Anaerobic Performance after a Low-Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Male Basketball Players


 ,
,  , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Participants
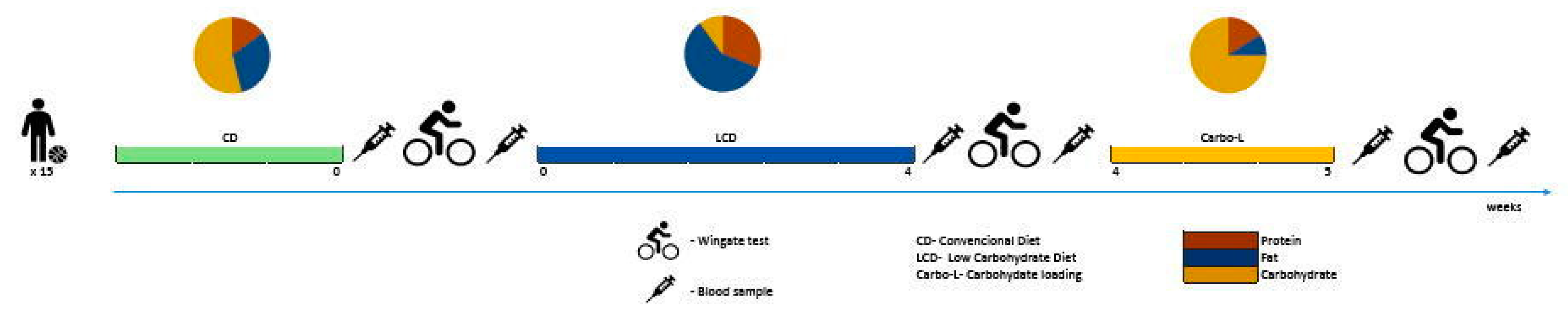
2.2. Experimental Design
2.2.1. Dietary Guidelines—Monitoring of Nutritional Intake
2.2.2. Training Program
2.3. Body Mass, Body Composition
2.4. Anaerobic Performance
2.5. Biochemical Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coyle, E.F.; Coggan, A.R.; Hemmert, M.K.; Ivy, J.L. Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. Appl. Physiol. 1986, 61, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Neufer, P.D.; Costill, D.L.; Flynn, M.G.; Kirwan, J.P.; Mitchell, J.B.; Houmard, J. Improvements in exercise performance: Effects of carbohydrate feedings and diet. J. Appl. Physiol. 1987, 62, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M. Exercise, muscle, and CHO metabolism. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. 4), 29–33. [Google Scholar] [CrossRef]
- Hawley, J.A.; Burke, L.M. Carbohydrate availability and training adaptation: Effect on cell metabolism. Sport Sci. Res. 2010, 38, 152–160. [Google Scholar] [CrossRef]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Endocrinol. Metab. 1993, 265, E380–E391. [Google Scholar] [CrossRef]
- Margolis, L.M.; Pasiakos, S.M. Optimizing intramuscular adaptations to aerobic exercise: Effects of carbohydrate restriction and protein supplementation on mitochondrial biogenesis. Adv. Nutr. 2013, 4, 657–664. [Google Scholar] [CrossRef]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef]
- Phinney, S.D.; Bistrian, B.R.; Evans, W.J.; Gervino, E.; Blackburn, G.L. The human metabolic response to chronic ketosis without caloric restriction: Preservation of submaximal exercise capability with reduced carbohydrate oxidation. Metabolism 1983, 32, 769–776. [Google Scholar] [CrossRef]
- Langfort, J.; Pilis, W.; Zarzeczny, R.; Nazar, K.; Kaciuba-Uściłko, H. Effect of low-carbohydrate-ketogenic diet on metabolic and hormonal responses to graded exercise in men. J. Physiol. Pharmacol. 1996, 47, 361–371. [Google Scholar] [PubMed]
- Phinney, S.D. Ketogenic diets and physical performance. Nutr. Metab. 2004, 1, 2. [Google Scholar] [CrossRef]
- Jeppesen, J.; Kiens, B. Regulation and limitations to fatty acid oxidation during exercise. J. Physiol. 2012, 590, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Zajac, A.; Poprzecki, S.; Maszczyk, A.; Czuba, M.; Michalczyk, M.; Zydek, G. The effects of a ketogenic diet on exercise metabolism and physical performance in off-road cyclists. Nutrients 2014, 6, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Rhyu, H.S.; Cho, S.Y. The effect of weight loss by ketogenic diet on the body composition, performance-related physical fitness factors and cytokines of Taekwondo athletes. J. Exerc. Rehabil. 2014, 10, 326–331. [Google Scholar] [CrossRef]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef]
