Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Treatments
2.3. Manufacturing Process of HLJG0701
2.4. AChE Activity Inhibition Assay
2.5. Morris Water Maze Task
2.6. Y-Maze Task
2.7. Passive Avoidance Task
2.8. AChE Activity and Contents of ACh in Brain Tissues
2.9. Statistical Analysis
3. Results
3.1. Analysis of HLJG0701
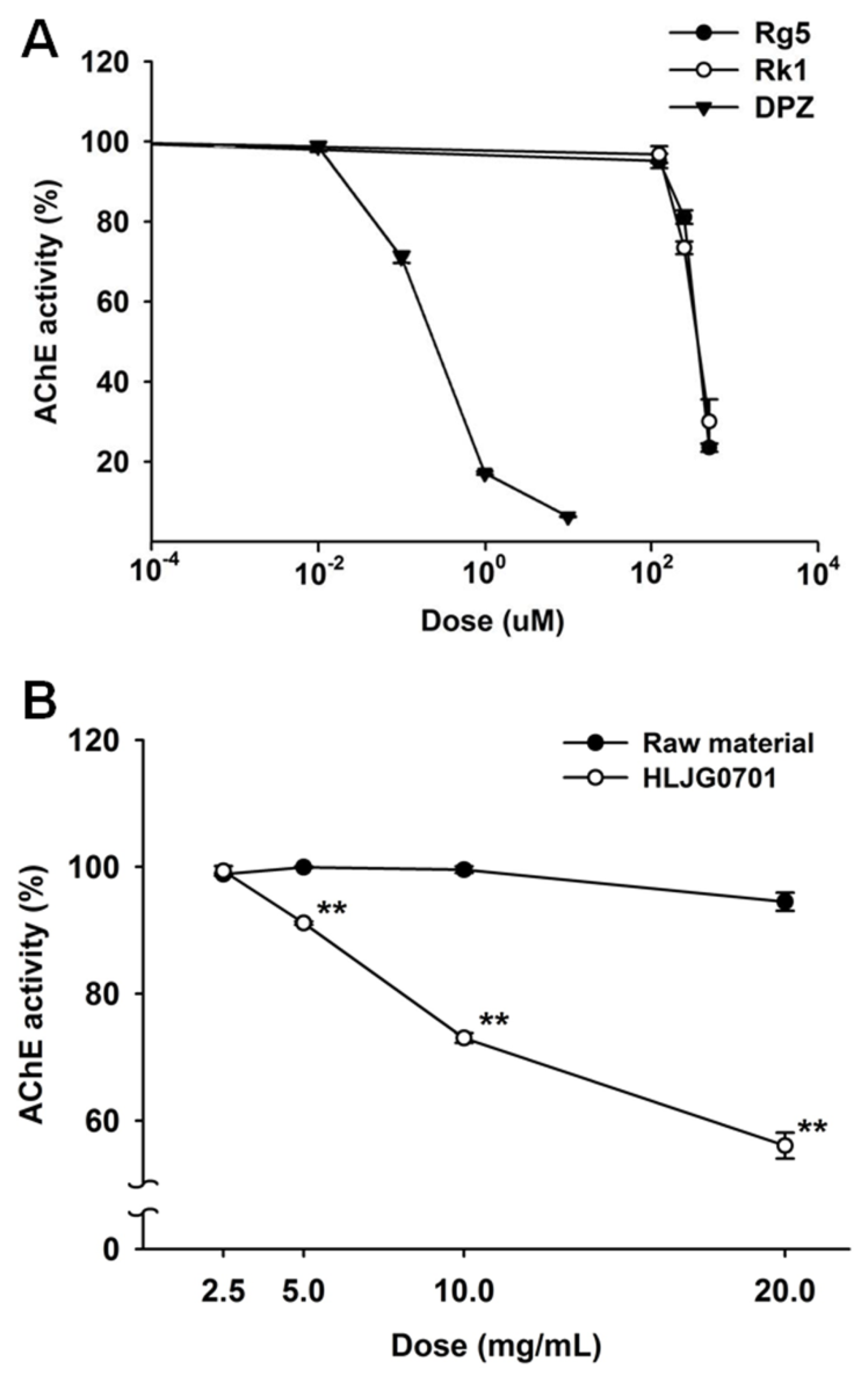
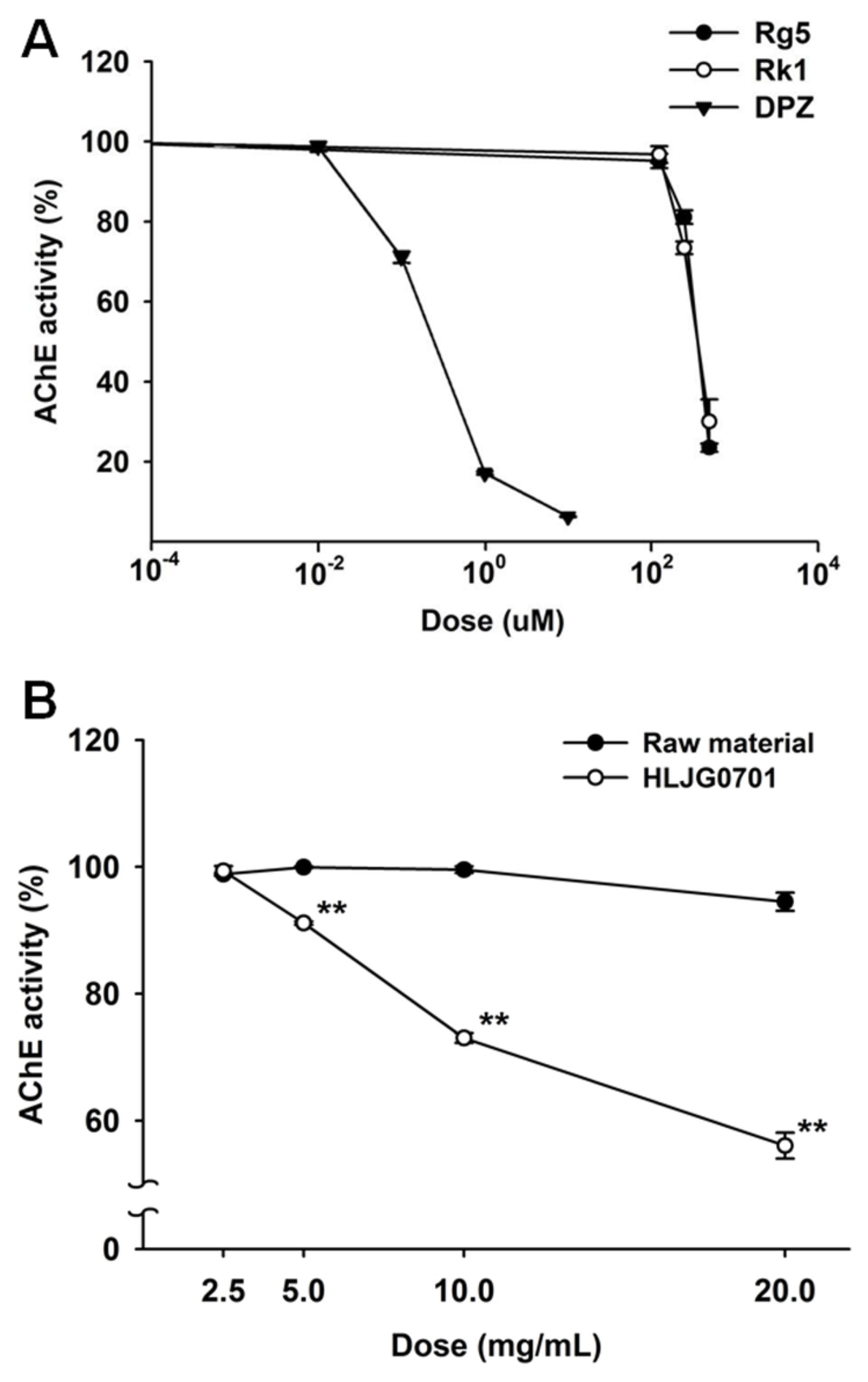
3.2. AChE Activity Inhibition Assay
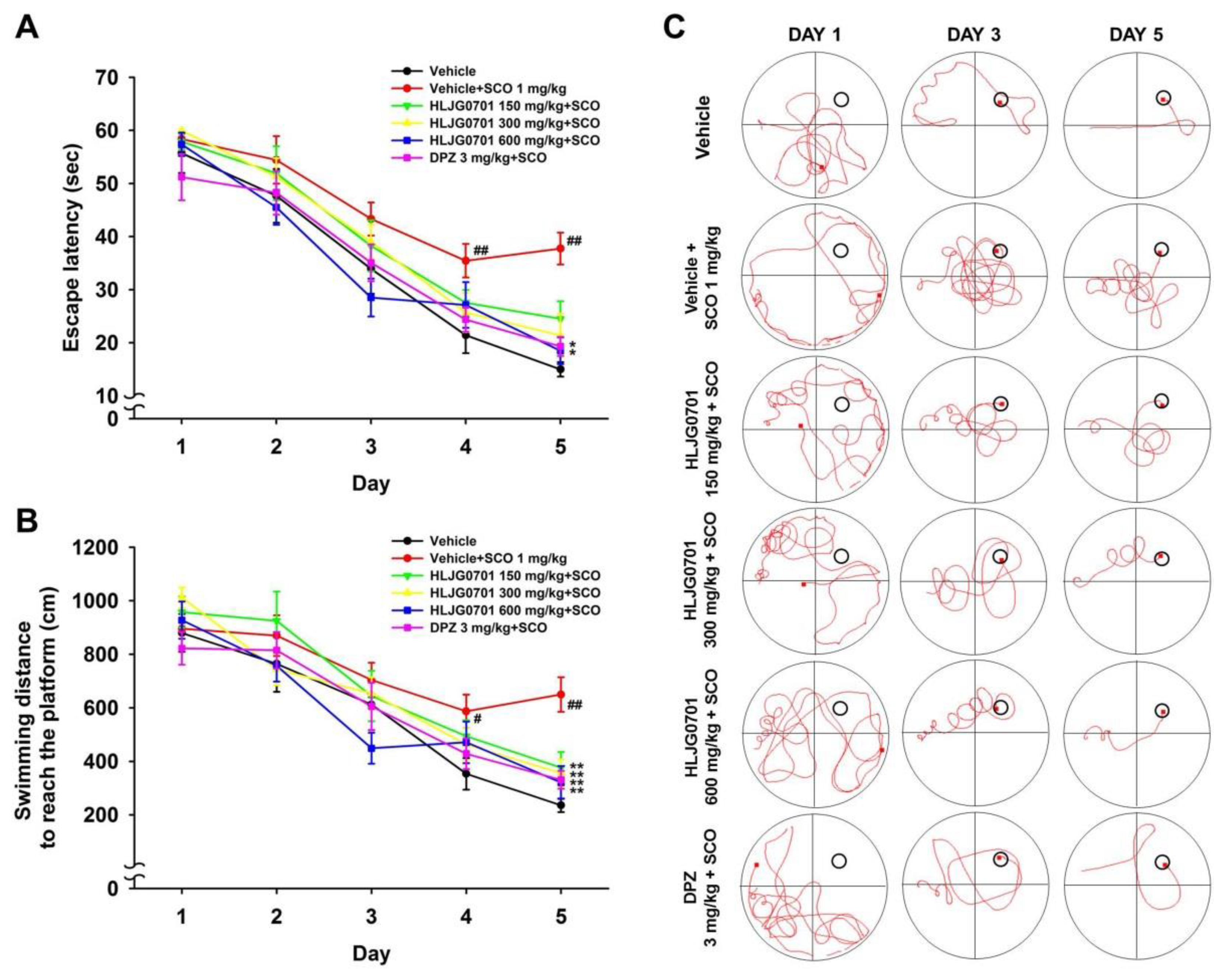
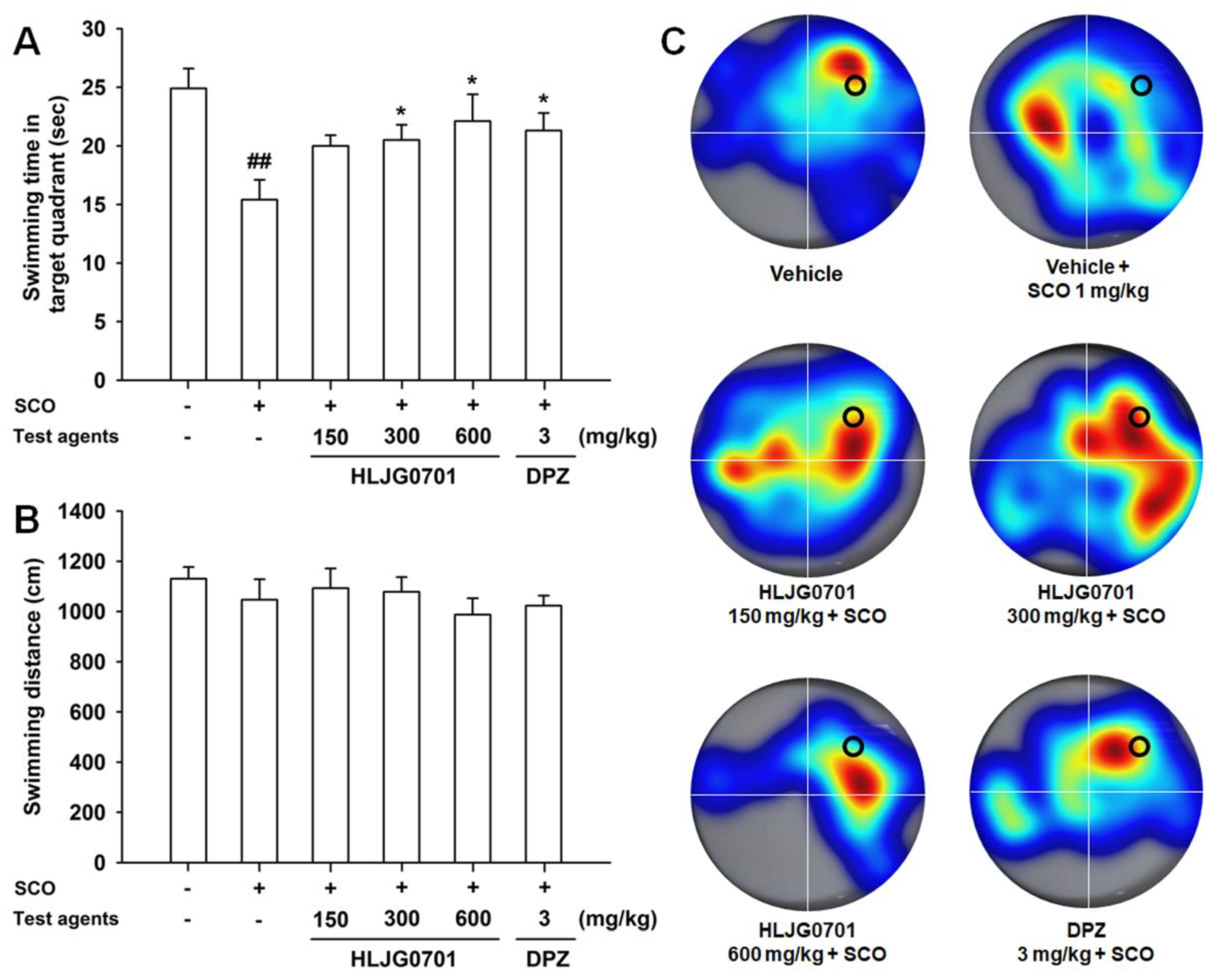
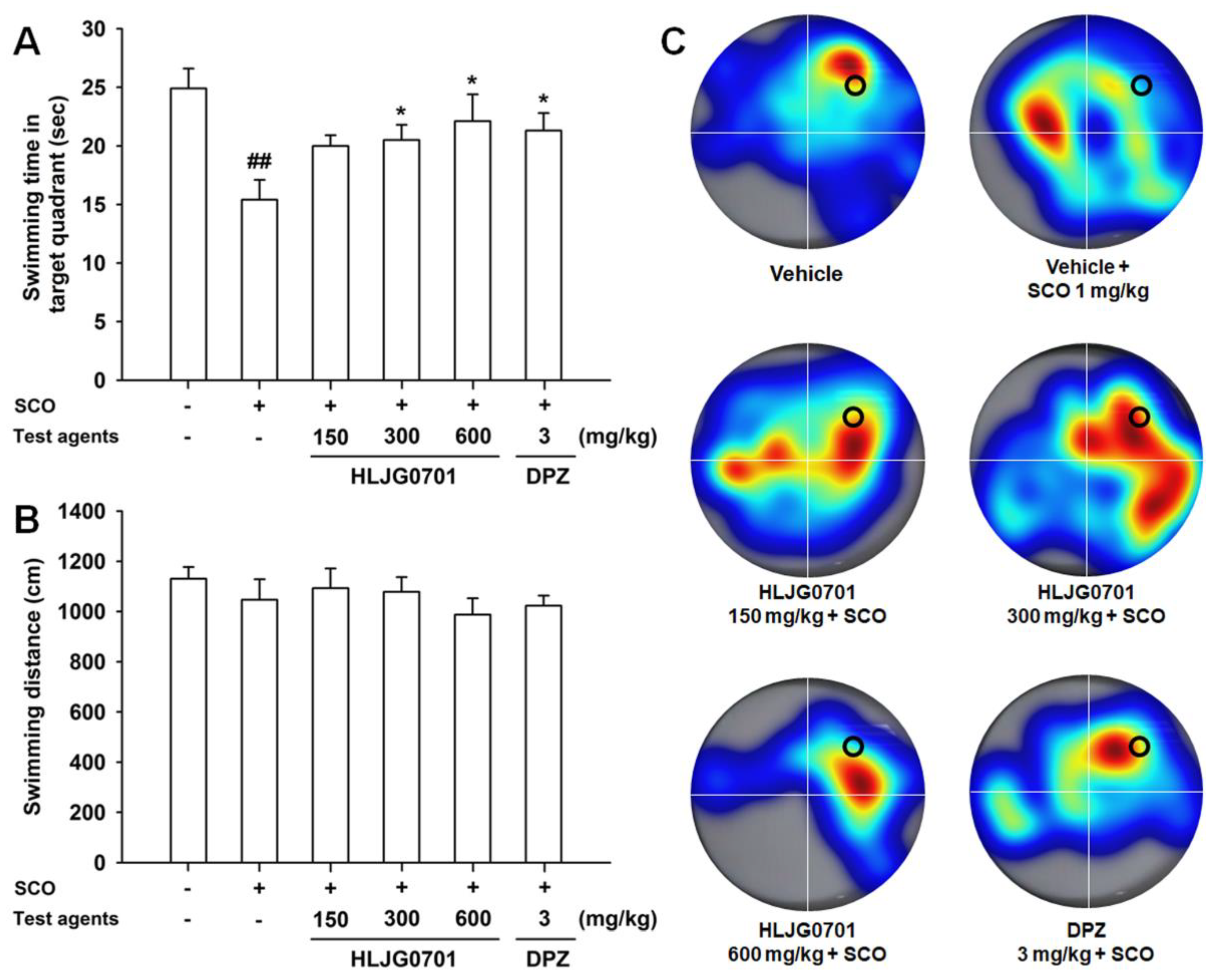
3.3. Morris Water Maze Task
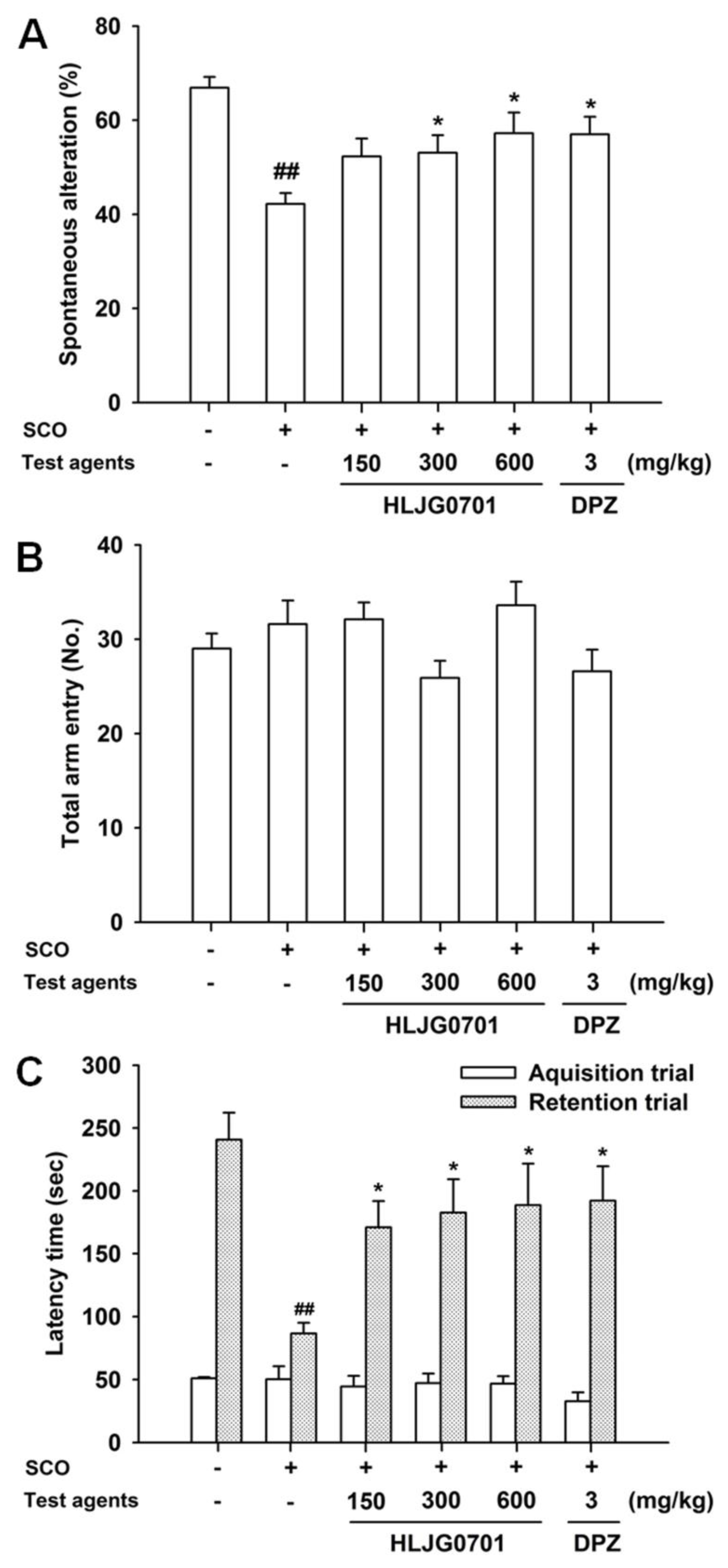
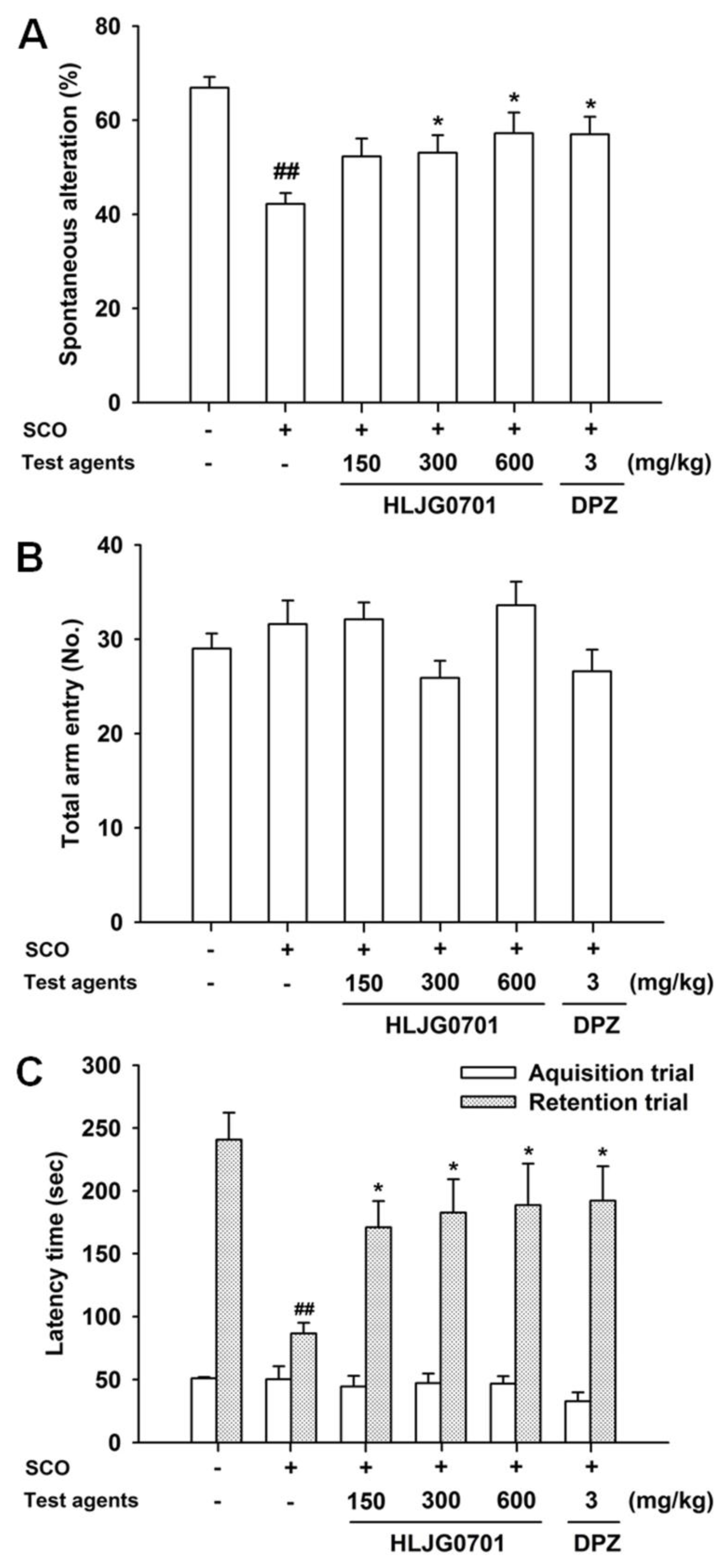
3.4. Y-Maze Task
3.5. Passive Avoidance Task
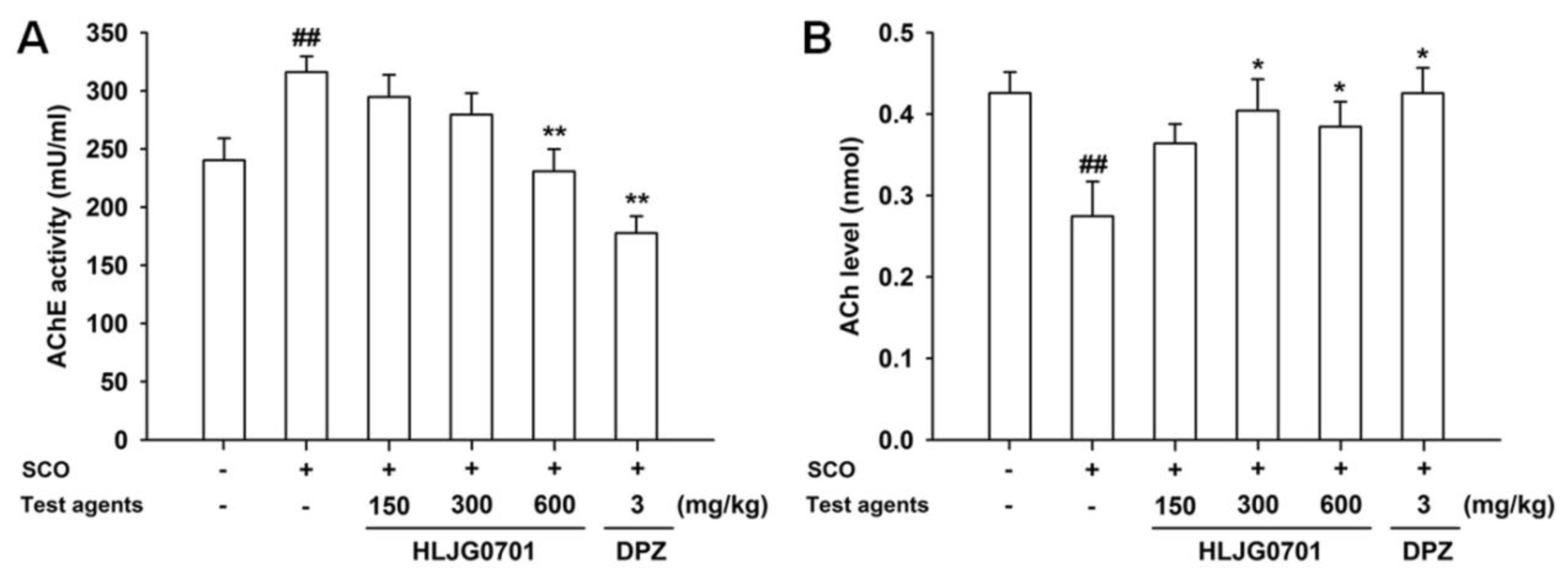
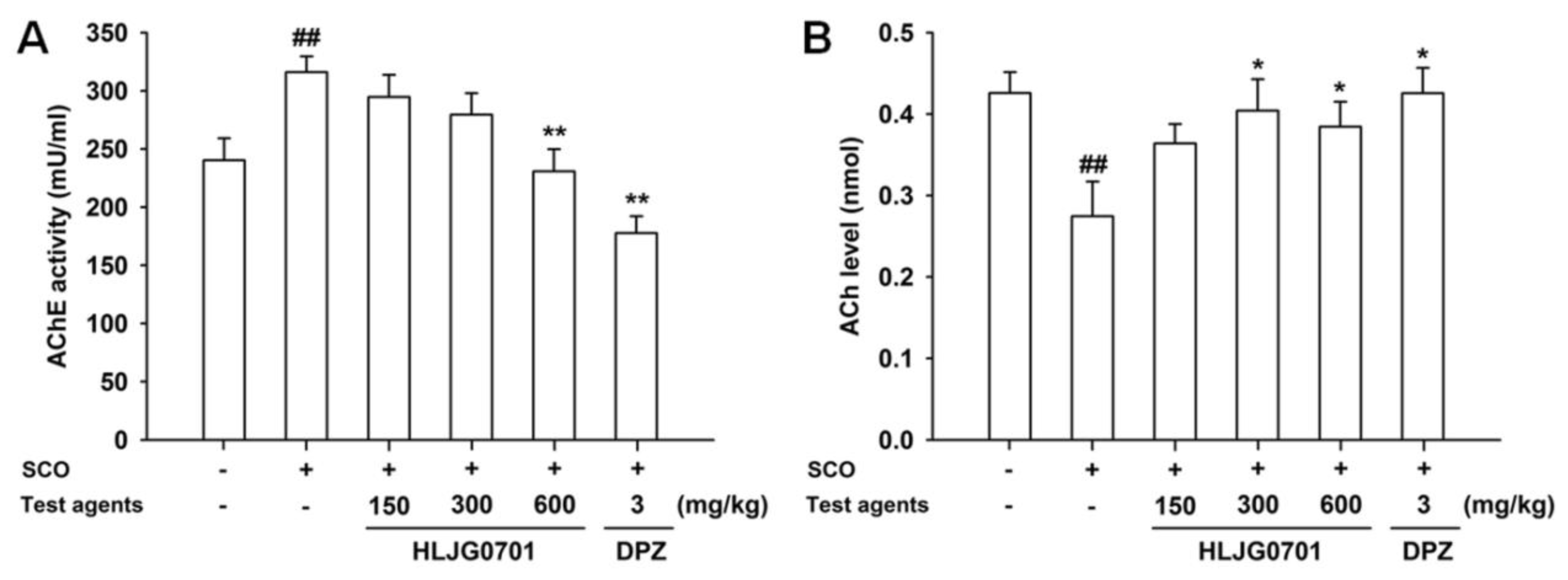
3.6. AChE Activity and ACh Contents in Brain Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food. Nutr. Res. 2009, 55, 1–99. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.H. Effects of panax ginseng in neurodegenerative diseases. J. Ginseng Res. 2012, 36, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.Y.; Zhang, J.; Yeo, S.J.; Myung, C.S.; Kim, H.M.; Kim, J.M.; Park, J.H.; Cho, J.S.; Kang, J.S. Memory enhancing and neuroprotective effects of selected ginsenosides. Arch. Pharm. Res. 2005, 28, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Shim, J.S.; Lee, S.Y.; Cho, W.H.; Hong, E.Y.; Lee, J.H.; Han, J.S.; Lee, H.J.; Lee, K.W. Rg3-enriched