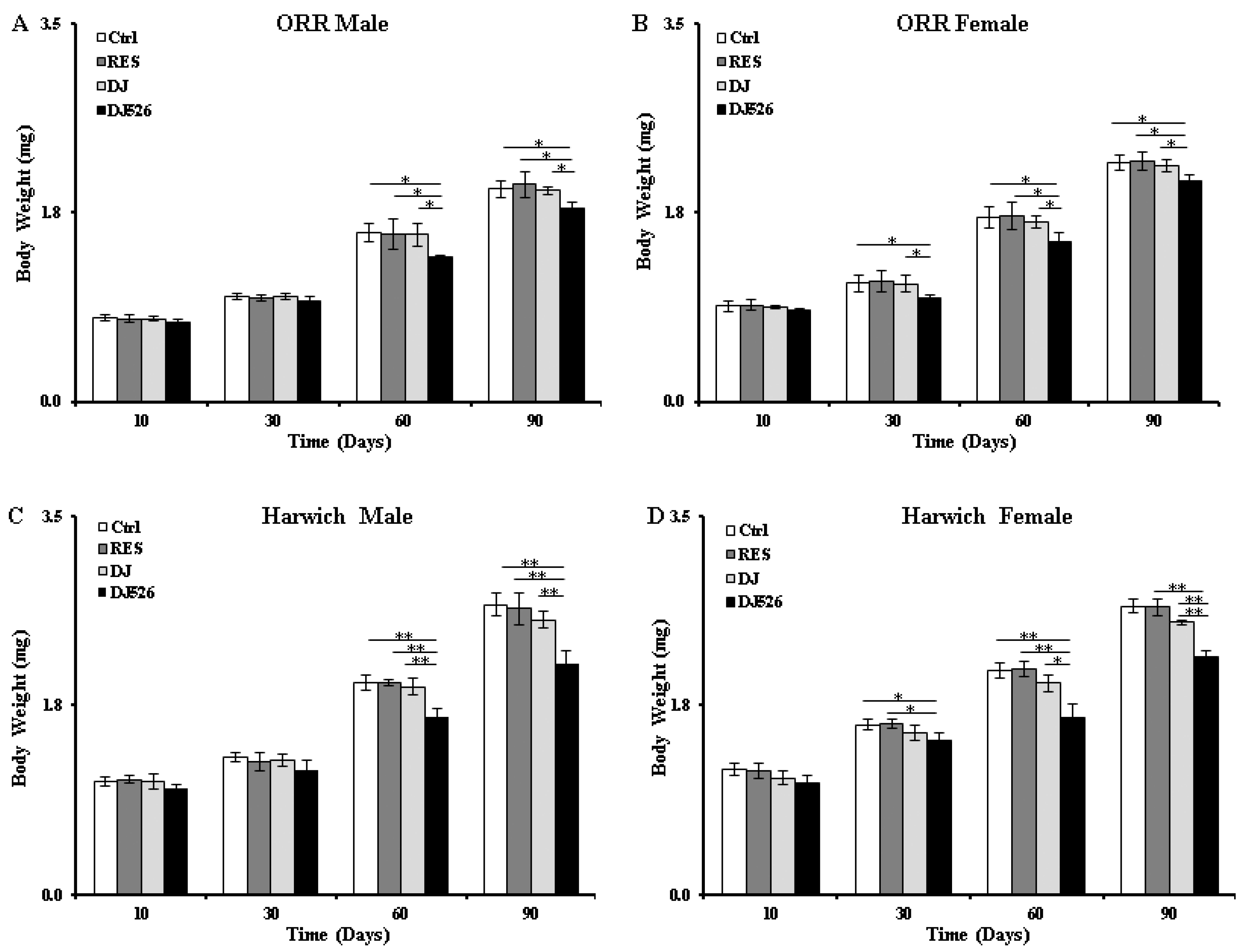
3.4. The Resveratrol Rice DJ526 Callus Maintained a Healthy Body Weight of D. melanogaster during Age Progression
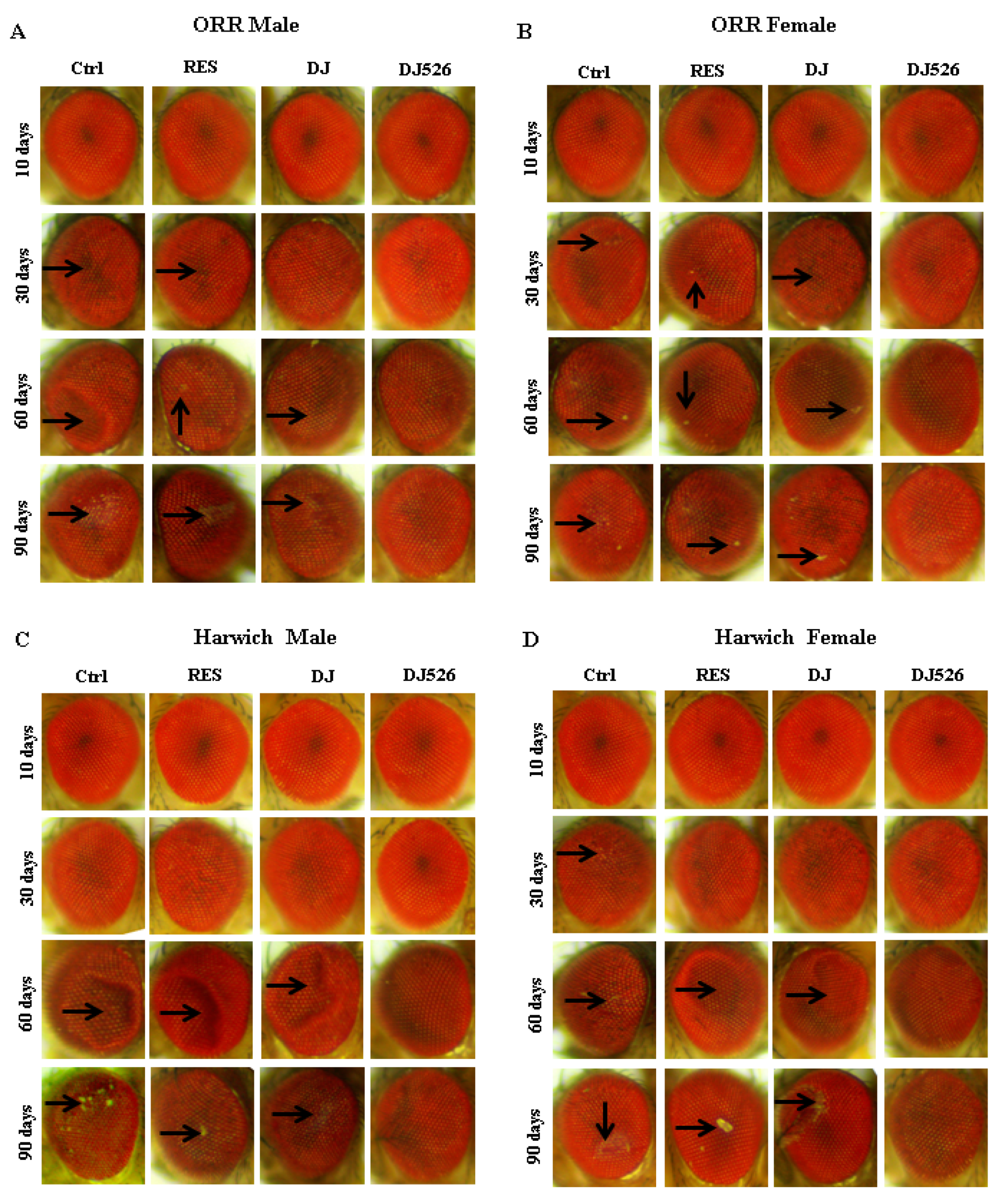
3.5. The Resveratrol-Enriched Rice DJ526 Callus Inhibited Eye Degeneration of D. melanogaster during Age Progression
3.6. The Resveratrol Rice DJ526 Callus Ameliorated Neurodegeneration of D. melanogaster during Age Progression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Vijg, J.; Campisi, J. Puzzles, promises and a cure for ageing. Nature 2008, 454, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T. A new definition of aging? Front. Genet. 2012, 3, 148. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. The aging process: Major risk factor for disease and death. Proc. Natl. Acad. Sci. USA 1991, 88, 5360–5363. [Google Scholar] [CrossRef] [PubMed]
- Fremont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Martin-Montalvo, A.; Mercken, E.M.; Mitchell, S.J.; Palacios, H.H.; Mote, P.L.; Scheibye-Knudsen, M.; Gomes, A.P.; Ward, T.M.; Minor, R.K.; Blouin, M.J.; et al. Metformin improves healthspan and lifespan in mice. Nat. Commun. 2013, 4, 2192–2200. [Google Scholar] [CrossRef]
- Morselli, E.; Galluzzi, L.; Kepp, O.; Criollo, A.; Maiuri, M.C.; Tavernarakis, N.; Madeo, F.; Kroemer, G. Autophagy mediates pharmacological lifespan extension by spermidine and resveratrol. Aging 2009, 1, 961–970. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Hubbard, B.P. Lifespan and healthspan extension by resveratrol. Biochem. Biophys. Acta 2015, 1852, 1209–1218. [Google Scholar] [CrossRef]
- Markus, M.A.; Morris, B.J. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin. Int. Aging 2008, 3, 331–339. [Google Scholar]
- Marchal, J.; Pifferi, F.; Aujard, F. Resveratrol in mammals: Effects on aging biomarkers, age-related diseases, and life span. Ann. N. Y. Acad. Sci. 2013, 1290, 67–73. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef]
