Vitamin B12 in Relation to Oxidative Stress: A Systematic Review


Abstract
1. Introduction
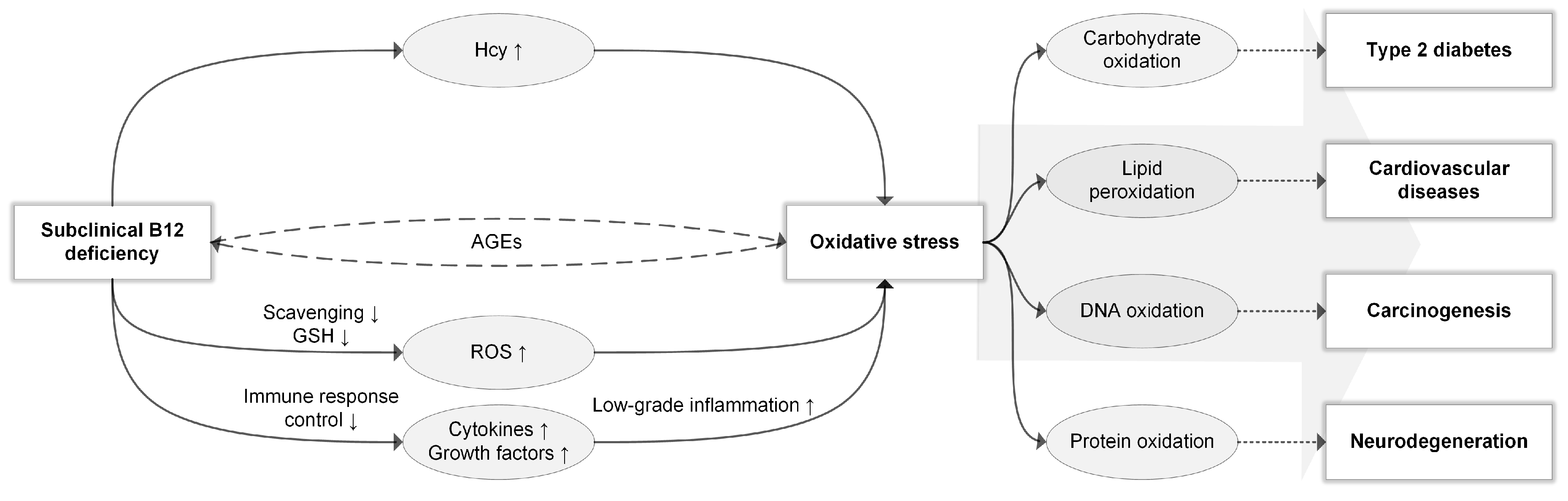
1.1. Subclinical B12 Deficiency
1.2. Oxidative Stress and Biomarkers
1.3. Antioxidant Properties of B12
2. Materials and Methods
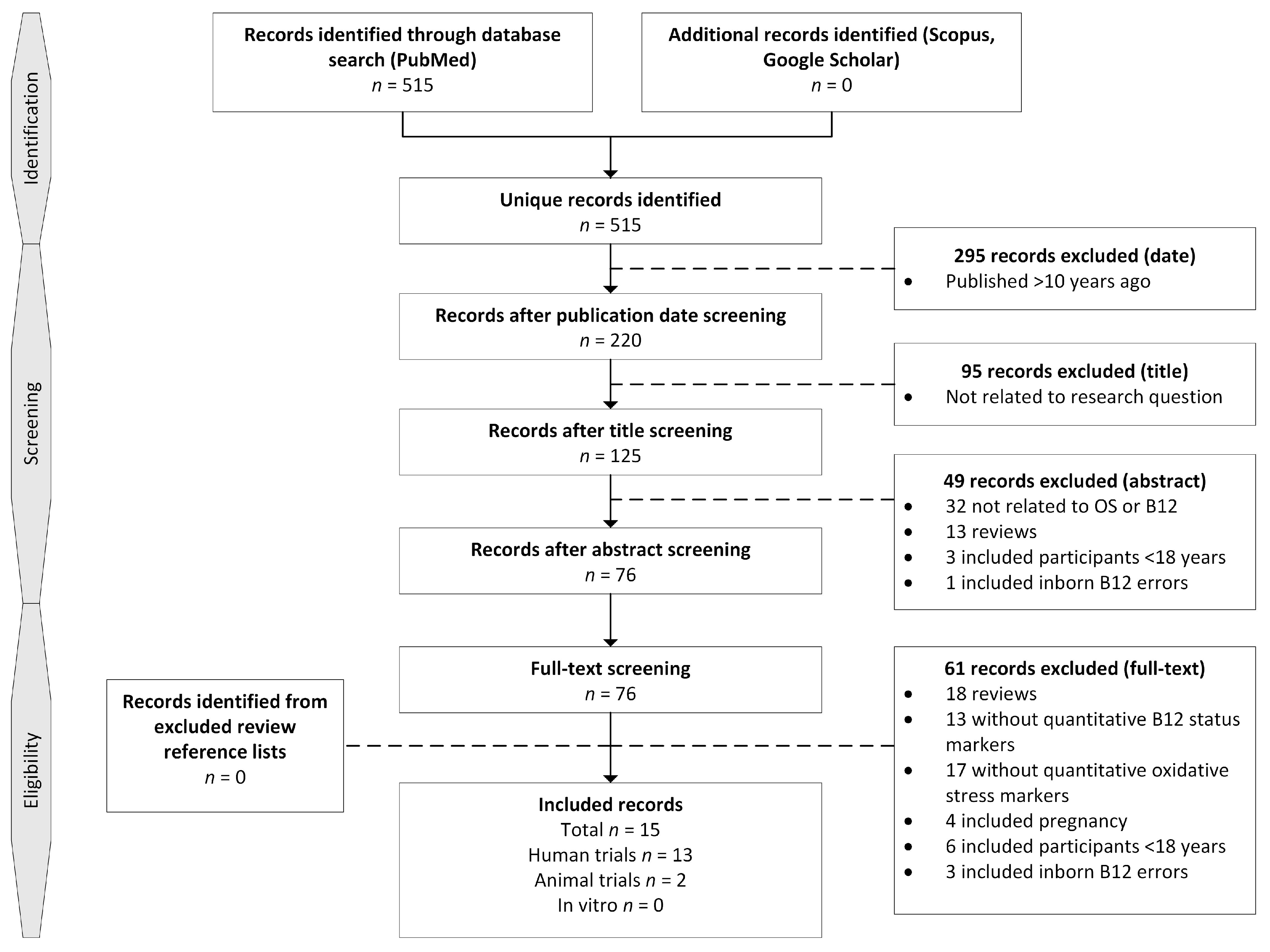
2.1. Identification of Records
2.2. In/Exclusion Criteria
2.3. Screening and Eligibility
2.4. Data Extraction
2.5. Quality Assessment
2.6. Data Analysis
3. Results
3.1. Study Characteristics
3.2. Overall B12 Status in Relation to Oxidative Stress
3.3. Subclinical B12 Deficiency in Relation to Oxidative Stress
4. Discussion
4.1. B12 Status in Relation to Oxidative Stress
4.2. Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Green, R.; Allen, L.H.; Bjørke-Monsen, A.-L.; Brito, A.; Guéant, J.-L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.-H.; et al. Vitamin B12 deficiency. Nat. Rev. Dis. Prim. 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Low micronutrient intake may accelerate the degenerative diseases of aging through allocation of scarce micronutrients by triage. Proc. Natl. Acad. Sci. USA 2006, 103, 17589–17594. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Prevention of mutation, cancer, and other age-associated diseases by optimizing micronutrient intake. J. Nucleic Acids 2010, 2010, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Optimal micronutrients delay mitochondrial decay and age-associated diseases. Mech. Ageing Dev. 2010, 131, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, W.; Hardy, G. Vitamin B12: The forgotten micronutrient for critical care. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, L.; Lysne, V.; Bjørke-Monsen, A.-L.; Behringer, S.; Grünert, S.C.; Spiekerkoetter, U.; Jacobsen, D.W.; Blom, H.J. Biomarkers and algorithms for the diagnosis of vitamin B12 deficiency. Front. Mol. Biosci. 2016, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Carmel, R. Diagnosis and management of clinical and subclinical cobalamin deficiencies: Why controversies persist in the age of sensitive metabolic testing. Biochimie 2013, 95, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, K.; Billa, G. Treatment of vitamin B12 deficiency–Methylcobalamine? Cyancobalamine? Hydroxocobalamin?—Clearing the confusion. Eur. J. Clin. Nutr. 2015, 69, 1–2. [Google Scholar] [CrossRef] [PubMed]
- (NIH), N.I. of H. Dietary Supplement Fact Sheet: Vitamin B12. Available online: https://ods.od.nih.gov/factsheets/VitaminB12-HealthProfessional/#h2 (accessed on 22 November 2017).
