Changes in Human Milk Fatty Acid Composition during Lactation: The Ulm SPATZ Health Study

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Data Collection and Measurements
2.3. Fatty Acid Analysis
2.4. Statistical Analysis
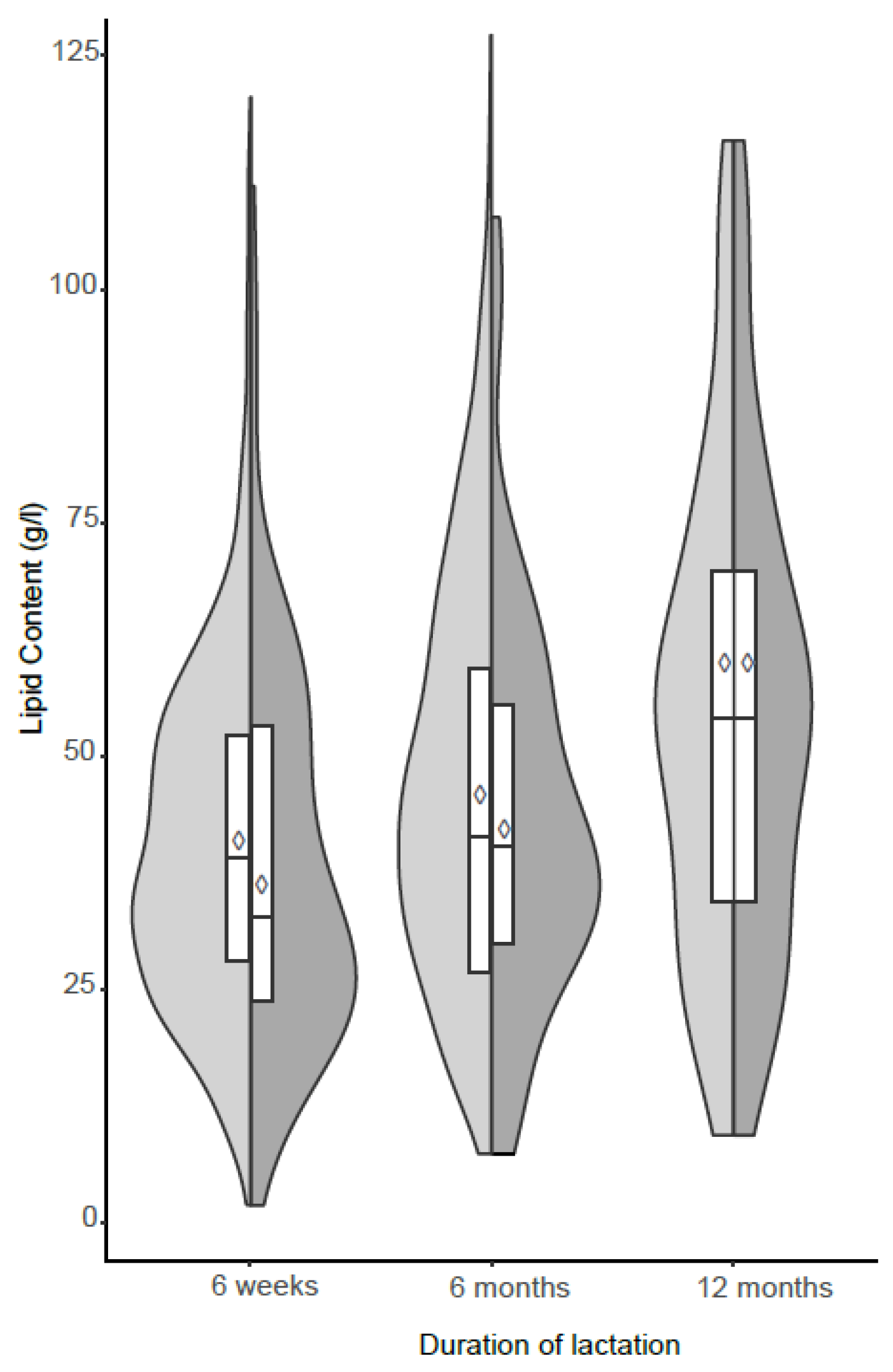
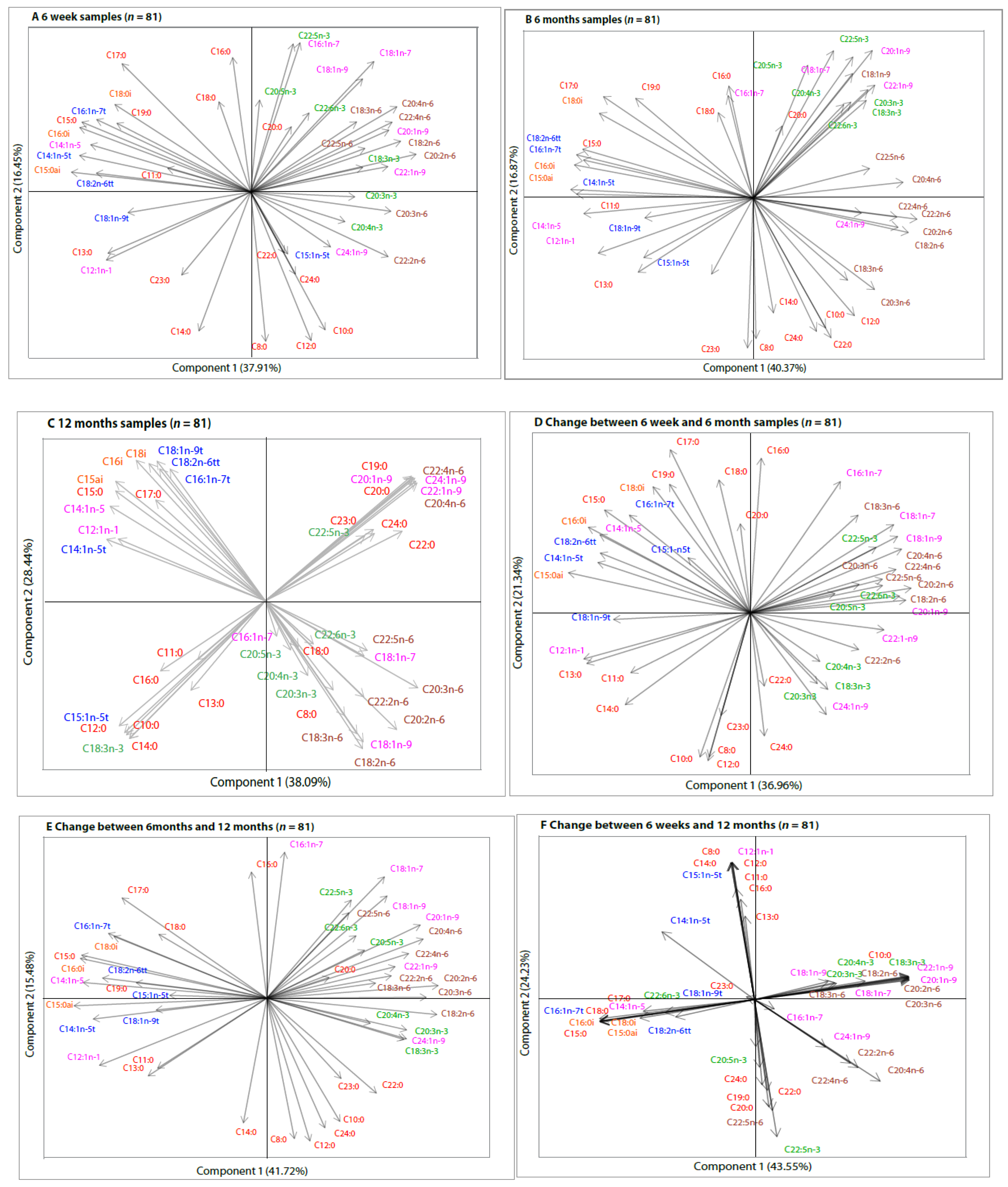
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatric Clin. 2013, 60, 49–74. [Google Scholar]
- Grote, V.; Verduci, E.; Vecchi, F.; Contarini, G.; Giovannini, M.; Koletzko, B. Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur. J. Clin. Nutr. 2016, 70, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Bhutta, Z.A.; Cai, W.; Cruchetm, S.; Guindim, M.E.; Fuchs, G.J. Compositional Requirements of Follow-Up Formula for Use in Infancy: Recommendations of an International Expert Group Coordinated by the Early Nutrition Academy. Ann. Nutr. Metab. 2013, 62, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Impact of maternal diet on human milk composition and neurological development of infants. Am. J. Clin. Nutr. 2014, 99, 734S–741S. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Dhar, P.; Ghosh, S. Comparative evaluation of essential fatty acid composition of mothers’ milk of some urban and suburban regions of West Bengal, India. Int. J. Food Sci. Nutr. 2012, 63, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Olafsdottir, A.S.; Thorsdottir, I.; Wagner, K.H.; Elmadfa, I. Polyunsaturated fatty acids in the diet and breast milk of lactating icelandic women with traditional fish and cod liver oil consumption. Ann. Nutr. Metab. 2006, 50, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, X.; Zhou, B.; Jiang, A.C.; Chai, L. An updated review of worldwide levels of docosahexaenoic and arachidonic acid in human breast milk by region. Public Health Nutr. 2016, 19, 2675–2687. [Google Scholar] [CrossRef]
