Neuroprotective Effects of Choline and Other Methyl Donors

Abstract
1. Introduction
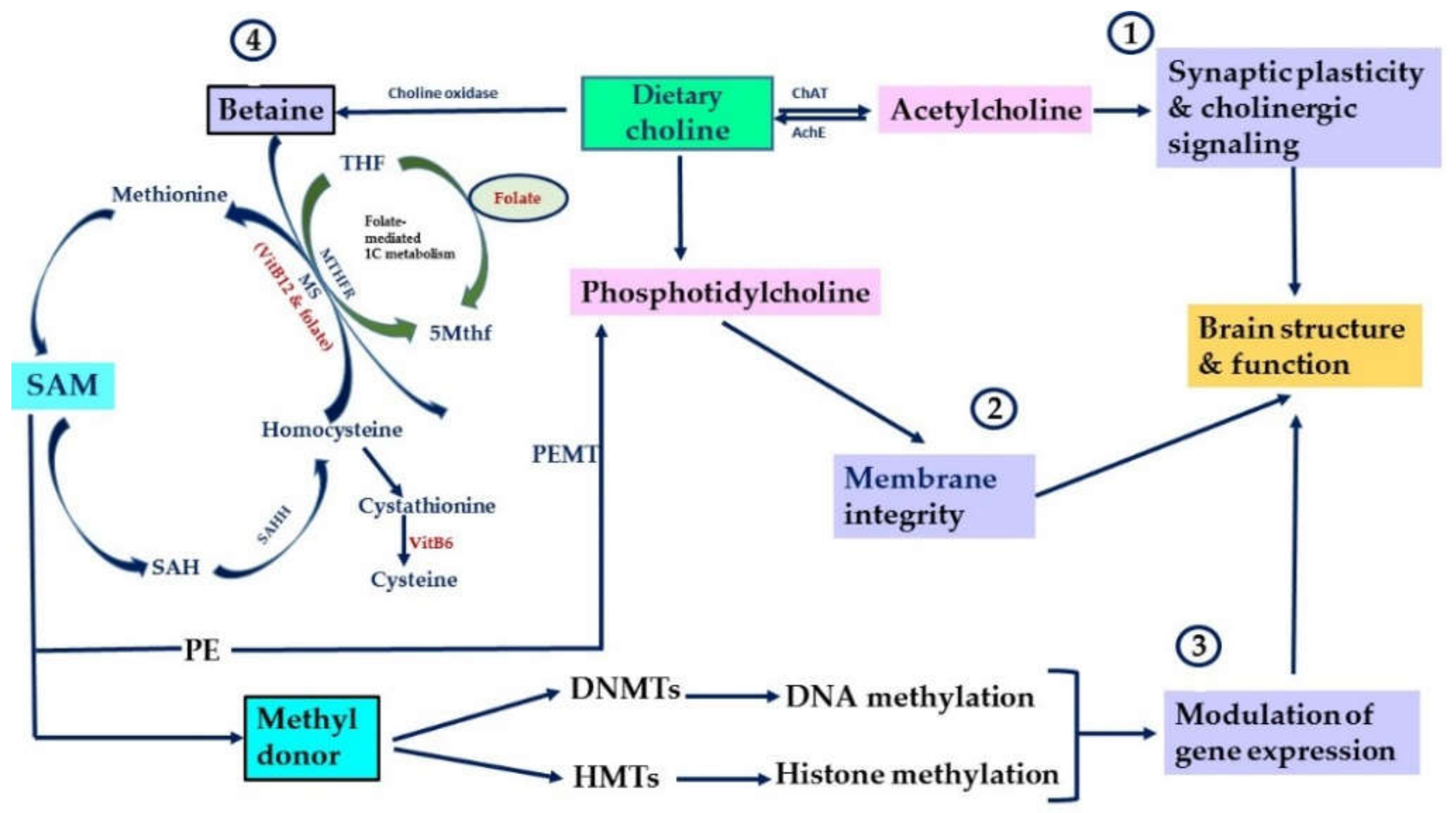
2. The Physiological Functions of Choline and Other Methyl Donors
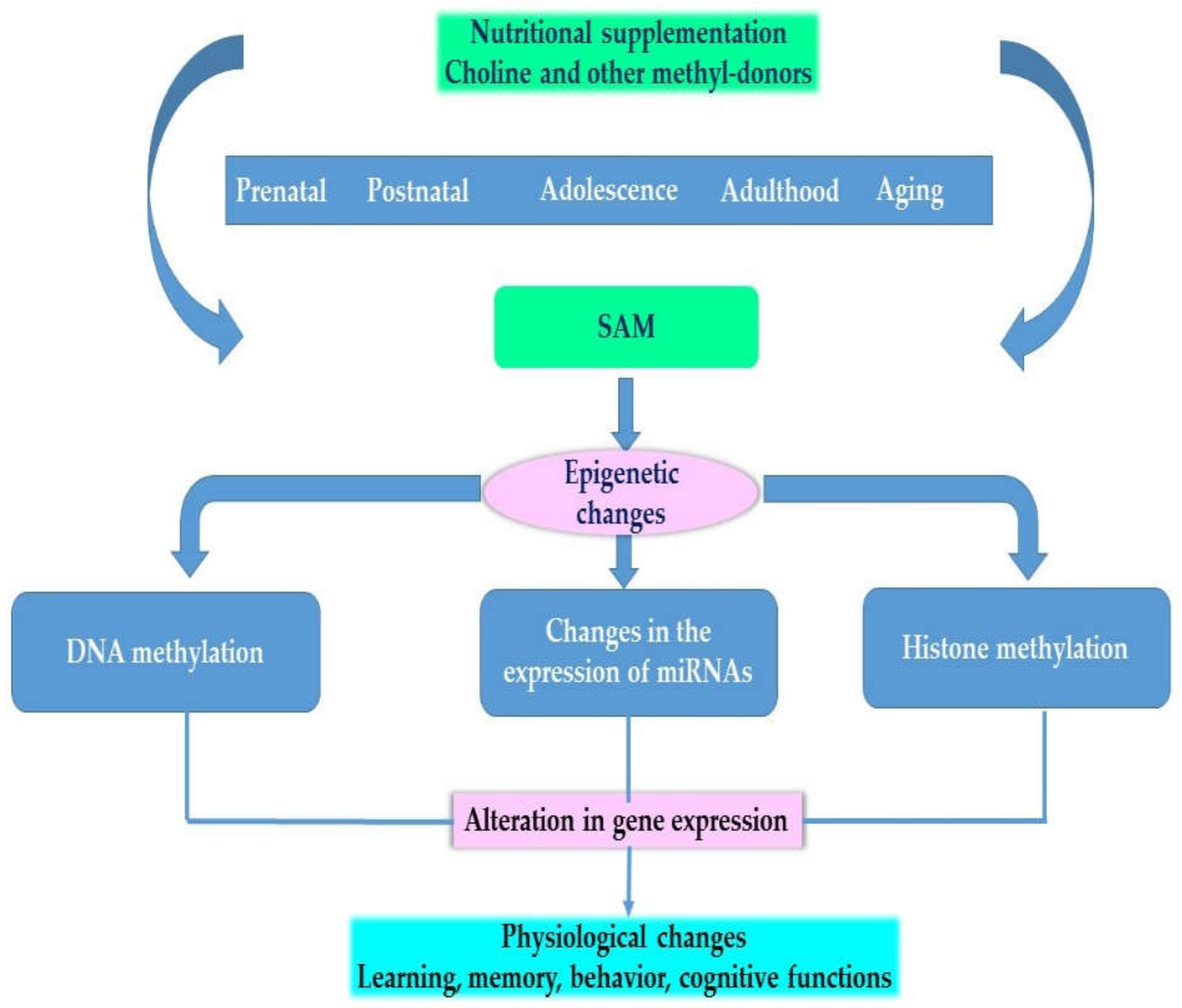
3. Choline and Other Methyl Donors as Modulators of Neuronal Plasticity Throughout Development
3.1. Choline, Other Methyl Donors and the Developing Brain
3.2. Choline, Other Methyl Donors and the Aging Brain
4. Adverse Effects of Inadequate Intake of Methyl Donors
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abdul, Q.A.; Yu, B.P.; Chung, H.Y.; Jung, H.A.; Choi, J.S. Epigenetic Modifications of Gene Expression by Lifestyle and Environment. Arch. Pharm. Res. 2017, 40, 1219–1237. [Google Scholar] [CrossRef]
- Mill, J.; Tang, T.; Kaminsky, Z.; Khare, T.; Yazdanpanah, S.; Bouchard, L.; Jia, P.; Assadzadeh, A.; Flanagan, J.; Schumacher, A.; et al. Epigenomic Profiling Reveals DNA-Methylation Changes Associated with Major Psychosis. Am. J. Hum. Genet. 2008, 82, 696–711. [Google Scholar] [CrossRef]
- Klengel, T.; Pape, J.; Binder, E.B.; Mehta, D. The Role of DNA Methylation in Stress-Related Psychiatric Disorders. Neuropharmacology 2014, 80, 115–132. [Google Scholar] [CrossRef]
- Prasad, R.; Jho, E.-H. A Concise Review of Human Brain Methylome during Aging and Neurodegenerative Diseases. BMB Rep. 2019, 52, 577–588. [Google Scholar] [CrossRef]
- Berdasco, M.; Esteller, M. Clinical Epigenetics: Seizing Opportunities for Translation. Nat. Rev. Genet. 2019, 20, 109–127. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Klatt, K.C.; Caudill, M.A. Choline. Adv. Nutr. 2018, 9, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Radziejewska, A.; Chmurzynska, A. Folate and Choline Absorption and Uptake: Their Role in Fetal Development. Biochimie 2019, 158, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S. Choline, other Methyl-Donors and Epigenetics. Nutrients 2017, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Bekdash, R.A. Choline, the Brain and Neurodegeneration: Insights from Epigenetics. Front. Biosci. 2018, 23, 1113–1143. [Google Scholar] [CrossRef] [PubMed]
- Cooney, C.A. Are Somatic Cells Inherently Deficient in Methylation Metabolism? A Proposed Mechanism for DNA Methylation Loss, Senescence and Aging. Growth Dev. Aging 1993, 57, 261–273. [Google Scholar] [PubMed]
- Montgomery, S.E.; Sepehry, A.A.; Wangsgaard, J.D.; Koenig, J.E. The Effect of S-Adenosylmethionine on Cognitive Performance in Mice: An Animal Model Meta-Analysis. PLoS ONE 2014, 9, e107756. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C. A Comprehensive Review of Eggs, Choline, and Lutein on Cognition Across the Life-Span. J. Am. Coll. Nutr. 2018, 37, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; Costa, K.A.D.; Franklin, P.D.; Alexander, E.A.; Lamont, J.T.; Sheard, N.F.; Beiser, A. Choline, an Essential Nutrient for Humans. FASEB J. 1991, 5, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Choline: Essential for Brain Development and Function. Adv. Pediatr. 1997, 44, 263–295. [Google Scholar] [PubMed]
- Zeisel, S.H.; Niculescu, M.D. Perinatal Choline Influences Brain Structure and Function. Nutr. Rev. 2006, 64, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Choline: Needed for Normal Development of Memory. J. Am. Coll. Nutr. 2000, 19 (Suppl. 5), 528S–531S. [Google Scholar] [CrossRef]
- Thomas, J.D.; Biane, J.S.; O’Bryan, K.A.; O’Neill, T.M.; Dominguez, H.D. Choline Supplementation Following Third-Trimester-Equivalent Alcohol Exposure Attenuates Behavioral Alterations in Rats. Behav. Neurosci. 2007, 121, 120–130. [Google Scholar] [CrossRef]
