American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties

 , , , , and
, , , , and 
Abstract
1. Introduction
2. American Ginseng: Cultivation, Characteristics, and Applications
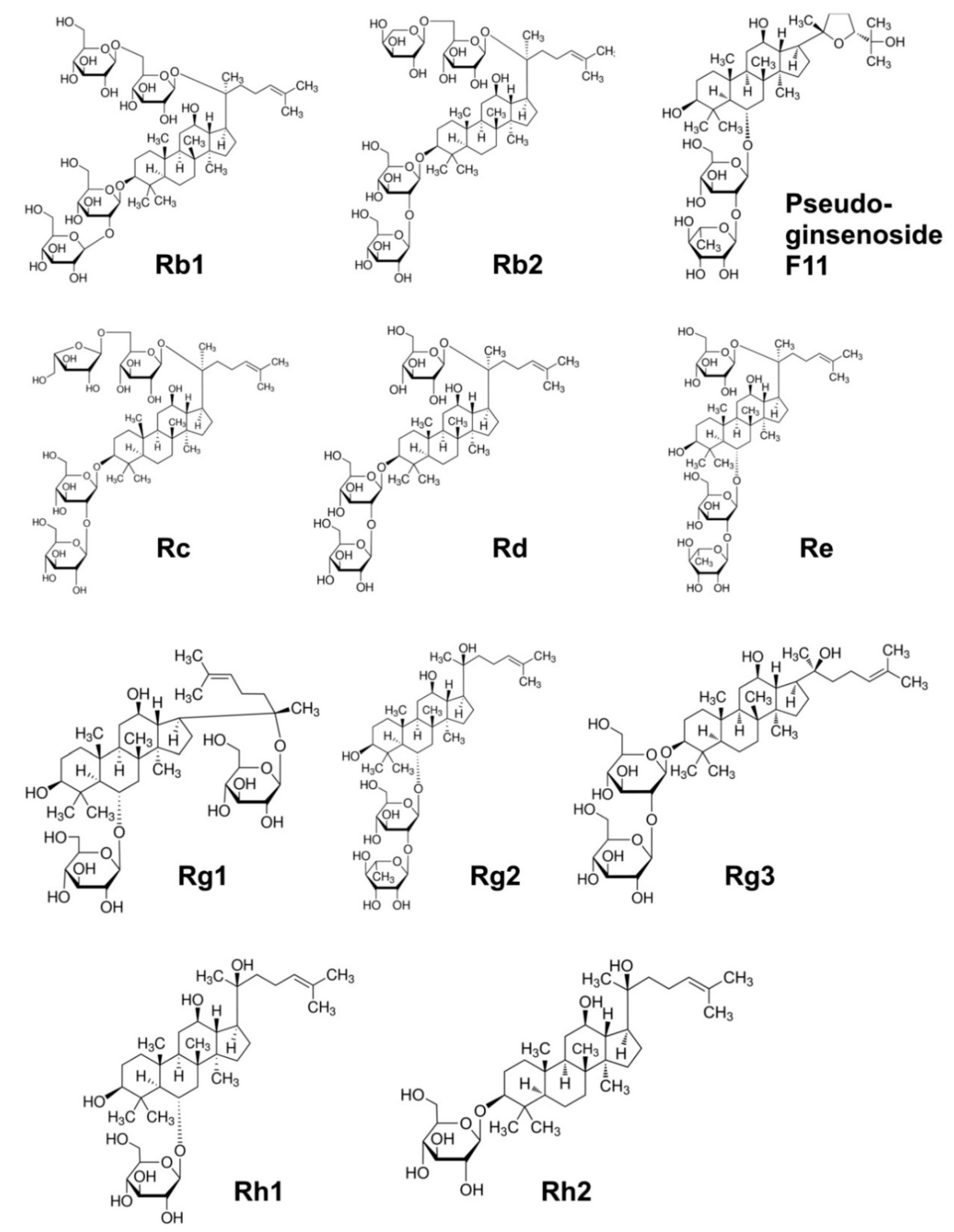
3. Bioactive Phytochemicals of American Ginseng
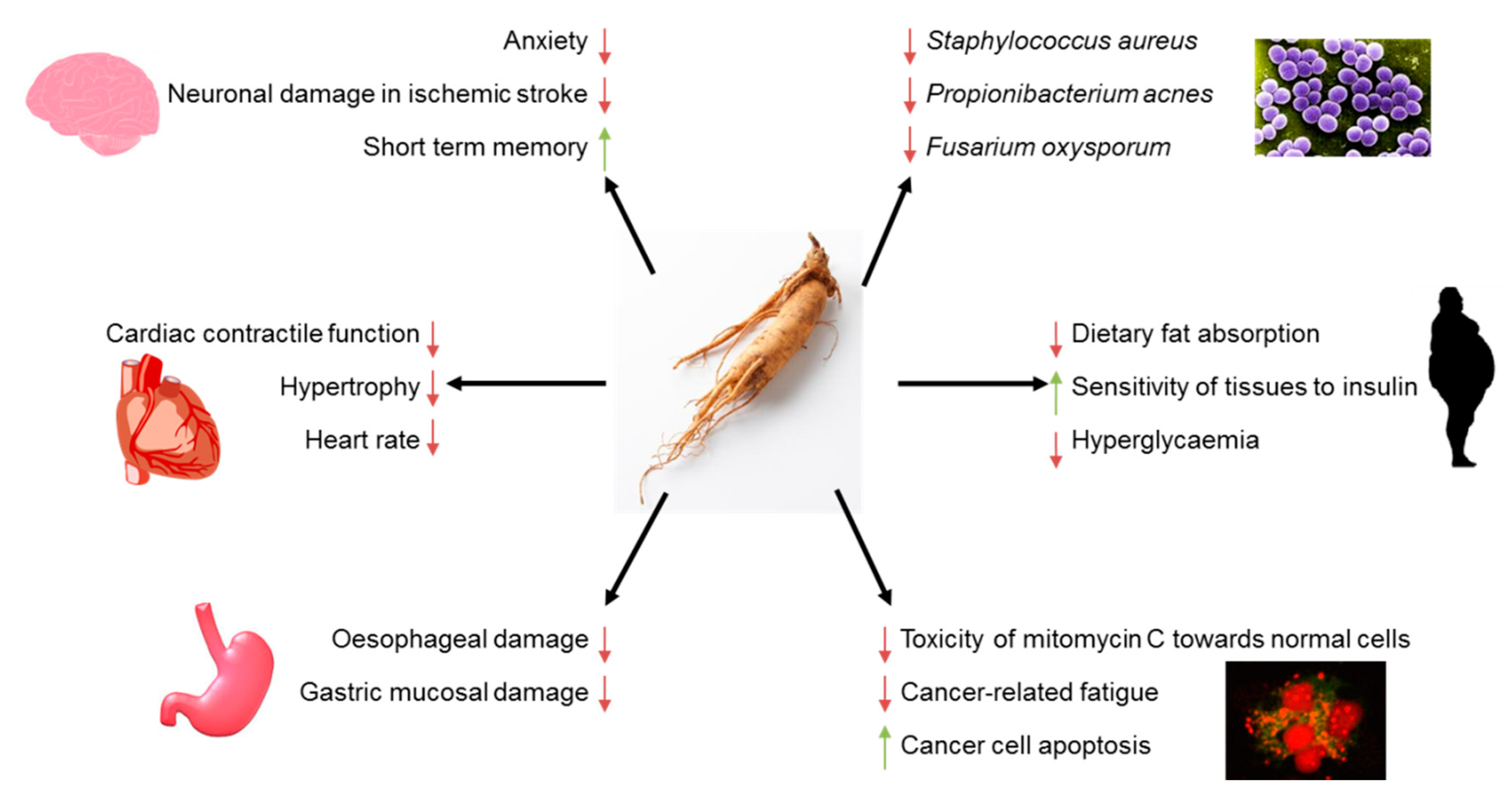
4. Pro-Health Effects of American Ginseng
4.1. Nervous System
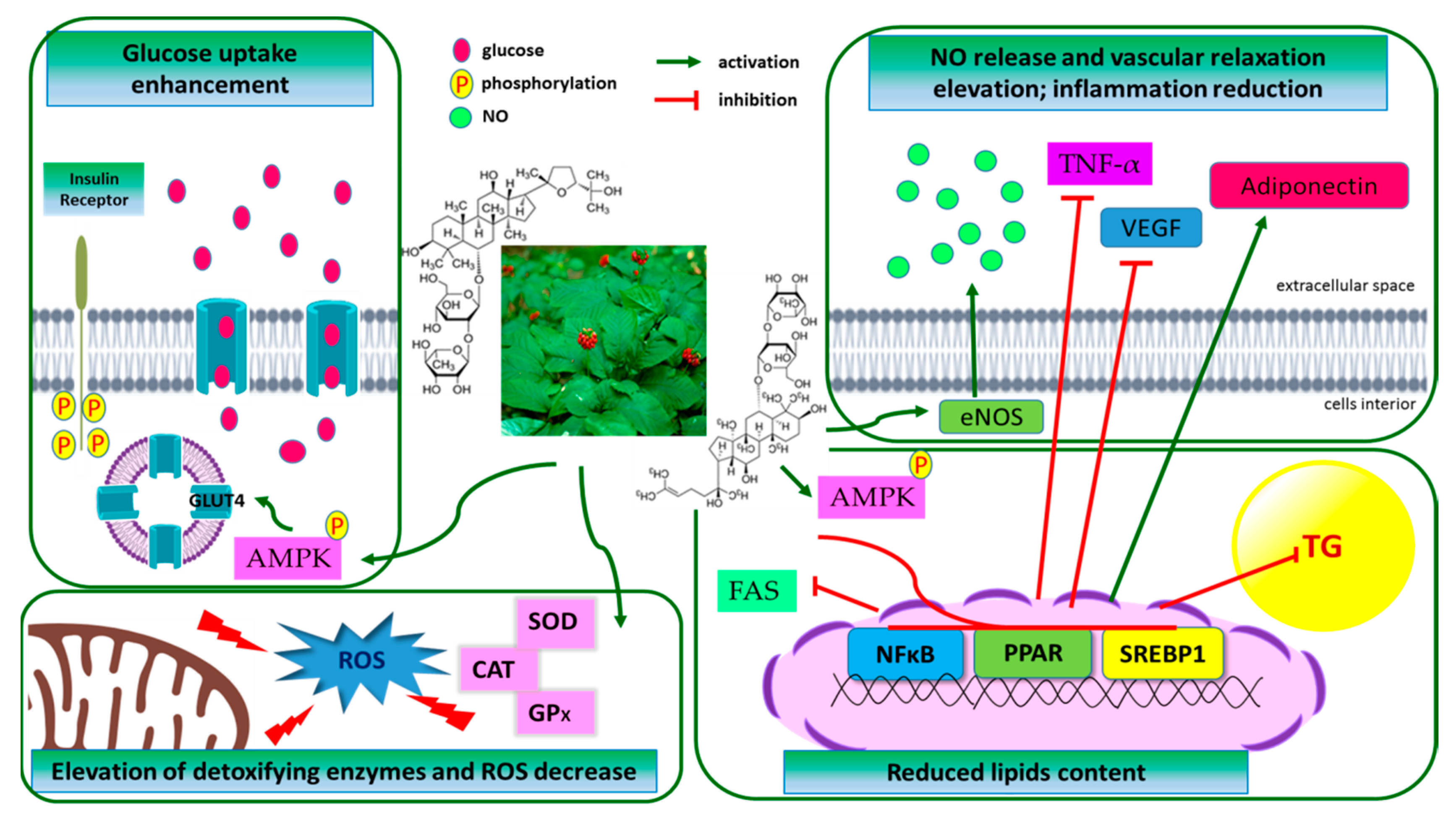
4.2. Cardiovascular and Gastrointestinal Systems
4.3. Anti-Cancer Activity
4.4. Anti-Diabetic Activity
4.5. Prevention of Obesity
4.6. Anti-Aging Properties
5. Interactions of American Ginseng with Microorganisms
5.1. Antimicrobial Action
5.2. Metabolism of American Ginseng Ginsenosides by Intestinal Microbiota
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ahmad, Z.; Hassan, S.S.; Azim, S. A therapeutic connection between dietary phytochemicals and ATP synthase. Curr. Med. Chem. 2017, 24, 3894–3906. [Google Scholar] [CrossRef]
- Lee, G.; Bae, H. Therapeutic effects of phytochemicals and medicinal herbs on depression. BioMed Res. Int. 2017, 2017, 6596241. [Google Scholar] [CrossRef] [PubMed]
- He, Y.S.; Sun, W.; Wang, C.Z.; Qi, L.W.; Yang, J.; Li, P.; Wen, X.D.; Yuan, C.S. Effects of American ginseng on pharmacokinetics of 5-fluorouracil in rats. Biomed. Chromatogr. 2014, 29, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ichikawa, T.; Jin, Y.; Hofseth, L.J.; Nagarkatti, P.; Nagarkatti, M.; Windust, A.; Cui, T. An essential role of Nrf2 in American ginseng-mediated anti-oxidative actions in cardiomyocytes. J. Ethnopharmacol. 2010, 130, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Shergis, J.L.; Di, Y.M.; Zhang, A.L.; Vlahos, R.; Helliwell, R.; Ye, J.M.; Xue, C.C. Therapeutic potential of Panax ginseng and ginsenosides in the treatment of chronic obstructive pulmonary disease. Complement. Ther. Med. 2014, 22, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Rotem, C.; Kaplan, B. Phyto-female complex for the relief of hot flushes, night sweats and quality of sleep: Randomized, controlled, double-blind pilot study. Gynecol. Endocrinol. 2007, 23, 117–122. [Google Scholar] [CrossRef]
- McGraw, J.B.; Lubbers, A.E.; Van der Voort, M.; Mooney, E.H.; Furedi, M.A.; Souther, S.; Turner, J.B.; Chandler, J. Ecology and conservation of ginseng (Panax quinquefolius) in a changing world. Ann. N. Y. Acad. Sci. 2013, 1286, 62–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Choi, H.K.; Brinckmann, J.A.; Jiang, X.; Huang, L. Chemical analysis of Panax quinquefolius (North American ginseng): A review. J. Chromatogr. A 2015, 1426, 1–15. [Google Scholar] [CrossRef]
- Qi, L.W.; Wang, C.Z.; Yuan, C.S. Ginsenosides from American ginseng: Chemical and pharmacological diversity. Phytochemistry 2011, 72, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.C. Molecular signaling of ginsenosides Rb1, Rg1, and Rg3 and their mode of actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Anderson, G.A.; Fernandez, T.G.; Doré, S. Efficacy and mechanism of Panax ginseng in experimental stroke. Front. Neurosci. 2019, 2013, 1–20. [Google Scholar] [CrossRef]