- Much, D.; Brunner, S.; Vollhardt, C.; Schmid, D.; Sedlmeier, E.-M.; Brüderl, M. Breast milk fatty acid profile in relation to infant growth and body composition: Results from the INFAT study. Pediatric Res. 2013, 74, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Szabo, E.; Boehm, G.; Beermann, C.; Weyermann, M.; Brenner, H.; Rothenbacher, D. Fatty acid profile comparisons in human milk sampled from the same mothers at the sixth week and the sixth month of lactation. J. Pediatric Gastroenterol. Nutr. 2010, 50, 316–320. [Google Scholar] [CrossRef]
- Fidler, N.; Sauerwald, T.; Pohl, A.; Demmelmair, H.; Koletzko, B. Docosahexaenoic acid transfer into human milk after dietary supplementation: A randomized clinical trial. J. Lipid Res. 2000, 41, 1376–1388. [Google Scholar]
- Agostoni, C.; Marangoni, F.; Lammardo, A.M.; Galli, C.; Giovannini, M.; Riva, E. Long-chain polyunsaturated fatty acid concentrations in human hindmilk are constant throughout twelve months of lactation. Adv. Exp. Med. Biol. 2001, 501, 157–161. [Google Scholar] [PubMed]
- Marangoni, F.; Agostoni, C.; Lammardo, A.M.; Giovannini, M.; Galli, C.; Riva, E. Polyunsaturated fatty acid concentrations in human hindmilk are stable throughout 12-months of lactation and provide a sustained intake to the infant during exclusive breastfeeding: An Italian study. Br. J. Nutr. 2000, 84, 103–109. [Google Scholar] [PubMed]
- Barreiro, R.; Regal, P.; López-Racamonde, O.; Cepeda, A.; Fente, C.A. Comparison of the fatty acid profile of Spanish infant formulas and Galician women breast milk. J. Physiol. Biochem. 2018, 74, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Aitchison, J.J.; Egozcue, J. Compositional Data Analysis: Where Are We and Where Should We Be Heading? Math. Geol. 2005, 37, 829–850. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Voortman, T.; Tielemans, M.J.; Stroobant, W.; Schoufour, J.D.; Kiefte-de Jong, J.C.; Steenweg-de Graaff, J. Plasma fatty acid patterns during pregnancy and child’s growth, body composition, and cardiometabolic health: The Generation R Study. Clin. Nutr. 2018, 37, 984–992. [Google Scholar] [CrossRef]