- Bekdash, R.A.; Zhang, C.; Sarkar, D.K. Gestational Choline Supplementation Normalized Fetal Alcohol-Induced Alterations in Histone Modifications, DNA Methylation, and Proopiomelanocortin (POMC) Gene Expression in β-Endorphin-Producing POMC Neurons of the Hypothalamus. Alcohol. Clin. Exp. Res. 2013, 37, 1133–1142. [Google Scholar] [CrossRef]
- Kennedy, B.C.; Tran, P.V.; Kohli, M.; Maertens, J.J.; Gewirtz, J.C.; Georgieff, M.K. Beneficial Effects of Postnatal Choline Supplementation on Long-Term Neurocognitive Deficit Resulting from Fetal-Neonatal Iron Deficiency. Behav. Brain Res. 2018, 336, 40–43. [Google Scholar] [CrossRef]
- Moreno, H.C.; de Brugada, I.; Carias, D.; Gallo, M. Long-Lasting Effects of Prenatal Dietary Choline Availability on Object Recognition Memory Ability in Adult Rats. Nutr. Neurosci. 2013, 16, 269–274. [Google Scholar] [CrossRef]
- Dauncey, M.J. Nutrition, the Brain and Cognitive Decline: Insights from Epigenetics. Eur. J. Clin. Nutr. 2014, 68, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Dauncey, M.J. Recent Advances in Nutrition, Genes and Brain Health. Proc. Nutr. Soc. 2012, 71, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Powers, B.E.; Kelley, C.M.; Velazquez, R.; Ash, J.A.; Strawderman, M.S.; Alldred, M.J.; Ginsberg, S.D.; Mufson, E.J.; Strupp, B.J. Maternal Choline Supplementation in a Mouse Model of Down Syndrome: Effects on Attention and Nucleus Basalis/Substantia Innominata Neuron Morphology in Adult Offspring. Neuroscience 2017, 340, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Strupp, B.J.; Powers, B.E.; Velazquez, R.; Ash, J.A.; Kelley, C.M.; Alldred, M.J.; Strawderman, M.; Caudill, M.A.; Mufson, E.J.; Ginsberg, S.D. Maternal Choline Supplementation: A Potential Prenatal Treatment for Down Syndrome and Alzheimer’s Disease. Curr. Alzheimer Res. 2016, 13, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Alldred, M.J.; Chao, H.M.; Lee, S.H.; Beilin, J.; Powers, B.E.; Petkova, E.; Strupp, B.J.; Ginsberg, S.D. CA1 Pyramidal Neuron Gene Expression Mosaics in the Ts65Dn Murine Model of Down Syndrome and Alzheimer’s Disease Following Maternal Choline Supplementation. Hippocampus 2018, 28, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Chin, E.W.M.; Lim, W.M.; Ma, D.; Rosales, F.J.; Goh, E.L.K. Choline Rescues Behavioural Deficits in a Mouse Model of Rett Syndrome by Modulating Neuronal Plasticity. Mol. Neurobiol. 2019, 56, 3882–3896. [Google Scholar] [CrossRef]
- Athanasopoulos, D.; Karagiannis, G.; Tsolaki, M. Recent Findings in Alzheimer Disease and Nutrition Focusing on Epigenetics. Adv. Nutr. 2016, 7, 917–927. [Google Scholar] [CrossRef]
- Blusztajn, J.K.; Mellott, T.J. Choline Nutrition Programs Brain Development via DNA and Histone Methylation. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 82–94. [Google Scholar] [CrossRef]
- Blusztajn, J.K.; Slack, B.E.; Mellott, T.J. Neuroprotective Actions of Dietary Choline. Nutrients 2017, 9, 815. [Google Scholar] [CrossRef]
- Chango, A.; Pogribny, I.P. Considering Maternal Dietary Modulators for Epigenetic Regulation and Programming of the Fetal Epigenome. Nutrients 2015, 7, 2748–2770. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and its Panel on Folate, Other B Vitamins, and Choline. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; The National Academies Collection: Reports funded by National Institutes of Health; National Academies Press (US): Washington, DC, USA, 1998. [Google Scholar]