- Hine, B.; Boggs, I.; Green, R.; Miller, J.W.; Hovey, R.C.; Humphrey, R.; Wheeler, T.T. Transcobalamin derived from bovine milk stimulates apical uptake of vitamin B12 into human intestinal epithelial cells. J. Cell. Biochem. 2014, 115, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- Bor, M.V.; Lydeking-Olsen, E.; Møller, J.; Nexø, E. A daily intake of approximately 6 μg vitamin B-12 appears to saturate all the vitamin B-12–related variables in Danish postmenopausal women. Am. J. Clin. Nutr. 2006, 83, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.; Grapov, D.; Fahrmann, J.; Harvey, D.; Green, R.; Miller, J.W.; Fedosov, S.N.; Shahab-Ferdows, S.; Hampel, D.; Pedersen, T.L.; et al. The human serum metabolome of vitamin B-12 deficiency and repletion, and associations with neurological function in elderly adults. J. Nutr. 2017, 147, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- McCaddon, A.; Regland, B.; Hudson, P.; Davies, G. Functional vitamin B 12 deficiency and Alzheimer disease. Neurology 2002, 58, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- McCaddon, A. Vitamin B12 in neurology and ageing; clinical and genetic aspects. Biochimie 2013, 95, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Rozycka, A.; Jagodzinski, P.P.; Kozubski, W.; Lianeri, M.; Dorszewska, J. Homocysteine level and mechanisms of injury in Parkinson’s Disease as related to MTHFR, MTR, and MTHFD1 genes polymorphisms and L-Dopa treatment. Curr. Genom. 2013, 14, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Firth, J.; Stubbs, B.; Sarris, J.; Rosenbaum, S.; Teasdale, S.; Berk, M.; Yung, A.R. The effects of vitamin and mineral supplementation on symptoms of schizophrenia: A systematic review and meta-analysis. Psychol. Med. 2017, 47, 1515–1527. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Czerska, M.; Mikołajewska, K.; Zieliński, M.; Gromadzińska, J.; Wąsowicz, W. Today’s oxidative stress markers. Med. Pracy 2015, 66, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Kregel, K.C.; Zhang, H.J. An integrated view of oxidative stress in aging: Basic mechanisms, functional effects, and pathological considerations. AJP Regul. Integr. Comp. Physiol. 2006, 292, R18–R36. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.-J.; Kundu, J.; Na, H.-K. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med. 2008, 74, 1526–1539. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Noori, S. An overview of oxidative stress and antioxidant defensive system. IUBMB Life 2012, 1, 241–244. [Google Scholar] [CrossRef]
- Voigt, A.; Rahnefeld, A.; Kloetzel, P.M.; Krüger, E. Cytokine-induced oxidative stress in cardiac inflammation and heart failure—How the ubiquitin proteasome system targets this vicious cycle. Front. Physiol. 2013, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M.; Nourhashemi, F. Proinflammatory cytokines, aging, and age-related diseases. J. Am. Med. Dir. Assoc. 2013, 14, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Guarner, V.; Rubio-Ruiz, M.E. Low-grade systemic inflammation connects aging, metabolic syndrome and cardiovascular disease. Interdiscip. Top. Gerontol. 2015, 40, 99–106. [Google Scholar] [PubMed]
- Collins, B.; Morrow, J.D.; Milne, G.L.; Swenberg, J.A. Comparison of three oxidative stress biomarkers in a sample of healthy adults. Biomarkers 2009, 14, 587–595. [Google Scholar]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.; Ziaja, I.; Grune, T. Protein oxidation and degradation during aging: Role in skin aging and neurodegeneration. Free Radic. Res. 2006, 40, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.H.S. Suppression of LDL oxidation by garlic compounds is a possible mechanism of cardiovascular health Benefit. J. Nutr. 2006, 136, 765S–768S. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, S.V.; Konkova, M.S.; Ershova, E.S.; Alekseeva, A.J.; Smirnova, T.D.; Stukalov, S.V.; Kozhina, E.A.; Shilova, N.V.; Zolotukhina, T.V.; Markova, Z.G.; et al. An exposure to the oxidized DNA enhances both instability of genome and survival in cancer cells. PLoS ONE 2013, 8, e77469. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.S.; Brasch, N.E.; Yun, J. Vitamin B12 protects against superoxide-induced cell injury in human aortic endothelial cells. Free Radic. Biol. Med. 2011, 51, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.; Almasieh, M.; Catrinescu, M.-M.; Levin, L.A. Cobalamin-associated superoxide scavenging in neuronal cells is a potential mechanism for vitamin B12–deprivation optic neuropathy. Am. J. Pathol. 2017, 188, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Karamshetty, V.; Acharya, J.D.; Ghaskadbi, S.; Goel, P. Mathematical modeling of glutathione status in type 2 diabetics with vitamin B12 deficiency. Front. Cell Dev. Biol. 2016, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Politis, A.; Olgiati, P.; Malitas, P.; Albani, D.; Signorini, A.; Polito, L.; De Mauro, S.; Zisaki, A.; Piperi, C.; Stamouli, E.; et al. Vitamin B12 levels in Alzheimer’s disease: Association with clinical features and cytokine production. J. Alzheimers Dis. 2010, 19, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Birch, C.S.; Brasch, N.E.; McCaddon, A.; Williams, J.H. A novel role for vitamin B(12): Cobalamins are intracellular antioxidants in vitro. Free Radic. Biol. Med. 2009, 47, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J. The oxidant stress of hyperhomocyst(e)inemia. J. Clin. Investig. 1996, 98, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, N. Mechanisms of homocysteine-induced oxidative stress. AJP Heart Circ. Physiol. 2005, 289, H2649–H2656. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Shannan, B.; Herrmann, W. Advanced glycation end products overload might explain intracellular cobalamin deficiency in renal dysfunction, diabetes and aging. Med. Hypotheses 2011, 77, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Gariballa, S.; Afandi, B.; Abuhaltem, M.; Yassin, J.; Habib, H.; Ibrahim, W. Oxidative damage and inflammation in obese diabetic Emirati subjects supplemented with antioxidants and B-vitamins: A randomized placebo-controlled trail. Nutr. Metab. 2013, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Muss, C.; Mosgoeller, W.; Endler, T. Neuroprotective impact of a vitamin trace element composition—A randomized, double blind, placebo controlled clinical trial with healthy volunteers. Neuroendocrinol. Lett. 2015, 36, 31–40. [Google Scholar] [PubMed]
- Hunaiti, A.; Al-Mustafa, A. Correlation between serum B12 levels and lipid peroxidation in B12 deficiency patients. J. Hum. Nutr. Food 2016, 4, 1100. [Google Scholar]
- Solomon, L.R. Low cobalamin levels as predictors of cobalamin deficiency: Importance of comorbidities associated with increased oxidative stress. Am. J. Med. 2016, 129, 115.e9. [Google Scholar] [CrossRef] [PubMed]
- Boanca, M.M.; Colosi, H.A.; Craciun, E.C.; Cra, E.C.; Boanca, M.M.; Colosi, H.A.; Craciun, E.C. The impact of the lacto-ovo vegetarian diet on the erythrocyte superoxide dismutase activity: A study in the Romanian population. Eur. J. Clin. Nutr. 2014, 68, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Solomon, L.R. Functional cobalamin (vitamin B12) deficiency: Role of advanced age and disorders associated with increased oxidative stress. Eur. J. Clin. Nutr. 2015, 69, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Al-Maskari, M.Y.; Waly, M.I.; Ali, A.; Al-Shuaibi, Y.S.; Ouhtit, A. Folate and vitamin B12 deficiency and hyperhomocysteinemia promote oxidative stress in adult type 2 diabetes. Nutrition 2012, 28, e23–e26. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Wang, M.-Y.; Lin, M.-C.; Lin, P.