- Logan, C.; Zittel, T.; Striebel, S.; Reister, F.; Brenner, H.; Rothenbacher, D. Changing Societal and Lifestyle Factors and Breastfeeding Patterns Over Time. Pediatrics 2016, 137, e20154473. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Beermann, C.; Green, A.; Möbius, M.; Schmitt, J.J.; Boehm, G. Lipid class separation by HPLC combined with GC FA analysis: Comparison of seed lipid compositions from different Brassica napus L. varieties. J. Am. Oil Chem. Soc. 2003, 80, 747–753. [Google Scholar] [CrossRef]
- Lucas, A.; Gibbs, J.A.; Lyster, R.L.; Baum, J.D. Creamatocrit: Simple clinical technique for estimating fat concentration and energy value of human milk. Br. Med. J. 1978, 1, 1018–1020. [Google Scholar] [CrossRef]
- Jones, E.; Dimmock, P.W.; Spencer, S.A. A randomised controlled trial to compare methods of milk expression after preterm delivery. Arch. Dis. Child.-Fetal Neonatal Ed. 2001, 85, F91–F95. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.A.; Brandt, S.; Wabitsch, M.; Brenner, H.; Wiens, F.; Stahl, B. New approach shows no association between maternal milk fatty acid composition and childhood wheeze or asthma. Allergy 2017, 72, 374–833. [Google Scholar] [CrossRef]
- Da Cunha, J.; Macedo da Costa, T.H.; Ito, M.K. Influences of maternal dietary intake and suckling on breast milk lipid and fatty acid composition in low-income women from Brasilia, Brazil. Early Hum. Dev. 2005, 81, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Mandel, D.; Lubetzky, R.; Dollberg, S.; Barak, S.; Mimouni, F.B. Fat and Energy Contents of Expressed Human Breast Milk in Prolonged Lactation. Pediatrics 2005, 116, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Keating, E.M.; Curtis, B.A.; Slusher, T.M. Maternal milk volume and breast milk expression: Implications for diet and nutrition in infants. In Handbook of Dietary and Nutritional Aspects of Human Breast Milk; Wageningen Academic Publishers: Wageningen, The Nederlands, 2013; Volume 5, pp. 193–214. [Google Scholar] [CrossRef]