ginseng extract ameliorates scopolamine-induced learning deficits in mice. BMC Complement. Altern. Med. 2016, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Park, B.Y.; Hwang, H.L.; Lee, J.N.; Sohn, S.O.; Lee, S.H.; Jung, M.Y.; Lim, H.I.; Park, H.W.; Lee, J.H. Evaluation of ginsenoside bioconversion of lactic acid bacteria isolated from kimchi. J. Ginseng Res. 2017, 41, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Lim, C.S.; Cha, B.C.; Choi, S.H.; Kwon, K.R. Component analysis of cultivated ginseng, cultivated wild ginseng, and wild ginseng and the change of ginsenoside components in the process of red ginseng. J. Pharmacopuncture 2010, 13, 63–77. [Google Scholar] [CrossRef]
- Nam, K.Y.; Kim, Y.S.; Shon, M.Y.; Park, J.D. Recent advances in studies on chemical constituents and biological activities of Korean black ginseng (Panax ginseng C. A. Meyer). Kor. J. Pharmacogn. 2015, 46, 173–188. [Google Scholar]
- Wahlsten, D.; Cooper, S.F.; Crabbe, J.C. Different rankings of inbred mouse strains on the Morris maze and a refined 4-arm water escape task. Behav. Brain Res. 2005, 165, 36–51. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-Stone, R.M. A new and rapid colormetric determination of cholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Charles, V.V.; Michael, T.W. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Shibata, S.; Tanaka, O.; Soma, K.; Lita, Y.; Ando, T.; Nakamura, H. Studies on saponins and sapogenins of ginseng: The structure of panaxatriol. Tetrahedron Lett. 1965, 3, 207–213. [Google Scholar] [CrossRef]
- Lorenzini, C.A.; Baldi, E.; Bucherelli, C.; Sacchetti, B.; Tassoni, G. Role of dorsal hippocampusin acquisition, consolidation and retrieval of rat’s passive avoidance response: A tetrodotoxin functional inactivation study. Brain Res. 1996, 730, 32–39. [Google Scholar] [CrossRef]
- Elshafay, A.; Tinh, N.X.; Salman, S.; Shaheen, Y.S.; Othman, E.B.; Elhady, M.T.; Kansakar, A.R.; Tran, L.; Van, L.; Hirayama, K.; et al. Ginsenoside Rk1 bioactivity: A systematic review. PeerJ 2017, 5, e3993. [Google Scholar] [CrossRef] [PubMed]
- Son, S.H.; Choi, S.M.; Hyung, S.J.; Yun, S.R.; Choi, M.S.; Shin, E.M.; Hong, Y.P. Induction and cultures of mountain ginseng adventitious roots and AFLP analysis for identifying mountain ginseng. Biotechnol. Bioprocess Eng. 1999, 4, 119–123. [Google Scholar] [CrossRef]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Huang, D.; Li, Y.; Zhang, M.; Ruan, S.; Zhang, H.; Wang, Y.; Hu, P. Tartaric acid induced conversion of protopanaxadiol to ginsenosides Rg3 and Rg5 and their in situ recoveries by integrated expanded bed adsorption chromatography. J. Sep. Sci. 2016, 39, 2995–3001. [Google Scholar] [CrossRef]