- McSwiney, F.T.; Wardrop, B.; Hyde, P.N.; Lafountain, R.A.; Volek, J.S.; Doyle, L. Keto-adaptation enhances exercise performance and body composition responses to training in endurance athletes. Metabolism 2018, 81, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.M.; Hamdan, H.; Torisky, D.M.; Akers, J.D. A low-carbohydrate ketogenic diet combined with 6-weeks of crossfit training improves body composition and performance. Int. J. Sports Exerc. Med. 2017, 3, 1–10. [Google Scholar] [CrossRef]
- Michalczyk, M.; Zajac, A.; Mikolajec, K.; Zydek, G.; Langfort, J. No Modification in Blood Lipoprotein Concentration but Changes in Body Composition after 4 Weeks of Low Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Basketball Players. J. Hum. Kinet. 2018, 65, 125–137. [Google Scholar] [CrossRef]
- Wroble, K.A.; Trott, M.N.; Schweitzer, G.G.; Rahman, R.S.; Kelly, P.V.; Weiss, E.P. Low carbohydrate, ketogenic diet impairs anaerobic exercise performance in exercise-trained women and men: A randomized-sequence crossover trial. J. Sport Med. Phys. Fit. 2018. [Google Scholar] [CrossRef]
- Urbain, P.; Strom, L.; Morawski, L.; Wehrle, A.; Deibert, P.; Bertz, H. Impact of a 6-week non-energy-restricted ketogenic diet on physical fitness, body composition and biochemical parameters in healthy adults. Nutr. Metab. 2017, 14, 17. [Google Scholar] [CrossRef]
- Paoli, A.; Grimaldi, K.; D’Agonisto, D. Ketogenic diet does not affect strength performance in elite artistic gymnasts. J. Int. Soc. Sport Nutr. 2012, 9, 34. [Google Scholar] [CrossRef]
- Langfort, J.; Zarzeczny, R.; Pilis, W.; Nazar, K.; Kaciuba-Uścitko, H. The effect of a low-carbohydrate diet on performance, hormonal and metabolic responses to a 30-s bout of supramaximal exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 76, 128–133. [Google Scholar] [CrossRef]
- Burke, L.M. Re-examinig high-fat diets for sport performance: Did we call the “nail in the coffin” to soon? Sports Med. 2015, 45 (Suppl. 1), S33–S49. [Google Scholar] [CrossRef] [PubMed]
- Havemann, L.; West, S.J.; Goedecke, J.H.; Macdonald, I.A.; St Clair Gibson, A.; Noakes, T.D.; Lambert, E.V. Fat adaptation followed by carbohydrate loading compromises high-intensity sprint performance. J. Appl. Physiol. 2006, 100, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Yeo, W.K.; Carey, A.; Burke, L.M.; Spriet, L.L.; Hawley, J. Fat adaptation in well-trained athletes: Effect of Cell Metabolism. Appl. Physiol. Nutr. Metab. 2011, 36, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Stellingweff, T.; Spriet, L.L.; Watt, M.; Kimber, N.; Hargreaves, M.; Hawley, J.; Burkey, L.M. Decreased PDH activation and glycogenolysis during exercise following fat adaptation with carbohydrate restoration. Am. J. Physiol. Endocrinol. Metab. 2006, 29, E380–E388. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover concept”. J. Appl. Physiol. 1994, 76, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Langfort, J.; Ploug, T.; Ihlemann, J.; Baranczuk, E.; Donsmark, M.; Górski, J.; Galbo, H. Additivity of adrenaline and contractions on hormone-sensitive lipase, but not on glycogen phosphorylase, in rat muscle. Acta Physiol. Scand. 2003, 178, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Bianco, A.; Grimaldi, K.A. The ketogenic diet and sport: A possible marriage? Exerc. Sport Sci. Rev. 2015, 43, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.; Volek, J.S.; Phinney, S.D. Low-carbohydrate diets for athletes: What evidence? Br. J. Sports Med. 2014, 48, 1077–1078. [Google Scholar] [CrossRef] [PubMed]