- Wallace, T.C.; Blusztajn, J.K.; Caudill, M.A.; Klatt, K.C.; Natker, E.; Zeisel, S.H.; Zelman, K.M. Choline: The Underconsumed and Underappreciated Essential Nutrient. Nutr. Today 2018, 53, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.M.; daCosta, K.A.; Kwock, L.; Stewart, P.W.; Lu, T.-S.; Stabler, S.P.; Allen, R.H.; Zeisel, S.H. Sex and Menopausal Status Influence Human Dietary Requirements for the Nutrient Choline. Am. J. Clin. Nutr. 2007, 85, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [PubMed]
- Ganz, A.B.; Klatt, K.C.; Caudill, M.A. Common Genetic Variants Alter Metabolism and Influence Dietary Choline Requirements. Nutrients 2017, 9, 837. [Google Scholar] [CrossRef]
- Resseguie, M.; Song, J.; Niculescu, M.D.; da Costa, K.-A.; Randall, T.A.; Zeisel, S.H. Phosphatidylethanolamine N-Methyltransferase (PEMT) Gene Expression Is Induced by Estrogen in Human and Mouse Primary Hepatocytes. FASEB J. 2007, 21, 2622–2632. [Google Scholar] [CrossRef]
- Iorio, E.; Caramujo, M.J.; Cecchetti, S.; Spadaro, F.; Carpinelli, G.; Canese, R.; Podo, F. Key Players in Choline Metabolic Reprograming in Triple-Negative Breast Cancer. Front. Oncol. 2016, 6, 205. [Google Scholar] [CrossRef]
- Wortmann, S.B.; Mayr, J.A. Choline-Related-Inherited Metabolic Diseases-A Mini Review. J. Inherit. Metab. Dis. 2019, 42, 237–242. [Google Scholar] [CrossRef]
- Mehedint, M.G.; Zeisel, S.H. Choline’s Role in Maintaining Liver Function: New Evidence for Epigenetic Mechanisms. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 339–345. [Google Scholar] [CrossRef]
- Zeisel, S.H. Dietary Choline Deficiency Causes DNA Strand Breaks and Alters Epigenetic Marks on DNA and Histones. Mutat. Res. 2012, 733, 34–38. [Google Scholar] [CrossRef]
- Shaw, G.M.; Carmichael, S.L.; Yang, W.; Selvin, S.; Schaffer, D.M. Periconceptional Dietary Intake of Choline and Betaine and Neural Tube Defects in Offspring. Am. J. Epidemiol. 2004, 160, 102–109. [Google Scholar] [CrossRef]
- Li, K.; Wahlqvist, M.L.; Li, D. Nutrition, One-Carbon Metabolism and Neural Tube Defects: A Review. Nutrients 2016, 8, 741. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeier, M.; da Costa, K.-A.; Fischer, L.M.; Zeisel, S.H. Genetic Variation of Folate-Mediated One-Carbon Transfer Pathway Predicts Susceptibility to Choline Deficiency in Humans. Proc. Natl. Acad. Sci. USA 2005, 102, 16025–16030. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bar, H.Y.; Yan, J.; Jones, S.; Brannon, P.M.; West, A.A.; Perry, C.A.; Ganti, A.; Pressman, E.; Devapatla, S.; et al. A Higher Maternal Choline Intake among Third-Trimester Pregnant Women Lowers Placental and Circulating Concentrations of the Antiangiogenic Factor Fms-like Tyrosine Kinase-1 (SFLT1). FASEB J. 2013, 27, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yan, J.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Devapatla, S.; Pressman, E.; Vermeylen, F.; Caudill, M.A. Maternal Choline Intake Alters the Epigenetic State of Fetal Cortisol-Regulating Genes in Humans. FASEB J. 2012, 26, 3563–3574. [Google Scholar] [CrossRef] [PubMed]
- Meck, W.H.; Williams, C.L. Metabolic Imprinting of Choline by Its Availability during Gestation: Implications for Memory and Attentional Processing across the Lifespan. Neurosci. Biobehav. Rev. 2003, 27, 385–399. [Google Scholar] [CrossRef]