- Ong, W.Y.; Farooqui, T.; Koh, H.L.; Farooqui, A.A.; Ling, E.A. Protective effects of ginseng on neurological disorders. Front. Aging Neurosci. 2015, 7, 129. [Google Scholar] [CrossRef]
- Wong, A.S.; Che, C.M.; Leung, K.W. Recent advances in ginseng as cancer therapeutics: A functional and mechanistic overview. Nat. Prod. Rep. 2015, 32, 256–272. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Park, J.S.; Jung, J.S.; Kim, D.H.; Kim, H.S. Anti-inflammatory effect of ginsenoside Rg5 in lipopolysaccharide-stimulated BV2 microglial cells. Int. J. Mol. Sci. 2013, 14, 9820–9833. [Google Scholar] [CrossRef]
- Radad, K.; Moldzio, R.; Rausch, W.D. Ginsenosides and their CNS targets. CNS Neurosci. Ther. 2011, 17, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Myhrer, T. Neurotransmitter systems involved in learning and memory in the rat: A meta-analysis based on studies of four behavioral tasks. Brain Res. Rev. 2003, 41, 268–287. [Google Scholar] [CrossRef]
- Drachman, D.A.; Leavitt, J. Human memory and the cholinergic system: A relationship to aging? Arch. Neurol. 1974, 30, 113–121. [Google Scholar] [CrossRef]
- Ibach, B.; Haen, E. Acetylcholinesterase inhibition in Alzheimer’s disease. Curr. Pharm. Des. 2004, 10, 231–251. [Google Scholar] [CrossRef]
- DeLaGarza, V.W. Pharmacologic treatment of Alzheimer’s disease: An update. Am. Fam. Phys. 2003, 68, 1365–1372. [Google Scholar]
