Biotin Is Required for the Zinc Homeostasis in the Skin


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Approval
2.2. Animals and Diets
2.3. Reagents and Antibodies
2.4. Quantification of Zinc Levels in the Serum
2.5. Histological Examination
2.6. Preparation of Epidermal Cell Suspensions
2.7. Flow Cytometry
2.8. Quantification of ATP Release from the Skin
2.9. ACD and ICD Responses
2.10. Quantitative Real-Time PCR Analysis
2.11. Statistics
3. Results
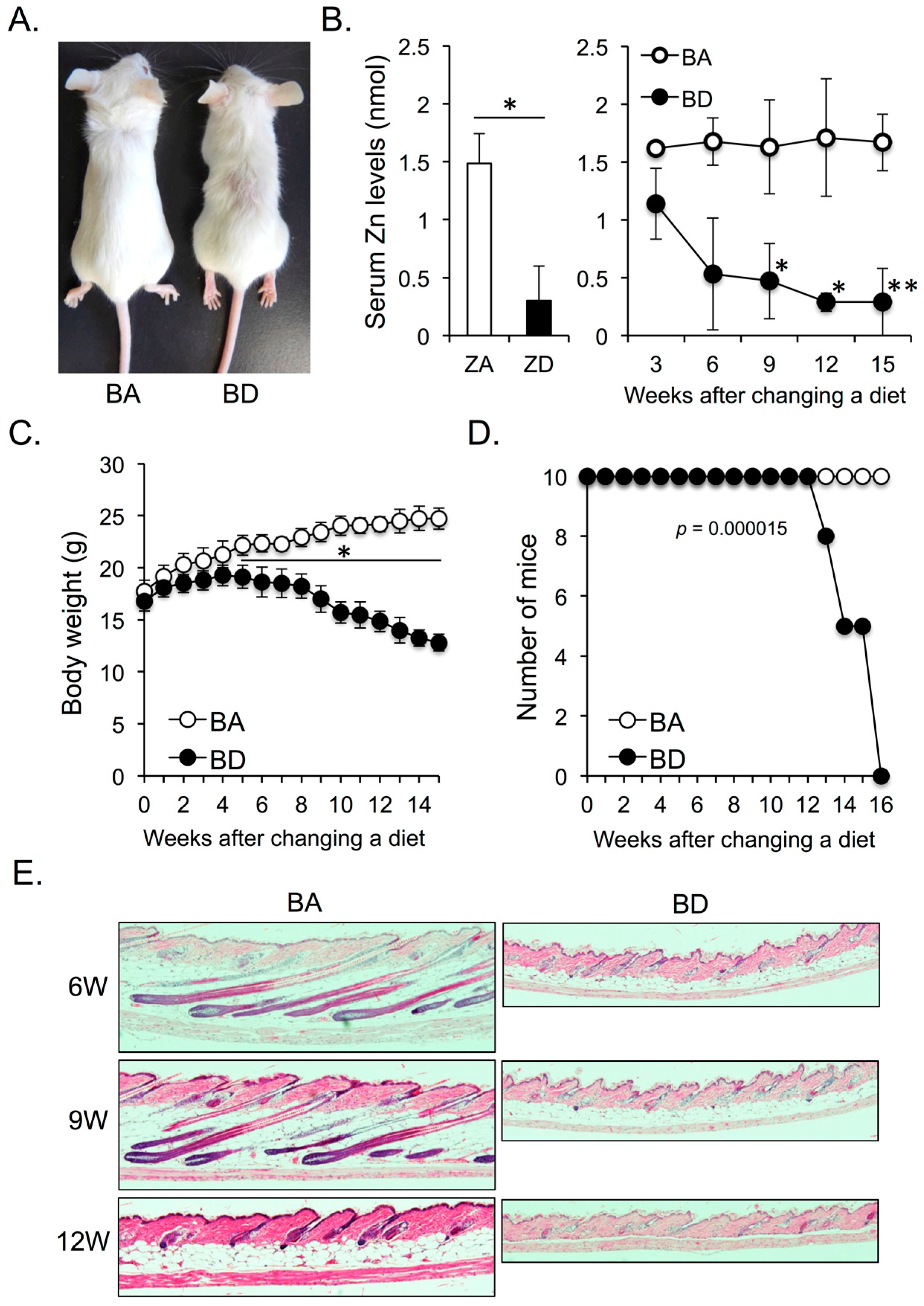
3.1. Dietary Biotin Deficiency Leads to Zinc Deficiency
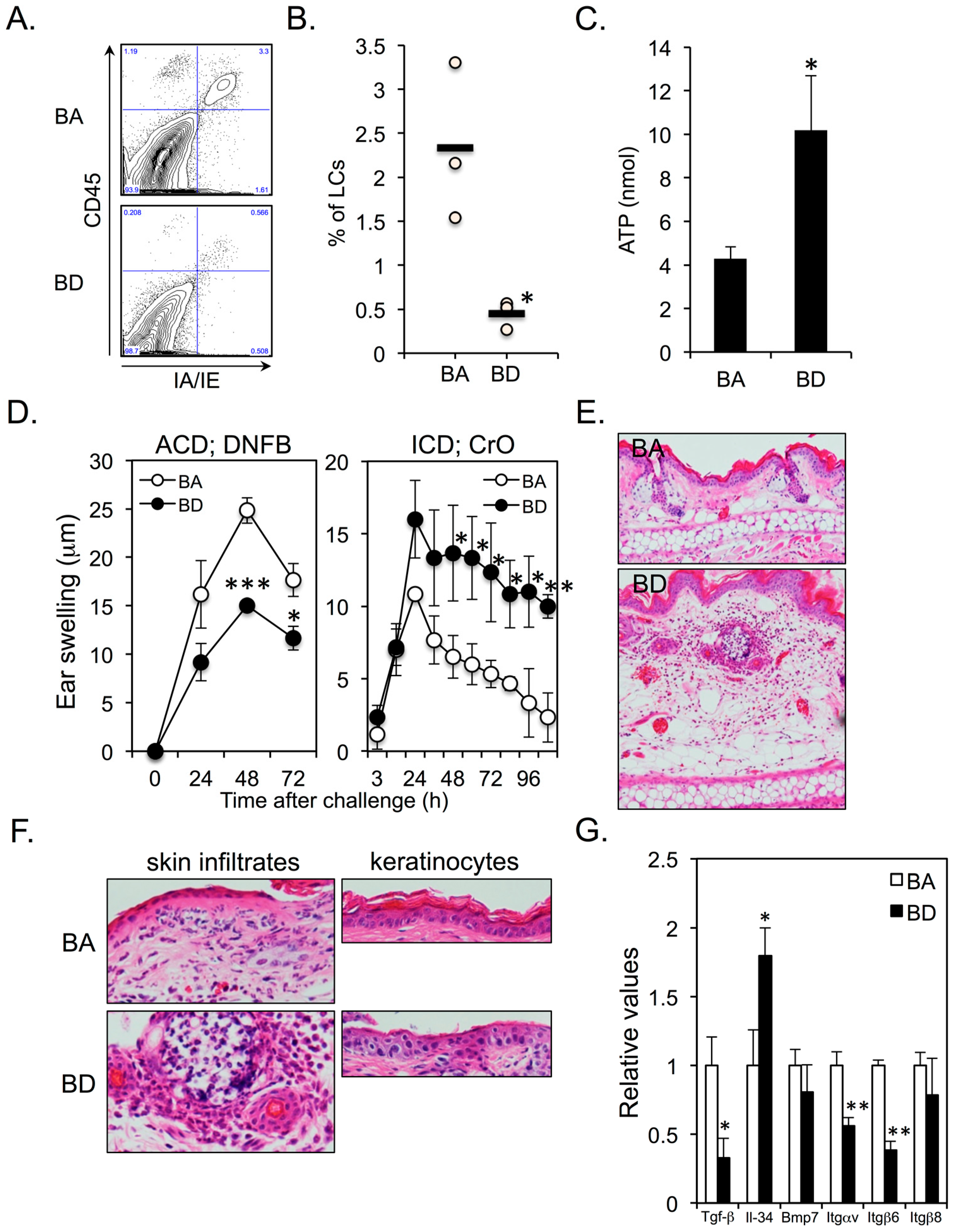
3.2. Dietary Biotin Deficiency Leads to the Development of ATP-Mediated Irritant Contact Dermatitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Leon-Del-Rio, A. Biotin in metabolism, gene expression, and human disease. J. Inherit. Metab. Dis. 2019. [Google Scholar] [CrossRef]
- Zempleni, J.; Wijeratne, S.S.; Hassan, Y.I. Biotin. Biofactors 2009, 35, 36–46. [Google Scholar] [CrossRef]
- Zempleni, J.; Hassan, Y.I.; Wijeratne, S.S. Biotin and biotinidase deficiency. Expert Rev. Endocrinol. Metab. 2008, 3, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.M. Skin manifestations of biotin deficiency. Semin. Dermatol. 1991, 10, 296–302. [Google Scholar]
- Wang, K.; Pugh, E.W.; Griffen, S.; Doheny, K.F.; Mostafa, W.Z.; al-Aboosi, M.M.; el-Shanti, H.; Gitschier, J. Homozygosity mapping places the acrodermatitis enteropathica gene on chromosomal region 8q24.3. Am. J. Hum. Genet. 2001, 68, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, B.; Kuo, Y.M.; Zemansky, J.; Gitschier, J. A novel member of a zinc transporter family is defective in acrodermatitis enteropathica. Am. J. Hum. Genet. 2002, 71, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Kury, S.; Dreno, B.; Bezieau, S.; Giraudet, S.; Kharfi, M.; Kamoun, R.; Moisan, J.P. Identification of SLC39A4, a gene involved in acrodermatitis enteropathica. Nat. Genet. 2002, 31, 239–240. [Google Scholar] [CrossRef]