- Meck, W.H.; Williams, C.L.; Cermak, J.M.; Blusztajn, J.K. Developmental Periods of Choline Sensitivity Provide an Ontogenetic Mechanism for Regulating Memory Capacity and Age-Related Dementia. Front. Integr. Neurosci. 2007, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Dorninger, F.; Moser, A.B.; Kou, J.; Wiesinger, C.; Forss-Petter, S.; Gleiss, A.; Hinterberger, M.; Jungwirth, S.; Fischer, P.; Berger, J. Alterations in the Plasma Levels of Specific Choline Phospholipids in Alzheimer’s Disease Mimic Accelerated Aging. J. Alzheimers Dis. 2018, 62, 841–854. [Google Scholar] [CrossRef]
- Zhu, H.; Yan, H.; Tang, N.; Li, X.; Pang, P.; Li, H.; Chen, W.; Guo, Y.; Shu, S.; Cai, Y.; et al. Impairments of Spatial Memory in an Alzheimer’s Disease Model via Degeneration of Hippocampal Cholinergic Synapses. Nat. Commun. 2017, 8, 1676. [Google Scholar] [CrossRef]
- Thomas, J.D.; Idrus, N.M.; Monk, B.R.; Dominguez, H.D. Prenatal Choline Supplementation Mitigates Behavioral Alterations Associated with Prenatal Alcohol Exposure in Rats. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 827–837. [Google Scholar] [CrossRef]
- Idrus, N.M.; Breit, K.R.; Thomas, J.D. Dietary Choline Levels Modify the Effects of Prenatal Alcohol Exposure in Rats. Neurotoxicol. Teratol. 2017, 59, 43–52. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Risbud, R.D.; Mattson, S.N.; Chambers, C.D.; Thomas, J.D. Randomized, Double-Blind, Placebo-Controlled Clinical Trial of Choline Supplementation in School-Aged Children with Fetal Alcohol Spectrum Disorders. Am. J. Clin. Nutr. 2016, 104, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.C.; Kolodny, N.H.; Nag, N.; Berger-Sweeney, J.E. Neurochemical Changes in a Mouse Model of Rett Syndrome: Changes over Time and in Response to Perinatal Choline Nutritional Supplementation. J. Neurochem. 2009, 108, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Powers, B.E.; Velazquez, R.; Kelley, C.M.; Ash, J.A.; Strawderman, M.S.; Alldred, M.J.; Ginsberg, S.D.; Mufson, E.J.; Strupp, B.J. Attentional Function and Basal Forebrain Cholinergic Neuron Morphology during Aging in the Ts65Dn Mouse Model of Down Syndrome. Brain Struct. Funct. 2016, 221, 4337–4352. [Google Scholar] [CrossRef] [PubMed]
- Ash, J.A.; Velazquez, R.; Kelley, C.M.; Powers, B.E.; Ginsberg, S.D.; Mufson, E.J.; Strupp, B.J. Maternal Choline Supplementation Improves Spatial Mapping and Increases Basal Forebrain Cholinergic Neuron Number and Size in Aged Ts65Dn Mice. Neurobiol. Dis. 2014, 70, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.E.; Ridgway, N.D. The Methylation of Phosphatidylethanolamine. Prog. Lipid Res. 1988, 27, 61–79. [Google Scholar] [CrossRef]
- Millington, W.R.; Wurtman, R.J. Choline Administration Elevates Brain Phosphorylcholine Concentrations. J. Neurochem. 1982, 38, 1748–1752. [Google Scholar] [CrossRef]
- Babb, S.M.; Ke, Y.; Lange, N.; Kaufman, M.J.; Renshaw, P.F.; Cohen, B.M. Oral Choline Increases Choline Metabolites in Human Brain. Psychiatry Res. 2004, 130, 1–9. [Google Scholar] [CrossRef]
- Michel, V.; Yuan, Z.; Ramsubir, S.; Bakovic, M. Choline Transport for Phospholipid Synthesis. Exp. Biol. Med. 2006, 231, 490–504. [Google Scholar] [CrossRef]
- Horn, A.K.E.; Horng, A.; Buresch, N.; Messoudi, A.; Härtig, W. Identification of Functional Cell Groups in the Abducens Nucleus of Monkey and Human by Perineuronal Nets and Choline Acetyltransferase Immunolabeling. Front. Neuroanat. 2018, 12, 45. [Google Scholar] [CrossRef]
- Blusztajn, J.K.; Wurtman, R.J. Choline and Cholinergic Neurons. Science 1983, 221, 614–620. [Google Scholar] [CrossRef]
- Mellott, T.J.; Huleatt, O.M.; Shade, B.N.; Pender, S.M.; Liu, Y.B.; Slack, B.E.; Blusztajn, J.K. Perinatal Choline Supplementation Reduces Amyloidosis and Increases Choline Acetyltransferase Expression in the Hippocampus of the APPswePS1dE9 Alzheimer’s Disease Model Mice. PLoS ONE 2017, 12, e0170450. [Google Scholar] [CrossRef]