- Bass, T.M.; Weinkove, D.; Houthoofd, K.; Gems, D.; Partridge, L. Effects of resveratrol on lifespan in Drosophila melanogaster and Caenorhabditis elegans. Mech. Ageing Dev. 2007, 128, 546–552. [Google Scholar] [CrossRef]
- Staats, S.; Wagner, A.E.; Kowalewski, B.; Rieck, F.T.; Soukup, S.T.; Kulling, S.E.; Rimbach, G. Dietary resveratrol does not affect life span, body composition, stress response, and longevity-related gene expression in drosophila melanogaster. Int. J. Mol. Sci. 2018, 19, 223. [Google Scholar] [CrossRef]
- Valenzano, D.R.; Terzibasi, E.; Genade, T.; Cattaneo, A.; Domenici, L.; Cellerino, A. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short-lived vertebrate. Curr. Biol. 2006, 16, 296–300. [Google Scholar] [CrossRef]
- Yu, X.; Li, G. Effects of resveratrol on longevity, cognitive ability and aging-related histological markers in the annual fish nothobranchius guentheri. Exp. Gerontol. 2012, 47, 940–949. [Google Scholar] [CrossRef]
- Genade, T.; Lang, D.M. Resveratrol extends lifespan and preserves glia but not neurons of the nothobranchius guentheri optic tectum. Exp. Gerontol. 2013, 48, 202–212. [Google Scholar] [CrossRef]
- Kim, E.; Ansell, C.M.; Dudycha, J.L. Resveratrol and food effects on lifespan and reproduction in the model crustacean daphnia. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2014, 321, 48–56. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell. Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Shin, W.C.; Ryu, H.S.; Lee, D.W.; Moon, E.; Seo, C.S.; Hwang, E.; Lee, H.S.; Ahn, M.H.; Jeon, Y.; et al. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE 2013, 8, e57930. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Chung, H.J.; Lee, H.K.; D’Souza, R.; Jeon, Y.; Kim, H.J.; Kweon, S.J.; Hong, S.T. Treatment of obesity with the resveratrol-enriched rice dj-526. Sci. Rep. 2014, 4, 3879. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Sharma, S.P.; Kim, H.J.; Baek, S.H.; Hong, S.T. The resveratrol-enriched rice dj526 boosts motor coordination and physical strength. Sci. Rep. 2016, 6, 23958. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Lee, H.K.; Kim, H.J.; Baek, S.H.; Hong, S.T. Gene expression profiles and physiological data from mice fed resveratrol-enriched rice DJ526. Sci. Data 2016, 3, 160114. [Google Scholar] [CrossRef] [PubMed]
- White, K.E.; Humphrey, D.M.; Hirth, F. The dopaminergic system in the aging brain of Drosophila. Front. Neurosci. 2010, 4, 205. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.C.; Bohmann, D. Assessing neurodegenerative phenotypes in drosophila dopaminergic neurons by climbing assays and whole brain immunostaining. J. Vis. Exp. 2013, e50339. [Google Scholar] [CrossRef]
- McPhee, J.S.; French, D.P.; Jackson, D.; Nazroo, J.; Pendleton, N.; Degens, H. Physical activity in older age: Perspectives for healthy ageing and frailty. Biogerontology 2016, 17, 567–580. [Google Scholar] [CrossRef]
- Cutler, T.; Sarkar, A.; Moran, M.; Steffensmeier, A.; Puli, O.R.; Mancini, G.; Tare, M.; Gogia, N.; Singh, A. Drosophila eye model to study neuroprotective role of creb binding protein (cbp) in alzheimer’s disease. PLoS ONE 2015, 10, e0137691. [Google Scholar] [CrossRef]
- Kounatidis, I.; Chtarbanova, S.; Cao, Y.; Hayne, M.; Jayanth, D.; Ganetzky, B.; Ligoxygakis, P. Nf-κb immunity in the brain determines fly lifespan in healthy aging and age-related neurodegeneration. Cell. Rep. 2017, 19, 836–848. [Google Scholar] [CrossRef]
- Bitterman, J.L.; Chung, J.H. Metabolic effects of resveratrol: Addressing the controversies. Cell. Mol. Life Sci. 2015, 72, 1473–1488. [Google Scholar]
- Kaeberlein, M.; McDonagh, T.; Heltweg, B.; Hixon, J.; Westman, E.A.; Caldwell, S.D.; Napper, A.; Curtis, R.; DiStefano, P.S.; Fields, S.; Bedalov, A.; et al. Substrate-specific activation of sirtuins by resveratrol. J. Biol. Chem. 2005, 280, 17038–17045. [Google Scholar] [CrossRef]
- Guarente, L.; Picard, F. Calorie restriction-the SIR2 connection. Cell 2005, 120, 473–482. [Google Scholar] [CrossRef]
