Resistant Maltodextrin Ameliorates Altered Hepatic Lipid Homeostasis via Activation of AMP-Activated Protein Kinase in a High-Fat Diet-Fed Rat Model



Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Liver Lipid Extraction
2.3. Detection of Triglyceride (TG), Cholesterol (TC), and Lipoprotein Levels, and Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT) Activity
2.4. Measurement of Hepatic Acetyl-CoA Carboxylase Activity (ACC)
2.5. Measurement of Hepatic Fatty Acid Synthase Activity (FAS)
2.6. Measurement of Hepatic HMG-CoA Reductase (HMGCR) Activity
2.7. Histological Examination
2.8. Protein Expression Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of RMD on the Changes of Body Weight, Liver Weight, Serum and Liver Lipids, and Liver Histopathology in HF-Diet-Fed Rats
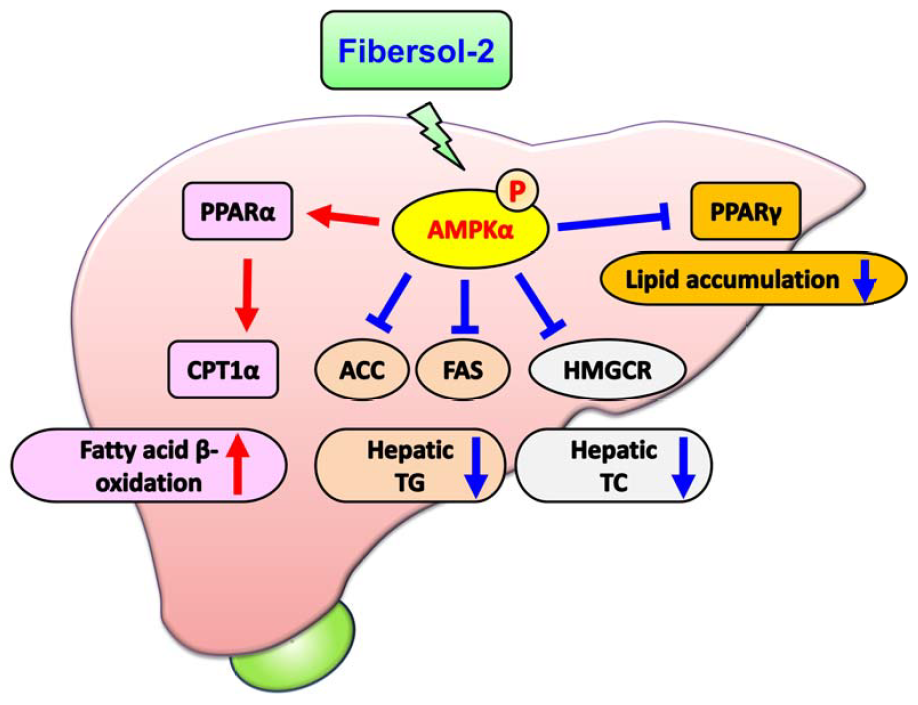
3.2. Effects of RMD on the Activities of Acetyl-CoA Carboxylase (ACC), Fatty Acid Synthase (FAS), and HMG-CoA Reductase (HMGCR) and Lipid Metabolism-Related Signaling Molecules in the Liver of HF-Diet-Fed Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hsu, C.S.; Kao, J.H. Non-alcoholic fatty liver disease: An emerging liver disease in Taiwan. J. Formos. Med. Assoc. 2012, 111, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P. Nonalcoholic fatty liver disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef]
- Rahimlou, M.; Ahmadnia, H.; Hekmatdoost, A. Dietary supplements and pediatric non-alcoholic fatty liver disease: Present and the future. World J. Hepatol. 2015, 7, 2597–2602. [Google Scholar] [CrossRef]
- Abenavoli, L. Non-alcoholic fatty liver disease and beneficial effects of dietary supplements. World J. Hepatol. 2015, 7, 1723–1724. [Google Scholar] [CrossRef] [PubMed]
- Silva Figueiredo, P.; Inada, A.C.; Ribeiro Fernandes, M.; Granja Arakaki, D.; Freitas, K.C.; Avellaneda Guimarães, R.C.; Aragão do Nascimento, V.; Aiko Hiane, P. An overview of novel dietary supplements and food ingredients in patients with metabolic syndrome and non-alcoholic fatty liver disease. Molecules 2018, 23, 877. [Google Scholar] [CrossRef] [PubMed]
- Ohkuma, K.; Wakabayashi, S. Fibersol-2: A soluble, non-digestible, starch-derived dietary fibre. In Advanced Dietary Fibre Technology; McCleary, B.V., Prosky, L., Eds.; Blackwell Science Ltd.: Oxford, UK, 2001; pp. 509–523. [Google Scholar]
- Kishimoto, Y.; Wakabayashi, S.; Tokunaga, K. Effects of long-term administration of indigestible dextrin on visceral fat accumulation. J. Jpn. Assoc. Dietary Fiber Res. 2000, 4, 59–65. [Google Scholar]
- Hashizume, C.; Kishimoto, Y.; Kanahori, S.; Yamamoto, T.; Okuma, K.; Yamamoto, K. Improvement effect of resistant maltodextrin in humans with metabolic syndrome by continuous administration. J. Nutr. Sci. Vitaminol 2012, 58, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar] [PubMed]
- Wang, H.; Wang, L.; Cheng, Y.; Xia, Z.; Liao, Y.; Cao, J. Efficacy of orlistat in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Biomed. Rep. 2018, 9, 90–96. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; Available online: https://doi.org/10.17226/12910 (accessed on 13 September 2018).