- Ros-Freixedes, R.; Estany, J. On the Compositional Analysis of Fatty Acids in Pork. J. Agric. Biol. Environ. Stat. 2014, 19, 136–155. [Google Scholar] [CrossRef]
- Imamura, F.; Sharp, S.J.; Koulman, A.; Schulze, M.B.; Kröger, J.; Griffin, J.L. A combination of plasma phospholipid fatty acids and its association with incidence of type 2 diabetes: The EPIC-InterAct case-cohort study. PLoS Med. 2017, 14, e1002409. [Google Scholar] [CrossRef] [PubMed]
- Siziba, L.P.; Baumgartner, J.; Ricci, C.; Jacobs, A.; Rothman, M.; Matsungo, T.M. Associations of plasma total phospholipid fatty acid patterns with feeding practices, growth, and psychomotor development in 6-month-old South African infants. Matern. Child Nutr. 2017, 15, e12763. [Google Scholar] [CrossRef]
- Brenna, J.T.; Plourde, M.; Stark, K.D.; Jones, P.J.; Lin, Y.-H. Best practices for the design, laboratory analysis, and reporting of trials involving fatty acids. Am. J. Clin. Nutr. 2018, 108, 211–227. [Google Scholar] [CrossRef]
- Butts, C.A.; Hedderley, D.I.; Herath, T.D.; Paturi, G.; Glyn-Jones, S.; Wiens, F. Human Milk Composition and Dietary Intakes of Breastfeeding Women of Different Ethnicity from the Manawatu-Wanganui Region of New Zealand. Nutrients 2018, 10, 1231. [Google Scholar] [CrossRef]
- Antonakou, A.; Skenderi, K.P.; Chiou, A.; Anastasiou, C.A.; Bakoula, C.; Matalas, A.L. Breast milk fat concentration and fatty acid pattern during the first six months in exclusively breastfeeding Greek women. Eur. J. Nutr. 2013, 52, 963–973. [Google Scholar] [CrossRef]
- Truthmann, J.; Richter, A.; Thiele, S.; Drescher, L.; Roosen, J.; Mensink, G.B. Associations of dietary indices with biomarkers of dietary exposure and cardiovascular status among adolescents in Germany. Nutr. Metab. 2012, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, B.; West, J.A.; Koulman, A. A Review of Odd-Chain Fatty Acid Metabolism and the Role of Pentadecanoic Acid (C15:0) and Heptadecanoic Acid (C17:0) in Health and Disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.; Thijs, C.; Rist, L.; Simões-Wüst, A.P.; Huber, M.; Steinhart, H. Trans