- Liu, J.; Lkhagva, E.; Chung, H.J.; Kim, H.J.; Hong, S.T. The pharmabiotic approach to treat hyperammonemia. Nutrients 2018, 10, 140. [Google Scholar] [CrossRef]
- Chung, H.J.; Nguyen, T.; Kim, H.J.; Hong, S.T. Gut Microbiota as a missing link between nutrients and traits of human. Front. Microbiol. 2018, 9, 1510. [Google Scholar] [CrossRef]
- Nguyen, T.T.B.; Jin, Y.Y.; Chung, H.J.; Hong, S.T. Pharmabiotics as an emerging medication for metabolic syndrome and its related diseases. Molecules 2017, 22, 1795. [Google Scholar] [CrossRef]
- Chung, H.J.; Yu, J.G.; Liu, M.J.; Shen, Y.F.; Sharma, S.P.; Jamal, M.A.; Yoo, J.H.; Kim, H.J.; Hong, S.T. Intestinal removal of free fatty acids from hosts by Lactobacilli for the treatment of obesity. FEBS Open Bio 2016, 6, 64–76. [Google Scholar] [CrossRef]
- Singh, P.; Chung, H.J.; Lee, I.A.; D’Souza, R.; Kim, H.J.; Hong, S.T. Elucidation of the anti-hyperammonemic mechanism of Lactobacillus amylovorus JBD401 by comparative genomic analysis. BMC Genom. 2018, 19, 292. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.; Park, S.; Kim, H.-J.; Lee, K.-J.; Kim, D.H.; Baek, S.-H.; Hong, S.-T. The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity). Nutrients 2019, 11, 983. https://doi.org/10.3390/nu11050983
Khan M, Park S, Kim H-J, Lee K-J, Kim DH, Baek S-H, Hong S-T. The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity). Nutrients. 2019; 11(5):983. https://doi.org/10.3390/nu11050983
Chicago/Turabian StyleKhan, Mousumee, Soyeon Park, Hyeon-Jin Kim, Kui-Jae Lee, Dea Heon Kim, So-Hyeon Baek, and Seong-Tshool Hong. 2019. "The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity)" Nutrients 11, no. 5: 983. https://doi.org/10.3390/nu11050983
APA StyleKhan, M., Park, S., Kim, H.-J., Lee, K.-J., Kim, D. H., Baek, S.-H., & Hong, S.-T. (2019). The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity). Nutrients, 11(5), 983. https://doi.org/10.3390/nu11050983