- Blusztajn, J.K.; Zeisel, S.H.; Wurtman, R.J. Developmental Changes in the Activity of Phosphatidylethanolamine N-Methyltransferases in Rat Brain. Biochem. J. 1985, 232, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Skripuletz, T.; Manzel, A.; Gropengießer, K.; Schäfer, N.; Gudi, V.; Singh, V.; Salinas Tejedor, L.; Jörg, S.; Hammer, A.; Voss, E.; et al. Pivotal Role of Choline Metabolites in Remyelination. Brain 2015, 138 Pt 2, 398–413. [Google Scholar] [CrossRef]
- Tayebati, S.K. Phospholipid and Lipid Derivatives as Potential Neuroprotective Compounds. Molecules 2018, 23, 2257. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. A Brief History of Choline. Ann. Nutr. Metab. 2012, 61, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Tayebati, S.K.; Marucci, G.; Santinelli, C.; Buccioni, M.; Amenta, F. Choline-Containing Phospholipids: Structure-Activity Relationships Versus Therapeutic Applications. Curr. Med. Chem. 2015, 22, 4328–4340. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, M.; Edlund, C.; Alafuzoff, I.; Kristensson, K.; Dallner, G. Lipid Composition in Different Regions of the Brain in Alzheimer’s Disease/Senile Dementia of Alzheimer’s Type. J. Neurochem. 1992, 59, 1646–1653. [Google Scholar] [CrossRef]
- Klein, J. Membrane Breakdown in Acute and Chronic Neurodegeneration: Focus on Choline-Containing Phospholipids. J. Neural. Transm. 2000, 107, 1027–1063. [Google Scholar] [CrossRef]
- Zhang, Y.; Kutateladze, T.G. Diet and the Epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef]
- Anderson, O.S.; Sant, K.E.; Dolinoy, D.C. Nutrition and Epigenetics: An Interplay of Dietary Methyl Donors, One-Carbon Metabolism and DNA Methylation. J. Nutr. Biochem. 2012, 23, 853–859. [Google Scholar] [CrossRef] [PubMed]
- McGarel, C.; Pentieva, K.; Strain, J.J.; McNulty, H. Emerging Roles for Folate and Related B-Vitamins in Brain Health across the Lifecycle. Proc. Nutr. Soc. 2015, 74, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Friso, S.; Udali, S.; De Santis, D.; Choi, S.-W. One-Carbon Metabolism and Epigenetics. Mol. Aspects Med. 2017, 54, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Meck, W.H.; Williams, C.L. Characterization of the Facilitative Effects of Perinatal Choline Supplementation on Timing and Temporal Memory. Neuroreport 1997, 8, 2831–2835. [Google Scholar] [CrossRef]
- Meck, W.H.; Smith, R.A.; Williams, C.L. Pre- and Postnatal Choline Supplementation Produces Long-Term Facilitation of Spatial Memory. Dev. Psychobiol. 1988, 21, 339–353. [Google Scholar] [CrossRef]
- Holmes-McNary, M.Q.; Loy, R.; Mar, M.H.; Albright, C.D.; Zeisel, S.H. Apoptosis Is Induced by Choline Deficiency in Fetal Brain and in PC12 Cells. Brain Res. Dev. Brain Res. 1997, 101, 9–16. [Google Scholar] [CrossRef]
- Niculescu, M.D.; Craciunescu, C.N.; Zeisel, S.H. Dietary Choline Deficiency Alters Global and Gene-Specific DNA Methylation in the Developing Hippocampus of Mouse Fetal Brains. FASEB J. 2006, 20, 43–49. [Google Scholar] [CrossRef]
- Kovacheva, V.P.; Mellott, T.J.; Davison, J.M.; Wagner, N.; Lopez-Coviella, I.; Schnitzler, A.C.; Blusztajn, J.K. Gestational Choline Deficiency Causes Global and Igf2 Gene DNA Hypermethylation by Up-Regulation of Dnmt1 Expression. J. Biol. Chem. 2007, 282, 31777–31788. [Google Scholar] [CrossRef]
- Mehedint, M.G.; Niculescu, M.D.; Craciunescu, C.N.; Zeisel, S.H. Choline Deficiency Alters Global Histone Methylation and Epigenetic Marking at the Re1 Site of the Calbindin 1 Gene. FASEB J. 2010, 24, 184–195. [Google Scholar] [CrossRef]