- Nakhjavani, M.; Hardingham, J.E.; Palethorpe, H.M.; Tomita, Y.; Smith, E.; Price, T.J.; Townsend, A.R. Ginsenoside Rg3: Potential molecular targets and therapeutic indication in metastatic breast cancer. Medicines 2019, 6, 17. [Google Scholar] [CrossRef]
- Yu, C.; Wang, C.Z.; Zhou, C.J.; Wang, B.; Han, L.; Zhang, C.F.; Wu, X.H.; Yuan, C.S. Adulteration and cultivation region identification of American ginseng using HPLC coupled with multivariate analysis. J. Pharm. Biomed. Anal. 2014, 99, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Jang, S.I.; Kim, Y.R.; Yang, K.E.; Yoon, S.J.; Lee, Z.W.; An, H.J.; Jang, I.S.; Choi, J.S.; Yoo, H.S. Anti-proliferative effects of ginsenosides extracted from mountain ginseng on lung cancer. Chin. J. Integr. Med. 2016, 2, 344–352. [Google Scholar] [CrossRef]
- Yang, L.; Yu, Q.T.; Ge, Y.Z.; Zhang, W.S.; Fan, Y.; Ma, C.W.; Liu, Q.; Qi, L.W. Distinct urine metabolome after Asian ginseng and American ginseng intervention based on GC-MS metabolomics approach. Sci. Rep. 2018, 6, 39045. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.R.; Nguyen, N.Q.; Lee, K.H.; Kim, Y.C.; Seo, J. Cytohistological study of the leaf structures of Panax ginseng Meyer and Panax quinquefolius L. J. Ginseng Res. 2016, 41, 463–468. [Google Scholar] [CrossRef][Green Version]
- Pan, Y.; Wang, X.; Sun, G.; Li, F.; Gong, X. Application of RAD sequencing for evaluating the genetic diversity of domesticated Panax notoginseng (Araliaceae). PloS ONE 2016, 11, e0166419. [Google Scholar] [CrossRef] [PubMed]
- Cruse-Sanders, J.M.; Hamrick, J.L. Genetic diversity in harvested and protected populations of wild American ginseng, Panax quinquefolius L. (Araliaceae). Am. J. Bot. 2004, 91, 540–548. [Google Scholar] [CrossRef]
- Jia, L.; Zhao, Y. Current evaluation of the millennium phytomedicine-ginseng (I): Etymology, pharmacognosy, phytochemistry, market and regulations. Curr. Med. Chem. 2009, 16, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Jia, C.; Liao, D.; Chen, X.; Li, X. Comparison of serum metabolite changes of radiated mice administered with Panax quinquefolium from different cultivation regions using UPLC-Q/TOF-MS based metabolomic approach. Molecules 2018, 23, 1014. [Google Scholar] [CrossRef] [PubMed]
- Souther, S.; Lechowicz, M.J.; McGraw, J.B. Experimental test for adaptive differentiation of ginseng populations reveals complex response to temperature. Ann. Bot. 2012, 110, 829–837. [Google Scholar] [CrossRef]
- Pengelly, A.; Bennett, K. Appalachian Plant Monographs: Panax quinquefolius L., American Ginseng. Available online: http://www.frostburg.edu/aces/appalachian-plants/ (accessed on 5 January 2019).