- Cochran, A.J.; Little, J.P.; Tarnopolsky, M.A.; Gibala, M.J. Carbohydrate feeding during recovery alters the skeletal muscle metabolic response to repeated sessions of high-intensity interval exercise in humans. J. Appl. Physiol. 2010, 108, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Van Praekers, K.; Szlufcik, K.; Nielsen, H.; Ramaekers, M.; Hespel, P. Beneficial metabolic adaptation due to endurance exercise training in the fasted state. J. Appl. Physiol. 2011, 110, 236–245. [Google Scholar]
- Yeo, W.K.; Paton, C.D.; Garnham, A.P.; Burke, L.M.; Carey, A.L.; Hawley, J.A. Skeletal muscle adaptation and performance responses to once a day versus twice every second day endurance training regimes. J. Appl. Physiol. 2008, 105, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Issurin, V.B. Benefits and limitations of block periodized training approaches to athletes’ preparation: A review. Sport Med. 2016, 46, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Issurin, V.B. New horizons for the methodology and physiology of training periodization. Sports Med. 2010, 40, 189–206. [Google Scholar] [CrossRef]
- Jarosz, M.; Traczyk, I.; Rychlik, E. Nutrition Norms for the Polish Population; Institute of Nutrition: Warsaw, Poland, 2012; pp. 18–31. [Google Scholar]
- Ortenblad, N.; Westerblad, H.; Nielsaen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G. Importance of the “crossover” concept in exercise metabolism. Clin. Exerc. Pharm. Physiol. 1997, 124, 889–895. [Google Scholar] [CrossRef]
- Hickson, R.C.; Rennie, M.J.; Conlee, R.K.; Winder, W.W.; Holloszy, J.O. Effects of increase plasma fatty acids on glycogen utilization and endurance. J. Appl. Physiol. 1997, 43, 829–833. [Google Scholar] [CrossRef]
- Erlenbusch, M.; Haub, M.; Munoz, K.; Mac Connie, S.; Stillwell, B. Effect of high-fat or high-carbohyddrate diets on endurance exercise: A meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 1–14. [Google Scholar] [CrossRef]
- Colombani, P.O.; Mannhart, C.; Mettler, S. Carbohydrates and exercise performance in nin-fasted athletes: A systematic review of studies mimicking real-life. Nutr. J. 2013, 12, 16. [Google Scholar] [CrossRef]
- Medbo, J.I.; Tabata, I. Anaerobic energy release in working muscle during 30 s to 3 min of exhausting bicycling. J. Appl. Physiol. 1993, 75, 1654–1660. [Google Scholar] [CrossRef]
- Mullins, G.; Hallam, C.L.; Broom, I. Ketosis, ketoacidosis and very-low-calorie diets: Putting the record straight. Nutr. Bull. 2011, 36, 397–402. [Google Scholar] [CrossRef]
- Fleming, J.; Sharman, M.J.; Avery, N.G.; Love, D.M.; Gómez, A.L.; Scheett, T.P.; Kraemer, W.J.; Volek, J.S. Endurance capacity and high-intensity exercises performance responses to a high-fat diet. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R. Metabolic interactions between glucose and fatty acids in humans. Am. J. Clin. Nutr. 1998, 67, 519S–526S. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kunth, S.T.; Dave, H.; Peters, J.R.; Fitts, R.H. Low cell pH depresses peak power in rat skeletal muscle fibres at both 30 degrees C and 15 degrees C: Implication for muscle fatigue. J. Physiol. 2006, 575, 887–899. [Google Scholar]