- Tomizawa, H.; Matsuzawa, D.; Ishii, D.; Matsuda, S.; Kawai, K.; Mashimo, Y.; Sutoh, C.; Shimizu, E. Methyl-Donor Deficiency in Adolescence Affects Memory and Epigenetic Status in the Mouse Hippocampus. Genes Brain Behav. 2015, 14, 301–309. [Google Scholar] [CrossRef]
- Tran, P.V.; Kennedy, B.C.; Lien, Y.-C.; Simmons, R.A.; Georgieff, M.K. Fetal Iron Deficiency Induces Chromatin Remodeling at the Bdnf Locus in Adult Rat Hippocampus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, 276–282. [Google Scholar] [CrossRef]
- Moon, J.; Chen, M.; Gandhy, S.U.; Strawderman, M.; Levitsky, D.A.; Maclean, K.N.; Strupp, B.J. Perinatal Choline Supplementation Improves Cognitive Functioning and Emotion Regulation in the Ts65Dn Mouse Model of Down Syndrome. Behav. Neurosci. 2010, 124, 346–361. [Google Scholar] [CrossRef]
- Nag, N.; Berger-Sweeney, J.E. Postnatal Dietary Choline Supplementation Alters Behavior in a Mouse Model of Rett Syndrome. Neurobiol. Dis. 2007, 26, 473–480. [Google Scholar] [CrossRef]
- Villamor, E.; Rifas-Shiman, S.L.; Gillman, M.W.; Oken, E. Maternal Intake of Methyl-Donor Nutrients and Child Cognition at 3 Years of Age. Paediatr Perinat. Epidemiol. 2012, 26, 328–335. [Google Scholar] [CrossRef]
- Boeke, C.E.; Gillman, M.W.; Hughes, M.D.; Rifas-Shiman, S.L.; Villamor, E.; Oken, E. Choline Intake during Pregnancy and Child Cognition at Age 7 Years. Am. J. Epidemiol. 2013, 177, 1338–1347. [Google Scholar] [CrossRef]
- Signore, C.; Ueland, P.M.; Troendle, J.; Mills, J.L. Choline Concentrations in Human Maternal and Cord Blood and Intelligence at 5 y of Age. Am. J. Clin. Nutr. 2008, 87, 896–902. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective Strategies for Neurological Disorders by Natural Products: An Update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef]
- Teather, L.A.; Wurtman, R.J. Dietary CDP-Choline Supplementation Prevents Memory Impairment Caused by Impoverished Environmental Conditions in Rats. Learn. Mem. 2005, 12, 39–43. [Google Scholar] [CrossRef]
- Cohen, B.M.; Renshaw, P.F.; Stoll, A.L.; Wurtman, R.J.; Yurgelun-Todd, D.; Babb, S.M. Decreased Brain Choline Uptake in Older Adults. An in Vivo Proton Magnetic Resonance Spectroscopy Study. JAMA 1995, 274, 902–907. [Google Scholar] [CrossRef]
- Cummings, J.; Scheltens, P.; McKeith, I.; Blesa, R.; Harrison, J.E.; Bertolucci, P.H.F.; Rockwood, K.; Wilkinson, D.; Wijker, W.; Bennett, D.A.; et al. Effect Size Analyses of Souvenaid in Patients with Alzheimer’s Disease. J. Alzheimers Dis. 2017, 55, 1131–1139. [Google Scholar] [CrossRef]
- West, R.L.; Lee, J.M.; Maroun, L.E. Hypomethylation of the Amyloid Precursor Protein Gene in the Brain of an Alzheimer’s Disease Patient. J. Mol. Neurosci. 1995, 6, 141–146. [Google Scholar] [CrossRef]
- Kennedy, B.P.; Bottiglieri, T.; Arning, E.; Ziegler, M.G.; Hansen, L.A.; Masliah, E. Elevated S-Adenosylhomocysteine in Alzheimer Brain: Influence on Methyltransferases and Cognitive Function. J. Neural Transm. 2004, 111, 547–567. [Google Scholar] [CrossRef]