-T. Associations between vitamin B-12 status and oxidative stress and inflammation in diabetic vegetarians and omnivores. Nutrients 2016, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Waly, M.I.; Ali, A.; Al-Nassri, A.; Al-Mukhaini, M.; Valliatte, J.; Al-Farsi, Y. Low nourishment of B-vitamins is associated with hyperhomocysteinemia and oxidative stress in newly diagnosed cardiac patients. Exp. Biol. Med. 2016, 241, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, O.; Ipçioğlu, O.M.; Gültepe, M.; Başoğglu, C. Altered red cell membrane compositions related to functional vitamin B(12) deficiency manifested by elevated urine methylmalonic acid concentrations in patients with schizophrenia. Ann. Clin. Biochem. 2008, 45, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Vener, C.; Novembrino, C.; Catena, F.B.; Fracchiolla, N.S.; Gianelli, U.; Savi, F.; Radaelli, F.; Fermo, E.; Cortelezzi, A.; Lonati, S.; et al. Oxidative stress is increased in primary and post-polycythemia vera myelofibrosis. Exp. Hematol. 2010, 38, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Güney, T.; Alişik, M.; Akinci, S.; Neşelioğlu, S.; Dilek, I.İ.; Erel, Ö.; Guney, T.; Alisik, M.; Akinci, S.; Neselioglu, S.; et al. Evaluation of oxidant and antioxidant status in patients with vitamin B12 deficiency. Turk. J. Med. Sci. 2015, 45, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Misra, U.K.; Kalita, J.; Singh, S.K.; Rahi, S.K. Oxidative stress markers in vitamin B12 deficiency. Mol. Neurobiol. 2017, 54, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sinha, J.K.; Putcha, U.K.; Raghunath, M. Severe but not moderate vitamin B12 deficiency impairs lipid profile, induces adiposity, and leads to adverse gestational outcome in female C57BL/6 mice. Front. Nutr. 2016, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Bito, T.; Misaki, T.; Yabuta, Y.; Ishikawa, T.; Kawano, T.; Watanabe, F. Vitamin B12 deficiency results in severe oxidative stress, leading to memory retention impairment in Caenorhabditis elegans. Redox Biol. 2017, 11, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Information, N.C. for B. Vitamin B-12. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/vitamin B-12#section=Top (accessed on 6 December 2017).
- Hozo, S.P.; Djulbegovic, B.; Hozo, I. Estimating the mean and variance from the median, range, and the size of a sample. BMC Med. Res. Methodol. 2005, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Rzepka, Z.; Respondek, M.; Rok, J.; Beberok, A.; Proinsias, K.Ó.; Gryko, D.; Wrześniok, D. Vitamin B12 deficiency induces imbalance in melanocytes homeostasis—A cellular basis of hypocobalaminemia pigmentary manifestations. Int. J. Mol. Sci. 2018, 19, 2845. [Google Scholar] [CrossRef] [PubMed]
- Danchin, A.; Braham, S. Coenzyme B12 synthesis as a baseline to study metabolite contribution of animal microbiota. Microb. Biotechnol. 2017, 10, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Ugarriza, R.; Palacios, G.; Alder, M.; González-Gross, M. A review of the cut-off points for the diagnosis of vitamin B12 deficiency in the general population. Clin. Chem. Lab. Med. 2015, 53, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, S.N.; Brito, A.; Miller, J.W.; Green, R.; Allen, L.H. Combined indicator of vitamin B12 status: Modification for missing biomarkers and folate status and recommendations for revised cut-points. Clin. Chem. Lab. Med. 2015, 53, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Vet. Res. 2016, 12, 166. [Google Scholar] [CrossRef] [PubMed]
- Pitocco, D.; Zaccardi, F.; Di Stasio, E.; Romitelli, F.; Santini, S.A.; Zuppi, C.; Ghirlanda, G. Oxidative stress, nitric oxide, and diabetes. Rev. Diabet. Stud. 2010, 7, 15–25. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Design | Country | Primary Goal | Study Groups (Mean Age ± SD, % Male) | B12 Markers | Oxidative Stress Markers | Supports Antioxidant Properties of B12 |