- Kawamura, T.; Ogawa, Y.; Nakamura, Y.; Nakamizo, S.; Ohta, Y.; Nakano, H.; Kabashima, K.; Katayama, I.; Koizumi, S.; Kodama, T.; et al. Severe dermatitis with loss of epidermal Langerhans cells in human and mouse zinc deficiency. J. Clin. Invest. 2012, 122, 722–732. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kawamura, T.; Shimada, S. Zinc and skin biology. Arch. Biochem. Biophys. 2016, 611, 113–119. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kinoshita, M.; Shimada, S.; Kawamura, T. Zinc and Skin Disorders. Nutrients 2018, 10, 199. [Google Scholar] [CrossRef]
- Mizumoto, N.; Mummert, M.E.; Shalhevet, D.; Takashima, A. Keratinocyte ATP release assay for testing skin-irritating potentials of structurally diverse chemicals. J. Invest. Dermatol. 2003, 121, 1066–1072. [Google Scholar] [CrossRef]
- Kashem, S.W.; Haniffa, M.; Kaplan, D.H. Antigen-Presenting cells in the skin. Annu. Rev. Immunol. 2017, 35, 469–499. [Google Scholar] [CrossRef]
- Georgiou, J.G.; Skarratt, K.K.; Fuller, S.J.; Martin, C.J.; Christopherson, R.I.; Wiley, J.S.; Sluyter, R. Human epidermal and monocyte-derived langerhans cells express functional P2X receptors. J. Invest. Dermatol. 2005, 125, 482–490. [Google Scholar] [CrossRef]
- Ho, C.L.; Yang, C.Y.; Lin, W.J.; Lin, C.H. Ecto-nucleoside triphosphate diphosphohydrolase 2 modulates local ATP-induced calcium signaling in human HaCaT keratinocytes. PLoS ONE 2013, 8, e57666. [Google Scholar] [CrossRef]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sevigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell-associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Kinoshita, M.; Mizumura, N.; Miyazaki, S.; Aoki, R.; Momosawa, A.; Shimada, S.; Kambe, T.; Kawamura, T. Purinergic molecules in the epidermis. J. Investig. Dermatol. 2018, 138, 2486–2488. [Google Scholar] [CrossRef] [PubMed]