Fatty Acids in Human Milk are an Indicator of Different Maternal Dietary Sources Containing Trans Fatty Acids. Lipids 2010, 45, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Visentainer, J.V.; Santos, O.O.; Maldaner, L.; Zappielo, C.; Neia, V.L. Lipids and Fatty Acids in Human Milk: Benefits and Analysis. Biochem. Health Benefits Fat. Acids 2018, 1–22. [Google Scholar] [CrossRef]
- Innis, S.M.; King, D.J. trans Fatty acids in human milk are inversely associated with concentrations of essential all-cis n−6 and n−3 fatty acids and determine trans, but not n−6 and n−3, fatty acids in plasma lipids of breast-fed infants. Am. J. Clin. Nutr. 1999, 70, 383–390. [Google Scholar] [CrossRef]
- Hudgins, L.C.; Hellerstein, M.; Seidman, C.; Neese, R.J.; Hirsch, J. Human fatty acid synthesis is stimulated by a eucaloric low fat, high carbohydrate diet. J. Clin. Investig. 1996, 97, 2081–2091. [Google Scholar] [CrossRef]
- Gardner, A.S.; Rahman, I.A.; Lai, C.T.; Hepworth, A.; Trengove, N.; Hartmann, P.E. Changes in Fatty Acid Composition of Human Milk in Response to Cold-Like Symptoms in the Lactating Mother and Infant. Nutrients 2017, 9, 1034. [Google Scholar] [CrossRef]
- Høstmark, A.T.; Haug, A. Percentages of oleic acid and arachidonic acid are inversely related in phospholipids of human sera. Lipids Health Dis. 2013, 12, 106. [Google Scholar] [CrossRef]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatric Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef]
- Hadley, K.; Ryan, A.; Forsyth, S.; Gautier, S.; Salem, N. The Essentiality of Arachidonic Acid in Infant Development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef]
 (6 weeks (n = 706), 6 months (n = 483), 12 months (n = 81)),
(6 weeks (n = 706), 6 months (n = 483), 12 months (n = 81)),  Sub-sampl e size (n = 81).
(6 weeks (n = 706), 6 months (n = 483), 12 months (n = 81)), Sub-sampl e size (n = 81).
Sub-sampl e size (n = 81).
(6 weeks (n = 706), 6 months (n = 483), 12 months (n = 81)), Sub-sampl e size (n = 81).