- Raederstorff, D.G.; Schlachter, M.F.; Elste, V.; Weber, P. Effect of EGCG on lipid absorption and plasma lipid levels in rats. J. Nutr. Biochem. 2003, 14, 326–332. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Yoshikawa, Y.; Miyazato, S.; Oga, H.; Yamada, T.; Tagami, H.; Hashizume, C.; Yamamoto, K. Effect of resistant maltodextrin on digestion and absorption of lipids. J. Health Sci. 2009, 55, 838–844. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Dyson, J.K.; Anstee, Q.M.; McPherson, S. Non-alcoholic fatty liver disease: A practical approach to diagnosis and staging. Frontline Gastroenterol. 2014, 5, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Yang, R.S.; Sheu, M.L.; Chan, D.C.; Yang, T.H.; Tsai, K.S.; Chiang, C.K.; Liu, S.H. Advanced glycation end-products induce skeletal muscle atrophy and dysfunction in diabetic mice via a RAGE-mediated, AMPK-down-regulated, Akt pathway. J. Pathol. 2016, 238, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Ho, B.Y.; Lee, C.L.; Lee, C.H.; Pan, T.M. Red mold rice prevents the development of obesity, dyslipidemia and hyperinsulinemia induced by high-fat diet. Int. J. Obes. 2008, 32, 1694–1704. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Wang, L.P.; Liu, S.H.; Chiang, M.T. Fish oil supplementation alleviates the altered lipid homeostasis in blood, liver, and adipose tissues in high-fat diet-fed rats. J. Agric. Food Chem. 2018, 66, 4118–4128. [Google Scholar] [CrossRef]
- Borovicka, J.; Schwizer, W.; Guttmann, G.; Hartmann, D.; Kosinski, M.; Wastiel, C.; Bischof-Delaloye, A.; Fried, M. Role of lipase in the regulation of postprandial gastric acid secretion and emptying of fat in humans: A study with orlistat, a highly specific lipase inhibitor. Gut 2000, 46, 774–781. [Google Scholar] [CrossRef]
- Mahmoud, R.H.; Elnour, W.A. Comparative evaluation of the efficacy of ginger and orlistat on obesity management, pancreatic lipase and liver peroxisomal catalase enzyme in male albino rats. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 75–83. [Google Scholar]
- Guo, J.; Jou, W.; Gavrilova, O.; Hall, K.D. Persistent diet-induced obesity in male C57BL/6 mice resulting from temporary obesigenic diets. PLoS ONE 2009, 4, e5370. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chang, T.C.; Liu, S.H.; Chiang, M.T. The regulatory effects of fish oil and chitosan on hepatic lipogenic signals in high-fat diet-induced obese rats. J. Food Drug Anal. 2017, 25, 919–930. [Google Scholar] [CrossRef]
- Biddinger, S.B.; Almind, K.; Miyazaki, M.; Kokkotou, E.; Ntambi, J.M.; Kahn, C.R. Effects of diet and genetic background on sterol regulatory element-binding protein-1c, stearoyl-CoA desaturase 1, and the development of the metabolic syndrome. Diabetes 2005, 54, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Meugnier, E.; Bossu, C.; Oliel, M.; Jeanne, S.; Michaut, A.; Sothier, M.; Brozek, J.; Rome, S.; Laville, M.; Vidal, H. Changes in gene expression in skeletal muscle in response to fat overfeeding in lean men. Obesity 2007, 15, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, S.; Satouchi, M.; Nogami, Y.; Ohkuma, K.; Matsuoka, A. Effect of resistant maltodextrin on cholesterol metabolism in rat. J. Jpn. Soc. Nutr. Food Sci. 1991, 44, 471–478. [Google Scholar] [CrossRef]