- Lim, W.; Mudge, K.W.; Vermeylen, F. Effects of population, age and cultivation methods on ginsenoside content of wild American ginseng (Panax quinquefolium). J. Agric. Food Chem. 2005, 53, 8498–8505. [Google Scholar] [CrossRef] [PubMed]
- Proctor, J.T.; Shelp, B.J. Effect of boron nutrition on American ginseng in field and in nutrient cultures. J. Ginseng Res. 2013, 38, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Bae, I. Quality Characteristics, changes in physiochemical properties and functional properties of camembert cheese containing red ginseng powder. Korean J. Food Sci. Anim. Resour. 2018, 38, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.T.; Yoo, K.M.; Lee, J.W.; Eom, S.H.; Hwang, I.K.; Lee, C.Y. Protective effect of steamed American ginseng (Panax quinquefolius L.) on V79-4 cells induced by oxidative stress. J. Ethnopharmacol. 2007, 111, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Dolot, M.; Smigielski, K.; Wesolowska, M. Analysis of chosen nutrients in American ginseng (Panax quinquefolium L.) cultivated in Poland. Sci. Pap. Tech. Univ. Lodz Food Chem. Biotechnol. 2006, 70, 53–63. [Google Scholar]
- Sengupta, S.; Toh, S.A.; Sellers, L.A.; Skepper, J.N.; Koolwijk, P.; Leung, H.W.; Yeung, H.W.; Wong, R.N.; Sasisekharan, R.; Fan, T.P. Modulating angiogenesis: The yin and the yang in ginseng. Circulation 2004, 110, 1219–1225. [Google Scholar] [CrossRef]
- Jung, J.; Kim, K.H.; Yang, K.; Bang, K.H.; Yang, T.J. Practical application of DNA markers for high-throughput authentication of Panax ginseng and Panax quinquefolius from commercial ginseng products. J. Ginseng Res. 2014, 38, 123–129. [Google Scholar] [CrossRef]
- Ma, Z.N.; Li, Y.Z.; Li, W.; Yan, X.T.; Yang, G.; Zhang, J.; Zhao, L.C.; Yang, L.M. Nephroprotective effects of saponins from leaves of Panax quinquefolius against cisplatin-induced acute kidney injury. Int. J. Mol. Sci. 2017, 18, 1407. [Google Scholar] [CrossRef]
- Nag, S.A.; Qin, J.; Wang, W.; Wang, M.H.; Wang, H.; Zhang, R. Ginsenosides as anticancer agents: In Vitro and in vivo activities, structure–activity relationships and molecular mechanisms of action. Front. Pharmacol. 2012, 3, 25. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Lipert, A.; Szymańska, G. Yeast extract stimulates ginsenoside production in hairy root cultures of American ginseng cultivated in shake flasks and nutrient sprinkle bioreactors. Molecules 2017, 22, 880. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Liu, J.; Wang, Z.; Zhang, J.; Cates, C.; Rousselle, T.; Meng, Q.; Li, J. The structure-activity relationship of ginsenosides on hypoxia-reoxygenation induced apoptosis of cardiomyocytes. Biochem. Biophys. Res. Commun. 2017, 494, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Z.; Hu, Y.; Wu, W.Y.; Ye, M.; Guo, D.A. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.B.; Wang, C.Z.; Wu, J.A.; Osinski, J.; Yuan, C.S. Determination of major ginsenosides in Panax quinquefolius (American ginseng) using high-performance liquid chromatography. Phytochem. Anal. 2005, 16, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Shukla, Y.N.; Tripathi, A.K.; Mehta, V.K. Feeding-deterrency of oleanolic acid isolated from Panax quinquefolium against lepidopterans. Phytother. Res. 1997, 11, 591–593. [Google Scholar] [CrossRef]
- Huang, X.; Liu, Y.; Zhang, N.; Sun, X.; Yue, H.; Chen, C.; Liu, S. UPLC Orbitrap HRMS analysis of Panax quinquefolium L. for authentication of Panax genus with chemometric methods. J. Chromatogr. Sci. 2018, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Chemical diversity of Panax ginseng, Panax quinquefolium, and Panax notoginseng. J Ginseng Res. 2012, 36, 1–15. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zhang, T.Y.; Xue, X.; Liu, D.M.; Zhang, H.T.; Yuan, L.L.; Liu, Y.L.; Yang, H.L.; Sun, S.B.; Zhang, C.; et al. Pseudoginsenoside-F11 attenuates cerebral ischemic injury by alleviating autophagic/lysosomal defects. CNS Neurosci. Ther. 2017, 23, 567–579. [Google Scholar] [CrossRef]
- Popovich, D.G.; Yeo, C.R.; Zhang, W. Ginsenosides derived from Asian (Panax ginseng), American ginseng (Panax quinquefolius) and potential cytoactivity. Int. J. Biomed. Pharma. Sci. 2012, 6, 56–62. [Google Scholar]
- Zhang, X.J.; Huang, L.L.; Cai, X.J.; Li, P.; Wang, Y.T.; Wan, J.B. Fatty acid variability in three medicinal herbs of Panax species. Chem. Cent. J. 2013, 7, 12. [Google Scholar] [CrossRef]
- Wang, M.; Guilbert, L.J.; Li, J.; Wu, Y.; Pang, P.; Basu, T.K.; Shan, J.J. A proprietary extract from north American ginseng (Panax quinquefolium) enhances IL-2 and IFN-gamma productions in murine spleen cells induced by Con-A. Int. Immunopharmacol. 2004, 4, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, X.; Yang, X.; Li, Y.; Yao, Y.; Lui, E.M.K.; Ren, G. Structural and anti-inflammatory characterization of a novel neutral polysaccharide from North American ginseng (Panax quinquefolius). Int. J. Biol. Macromol. 2015, 74, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczuk, A.; Wolski, T.; Hołderna-Kędzia, E. Estimation of the chemical composition and antimicrobial and antioxidant activity of extracts received from leaves and roots of American ginseng (Panax quinquefolium L.). Herba Pol. 2006, 52, 79–90. [Google Scholar]
- Cui, S.; Wu, J.; Wang, J.; Wang, X. Discrimination of American ginseng and Asian ginseng using electronic nose and gas chromatography mass spectrometry coupled with chemometrics. J. Ginseng Res. 2017, 41, 85–95. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Szymańska, G.; Wajs-Bonikowska, A.; Bonikowski, R.; Sienkiewicz, M. The increase of triterpene saponin production induced by trans-anethole in hairy root cultures of Panax quinquefolium. Molecules 2018, 23, 2674. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Cai, Y.; Anderson, S.; Yuan, C.S. Ginseng metabolites on cancer chemoprevention: An angiogenesis link? Diseases 2015, 3, 193–204. [Google Scholar] [CrossRef]
- Sun, J.; Chen, P. Differentiation of Panax quinquefolius grown in the USA and China using LC/MS-based chromatographic fingerprinting and chemometric approaches. Anal. Bioanal. Chem. 2011, 399, 1877–1889. [Google Scholar] [CrossRef]
- Sun, X.; Chen, P.; Cook, S.L.; Jackson, G.P.; Harnly, J.M.; Harrington, P.B. Classification of cultivation locations of Panax quinquefolius L samples using high performance liquid chromatography-electrospray ionization mass spectrometry and chemometric analysis. Anal. Chem. 2012, 84, 3628–3634. [Google Scholar] [CrossRef] [PubMed]
- Kochan, E.; Kołodziej, B.; Gadomska, G.; Chmiel, A. Content of ginsenosides in Panax quinquefolium from field cultivation. Herba Pol. 2004, 50, 20–27. [Google Scholar] [CrossRef]
- Xiao, D.; Yue, H.; Xiu, Y.; Sun, X.; Wang, Y.B.; Liu, S.Y. Accumulation characteristics and correlation analysis of five ginsenosides with different cultivation ages from different regions. J. Ginseng Res. 2015, 39, 338–344. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Wang, L.; Chen, X.; Pang, X.; Han, J. A Nucleotide signature for the identification of American ginseng and its products. Front. Plant Sci. 2016, 7, 319. [Google Scholar] [CrossRef]
- Mancuso, C.; Santangelo, R. Panax ginseng and Panax quinquefolius: From pharmacology to toxicology. Food Chem. Toxicol. 2017, 107, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Wolski, T.; Ludwiczuk, A.; Baj, T.; Głowniak, K.; Świątek, M. Genus Panax—Taxonomy, chemical composition, pharmacological effects, medicinal application and phytochemical analysis of aerial and underground parts of American ginseng—Panax quinquefolium l. Part I. Post. Fitoterapii 2008, 2, 94–114. [Google Scholar]