- Coyle, J.T.; Price, D.L.; DeLong, M.R. Alzheimer’s disease: A disorder of cortical cholinergic innervation. Science 1983, 219, 1184–1190. [Google Scholar] [CrossRef]
- Jeong, E.J.; Lee, K.Y.; Kim, S.H.; Sung, S.H.; Kim, Y.C. Cognitive-enhancing and antioxidant activities of iridoid glycosides from Scrophularia buergeriana in scopolamine-treated mice. Eur. J. Pharmacol. 2008, 588, 78–84. [Google Scholar] [CrossRef]
- Sunita, S.; Sharlene, R.; Holly, B.B. Assessment of spatial memory in mice. Life Sci. 2010, 87, 521–536. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.; Seo, S.G.; Choi, B.R.; Han, J.S.; Lee, K.W.; Kim, J. Sulforaphane alleviates scopolamine-induced memory impairment in mice. Pharmacol. Res. 2014, 85, 23–32. [Google Scholar] [CrossRef]
- Araujo, J.A.; Studzinski, C.M.; Milgram, N.W. Further evidence for the cholinergic hypothesis of aging and dementia from the canine model of aging. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 411–422. [Google Scholar] [CrossRef]
- Tariot, P.N.; Patel, S.V.; Cox, C.; Henderson, R.E. Age-related decline in central cholinergic function demonstrated with scopolamine. Psychopharmacology 1996, 125, 50–56. [Google Scholar] [CrossRef]
- Kocahan, S.; Doğan, Z. Mechanisms of Alzheimer’s disease pathogenesis and prevention: The brain, neural pathology, N-methyl-D-aspartate receptors, tau protein and other risk factors. Clin. Psychopharmacol. Neurosci. 2017, 15, 1–8. [Google Scholar] [CrossRef]