- Maverakis, E.; Fung, M.A.; Lynch, P.J.; Draznin, M.; Michael, D.J.; Ruben, B.; Fazel, N. Acrodermatitis enteropathica and an overview of zinc metabolism. J. Am. Acad. Dermatol. 2007, 56, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, T.A.; Letterio, J.J.; Farr, A.G.; Udey, M.C. A role for endogenous transforming growth factor beta 1 in Langerhans cell biology: The skin of transforming growth factor beta 1 null mice is devoid of epidermal Langerhans cells. J. Exp. Med. 1996, 184, 2417–2422. [Google Scholar] [CrossRef]
- Wang, Y.; Szretter, K.J.; Vermi, W.; Gilfillan, S.; Rossini, C.; Cella, M.; Barrow, A.D.; Diamond, M.S.; Colonna, M. IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nat. Immunol. 2012, 13, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, N.; Bauer, T.; Modak, M.; Wagner, K.; Schuster, C.; Koffel, R.; Seyerl, M.; Stockl, J.; Elbe-Burger, A.; Graf, D.; et al. Identification of bone morphogenetic protein 7 (BMP7) as an instructive factor for human epidermal Langerhans cell differentiation. J. Exp. Med. 2013, 210, 2597–2610. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Mu, Z.; Dabovic, B.; Jurukovski, V.; Yu, D.; Sung, J.; Xiong, X.; Munger, J.S. Absence of integrin-mediated TGFbeta1 activation in vivo recapitulates the phenotype of TGFbeta1-null mice. J. Cell Biol. 2007, 176, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Aluwihare, P.; Mu, Z.; Zhao, Z.; Yu, D.; Weinreb, P.H.; Horan, G.S.; Violette, S.M.; Munger, J.S. Mice that lack activity of alphavbeta6- and alphavbeta8-integrins reproduce the abnormalities of Tgfb1- and Tgfb3-null mice. J. Cell Sci. 2009, 122, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.M. Evidence for a pathogenic role of omega 6 polyunsaturated fatty acid in the cutaneous manifestations of biotin deficiency. J. Pediatr. Gastroenterol. Nutr. 1990, 10, 222–229. [Google Scholar] [CrossRef]