{kind=link}
{kind=link}
| Characteristic | All 6 Weeks Samples (n = 706) | All 6 Months Samples (n = 483) | All 12 Months Samples (n = 83) | |||
|---|---|---|---|---|---|---|
| n | % or Mean | n | % or Mean | n | % or Mean | |
| Age | 706 | 33.1 | 483 | 33.5 | 83 | 34.4 |
| Gestation period | ||||||
| Early (37 or 38 weeks) | 146 | 22.5% | 96 | 21.5% | 18 | 22.8% |
| Full (39 or 40 weeks) | 397 | 61.3% | 275 | 61.5% | 47 | 59.5% |
| Late or Post (41 or 42 weeks) | 105 | 16.2% | 76 | 17.0% | 14 | 17.7% |
| Parity (n births of fetus ≥24 weeks) | ||||||
| 0 births | 368 | 52.2% | 252 | 52.2% | 39 | 47.0% |
| ≥1 birth | 337 | 47.8% | 231 | 47.8% | 44 | 53.0% |
| Delivery mode | ||||||
| Vaginal spontaneous | 480 | 68.1% | 344 | 71.2% | 60 | 72.3% |
| Elective caesarean | 69 | 9.8% | 39 | 8.1% | 9 | 10.8% |
| Emergency caesarean | 90 | 12.8% | 56 | 11.6% | 9 | 10.8% |
| Vaginal assisted | 66 | 9.4% | 44 | 9.1% | 5 | 6.0% |
| Maternal pre-pregnancy body mass index (BMI) category | ||||||
| Underweight (BMI < 18.50) | 12 | 1.8% | 6 | 1.3% | 3 | 3.8% |
| Normal (18.50 ≤ BMI < 25.00) | 443 | 64.8% | 325 | 69.3% | 55 | 68.8% |
| Overweight (25.00 ≤ BMI < 30.00) | 150 | 21.9% | 93 | 19.8% | 15 | 18.8% |
| Obese (BMI > 30.00) | 79 | 11.5% | 45 | 9.6% | 7 | 8.8% |
| Maternal BMI category (6 weeks) | ||||||
| Underweight | 6 | 1.0% | 3 | 0.7% | ||
| Normal | 364 | 58.0% | 268 | 60.4% | 45 | 56.3% |
| Overweight | 189 | 30.1% | 131 | 29.5% | 27 | 33.8% |
| Obese | 69 | 11.0% | 42 | 9.5% | 8 | 10.0% |
| Maternal BMI category (6 months) | ||||||
| Underweight | 15 | 2.6% | 10 | 2.4% | 3 | 3.9% |
| Normal | 360 | 62.8% | 277 | 66.4% | 50 | 64.9% |
| Overweight | 136 | 23.7% | 92 | 22.1% | 18 | 23.4% |
| Obese | 62 | 10.8% | 38 | 9.1% | 6 | 7.8% |
| Education level | ||||||
| Low | 43 | 6.2% | 16 | 3.4% | 3 | 3.7% |
| Intermediate | 196 | 28.2% | 115 | 24.2% | 13 | 16.0% |
| High | 455 | 65.6% | 345 | 72.5% | 65 | 80.2% |
| Occupation | ||||||
| Leadership/professionals | 148 | 30.3% | 102 | 30.2% | 20 | 34.5% |
| Intermediate position | 223 | 45.6% | 160 | 47.3% | 27 | 46.6% |
| Skilled manuals/non-manuals | 88 | 18.0% | 63 | 18.6% | 8 | 13.8% |
| Unskilled/semi-skilled | 18 | 3.7% | 5 | 1.5% | 1 | 1.7% |
| Self-employed | 12 | 2.5% | 8 | 2.4% | 2 | 3.4% |
| Indicator food index (child, age 4) | ||||||
| Neutral | 221 | 44.2% | 153 | 40.7% | 28 | 38.9% |
| Favorable | 279 | 55.8% | 223 | 59.3% | 44 | 61.1% |
| Indicator food index (child, age 5) | ||||||
| Neutral | 198 | 45.3% | 150 | 44.9% | 29 | 43.9% |
| Favorable | 237 | 54.2% | 182 | 54.5% | 37 | 56.1% |