- Huang, Y.; Mucke, L. Alzheimer mechanisms and therapeutic strategies. Cell 2012, 148, 1204–1222. [Google Scholar] [CrossRef]
- Mahley, R.W. Apolipoprotein E: Cholesterol transport protein with expanding role in cell biology. Science 1988, 240, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Cong, W.N.; Ji, S.; Rothman, S.; Maudsley, S.; Martin, B. Metabolic dysfunction in Alzheimer’s disease and related neurodegenerative disorders. Curr. Alzheimer Res. 2012, 9, 5–17. [Google Scholar] [CrossRef]
- Lee, H.J.; Park, M.K.; Seo, Y.R. Pathogenic mechanisms of heavy metal induced-Alzheimer’s disease. Toxicol. Environ. Health Sci. 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the cholinergic system. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef]
- Shenghui, C.; Junfei, G.; Liang, F.; Jiping, L.; Minghua, Z.; Xiaobin, J.; Min, L.; Danian, Y. Ginsenoside Rg5 improves cognitive dysfunction and beta-amyloid deposition in STZ-induced memory impaired rats via attenuating neuroinflammatory responses. Int. Immunopharmacol. 2014, 19, 317–326. [Google Scholar] [CrossRef]
- Siraj, F.M.; Natarajan, S.; Huq, M.A.; Kim, Y.J.; Yang, D.C. Structural investigation of ginsenoside Rf with PPARg major transcriptional factor of adipogenesis and its impact on adipocyte. J. Ginseng Res. 2015, 39, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Yesmin Simu, S.; Ahn, S.; Castro-Aceituno, V.; Yang, D.C. Ginsenoside Rg5: Rk1 exerts an anti-obesity effect on 3T3-L1 cell line by the downregulation of PPARγ and CEBPα. Iran J. Biotechnol. 