- Proud, V.K.; Rizzo, W.B.; Patterson, J.W.; Heard, G.S.; Wolf, B. Fatty acid alterations and carboxylase deficiencies in the skin of biotin-deficient rats. Am. J. Clin. Nutr. 1990, 51, 853–858. [Google Scholar] [CrossRef]
- Da Silva, G.L.; Sperotto, N.D.; Borges, T.J.; Bonorino, C.; Takyia, C.M.; Coutinho-Silva, R.; Campos, M.M.; Zanin, R.F.; Morrone, F.B. P2X7 receptor is required for neutrophil accumulation in a mouse model of irritant contact dermatitis. Exp. Dermatol. 2013, 22, 184–188. [Google Scholar] [CrossRef]
- Matsusue, S.; Kashihara, S.; Takeda, H.; Koizumi, S. Biotin deficiency during total parenteral nutrition: Its clinical manifestation and plasma nonesterified fatty acid level. JPEN J. Parenter. Enteral. Nutr. 1985, 9, 760–763. [Google Scholar] [CrossRef]
- Higuchi, R.; Mizukoshi, M.; Koyama, H.; Kitano, N.; Koike, M. Intractable diaper dermatitis as an early sign of biotin deficiency. Acta Paediatr. 1998, 87, 228–229. [Google Scholar] [CrossRef]
- Lagier, P.; Bimar, P.; Seriat-Gautier, S.; Dejode, J.M.; Brun, T.; Bimar, J. Zinc and biotin deficiency during prolonged parenteral nutrition in the infant. Presse. Med. 1987, 16, 1795–1797. [Google Scholar] [PubMed]
- Khalidi, N.; Wesley, J.R.; Thoene, J.G.; Whitehouse, W.M., Jr.; Baker, W.L. Biotin deficiency in a patient with short bowel syndrome during home parenteral nutrition. JPEN J. Parenter. Enteral. Nutr. 1984, 8, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Noda, E.; Koyama, Y.; Shirai, T.; Horino, A.; Juri, T.; Koike, M. Biotin deficiency in an infant fed with amino acid formula and hypoallergenic rice. Acta. Paediatr. 1996, 85, 872–874. [Google Scholar] [CrossRef]
- Fujimoto, W.; Inaoki, M.; Fukui, T.; Inoue, Y.; Kuhara, T. Biotin deficiency in an infant fed with amino acid formula. J. Dermatol. 2005, 32, 256–261. [Google Scholar] [CrossRef] [PubMed]