- Sanders, F.W.; Griffin, J.L. De novo lipogenesis in the liver in health and disease: More than just a shunting yard for glucose. Biol. Rev. Camb. Philos. Soc. 2016, 91, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Kapoor, A.; Fuchs, M.; Mirshahi, F.; Zhou, H.; Maher, J.; Kellum, J.; Warnick, R.; Contos, M.J.; Sanyal, A.J. Increased hepatic synthesis and dysregulation of cholesterol metabolism is associated with the severity of nonalcoholic fatty liver disease. Cell Metab. 2012, 15, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Ruderman, N.B. Malonyl-CoA and AMP-activated protein kinase: An expanding partnership. Mol. Cell Biochem. 2003, 253, 65–70. [Google Scholar] [CrossRef]
- Dagon, Y.; Avraham, Y.; Berry, E.M. AMPK activation regulates apoptosis, adipogenesis, and lipolysis by eIF2alpha in adipocytes. Biochem. Biophys. Res. Commun. 2006, 340, 43–47. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chan, I.L.; Yang, T.H.; Liu, S.H.; Chiang, M.T. Supplementation of chitosan alleviates high-fat diet-enhanced lipogenesis in rats via adenosine monophosphate (AMP)-activated protein kinase activation and inhibition of lipogenesis-associated genes. J. Agric. Food Chem. 2015, 63, 2979–2988. [Google Scholar] [CrossRef]
- Lee, W.J.; Kim, M.; Park, H.S.; Kim, H.S.; Jeon, M.J.; Oh, K.S.; Koh, E.H.; Won, J.C.; Kim, M.S.; Oh, G.T.; et al. AMPK activation increases fatty acid oxidation in skeletal muscle by activating PPARα and PGC-1. Biochem. Biophys. Res. Commun. 2006, 340, 291–295. [Google Scholar] [CrossRef]
- Minnich, A.; Tian, N.; Byan, L.; Bilder, G. A potent PPARα agonist stimulates mitochondrial fatty acid β-oxidation in liver and skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E270–E279. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lee, H.E.; Lee, J.Y. A pharmacological inhibitor of NLRP3 inflammasome prevents non-alcoholic fatty liver disease in a mouse model induced by high fat diet. Sci. Rep. 2016, 6, 24399. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | NC | HF | HO | FS |
|---|---|---|---|---|
| Corn starch | 64.8 | 49.1 | 49.1 | 49.1 |
| Casein | 20 | 20 | 20 | 20 |
| Lard | 3 | 18 | 18 | 18 |
| Soybean oil | 2 | 2 | 2 | 2 |
| Vitamin 1 | 1 | 1 | 1 | 1 |
| Mineral 2 | 4 | 4 | 4 | 4 |
| Cholesterol | - | 0.5 | 0.5 | 0.5 |
| Cholic acid | - | 0.2 | 0.2 | 0.2 |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 |
| Cellulose | 5 | 5 | 5 | - |
| Fibersol-2 | - | - | - | 5 |
| Orlistat | - | - | 0.2 | - |
| Total calories (kcal/100 g) | 394.2 | 466.4 | 466.4 | 466.4 |
| Carbohydrate (% kcal) | 68.29 | 44.25 | 44.25 | 44.25 |
| Protein (% kcal) | 20.29 | 17.15 | 17.15 | 17.15 |
| Fat (% kcal) | 11.42 | 38.6 | 38.6 | 38.6 |
| Parameters | NC | HF | HO | FS |
|---|---|---|---|---|
| Body weight (g) | 616.2 ± 22.5 | 638.0 ± 33.6 * | 552.5 ± 35.6 **,## | 616.5 ± 43.8 |
| Body weight gain (g) | 115.5 ± 14.7 | 107.2 ± 16.6 | 34.3 ± 15.2 **,## | 106.4 ± 22.5 |