2017, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Cao, H.; Cui, X.; Zheng, W.; Wang, S.; Yu, J.; Chen, Z. Silymarin’s inhibition and treatment effects for Alzheimer’s disease. Molecules 2019, 24, 1748. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Park, S.K.; Kang, J.Y.; Park, S.B.; Yoo, S.K.; Han, H.J.; Cho, K.H.; Kim, J.C.; Heo, H.J. Green tea seed oil suppressed Aβ1-42-induced behavioral and cognitive deficit via the Aβ-Related Akt Pathway. Int. J. Mol. Sci. 2019, 20, 1865. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.W.; Son, H.J.; Um, M.Y.; Kim, I.H.; Han, D.; Cho, S.; Lee, C.H. Enhanced cognitive effects of demethoxycurcumin, a natural derivative of curcumin on scopolamine-Induced memory impairment in mice. Molecules 2016, 21, 1022. [Google Scholar] [CrossRef]
- Lin, M.K.; Lee, M.S.; Huang, H.C.; Cheng, T.J.; Cheng, Y.D.; Wu, C.R. Cuscuta chinensis and C. campestris attenuate scopolamine-induced memory deficit and oxidative damage in mice. Molecules 2018, 23, 3060. [Google Scholar] [CrossRef]
- OECD. Addressing Dementia: The OECD Response, OECD Health Policy Studies; OECD Publishing: Paris, France, 2015. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Contents (mg/g) | |||