- Dipla, K.; Makri, M.; Zafeiridis, A.; Soulas, D.; Tsalouhidou, S.; Mougios, V.; Kellis, S. An isoenergetic high-protein, moderate-fat diet does not compromise strength and fatigue during resistance exercise in women. Br. J. Nutr. 2008, 100, 283–286. [Google Scholar] [CrossRef]
- Grasii, B.; Rossiter, H.B.; Zoladz, J.A. Skeletal muscle fatigue and decreased efficiency: Two sides of the same coin? Exerc. Sport Sci. Rev. 2015, 43, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Korzeniewski, B.; Grassi, B. Training-induced acceleration of oxygen uptake kinetics in skeletal muscle: The underlying mechanisms. J. Physiol. Pharmacol. 2006, 57 (Suppl. 10), 67–84. [Google Scholar]
- Carey, A.L.; Staudacher, H.M.; Cummings, N.K.; Stepto, N.K.; Nikolopoulos, V.; Burke, L.M.; Hawley, J.A. Effects of fat adaptation carbohydrate restoration on prolonged endurance exercise. J. Appl. Phyiol. 2001, 91, 15–22. [Google Scholar] [CrossRef]
- Ormsbee, M.J.; Bach, C.W.; Baur, D.A. Pre-exercise nutrition: The role of macronutrients, modified starches and supplements on metabolism and endurance performance. Nutrients 2014, 6, 1782–1808. [Google Scholar] [CrossRef] [PubMed]
- Holness, M.J.; Kraus, A.; Harris, R.A.; Sugden, M.C. Target upregulation of pyruvate dehydrogenase kinase (PDK)-4 in slow-twitch skeletal muscle underlies the stable modification of the regulatory characteristics of PDK induced by high-fat feeding. Diabetes 2000, 49, 775–781. [Google Scholar] [CrossRef]
- Cox, P.J.; Kirk, T.; Ashmore, T.; Willerton, K.; Evans, R.; Smith, A.; Murray, A.J.; Stubbs, B.; West, J.; McLure, S.W.; et al. Nutritional ketosis alters fuel preference and thereby endurance performance in athletes. Cell Metab. 2016, 24, 256–268. [Google Scholar] [CrossRef] [PubMed]
- De Pergola, G. The adipose tissue metabolism: Role of testosterone and dehydroepiandrosterone. Int. J. Obes. Relat. Metab. Disord. 2000, 24 (Suppl. 2), S59–S63. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Kraemer, W.J.; Bush, J.A.; Incledon, T.; Boetes, M. Testosterone and cortisol in relationship to dietary nutrient band resistance exercise. J. Appl. Physiol. 1997, 82, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Westman, E.C.; Mavropoulos, J.; Yancy, W.S.; Volek, J.S. A review of low-carbohydrate ketogenic diets. Curr. Atheroscler. Rep. 2003, 5, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Berggren, A.; Ehrnborg, C.; Rosen, T.; Ellegard, L.; Bengtsson, B.A.; Caidahl, K. Short-term administration of supraphysiological recombinant human growth hormone (GH) does not increase maximum endurance capacity in healthy, active young men and women with normal GH-insulin-like growth factor I axes. J. Clin. Endocrinol. Metab. 2005, 90, 3268–3273. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variables | After CD | After LCD | After Carbo-L |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| BM (kg) | 92.18 ± 5.17 | 90.38 ± 3.12 * | 91.82 ± 4.32 |