|---|---|---|---|---|---|---|---|
| Misra et al. 2017 [59] | CC Human | India | Assess oxidative stress markers in B12-deficient patients compared to healthy volunteers and examine correlation among included markers. | 51 serum B12 deficient patients (45.78 ± 2.19 years, 70.6% male) 53 healthy volunteers (44.28 ± 2.20 years, 77.8% male) | Serum B12 Serum Hcy | Plasma GSH Serum TAC Plasma MDA | Yes |
| Waly et al. 2016 [55] | CC Human | Egypt | Evaluate B-vitamins in relation to hyperhomocysteinemia and oxidative stress in cardiac patients compared to healthy volunteers. | 25 cardiac patients (54.72 ± 10.3 years, 48.0% male) 25 healthy volunteers (53.81 ± 6.8 years, gender not specified) | Serum B12 Serum Hcy | Serum GSH Serum TAC Serum NN Serum MDA | Yes |
| Boanca et al. 2014 [51] | CC Human | Romania | Evaluate the impact of the lacto-ovo diet on oxidative stress compared to non-vegetarians regarding B12 status. | 48 lacto-ovo vegetarians (28.4 ± 8.6 years, 33.3% male) 38 non-vegetarian volunteers (29.8 ± 1 0.1 years, 34.2% male) | Serum B12 | Erythrocyte SOD Serum MDA | Unclear |
| Özcan et al. 2008 [56] | CC Human | Turkey | Investigate the relationship between B12 status and cell membrane composition regarding oxidative stress, cholesterol, and phospholipid content in schizophrenic patients compared to healthy volunteers. | 18 schizophrenic patients (31 ± 7 years, 55.6% male) 20 healthy volunteers (30 ± 8 years, 50.0% male) | Serum B12 Urine MMA Plasma Hcy | Membrane MDA | Yes |
| Vener et al. 2010 [57] | CC Human | Italy | Determine if folate and/or B12 depletion can lead to hyperhomocysteinemia and contribute to oxidative stress in chronic myeloproliferative disorders compared to healthy volunteers. | 51 CMPD patients (median 64 years (range 40–84 years), 52.9% male) 53 healthy volunteers (median 50 years (range 30–84 years), 64.2% male) | Serum B12 Serum holo-TC Serum Hcy | Serum TAC Serum ROS | Yes |
| Al-Maskari et al. 2012 [53] | CC Human | Oman | Evaluate the status of folate and B12 in relation to serum Hcy and oxidative stress indices in T2D patients compared to healthy volunteers. | 50 T2D patients (51.43 ± 7.9 years, 50% male) 50 healthy controls (48.94 ± 6.02 years, 50% male) | Serum B12 Serum Hcy | Serum GSH Serum TAS Serum catalase Serum GPx Serum SOD | Yes |
| Güney et al. 2015 [58] | CC with one-armed intervention Human | Turkey | Determine the effect of B12 deficiency on oxidative stress compared to healthy controls, and to determine the effect of 1-month cyano-Cbl treatment for B12 deficient patients on oxidative stress (without placebo or control). | 40 B12 deficient patients (43.1 ± 1 5.9 years, 25% male) 40 healthy controls (40.1 ± 16.9 years, 42.5% male) | Serum B12 Serum MMA Urine MMA Plasma Hcy | Plasma TOS Plasma TAS OSI | No |
| Lee et al. 2016 [54] | CS Human | Taiwan | Investigate the correlation between vitamin B12 status and oxidative stress and inflammation in diabetic vegetarians and omnivores. | 54 T2D vegetarians (65.1 ± 11.3 years, 30% male) 100 T2D omnivores (57.5 ± 10.5 years, 45% male) | Serum B12 | Plasma MDA Serum Ox-LDL-C Erythrocyte SOD Erythrocyte GPx Erythrocyte catalase | Unclear |
| Hunaiti et al. 2016 [49] | CS Human | Jordan | Assess the impact of B12 deficiency on lipid peroxidation and antioxidant capacity in patients with symptoms and signs of B12 deficiency. | 24 patients with B12 deficiency (mean 53 years, range 36–76 years, 37.5% male) | Serum B12 Serum Hcy | Serum MDA Serum TAC | Yes |
| Gariballa et al. 2013 [47] | RCT Human | UAE | Test the effect of 3 months B-vitamins and antioxidant supplementation compared to placebo on antioxidant capacity and oxidative stress in obese T2D patients. | 50 T2D patients allocated to intervention (median 52 years, Q1–Q3 44–56 years, 46% male) 50 T2D patients allocated to placebo (median 51 years, Q1–Q3 42–60 years, 36% male) | Serum B12 Plasma Hcy | Protein carbonyl Plasma MDA Plasma GSH | Unclear |