- Chen, C.; Chiou, W.; Zhang, J. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin. 2008, 29, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Park, J.D.; Rhee, D.K.; Lee, Y.H. Biological activities and chemistry of saponins from Panax ginseng C.A. Meyer. Phytochem. Rev. 2005, 4, 159–175. [Google Scholar] [CrossRef]
- Rokot, N.T.; Kairupan, T.S.; Cheng, C.-H.; Runtuwene, J.; Kapantow, N.H.; Amitani, M.; Morinaga, A.; Amitani, H.; Asakawa, A.; Inui, A. A Role of ginseng and its constituents in the treatment of central nervous system disorders. Evid. Based Complement. Alternat. Med. 2016, 2016, 2614742. [Google Scholar] [CrossRef] [PubMed]
- Popovich, D.G.; Kitts, D.D. Generation of ginsenosides Rg3 and Rh2 from North American ginseng. Phytochemistry 2004, 65, 337–344. [Google Scholar] [CrossRef]
- Rasheed, N.; Tyagi, E.; Ahmad, A.; Siripurapu, K.B.; Lahiri, S.; Shukla, R.; Palit, G. Involvement of monoamines and proinflammatory cytokines in mediating the anti-stress effects of Panax quinquefolium. J. Ethnopharmacol. 2008, 117, 257–262. [Google Scholar] [CrossRef]
- Xu, H.; Yu, X.; Qu, S.; Chen, Y.; Wang, Z.; Sui, D. In vive and in vitro cardioprotective effects of Panax quinquefolium 20(S)-protopanaxadiol saponins (PQDS), isolated from Panax quinquefolium. Pharmazie 2013, 68, 287–292. [Google Scholar]
- Wang, M.; Guilbert, L.J.; Ling, L.; Li, J.; Wu, Y.; Xu, S.; Pang, P.; Shan, J.J. Immunomodulating activity of CVT-E002, a proprietary extract from North American ginseng (Panax quinquefolium). J. Pharma Pharmacol. 2001, 53, 1515–1523. [Google Scholar] [CrossRef]
- Xie, G.; Wang, C.Z.; Yu, C.; Qiu, Y.; Wen, X.D.; Zhang, C.F.; Yuan, C.S.; Jia, W. Metabonomic profiling reveals cancer chemopreventive effects of American ginseng on colon carcinogenesis in Apc(Min/+) mice. J. Proteome Res. 2015, 14, 3336–3347. [Google Scholar] [CrossRef]
- Kochan, E.; Wasiela, M.; Sienkiewicz, M. The production of ginsenosides in hairy root cultures of American ginseng, Panax quinquefolium L. and their antimicrobial activity. In Vitro Cell Dev. Biol. Plant 2012, 49, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.; Guo, H.; Cha, Y.; Ban, Y.H.; Seo, D.W.; Choi, Y.; Kim, T.S.; Lee, S.P.; Kim, J.C.; Choi, E.K.; et al. Cereboost™ an American ginseng extract improves cognitive function via up-regulation of choline acetyltransferase expression and neuroprotection. Regul. Toxicol. Pharmacol. 2016, 78, 53–58. [Google Scholar] [CrossRef]
- Bandelow, B.; Michaelis, S. Epidemiology of anxiety disorders in the 21st century. Dialogues Clin. Neurosci. 2015, 17, 327–335. [Google Scholar] [PubMed]
- Wei, X.Y.; Yang, J.Y.; Wang, J.H.; Wu, C.F. Anxiolytic effect of saponins from Panax quinquefolium in mice. J. Ethnopharmacol. 2007, 111, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.; Ossoukhova, A.; Owen, L.A.; Pipingas, A.; He, K.; Roller, M.; Stough, C. Effects of American ginseng (Panax quinquefolius) on neurocognitive function: An acute, randomized, double-blind, placebo-controlled, crossover study. Psychopharmacology 2012, 212, 345–356. [Google Scholar] [CrossRef]
- Liu, D.; Li, B.; Liu, Y.; Attele, A.S.; Kyle, J.W.; Yuan, C.S. Voltage-dependent inhibition of brain Na+ channels by American ginseng. Eur. J. Pharmacol. 2001, 413, 47–54. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Chu, S.; Wang, Y.; Wang, X.; Chen, N.; Zhang, J. Improvement of memory in mice and increase of hippocampal excitability in rats by ginsenoside Rg1’s metabolites ginsenoside Rh1 and protopanaxatriol. J. Pharmacol. Sci. 2009, 109, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Liu, Y.L.; Song, M.; Liu, W.; Wang, J.H.; Li, X.; Yang, J.Y. Protective effects of pseudoginsenoside-F11 on methamphetamine-induced neurotoxicity in mice. Pharmacol. Biochem. Behav. 2003, 76, 103–109. [Google Scholar] [CrossRef]
- Bowie, L.E.; Roscoe, W.A.; Lui, E.M.K.; Smith, R.; Karlik, S.J. Effects of an aqueous extract of North American ginseng on MOG (35–55)-induced EAE in mice. Can. J. Physiol. Pharmacol. 2012, 90, 933–939. [Google Scholar] [CrossRef]
- Sloley, B.D.; Pang, P.K.; Huang, B.H.; Ba, F.; Li, F.L.; Benishin, C.G.; Greenshaw, A.J.; Han, J.J. American ginseng extract reduces scopolamine-induced amnesia in a spatial learning task. J. Psychiat. Neurosci. 1999, 24, 442–452. [Google Scholar]
- Wang, J.Y.; Yang, J.Y.; Wang, F.; Fu, S.Y.; Hou, Y.; Jiang, B.; Ma, J.; Song, C.; Wu, C.F. Neuroprotective effect of pseudoginsenoside-f11 on a rat model of Parkinson's disease induced by 6-hydroxydopamine. Evid. Based Complement. Alternat. Med. 2013, 2013, 152798. [Google Scholar] [CrossRef]
- Dou, H.C.; Chen, J.Y.; Ran, T.F.; Jiang, W.M. Panax quinquefolius saponin inhibits endoplasmic reticulum stress-mediated apoptosis and neurite injury and improves functional recovery in a rat spinal cord injury model. Biomed. Pharmacother. 2018, 102, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Lin, H.; Miyamoto, Y.; Wu, C.; Yang, J.; Uno, K.; Nitta, A. Pseudoginsenoside-F11 inhibits methamphetamine-induced behaviors by regulating dopaminergic and GABAergic neurons in the nucleus accumbens. Psychopharmacology 2015, 233, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.S.; Wang, X.; Wu, J.A.; Attele, A.S.; Xie, J.T.; Gu, M. Effects of Panax quinquefolius L. on brainstem neuronal activities: Comparison between Wisconsin-cultivated and Illinois-cultivated roots. Phytomedicine 2011, 8, 178–183. [Google Scholar] [CrossRef]
- Chanana, P.; Kumar, A. GABA-BZD receptor modulating mechanism of Panax quinquefolius against 72-h sleep deprivation induced anxiety like behavior: Possible roles of oxidative stress, mitochondrial dysfunction and neuroinflammation. Front. Neurosci. 2016, 10, 84. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, M.; Lu, Z.; Wang, Y.; Yu, X.; Sui, D.; Fu, L. Ginsenoside Rg3 induces ginsenoside Rb1—Comparable cardioprotective effects independent of reducing blood pressure in rats. Exp. Ther. Med. 2017, 14, 4977–4985. [Google Scholar] [CrossRef]
- Jiang, M.; Murias, J.M.; Chrones, T.; Sims, S.M.; Lui, E.; Noble, E.G. American ginseng acutely regulates contractile function of rat heart. Front. Pharmacol. 2014, 5, 43. [Google Scholar] [CrossRef]
- Xu, H.; Yu, X.; Qu, S.; Chen, Y.; Wang, Z.; Sui, D. Protective effect of Panax quinquefolium 20(S)-protopanaxadiol saponins, isolated from Panax quinquefolium on permanent focal cerebral ischemic injury in rats. Exp. Ther. Med. 2014, 7, 165–170. [Google Scholar] [CrossRef]
- Tang, X.; Gan, X.T.; Rajapurohitam, V.; Huang, C.X.; Xue, J.; Lui, E.M.K.; Karmazyn, M. North American ginseng (Panax quinquefolius) suppresses β-adrenergic-dependent signalling, hypertrophy, and cardiac dysfunction. Can. J. Physiol. Pharmacol. 2016, 94, 1325–1335. [Google Scholar] [CrossRef]
- Wu, Y.; Lu, X.; Xiang, F.L.; Lui, E.M.K.; Feng, Q. North American ginseng protects the heart from ischemia and reperfusion injury via upregulation of endothelial nitric oxide synthase. Pharmacol. Res. 2011, 64, 195–202. [Google Scholar] [CrossRef]
- Wu, Y.; Qin, C.; Lu, X.; Marchiori, J.; Feng, Q. North American ginseng inhibits myocardial NOX2-ERK1/2 signaling and tumor necrosis factor-alpha expression in endotoxemia. Pharmacol. Res. 2016, 111, 217–225. [Google Scholar] [CrossRef]
- Singh, P.; Singh, N.; Sengupta, S.; Palit, G. Ameliorative effects of Panax quinquefolium on experimentally induced reflux oesophagitis in rats. Indian J. Med. Res. 2012, 135, 407–413. [Google Scholar]
- Huang, C.C.; Chen, Y.M.; Wang, D.C.; Chiu, C.C.; Lin, W.T.; Huang, C.Y.; Hsu, M.C. Cytoprotective effect of American ginseng in a rat ethanol gastric ulcer model. Molecules 2013, 19, 316–326. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/cancer/en/ (accessed on 4 February 2019).