| BMI (kg/m2) | 24.48 ± 0.18 | 23.93 ± 0.17 | 24.19 ± 0.21 |
| FFM (kg) | 79.62 ± 4.88 | 78.20 ± 3.65 | 79.92 ± 3.84 # |
| FM (%) | 12.42 ± 2.25 | 11.1 ± 1.25 * | 11.8 ± 1.23 |
| Nutrients | CD | LCD | Carbo-L |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| Carbohydrates (%) | 54 ± 6.1 | 10 ± 0.5 | 75 ± 3 |
| Protein (%) | 15 ± 6.3 | 31 ± 2.3 | 16 ± 3 |
| Fat (%) | 31 ± 4.3 | 59 ± 3.6 | 9 ± 1.6 |
| SFAs (g) | 48 ± 6.1 | 30 ± 4.2 | 11 ± 2.4 |
| MUFAs (g) | 61 ± 5.2 | 128 ± 12.3 | 13 ± 1.6 |
| PUFAs (g) | 20 ± 2 | 68 ± 4.5 | 10 ± 1.7 |
| n-3 (g) | 3.2 ± 0.2 | 24.4 ± 0.6 | 1.8 ± 0.3 |
| n-6 (g) | 16.1 ± 6 | 47.7 ± 2.7 | 7.5 ± 1.2 |
| n-6/n-3 | 5 ± 1 | 2 ± 1 | 4 ± 1 |
| TEI (kcal) | 3740 ± 53 | 3758 ± 42 | 3752 ± 15 |
| TEI (kJ) | 15,658.63 ± 221 | 15,733.99 ± 175 | 15,708.87 ± 62 |
| Variables | CD | LCD | Carbo-L |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| TPP (s) | 2.65 ± 0.61 | 2.73 ± 0.57 | 2.58 ± 0.39 |
| PP (W/kg) | 20.35 ± 3.44 | 19.94 ± 3.42 | 20.87 ± 0.39 |
| TW/kg (J/kg) | 301.17 ± 12.42 | 266.69 ± 6.46 * | 302.46 ± 8.50 # |
| Variables | CD | LCD | Carbo-L | |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | ||
| LA (mmol/L) | Rest | 1.65 ± 0.06 | 1.26 ± 0.01 * | 1.69 ± 0.04 # |
| Post exercise | 9.47 ± 1.04 & | 8.36 ± 0.62 & | 9.62 ± 0.54 & | |
| pH (−Log[H+]) | Rest | 7.412 ± 0.003 | 7.381 ± 0.001 * | 7.420 ± 0.01 # |
| Post exercise | 7.275 ± 0.005 & | 7.322 ± 0.03 & | 7.261 ± 0.008 & | |
| HCO3− (mmol/L) | Rest | 24.10 ± 0.07 | 23.72 ± 0.11 | 24.48 ± 0.07 |
| Post exercise | 12.80 ± 0.09 & | 13.12 ± 0.14 & | 12.7 ± 0.05 & | |
| β-HB (mmol/L) | Rest | 0.041 ± 0.02 * | 0.161 ± 0.11 | 0.035 ± 0.01 # |
| Variables | CD | LCD | Carbo-L |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| Testosterone (nmol/L) | 546.67 ± 167.16 | 642.37 ± 194.47 * | 643.14 ± 186.52 $ |
| Growth hormone (ng/mL) | 0.15 ± 0.07 | 0.21 ± 0.09 * | 0.11 ± 0.08 # |
| Insulin (IU/mL) | 5.49 ± 3.25 | 3.99 ± 2.61 * | 7.28 ± 3.65 # |
| Cortisol (µg/dL) | 16.38 ± 6.81 | 16.22 ± 6.40 | 16.02 ± 5.79 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalczyk, M.M.; Chycki, J.; Zajac, A.; Maszczyk, A.; Zydek, G.; Langfort, J. Anaerobic Performance after a Low-Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Male Basketball Players. Nutrients 2019, 11, 778. https://doi.org/10.3390/nu11040778
Michalczyk MM, Chycki J, Zajac A, Maszczyk A, Zydek G, Langfort J. Anaerobic Performance after a Low-Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Male Basketball Players. Nutrients. 2019; 11(4):778. https://doi.org/10.3390/nu11040778
Chicago/Turabian StyleMichalczyk, Małgorzata Magdalena, Jakub Chycki, Adam Zajac, Adam Maszczyk, Grzegorz Zydek, and Józef Langfort. 2019. "Anaerobic Performance after a Low-Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Male Basketball Players" Nutrients 11, no. 4: 778. https://doi.org/10.3390/nu11040778
APA StyleMichalczyk, M. M., Chycki, J., Zajac, A., Maszczyk, A., Zydek, G., & Langfort, J. (2019). Anaerobic Performance after a Low-Carbohydrate Diet (LCD) Followed by 7 Days of Carbohydrate Loading in Male Basketball Players. Nutrients, 11(4), 778. https://doi.org/10.3390/nu11040778