- Remington, R.; Chan, A.; Paskavitz, J.; Shea, T.B. Efficacy of a Vitamin/Nutriceutical Formulation for Moderate-Stage to Later-Stage Alzheimer’s Disease: A Placebo-Controlled Pilot Study. Am. J. Alzheimers Dis. Other Demen. 2009, 24, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Paskavitz, J.; Remington, R.; Rasmussen, S.; Shea, T.B. Efficacy of a Vitamin/Nutriceutical Formulation for Early-Stage Alzheimer’s Disease: A 1-Year, Open-Label Pilot Study with an 16-Month Caregiver Extension. Am. J. Alzheimers Dis. Other Demen. 2008, 23, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Tayebati, S.K.; Amenta, F. Choline-Containing Phospholipids: Relevance to Brain Functional Pathways. Clin. Chem. Lab. Med. 2013, 51, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Traini, E.; Bramanti, V.; Amenta, F. Choline Alphoscerate (Alpha-Glyceryl-Phosphoryl-Choline) an Old Choline- Containing Phospholipid with a Still Interesting Profile as Cognition Enhancing Agent. Curr. Alzheimer Res. 2013, 10, 1070–1079. [Google Scholar] [CrossRef] [PubMed]
- Teather, L.A.; Wurtman, R.J. Dietary Cytidine (5’)-Diphosphocholine Supplementation Protects against Development of Memory Deficits in Aging Rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 711–717. [Google Scholar] [CrossRef]
- Velazquez, R.; Ferreira, E.; Knowles, S.; Fux, C.; Rodin, A.; Winslow, W.; Oddo, S. Lifelong Choline Supplementation Ameliorates Alzheimer’s Disease Pathology and Associated Cognitive Deficits by Attenuating Microglia Activation. Aging Cell 2019, 18, 13037. [Google Scholar] [CrossRef]
- Velazquez, R.; Ferreira, E.; Winslow, W.; Dave, N.; Piras, I.S.; Naymik, M.; Huentelman, M.J.; Tran, A.; Caccamo, A.; Oddo, S. Maternal Choline Supplementation Ameliorates Alzheimer’s Disease Pathology by Reducing Brain Homocysteine Levels across Multiple Generations. Mol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Froese, D.S.; Fowler, B.; Baumgartner, M.R. Vitamin B12, Folate, and the Methionine Remethylation Cycle-Biochemistry, Pathways, and Regulation. J. Inherit. Metab. Dis. 2018, 42, 673–685. [Google Scholar] [CrossRef]
- Kovalska, M.; Tothova, B.; Kovalska, L.; Tatarkova, Z.; Kalenska, D.; Tomascova, A.; Adamkov, M.; Lehotsky, J. Association of Induced Hyperhomocysteinemia with Alzheimer’s Disease-Like Neurodegeneration in Rat Cortical Neurons After Global Ischemia-Reperfusion Injury. Neurochem. Res. 2018, 43, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Bahous, R.H.; Cosín-Tomás, M.; Deng, L.; Leclerc, D.; Malysheva, O.; Ho, M.-K.; Pallàs, M.; Kaliman, P.; Bedell, B.J.; Caudill, M.A.; et al. Early Manifestations of Brain Aging in Mice Due to Low Dietary Folate and Mild MTHFR Deficiency. Mol. Neurobiol. 2018, 6, 4175–4191. [Google Scholar] [CrossRef] [PubMed]
- Van Wijk, N.; Broersen, L.M.; de Wilde, M.C.; Hageman, R.J.J.; Groenendijk, M.; Sijben, J.W.C.; Kamphuis, P.J.G.H. Targeting Synaptic Dysfunction in Alzheimer’s Disease by Administering a Specific Nutrient Combination. J. Alzheimers Dis. 2014, 38, 459–479. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; Twisk, J.W.R.; Blesa, R.; Scarpini, E.; von Arnim, C.A.F.; Bongers, A.; Harrison, J.; Swinkels, S.H.N.; Stam, C.J.; de Waal, H.; et al. Efficacy of Souvenaid in Mild Alzheimer’s Disease: Results from a Randomized, Controlled Trial. J. Alzheimers Dis. 2012, 31, 225–236. [Google Scholar] [CrossRef] [PubMed]
- de Waal, H.; Stam, C.J.; Lansbergen, M.M.; Wieggers, R.L.; Kamphuis, P.J.G.H.; Scheltens, P.; Maestú, F.; van Straaten, E.C.W. The Effect of Souvenaid on Functional Brain Network Organisation in Patients with Mild Alzheimer’s Disease: A Randomised Controlled Study. PLoS ONE 2014, 9, e86558. [Google Scholar] [CrossRef]
- van Wijk, N.; Slot, R.E.R.; Duits, F.H.; Strik, M.; Biesheuvel, E.; Sijben, J.W.C.; Blankenstein, M.A.; Bierau, J.; van der Flier, W.M.; Scheltens, P.; et al. Nutrients Required for Phospholipid Synthesis Are Lower in Blood and Cerebrospinal Fluid in Mild Cognitive Impairment and Alzheimer’s Disease Dementia. Alzheimers Dement. 2017, 8, 139–146. [Google Scholar] [CrossRef]