- Zhao, Y.; Hu, X.; Zuo, X.; Wang, M. Chemopreventive effects of some popular phytochemicals on human colon cancer: A review. Food Funct. 2018, 9, 4548–4568. [Google Scholar] [CrossRef]
- Dandawate, P.R.; Subramaniam, D.; Jensen, R.A.; Anant, S. Targeting cancer stem cells and signaling pathways by phytochemicals: Novel approach for breast cancer therapy. Semin. Cancer Biol. 2016, 40-41, 192–208. [Google Scholar] [CrossRef]
- Rais, J.; Jafri, A.; Siddiqui, S.; Tripathi, M.; Arshad, M. Phytochemicals in the treatment of ovarian cancer. Front. Biosci. 2017, 9, 67–75. [Google Scholar] [CrossRef]
- Kumar, N.B.; Krischer, J.P.; Allen, K.; Riccardi, D.; Besterman-Dahan, K.; Salup, R.; Kang, L.; Xu, P.; Pow-Sang, J. Safety of purified isoflavones in men with clinically localized prostate cancer. Nutr. Cancer 2007, 59, 169–175. [Google Scholar] [CrossRef]
- Nishino, H. Phytochemicals in hepatocellular cancer prevention. Nutr. Cancer 2009, 61, 789–791. [Google Scholar] [CrossRef]
- Yang, J.; Yuan, D.; Xing, T.; Su, H.; Zhang, S.; Wen, J.; Bai, Q.; Dang, D. Ginsenoside Rh2 inhibiting HCT116 colon cancer cell proliferation through blocking PDZ-binding kinase/T-LAK cell-originated protein kinase. J. Ginseng Res. 2016, 40, 400–408. [Google Scholar] [CrossRef]
- Yao, H.; Wan, J.Y.; Zeng, J.; Huang, W.H.; Sava-Segal, C.; Li, L.; Niu, X.; Wang, Q.; Wang, C.Z.; Yuan, C.S. Effects of compound K, an enteric microbiome metabolite of ginseng, in the treatment of inflammation associated colon cancer. Oncol. Lett. 2018, 15, 8339–8348. [Google Scholar] [CrossRef]
- Gao, J.L.; Lv, G.Y.; He, B.C.; Zhang, B.Q.; Zhang, H.; Wang, N.; Wang, C.Z.; Du, W.; Yuan, C.S.; He, T.C. Ginseng saponin metabolite 20(S)-protopanaxadiol inhibits tumor growth by targeting multiple cancer signaling pathways. Oncol. Rep. 2013, 30, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wen, X.D.; Zhang, Z.; Zhang, C.F.; Wu, X.H.; Martin, A.; Du, W.; He, T.C.; Wang, C.Z.; Yuan, C.S. American ginseng attenuates azoxymethane/dextran sodium sulfate-induced colon carcinogenesis in mice. J. Ginseng Res. 2014, 39, 14–21. [Google Scholar] [CrossRef]
- Qu, Y.; Wang, Z.; Zhao, F.; Liu, J.; Zhang, W.; Li, J.; Xu, H. AFM-detected apoptosis of hepatocellular carcinoma cells induced by American ginseng root water extract. Micron 2018, 104, 1–7. [Google Scholar] [CrossRef]
- King, M.; Murphy, L. American ginseng (Panax quinquefolius L.) extract alters mitogen-activated protein kinase cell signaling and inhibits proliferation of MCF-7-cells. J. Exp. Ther. Oncol. 2007, 6, 147–155. [Google Scholar] [PubMed]
- Li, B.; Wang, C.Z.; He, T.C.; Yuan, C.S.; Du, W. Antioxidants potentiate American ginseng-induced killing of colorectal cancer cells. Cancer Lett. 2010, 289, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Blasiak, J. DNA-damaging anticancer drugs - A Perspective for DNA repair- oriented therapy. Curr. Med. Chem. 2017, 24, 1488–1503. [Google Scholar] [CrossRef]
- Pawar, A.A.; Tripathi, D.N.; Ramarao, P.; Jena, G. Protective effects of American ginseng (Panax quinquefolium) against mitomycin C induced micronuclei in mice. Phytother. Res. 2007, 21, 1221–1227. [Google Scholar] [CrossRef]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- Ma, Z.N.; Liu, Z.; Wang, Z.; Ren, S.; Tang, S.; Wang, Y.P.; Xiao, S.Y.; Chen, C.; Li, W. Supplementation of American ginseng berry extract mitigated cisplatin-evoked nephrotoxicity by suppressing ROS-mediated activation of MAPK and NF-kappaB signaling pathways. Food Chem. Toxicol. 2017, 110, 62–73. [Google Scholar] [CrossRef]
- Mehendale, S.; Aung, H.; Wang, A.; Yin, J.J.; Wang, C.Z.; Xie, J.T.; Yuan, C.S. American ginseng berry extract and ginsenoside Re attenuate cisplatin-induced kaolin intake in rats. Cancer Chemother. Pharmacol. 2005, 56, 63–69. [Google Scholar] [CrossRef]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef]
- Lee, T.K.; O'Brien, K.F.; Wang, W.; Sheng, C.; Wang, T.; Johnke, R.M.; Allison, R.R. American ginseng modifies Cs-induced DNA damage and oxidative stress in human lymphocytes. Open Nucl. Med. J. 2009, 1, 1–8. [Google Scholar] [CrossRef]
- Lee, T.K.; O’Brien, K.F.; Wang, W.; Johnke, R.M.; Sheng, C.; Benhabib, S.M.; Wang, T.; Allison, R.R. Radioprotective effect of American ginseng on human lymphocytes at 90 minutes postirradiation: A study of 40 cases. J. Altern. Complement. Med. 2010, 16, 561–567. [Google Scholar] [CrossRef]
- Wang, C.Z.; Li, X.L.; Wang, Q.F.; Mehendale, S.R.; Fishbein, A.B.; Han, A.H.; Sun, S.; Yuan, C.S. The mitochondrial pathway is involved in American ginseng-induced apoptosis of SW-480 colon cancer cells. Oncol. Rep. 2009, 21, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.E. Cancer-related fatigue—Mechanisms, risk factors, and treatments. Nat. Rev. Clin. Oncol. 2014, 11, 597–609. [Google Scholar] [CrossRef]
- Horneber, M.; Fischer, I.; Dimeo, F.; Rüffer, J.U.; Weis, J. Cancer-related fatigue: Epidemiology, pathogenesis, diagnosis, and treatment. Dtsch. Arztebl. Int. 2012, 109, 161–171. [Google Scholar] [CrossRef]
- Barton, D.L.; Liu, H.; Dakhil, S.R.; Linquist, B.; Sloan, J.A.; Nichols, C.R.; McGinn, T.W.; Stella, P.J.; Seeger, G.R.; Sood, A.; Loprinzi, C.L. Wisconsin ginseng (Panax quinquefolius) to improve cancer-related fatigue: A randomized, double-blind trial, N07C2. J. Natl. Cancer Inst. 2013, 105, 1230–1238. [Google Scholar] [CrossRef]
- Yang, C.S.; Ko, S.R.; Cho, B.G. The ginsenoside metabolite compound K, a novel agonist of glucocorticoid receptor, induces tolerance to endotoxin induced lethal shock. J. Cell. Mol. Med. 2008, 12, 1739–1753. [Google Scholar] [CrossRef]
- Jin, Y.; Hofseth, A.B.; Cui, X. American ginseng suppresses colitis through p53-mediated apoptosis of inflammatory cells. Cancer Prev. Res. 2010, 3, 339–347. [Google Scholar] [CrossRef]
- Kang, A.; Hao, H.; Zheng, X. Peripheral anti-inflammatory effects explain the ginsenosides paradox between poor brain distribution and antidepression efficacy. J. Neuroinflammat. 2011, 8, 100. [Google Scholar] [CrossRef]
- Kan, J.; Velliquette, R.A.; Grann, K.; Burns, C.R.; Scholten, J.; Tian, F.; Zhang, Q.; Gui, N. A novel botanical formula prevents diabetes by improving insulin resistance. BMC Complement. Altern. Med. 2017, 17, 352. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Ji, L.L. Chronic ginseng consumption attenuates age-associated oxidative stress in rats. J. Nutr. 2003, 133, 3603–3609. [Google Scholar] [CrossRef]
- Luo, J.Z.; Luo, L. Ginseng on hyperglycemia: Effects and mechanisms. Evid. Based Complement. Alternat. Med. 2009, 6, 423–427. [Google Scholar] [CrossRef]
- Yoo, K.M.; Lee, C.; Lo, Y.M.; Moon, B.-K. The hypoglycemic effects of American red ginseng (Panax quinquefolius L.) on a diabetic mouse model. J. Food Sci. 2012, 77, 147–152. [Google Scholar] [CrossRef]