| Muss et al. 2015 [48] | RCT Human | Austria | Test the neuroprotective effect of verum supplementation (measured at 3 and 6 months) compared to placebo with a focus on oxidative stress in healthy volunteers. | 116 volunteers allocated to intervention (42.4 ± 15.2 years, 39.7% male) 43 volunteers allocated to placebo (45.8 ± 15.5 years, 25.6% male) | Serum B12 Serum Hcy | Serum FORD Serum FORT Serum lipid peroxidation Serum SOD | Unclear |
| Solomon 2015 [52] | RS Human | USA | Explore the association between functional B12 deficiency and oxidative stress in elderly and younger patients. | 170 community-dwelling adults with serum B12 ≥ 400 pg/mL were retrospectively reviewed and stratified <70 years (n = 100, 42% male) and ≥70 years (n = 70, 54.3% male) | Serum MMA Plasma Hcy | Oxidant risks | Yes |
| Solomon 2016 [50] | RS Human | USA | Examine the relationship between MMA/Hcy and oxidant risks in community-dwelling adults with low (≤200 pg/mL) and low-normal (201-300 pg/mL) serum B12. | 49 participants with low serum B12 (57 ± 19 years, 29% male) 187 participants with low-normal serum B12 (56 ± 17 years, 39% male) | Serum MMA Plasma Hcy | Oxidant risks | Unclear |
| Ghosh et al. 2016 [60] | Animal | Japan | Evaluate differential effects of severe and moderate B12 deficiency on several factors, including oxidative stress. | 10 severely B12 deficient mice 10 moderately deficient mice 10 control mice | Plasma Hcy | Liver MDA Liver protein carbonyl Liver SOD Liver catalase | Yes |
| Bito et al. (2017) [61] | Animal | India | Clarify levels of oxidative stress and induced damage when B12 deficiency is present using a C. elegans animal model. In addition, the relationship between B12 deficiency and memory impairment is investigated. | 12 B12-deficient worms 12 control worms | Plasma Hcy | H2O2 NN MDA Protein carbonyls GSH Total SOD Catalase GPx | Yes |
| Serum B12 A (pmol/L, Mean ± SD) | Antioxidant Marker B,* (% Difference between Groups) | Pro-Oxidant Marker B,* (% Difference between Groups) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Author (Year) | Subclinical Group | Normal Group | Supports B12 as Antioxidant | SOD | GSH | CAT | GPx | TAC TAS | TOS | OSI | NN | MDA |
| Misra (2017) [59] | 172.4 ± 17.8 | 304.0 ± 213.6 | Yes | † | −9.3 * | † | † | −9.0 * | † | † | † | 28.8 * |
| Waly (2016) [55] | 154.4 ± 10.9 | 272.9 ± 23.7 | Yes | † | −111.6 * | † | † | −54.9 * | † | † | 113.6 * | 76.2 * |
| Boanca (2014) [51] | 125.7 ± 44.7 | 282.5 ± 90.2 | Unclear | −6.1 * | † | † | † | † | † | † | † | −16.3 * |
| Al-Maskari (2012) [53] | 186.9 ± 19.0 | 508.0 ± 30.2 | Yes | −70.9 * | −78.3 * | −92.7 * | −66.6 * | −90.9 * | † | † | † | † |
| Güney (2015) [58] | 136.8 ± 40.3 | 562.3 ± 314.8 | No | † | † | † | † | 0.0 | −8.0 | −40.0 | † | † |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van de Lagemaat, E.E.; de Groot, L.C.P.G.M.; van den Heuvel, E.G.H.M. Vitamin B12 in Relation to Oxidative Stress: A Systematic Review. Nutrients 2019, 11, 482. https://doi.org/10.3390/nu11020482
van de Lagemaat EE, de Groot LCPGM, van den Heuvel EGHM. Vitamin B12 in Relation to Oxidative Stress: A Systematic Review. Nutrients. 2019; 11(2):482. https://doi.org/10.3390/nu11020482
Chicago/Turabian Stylevan de Lagemaat, Erik E., Lisette C.P.G.M. de Groot, and Ellen G.H.M. van den Heuvel. 2019. "Vitamin B12 in Relation to Oxidative Stress: A Systematic Review" Nutrients 11, no. 2: 482. https://doi.org/10.3390/nu11020482
APA Stylevan de Lagemaat, E. E., de Groot, L. C. P. G. M., & van den Heuvel, E. G. H. M. (2019). Vitamin B12 in Relation to Oxidative Stress: A Systematic Review. Nutrients, 11(2), 482. https://doi.org/10.3390/nu11020482