- Fransquet, P.D.; Ryan, J. The Current Status of Blood Epigenetic Biomarkers for Dementia. Crit. Rev. Clin. Lab. Sci. 2019, 56, 435–457. [Google Scholar] [CrossRef]
- Mapstone, M.; Cheema, A.K.; Fiandaca, M.S.; Zhong, X.; Mhyre, T.R.; MacArthur, L.H.; Hall, W.J.; Fisher, S.G.; Peterson, D.R.; Haley, J.M.; et al. Plasma Phospholipids Identify Antecedent Memory Impairment in Older Adults. Nat. Med. 2014, 20, 415–418. [Google Scholar] [CrossRef]
- Bahous, R.H.; Jadavji, N.M.; Deng, L.; Cosín-Tomás, M.; Lu, J.; Malysheva, O.; Leung, K.-Y.; Ho, M.-K.; Pallàs, M.; Kaliman, P.; et al. High Dietary Folate in Pregnant Mice Leads to Pseudo-MTHFR Deficiency and Altered Methyl Metabolism, with Embryonic Growth Delay and Short-Term Memory Impairment in Offspring. Hum. Mol. Genet. 2017, 26, 888–900. [Google Scholar] [CrossRef]
- Sittig, L.J.; Herzing, L.B.K.; Xie, H.; Batra, K.K.; Shukla, P.K.; Redei, E.E. Excess Folate during Adolescence Suppresses Thyroid Function with Permanent Deficits in Motivation and Spatial Memory. Genes Brain Behav. 2012, 11, 193–200. [Google Scholar] [CrossRef]
- Valera-Gran, D.; García de la Hera, M.; Navarrete-Muñoz, E.M.; Fernandez-Somoano, A.; Tardón, A.; Julvez, J.; Forns, J.; Lertxundi, N.; Ibarluzea, J.M.; Murcia, M.; et al. Folic Acid Supplements during Pregnancy and Child Psychomotor Development after the First Year of Life. JAMA Pediatr. 2014, 168, 142611. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, G.S.; Scarmeas, N. Dietary Interventions in Mild Cognitive Impairment and Dementia. Dialogues Clin. Neurosci. 2019, 21, 69–82. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Description | Outcomes | References |
|---|---|---|
| Prenatal and postnatal choline supplementation | Improved memory-related tasks in offspring | [46,47,75,76] |
| Deficiency of folate, methionine and choline (FMCD mice) | Impaired learning and memory Hypermethylation of Gria1 promoter | [81] |
| Gestational choline supplementation in iron-deficient (ID) rat | Alteration in hippocampal Bdnf promoter methylation | [82] |
| Perinatal choline supplementation in a mouse model of Down syndrome | Improved cognitive functions in offspring | [83] |
| Long-term dietary supplementation of choline in an APP/PS1 mouse model of AD Choline supplementation in a mouse model of AD | Reduced accumulation of amyloid plaques in Reduced α7nAchR expression Reduced microglia activation Improved spatial memory Improved cognitive functions and reduced levels of homocysteine across generations | [99,100] |
| Low dietary folate in Mthfr+/+ mice | Altered expression of synaptic markers, Bdnf and epigenetic enzymes | [103] |
| Supplementation of choline, VitB12, VitB6 and folate in early AD patients | Improved memory and enhanced synaptic signaling | [104] |
| Supplementation of SOUVENAID in early AD patients | Improved memory scores | [105,106] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bekdash, R.A. Neuroprotective Effects of Choline and Other Methyl Donors. Nutrients 2019, 11, 2995. https://doi.org/10.3390/nu11122995
Bekdash RA. Neuroprotective Effects of Choline and Other Methyl Donors. Nutrients. 2019; 11(12):2995. https://doi.org/10.3390/nu11122995
Chicago/Turabian StyleBekdash, Rola A. 2019. "Neuroprotective Effects of Choline and Other Methyl Donors" Nutrients 11, no. 12: 2995. https://doi.org/10.3390/nu11122995
APA StyleBekdash, R. A. (2019). Neuroprotective Effects of Choline and Other Methyl Donors. Nutrients, 11(12), 2995. https://doi.org/10.3390/nu11122995