- Lin, E.; Wang, Y.; Mehendale, S.; Sun, S.; Wang, C.Z.; Xie, J.T.; Aung, H.H.; Yuan, C.S. Antioxidant protection by American ginseng in pancreatic beta-cells. Am. J. Chin. Med. 2008, 36, 981–988. [Google Scholar] [CrossRef]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Xie, J.T.; Wang, C.Z.; Ni, M.; Wu, J.A.; Mehendale, S.R.; Aung, H.H.; Foo, A.; Yuan, C.S. American ginseng berry juice intake reduces blood glucose and body weight in ob/ob mice. J. Food Sci. 2007, 72, S590–S594. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.H.; Xie, J.T.; Vanden Hoek, T.L.; Mehendale, S.; Aung, H.; Li, C.Q.; Qin, Y.; Schumacker, P.T.; Becker, L.B.; Yuan, C.S. Antioxidant effects of American ginseng berry extract in cardiomyocytes exposed to acute oxidant stress. Biochim. Biophys. Acta 2004, 1670, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.T.; Shao, Z.H.; Vanden Hoek, T.L.; Chang, W.T.; Li, J.; Mehendale, S.; Wang, C.Z.; Hsu, C.W.; Becker, L.B.; Yin, J.J.; et al. Antioxidant effects of ginsenoside Re in cardiomyocytes. Eur. J. Pharmacol. 2006, 532, 201–207. [Google Scholar] [CrossRef]
- Mehendale, S.R.; Wang, C.Z.; Shao, Z.H.; Li, C.Q.; Xie, J.T.; Aung, H.H.; Yuan, C.S. Chronic pretreatment with American ginseng berry and its polyphenolic constituents attenuate oxidant stress in cardiomyocytes. Eur. J. Pharmacol. 2006, 553, 209–214. [Google Scholar] [CrossRef]
- Wang, J.-S.; Yin, H.-J.; Guo, C.-Y.; Huang, Y.; Xia, C.-D.; Liu, Q. Influence of high blood glucose fluctuation on endothelial function of type 2 diabetes mellitus rats and effects of Panax quinquefolius saponin of stem and leaf. Chin. J. Integr. Med. 2013, 19, 217–222. [Google Scholar] [CrossRef]
- Sen, S.; Chen, S.; Feng, B.; Yuexiu, W.; Lui, E.M.K.; Chakrabarti, S. American ginseng (Panax quinquefolius) prevents glucose-induced oxidative stress and associated endothelial cell abnormalities. Phytomedicine 2011, 18, 1110–1117. [Google Scholar] [CrossRef]
- Mucalo, I.; Jovanovski, E.; Rahelić, D.; Božikov, V.; Romić, Z.; Vuksan, V. Effect of American ginseng (Panax quinquefolius L.) on arterial stiffness in subjects with type-2 diabetes and concomitant hypertension. J. Ethnopharm. 2013, 150, 148–153. [Google Scholar] [CrossRef]
- Xu, Z.; Beljan-Zdravkovic, U.; Bateman, R.M.; Jenkins, A.L.; Sievenpiper, J.L.; Stavro, M.P.; Vuksan, V. American ginseng increases plasma nitric oxide concentration in type 2 diabetes. Can. J. Diabet. Care. 2000, 24, 38. [Google Scholar]
- Sen, S.; Chen, S.; Wu, Y.; Feng, B.; Lui, E.K.; Chakrabarti, S. Preventive effects of North American ginseng (Panax quinquefolius) on diabetic retinopathy and cardiomyopathy. Phytother. Res. 2013, 27, 290–298. [Google Scholar] [CrossRef]
- Amin, K.A.; Awad, E.M.; Nagy, M.A. Effects of Panax quinquefolium on streptozotocin-induced diabetic rats: role of C-peptide, nitric oxide and oxidative stress. Int. J. Clin. Exp. Med. 2011, 2, 136–147. [Google Scholar]
- Vuksan, V.; Sievenpiper, J.L.; Koo, V.Y.Y.; Francis, T.; Beljan-Zdravkovic, U.; Xu, Z.; Vidgen, E. American ginseng (Panax quinquefolius L) reduces postprandial glycemia in nondiabetic subjects and subjects with type 2 diabetes mellitus. Arch. Intern. Med. 2000, 160, 1009–1013. [Google Scholar] [CrossRef]
- Vuksan, V.; Sievenpiper, J.L.; Wong, J.; Xu, Z.; Beljan-Zdravkovic, U.; Arnason, J.T.; Assinewe, V.; Stavro, M.P.; Jenkins, A.L.; Leiter, L.A.; Francis, T. American ginseng (Panax quinquefolius L.) attenuates postprandial glycemia in a time-dependent but not dose-dependent manner in healthy individuals. Am. J. Clin. Nutr. 2001, 73, 753–758. [Google Scholar] [CrossRef]
- De Souza, L.R.; Jenkins, A.L.; Jovanovski, E.; Rahelić, D.; Vuksan, V. Ethanol extraction preparation of American ginseng (Panax quinquefolius L.) and Korean red ginseng (Panax ginseng C.A. Meyer): Differential effects on postprandial insulinemia in healthy individuals. J. Ethnopharm. 2015, 159, 55–61. [Google Scholar] [CrossRef]
- Mucalo, I.; Jovanovski, E.; Vuksan, V.; Dikov, V.B.; Romi, C.; Raheli, D. American ginseng extract (Panax quinquefolius L.) is safe in long-term use in type 2 diabetic patients. Evid. Based Complement. Altern. Med. 2014, 2014, 969168. [Google Scholar] [CrossRef] [PubMed]
- Vuksan, V.; Xu, Z.Z.; Jovanovski, E.; Jenkins, A.L.; Beljan-Zdravkovic, U.; Sievenpiper, J.L.; Stavro, P.M.; Zurbau, A.; Duvnjak, L.; Li, M.Z.C. Efficacy and safety of American ginseng (Panax quinquefolius L.) extract on glycemic control and cardiovascular risk factors in individuals with type 2 diabetes: A double-blind, randomized, crossover clinical trial. Eur. J. Nutr. 2018, 58, 1237–1245. [Google Scholar] [CrossRef]
- Jenkins, A.L.; Morgan, L.M.; Bishop, J.; Jovanovski, E.; Jenkins, D.J.A.; Vuksan, V. Co-administration of a konjac-based fibre blend and American ginseng (Panax quinquefolius L.) on glycaemic control and serum lipids in type 2 diabetes: A randomized controlled, cross-over clinical trial. Eur. J. Nutr. 2018, 57, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Virgous, C.; Si, H. Ginseng and obesity: Observations and understanding in cultured cells, animals and humans. J. Nutr. Biochem. 2017, 44, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zheng, Y.; Han, L.; Wang, H.; Saito, M.; Ling, M.; Kimura, Y.; Feng, Y. Saponins (ginsenosides) from stems and leaves of Panax quinquefolium prevented high-fat diet-induced obesity in mice. Phytomedicine 2008, 15, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, J.-Z.; Liu, W.-C.; Zheng, Y.-N. Anti-obesity effects of protopanaxatriol type ginsenosides isolated from American ginseng leaves in mice fed a high-fat diet. Int. J. Biomed. Pharm. Sci. 2012, 6, 106–112. [Google Scholar] [CrossRef]
- Yeo, C.R.; Lee, S.-M.; Popovich, D.G. Ginseng (Panax quinquefolius) Reduces cell growth, lipid acquisition and increases adiponectin expression in 3T3-L1 cells. Evid. Based Complem. Altern. Med. 2011, 2011, 610625. [Google Scholar] [CrossRef]
- Hwang, J.T.; Lee, M.S.; Kim, H.J.; Sung, M.J.; Kim, H.Y.; Kim, M.S.; Kwon, D.Y. Antiobesity effect of ginsenoside Rg3 involves the AMPK and PPAR-γ signal pathways. Phytotherapy Res. 2009, 23, 262–266. [Google Scholar] [CrossRef]
- Wu, G.Y.; Yi, J.Y.; Liu, L.; Wang, P.C.; Zhang, Z.J.; Li, Z. Pseudoginsenoside F11, a novel partial PPAR gamma agonist, promotes adiponectin oligomerization and secretion in 3T3-L1 adipocytes. PPAR Res. 2013, 2013, 701017. [Google Scholar] [CrossRef]
- Wilson, S.A.F.; Wong, M.H.T.; Stryjecki, S.; De Boer, A.; Lui, E.M.K.; Mutch, D.M. Unraveling the adipocyte inflammomodulatory pathways activated by North American ginseng. Int. J. Obes. 2013, 3711, 350–356. [Google Scholar] [CrossRef]
- Garbett, J.; Wilson, S.A.F.; Ralston, J.C.; De Boer, A.A.; Lui, E.M.K.; Wright, D.C.; Mutch, D.M. North American ginseng influences adipocyte macrophage crosstalk regulation of inflammatory gene expression. J. Ginseng. Res. 2016, 40, 141–150. [Google Scholar] [CrossRef]
- Singh, R.K.; Lui, E.; Wright, D.; Taylor, A.; Bakovic, M. Alcohol extract of North American ginseng (Panax quinquefolius) reduces fatty liver, dyslipidemia, and other complications of metabolic syndrome in a mouse model. Can. J. Physiol. Pharmacol. 2017, 95, 1046–1057. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Xing, N.; Han, D.; Kuang, H.; Ge, P. American ginseng regulates gene expression to protect against premature ovarian failure in rats. Biomed Res. Int. 2015, 2015, 767124. [Google Scholar] [CrossRef]