|---|---|---|---|---|
| Lot #1 | Lot #2 | Lot #3 | Average | |
| Ginsenoside Rg5 | 21.66 ± 0.13 | 21.63 ± 0.08 | 21.15 ± 0.22 | 21.48 ± 0.29 |
| Ginsenoside Rk1 | 18.89 ± 0.17 | 18.85 ± 0.09 | 18.39 ± 0.22 | 18.71 ± 0.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, K.S.; Choi, Y.O.; Lee, S.M.; Ryu, H.Y.; Kang, S.J.; Yeon, Y.; Kim, Y.R.; Lee, J.G.; Kim, C.J.; Lee, Y.j.; et al. Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice. Nutrients 2019, 11, 1120. https://doi.org/10.3390/nu11051120
An KS, Choi YO, Lee SM, Ryu HY, Kang SJ, Yeon Y, Kim YR, Lee JG, Kim CJ, Lee Yj, et al. Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice. Nutrients. 2019; 11(5):1120. https://doi.org/10.3390/nu11051120
Chicago/Turabian StyleAn, Kyu Sup, Yeo Ok Choi, So Min Lee, Hyeon Yeol Ryu, Su Jin Kang, Yong Yeon, Yu Ri Kim, Jae Geun Lee, Chul Joong Kim, Ye ji Lee, and et al. 2019. "Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice" Nutrients 11, no. 5: 1120. https://doi.org/10.3390/nu11051120
APA StyleAn, K. S., Choi, Y. O., Lee, S. M., Ryu, H. Y., Kang, S. J., Yeon, Y., Kim, Y. R., Lee, J. G., Kim, C. J., Lee, Y. j., Kang, B. J., Choi, J. E., & Song, K. S. (2019). Ginsenosides Rg5 and Rk1 Enriched Cultured Wild Ginseng Root Extract Bioconversion of Pediococcus pentosaceus HLJG0702: Effect on Scopolamine-Induced Memory Dysfunction in Mice. Nutrients, 11(5), 1120. https://doi.org/10.3390/nu11051120