| Food intake (g/day) | 31.2 ± 5.2 | 21.5 ± 0.8 ** | 25.5 ± 2.0 *,## | 20.9 ± 1.5 * |
| Feed efficiency (%) 1 | 3.9 ± 0.7 | 4.9 ± 0.7 * | 1.5 ± 0.7 **,## | 5.1 ± 0.7 * |
| Water intake (mL/day) | 30.0 ± 6.5 | 28.0 ± 12.6 | 40.9 ± 6.9 **,# | 28.3 ± 11.5 |
| Parameters | NC | HF | HO | FS |
|---|---|---|---|---|
| Liver weight (g) | 17.0 ± 1.4 | 36.2 ± 2.9 ** | 28.3 ± 4.5 **,## | 33.5 ± 3.4 ** |
| Relative liver weight (g/100 g BW) | 2.7 ± 0.2 | 5.8 ± 0.5 ** | 5.0 ± 0.4 **,## | 5.3 ± 0.4 ** |
| Adipose tissue weight (g) | 32.0 ± 4.7 | 28.5 ± 7.5 | 19.7 ± 7.4 **# | 29.5 ± 6.2 |
| Relative adipose tissue weight (g/100 g BW) | 5.2 ± 0.7 | 4.5 ± 1.0 | 3.4 ± 1.0 *# | 4.7 ± 0.8 |
| Parameters | NC | HF | HO | FS |
|---|---|---|---|---|
| Triglyceride (mg/dL) | 137.9 ± 44.6 | 23.1 ± 7.5 ** | 60.9 ± 21.8 **,## | 27.9 ± 9.0 ** |
| Total cholesterol (mg/dL) | 90.1 ± 9.3 | 115.6 ± 16.2 * | 117.3 ± 10.9 ** | 77.6 ± 14.7 # |
| HDL-C (mg/dL) | 54.2 ± 9.6 | 42.2 ± 10.0 * | 28.8 ± 8.3 **,# | 31.7 ± 5.4 **,# |
| LDL-C (mg/dL) | 22.7 ± 6.6 | 55.0 ± 10.7 ** | 67.8 ± 13.0 ** | 32.1 ± 13.5 ## |
| HDL-C/LDL-C ratio | 2.6 ± 0.9 | 0.8 ± 0.3 ** | 0.4 ± 0.2 **,# | 1.2 ± 0.5 ** |
| AST (U/L) | 15.6 ± 2.0 | 82.2 ± 36.0 ** | 47.4 ± 21.2 **,# | 50.8 ± 18.0 **,# |
| ALT (U/L) | 8.6 ± 3.5 | 37.4 ± 5.9 * | 29.3 ± 13.9 **,# | 25.3 ± 8.9 **,# |
| Parameters | NC | HF | HO | FS |
|---|---|---|---|---|
| Total cholesterol (mg/g liver) | 3.8 ± 2.3 | 138.4 ± 25.5 ** | 101.4 ± 16.5 **,# | 119.7 ± 17.0 **,# |
| Triglyceride (mg/g liver) | 14.0 ± 3.9 | 75.2 ± 10.2 ** | 41.6 ± 10.7 **,## | 57.8 ± 15.5 **,# |
| Parameters | NC | HF | HO | FS |
|---|---|---|---|---|
| Fecal wet weight (g/day) | 2.0 ± 0.2 | 1.9 ± 0.2 | 5.7 ± 0.7 **,## | 1.4 ± 0.2 **,## |
| Fecal dry weight (g/day) | 1.9 ± 0.2 | 1.7 ± 0.1 * | 4.0 ± 0.4 **,## | 1.0 ± 0.1 **,## |
| Triglyceride (mg/g feces) | 2.5 ± 1.4 | 3.1 ± 0.7 * | 138.1 ± 46.8 **,## | 4.7 ± 0.9 **,## |
| Total cholesterol (mg/g feces) | 2.4 ± 0.6 a | 12.8 ± 3.1 ** | 13.9 ± 2.1 ** | 24.6 ± 4.1 **,## |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-H.; Chiu, C.-Y.; Huang, L.-H.; Chiang, M.-T. Resistant Maltodextrin Ameliorates Altered Hepatic Lipid Homeostasis via Activation of AMP-Activated Protein Kinase in a High-Fat Diet-Fed Rat Model. Nutrients 2019, 11, 291. https://doi.org/10.3390/nu11020291
Liu S-H, Chiu C-Y, Huang L-H, Chiang M-T. Resistant Maltodextrin Ameliorates Altered Hepatic Lipid Homeostasis via Activation of AMP-Activated Protein Kinase in a High-Fat Diet-Fed Rat Model. Nutrients. 2019; 11(2):291. https://doi.org/10.3390/nu11020291
Chicago/Turabian StyleLiu, Shing-Hwa, Chen-Yuan Chiu, Lin-Hui Huang, and Meng-Tsan Chiang. 2019. "Resistant Maltodextrin Ameliorates Altered Hepatic Lipid Homeostasis via Activation of AMP-Activated Protein Kinase in a High-Fat Diet-Fed Rat Model" Nutrients 11, no. 2: 291. https://doi.org/10.3390/nu11020291
APA StyleLiu, S.-H., Chiu, C.-Y., Huang, L.-H., & Chiang, M.-T. (2019). Resistant Maltodextrin Ameliorates Altered Hepatic Lipid Homeostasis via Activation of AMP-Activated Protein Kinase in a High-Fat Diet-Fed Rat Model. Nutrients, 11(2), 291. https://doi.org/10.3390/nu11020291