- Fernandez-Moriano, C.; Gonzalez-Burgos, E.; Iglesias, I.; Lozano, R.; Gomez-Serranillos, M.P. Evaluation of the adaptogenic potential exerted by ginsenosides Rb1 and Rg1 against oxidative stress-mediated neurotoxicity in an in vitro neuronal model. PLoS ONE 2017, 12, e0182933. [Google Scholar] [CrossRef]
- Murphy, L.L.; Cadena, R.S.; Chavez, D.; Ferraro, J.S. Effect of American ginseng (Panax quinquefolium) on male copulatory behavior in the rat. Physiol. Behav. 1998, 64, 445–450. [Google Scholar] [CrossRef]
- Cai, W.; Zhang, K.; Li, P.; Zhu, L.; Xu, J.; Yang, B.; Hu, X.; Lu, Z.; Chen, J. Dysfunction of the neurovascular unit in ischemic stroke and neurodegenerative diseases: An aging effect. Ageing Res. Rev. 2017, 34, 77–87. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wallace, B.K.; Yuen, N.; Jenkins, D.P.; Wulff, H.; O’Donnell, M.E. Blood-brain barrier KCa3.1 channels: Evidence for a role in brain Na uptake and edema in ischemic stroke. Stroke 2015, 46, 237–244. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K.; Kipp, M. Oxidative stress in multiple sclerosis: Central and peripheral mode of action. Exp. Neurol. 2016, 277, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.; Iliffe, S. Alzheimer’s disease. BMJ 2009, 338, b158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Erikson, C.; Brun, A. Cortical synaptic changes and gliosis in normal aging, Alzheimer’s disease and frontal lobe degeneration. Dementia 1996, 7, 128–134. [Google Scholar] [CrossRef]
- Yang, L.; Hao, J.; Zhang, J.; Xia, W.; Dong, X.; Hu, X.; Kong, F.; Cui, X. Ginsenoside Rg3 promotes beta-amyloid peptide degradation by enhancing gene expression of neprilysin. J. Pharm. Pharmacol. 2009, 61, 375–380. [Google Scholar] [CrossRef]
- Al-Hazmi, M.A.; Rawi, S.M.; Arafa, N.M.; Wagas, A.; Montasser, A.O. The potent effects of ginseng root extract and memantine on cognitive dysfunction in male albino rats. Toxicol. Ind. Health 2015, 31, 494–509. [Google Scholar] [CrossRef]
- Nitta, H.; Matsumoto, K.; Shimizu, M.; Ni, X.H.; Watanabe, H. Panax ginseng extract improves the performance of aged Fischer 344 rats in radial maze task but not in operant brightness discrimination task. Biol. Pharm. Bull. 1995, 18, 1286–1288. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef]
- Jin, K. Modern biological theories of aging. Aging Dis. 2010, 1, 72–74. [Google Scholar]
- Shi, S.; Shi, R.; Hashizume, K. American ginseng improves neurocognitive function in senescence-accelerated mice: Possible role of the upregulated insulin and choline acetyltransferase gene expression. Geriatr. Gerontol. Int. 2012, 12, 123–130. [Google Scholar] [CrossRef]
- Sienkiewicz, M.; Głowacka, A.; Kowalczyk, E.; Kochan, E. The activity of different extracts from Panax quinquefolium L. cultures against pathogenic Staphylococcus aureus with respect to ginsenoside content. Arch. Biol. Sci. 2015, 67, 1277–1284. [Google Scholar] [CrossRef]
- Wang, L.; Yang, X.; Yu, X.; Yao, Y.; Ren, G. Evaluation of antibacterial and anti-inflammatory activities of less polar ginsenosides produced from polar ginsenosides by heat-transformation. J. Agric. Food Chem. 2013, 61, 12274–12282. [Google Scholar] [CrossRef]
- Alipour, M.; Omri, A.; Suntres, Z.E. Ginseng aqueous extract attenuates the production of virulence factors, stimulates twitching and adhesion, and eradicates biofilms of Pseudomonas aeruginosa. Can. J. Physiol. Pharmacol. 2011, 89, 419–427. [Google Scholar] [CrossRef]
- Xue, P.; Yao, Y.; Yang, X.S.; Feng, J.; Ren, G.X. Improved antimicrobial effect of ginseng extract by heat transformation. J. Ginseng Res. 2017, 41, 180–187. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B. Quinqueginsin, a novel protein with anti-human immunodeficiency virus, antifungal, ribonuclease and cell-free translation-inhibitory activities from American ginseng roots. Biochem. Biophys. Res. Commun. 2000, 269, 203–208. [Google Scholar] [CrossRef]
- Wang, C.Z.; Du, G.J.; Zhang, Z.; Wen, X.D.; Calway, T.; Zhen, Z.; Musch, M.W.; Bissonnette, M.; Chang, E.B.; Yuan, C.S. Ginsenoside compound K, not Rb1, possesses potential chemopreventive activities in human colorectal cancer. Int. J. Oncol. 2012, 40, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Gut microbiota-mediated pharmacokinetics of ginseng saponins. J. Ginseng Res. 2017, 42, 255–263. [Google Scholar] [CrossRef]
- Wan, J.Y.; Liu, P.; Wang, H.Y.; Qi, L.W.; Wang, C.Z.; Li, P.; Yuan, C.S. Biotransformation and metabolic profile of American ginseng saponins with human intestinal microflora by liquid chromatography quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2013, 1286, 83–92. [Google Scholar] [CrossRef]
- Wan, J.Y.; Wang, C.Z.; Liu, Z.; Zhang, Q.H.; Musch, M.W.; Bissonnette, M.; Chang, E.B.; Li, P.; Qi, L.W.; Yuan, C.S. Determination of American ginseng saponins and their metabolites in human plasma, urine and feces samples by liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1015–1016, 62–73. [Google Scholar] [CrossRef]
- Cimo, A.; Soltani, M.; Lui, E.; Hekmat, S. Fortification of probiotic yogurt with ginseng (Panax quinquefolius) extract. J. Food Nutr. Disor. 2013, 2, 2. [Google Scholar] [CrossRef]
- Wang, C.Z.; Yu, C.; Wen, X.D.; Chen, L.; Zhang, C.F.; Calway, T.; Qiu, Y.; Wang, Y.; Zhang, Z.; Anderson, S.; et al. American ginseng attenuates colitis-associated colon carcinogenesis in mice: Impact on gut microbiota and metabolomics. Cancer Prev. Res. 2016, 9, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Zhang, Y.; Zhao, H.; Wang, J.; Gao, X.; Chen, J.; Fu, B.; Shen, Y.; Miao, F.; Zhang, J.; Teng, G. Non-Invasive monitoring of CNS MHC-I molecules in ischemic stroke mice. Theranostics 2017, 7, 2837–2848. [Google Scholar] [CrossRef]
- Chua, Y.T.; Ang, X.L.; Zhong, X.M.; Khoo, K.S. Interaction between warfarin and Chinese herbal medicines. Singapore Med. J. 2015, 56, 11–18. [Google Scholar] [CrossRef]
- Assinewe, V.V.; Amason, J.T.; Aubry, A.; Mullin, J.; Lemaire, I. Extractable polysaccharides of Panax quinquefolius L. (North American ginseng) root stimulate TNFalpha production by alveolar macrophages. Phytomedicine 2002, 9, 398–404. [Google Scholar] [CrossRef]
- Yu, X.H.; Liu, Y.; Wu, X.L.; Liu, L.Z.; Fu, W.; Song, D.D. Isolation, purification, characterization and immunostimulatory activity of polysaccharides derived from American ginseng. Carbohydr. Polym. 2017, 156, 9–18. [Google Scholar] [CrossRef]
- Lui, E.M.K.; Azike, C.G.; Guerrero-Analco, J.A.; Romeh, A.A.; Pei, P.; Kaldas, S.J.; Arnason, J.T.; Charpentier, P.A. Bioactive polysaccharides of American ginseng Panax quinquefolius L. In Modulation of Immune Function: Phytochemical and Pharmacological Characterization, The Complex World of Polysaccharides; Karunaratne, D.N., Ed.; Intech Open: London, UK, 2012; pp. 513–518. [Google Scholar]
- Azike, C.G. American ginseng modulation of immune function and phytochemical analysis, Doctor of Philosophy, The University of Western Ontario, London, Ontario, Canada. May 2014. Available online: https://ir.lib.uwo.ca/cgi/viewcontent.cgi?article=3449&context=etd (accessed on 26 April 2019).