- El-Esawy, F.M.; Hussein, M.S.; Ibrahim Mansour, A. Serum biotin and zinc in male androgenetic alopecia. J. Cosmet. Dermatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta. 2008, 1783, 673–694. [Google Scholar] [CrossRef]
- Zimmermann, H. Extracellular metabolism of ATP and other nucleotides. Naunyn. Schmiedebergs. Arch. Pharmacol. 2000, 362, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.A.; Miyazaki, S.; Kobayashi, M.; Nishino, K.; Goto, T.; Matsunaga, M.; Ooi, M.; Shirakawa, H.; Tani, F.; Kawamura, T.; et al. Zinc deficiency causes delayed ATP clearance and adenosine generation in rats and cell culture models. Commun. Biol. 2018, 1, 113. [Google Scholar] [CrossRef]
- Kaplan, D.H. Ontogeny and function of murine epidermal Langerhans cells. Nat. Immunol. 2017, 18, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, J.; Beura, L.K.; Bobr, A.; Astry, B.; Chicoine, B.; Kashem, S.W.; Welty, N.E.; Igyarto, B.Z.; Wijeyesinghe, S.; Thompson, E.A.; et al. Stromal cells control the epithelial residence of DCs and memory T cells by regulated activation of TGF-beta. Nat. Immunol. 2016, 17, 414–421. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogawa, Y.; Kinoshita, M.; Sato, T.; Shimada, S.; Kawamura, T. Biotin Is Required for the Zinc Homeostasis in the Skin. Nutrients 2019, 11, 919. https://doi.org/10.3390/nu11040919
Ogawa Y, Kinoshita M, Sato T, Shimada S, Kawamura T. Biotin Is Required for the Zinc Homeostasis in the Skin. Nutrients. 2019; 11(4):919. https://doi.org/10.3390/nu11040919
Chicago/Turabian StyleOgawa, Youichi, Manao Kinoshita, Takuya Sato, Shinji Shimada, and Tatsuyoshi Kawamura. 2019. "Biotin Is Required for the Zinc Homeostasis in the Skin" Nutrients 11, no. 4: 919. https://doi.org/10.3390/nu11040919
APA StyleOgawa, Y., Kinoshita, M., Sato, T., Shimada, S., & Kawamura, T. (2019). Biotin Is Required for the Zinc Homeostasis in the Skin. Nutrients, 11(4), 919. https://doi.org/10.3390/nu11040919