- Lü, J.M.; Yao, Q.; Chen, C. Ginseng compounds: An update on their molecular mechanisms and medical applications. Curr. Vasc. Pharmacol. 2009, 7, 293–302. [Google Scholar] [CrossRef]
- Kim, J.H. Cardiovascular Diseases and Panax ginseng: A Review on molecular mechanisms and medical applications. J. Ginseng Res. 2012, 36, 16–26. [Google Scholar] [CrossRef]
- Sodrul, I.M.D.; Wang, C.; Chen, X.; Du, J.; Sun, H. Role of ginsenosides in reactive oxygen species-mediated anticancer therapy. Oncotarget 2018, 9, 2931–2950. [Google Scholar] [CrossRef]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a North American ginseng extract. Mol. Cell. Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef]
- Chan, L.Y.; Chiu, P.Y.; Lau, T.K. An in-vitro study of ginsenoside Rb1-induced teratogenicity using a whole rat embryo culture model. Hum. Reprod. 2003, 18, 2166–3268. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xu, Y.; Yin, H.; Wang, J.; Chen, K.; Li, Y. Developmental toxicity research of ginsenoside Rb1 using a whole mouse embryo culture model. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 74, 207–209. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pharmacological Action | Ginsenoside | Reference |
|---|---|---|
| Positively affects memory processes; induces synthesis of acetylcholine in the hippocampus by stimulating choline acetyltransferase; induces apoptosis and inhibits angiogenesis in cancer cells; inhibits the release of inflammatory leukotrienes; reversibly and tonically blocks voltage-dependent Na+ channels in the brain reducing detrimental effects of hypoxia; downregulates the COX-2 gene; stabilises neutrophils and lymphocytes; inhibits the release of histamine; blocks calcium channels and stabilised the heart; reduces blood sugar levels; anti-diabetic, insulin-sensitising and anti-obesity actions; neurotropic, neuroprotective, oestrogen-like activity; stimulates GABA receptors and induces a depressive effect on brain function, which underlines its calming, anxiolytic, sleeping, relaxing and antipsychotic effects | Rb1, Rb2, Rc | [9,22,53,54,55] |
| Stimulates superoxide dismutase; inhibits angiogenesis in cancer; prevents diabetes; lowers cholesterol and triglycerides levels, activates lipolysis; corticotropic and oestrogenic activity | Rb2 | [9,54,55] |
| Inhibits proliferation of breast cancer cells; induces corticotropic effects | Rc | [9,22,54] |
| Promotes neurites outgrowth, an important process for neuronal repair; induces corticotropic effects | Rd, Rc, Re | [9,22,54,55,56,57] |
| Scavenges hydroxyl radicals and degrades H2O2; reduces blood sugar levels; induces cardioprotective effects; activates cGMP and relaxes smooth muscles | Re | [9,22,54,55,56] |
| Downregulates the COX-2 gene; stabilises neutrophils and lymphocytes; inhibits histamine release; inhibits platelet-induced activation of thromboxane; increases insulin receptors; increases T-helper lymphocytes; inhibits release of endothelin and relaxation of the smooth muscle of blood vessels; activates cyclic guanosine monophosphate and relaxes the smooth muscle (hypotensive effect); blocks calcium channels and stabilised the heart; reduces blood sugar levels | Rg1 | [22,55,56] |
| Inhibits neuronal acetylcholine | Rg2 | [22] |
| Inhibits platelet aggregation induced by thrombin; relaxes the smooth muscle of the blood vessels by activating the K+ channels and releases Ca2+; inhibits progression of tumours and reduces drug resistance of cancer cells; inhibits endothelin and relaxation of the smooth muscle of blood vessels; induces hypotensive effect; downregulates the COX-2 gene; stabilises neutrophils and lymphocytes; inhibits histamine release; modulates mitogen-activated protein kinases, thus inhibiting the spread of cancer cells | Rg3 | [22,53,55,56,58] |
| Activates oestrogen receptor; inhibits proliferation of cancer cells and induces apoptosis | Rh1 | [22,58] |
| Inhibits breast, liver and prostate cancer cells proliferation | Rh2 | [22,58] |
| Assists memory improvement; induces neuroprotective effects | Pseudoginse-noside F11 | [22] |
| Effect | Reference |
|---|---|
| Mitigation of symptoms of Alzheimer disease in mice by AChE-acetyltransferase upregulation | [64] |
| Anxiolytic effect without influence on locomotion abilities in mice | [66] |
| Improvement of cognitive performance, predominantly working memory and calmness in young adults | [67] |
| Neuroprotective effect on tsA-201 cells transfected with cDNA expressing α subunits of the Brain2a Na+ channel during ischaemia by blocking Na+ channel | [68] |
| Rg1 mitigation of memory impairment in mice and increase of hippocampal excitability in anaesthetised rats | [69] |
| Mitigation of anxiety caused by methamphetamine abuse in rats | [70] |
| Decrease of iNOS synthase and demyelination scores in the central nervous system in mice with experimental autoimmune encephalomyelitis | [71] |
| Protection against scopolamine-induced memory deficits in rats | [72] |
| Neuroprotection of pseudoginsenoside F11 on 6-hydroxydopamine-induced Parkinson's disease in rats | [73] |
| Inhibition of neuronal apoptosis and decrease of neurite damage via regulation of endoplasmic reticulum stress after acute spinal cord injury in rats | [74] |
| F11 inhibition of hyper locomotion and increase of extracellular dopamine release by GABAergic neurons and μ-opioid receptors regulation in mice | [75] |
| Modulation of brain function by inhibition of neuronal discharge frequency in brainstem unitary activity in rats | [76] |
| Neuroprotection on anxiety-like behaviour induced by sleep deprivation in mice | [77] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczuka, D.; Nowak, A.; Zakłos-Szyda, M.; Kochan, E.; Szymańska, G.; Motyl, I.; Blasiak, J. American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties. Nutrients 2019, 11, 1041. https://doi.org/10.3390/nu11051041
Szczuka D, Nowak A, Zakłos-Szyda M, Kochan E, Szymańska G, Motyl I, Blasiak J. American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties. Nutrients. 2019; 11(5):1041. https://doi.org/10.3390/nu11051041
Chicago/Turabian StyleSzczuka, Daria, Adriana Nowak, Małgorzata Zakłos-Szyda, Ewa Kochan, Grażyna Szymańska, Ilona Motyl, and Janusz Blasiak. 2019. "American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties" Nutrients 11, no. 5: 1041. https://doi.org/10.3390/nu11051041
APA StyleSzczuka, D., Nowak, A., Zakłos-Szyda, M., Kochan, E., Szymańska, G., Motyl, I., & Blasiak, J. (2019). American Ginseng (Panax quinquefolium L.) as a Source of Bioactive Phytochemicals with Pro-Health Properties. Nutrients, 11(5), 1041. https://doi.org/10.3390/nu11051041