| Unfavorable | 2 | 0.5% | 2 | 0.6% | ||
| Maternal alcohol consumption | ||||||
| Never drinker | 165 | 23.4% | 114 | 23.6% | 40 | 48.2% |
| Abstinent drinker | 203 | 28.8% | 173 | 35.8% | 24 | 28.9% |
| Resumed drinking at 6 weeks to 6 months | 150 | 21.2% | 90 | 18.6% | 8 | 9.6% |
| Resumed drinking by 6 weeks | 115 | 16.3% | 73 | 15.1% | 10 | 12.0% |
| Undetermined drinking | 73 | 10.3% | 33 | 6.8% | 1 | 1.2% |
| Smoking relapse status by 2 years | ||||||
| Non-smoker | 539 | 78.3% | 394 | 82.8% | 74 | 89.2% |
| Relapse by 6 weeks | 48 | 7.0% | 16 | 3.4% | 2 | 2.4% |
| Relapse by 6 months | 22 | 3.2% | 7 | 1.5% | ||
| Relapse 6 months to 2 years | 27 | 3.9% | 18 | 3.8% | ||
| Abstained up to 2 years | 52 | 7.6% | 41 | 8.6% | 7 | 8.4% |
| Fatty Acid | Common Name | 6 Weeks | 6 Months | 12 Months | p Value a | p Value b | p Value c | |||
|---|---|---|---|---|---|---|---|---|---|---|
| C8:0 | Caprylic | −0.07 | (0.57) | 0.06 | (0.47) | 0.01 | (0.47) | 0.2109 | 0.3500 | 0.7188 |
| C10:0 | Capric | 2.03 | (0.26) | 2.09 | (0.22) | 2.10 | (0.25) | 0.1135 | 0.9546 | 0.1166 |
| C11:0 | Undecylic | −2.25 | (0.33) | −2.21 | (0.34) | −2.28 | (0.34) | 0.3313 | 0.1943 | 0.6914 |
| C12:0 | Lauric | 3.25 | (0.36) | 3.42 | (0.31) | 3.67 | (0.29) | 0.0021 | <.0001 | <.0001 |
| C13:0 | Tridecylic | −1.61 | (0.24) | −1.55 | (0.42) | −1.50 | (0.59) | 0.0125 | 0.0357 | <.0001 |
| C14:0 | Myristic | 3.50 | (0.23) | 3.68 | (0.20) | 3.98 | (0.26) | <.0001 | <.0001 | <.0001 |
| C15:0 | Pentadecylic | 0.80 | (0.25) | 0.86 | (0.26) | 0.82 | (0.25) | 0.1090 | 0.4683 | 0.3561 |
| C16:0 | Palmitic | 4.88 | (0.15) | 4.90 | (0.14) | 4.91 | (0.15) | 0.6024 | 0.3884 | 0.1903 |
| C17:0 | Margaric | 0.61 | (0.17) | 0.64 | (0.17) | 0.65 | (0.16) | 0.1620 | 0.6212 | 0.0366 |
| C18:0 | Stearic | 3.69 | (0.21) | 3.72 | (0.21) | 3.72 | (0.18) | 0.4243 | 0.8974 | 0.3372 |
| C19:0 | Nonadecylic acid | −1.84 | (0.27) | −1.80 | (0.26) | −1.77 | (0.21) | 0.1869 | 0.9213 | 0.1300 |
| C20:0 | Arachidic | 0.16 | (0.21) | 0.14 | (0.21) | 0.16 | (0.23) | 0.4986 | 0.6791 | 0.7771 |
| C22:0 | Behenic | −0.68 | (0.26) | −0.70 | (0.24) | −0.64 | (0.34) | 0.5421 | 0.5839 | 0.9626 |
| C23:0 | Tricosylic | −3.93 | (1.99) | −3.36 | (1.63) | −3.63 | (1.94) | 0.0489 | 0.3875 | 0.2778 |
| C24:0 | Lignoceric | −1.16 | (0.51) | −1.18 | (0.46) | −1.05 | (0.47) | 0.5388 | 0.0927 | 0.2675 |
| C12:1n-1 | −2.23 | (0.34) | −2.18 | (0.35) | −2.20 | (0.38) | 0.4302 | 0.8789 | 0.5050 | |
| C14:1n-5 | Myristoleic | 0.48 | (0.28) | 0.53 | (0.33) | 0.45 | (0.33) | 0.3022 | 0.1776 | 0.7063 |
| C16:1n-7 | Palmitoleic | 2.55 | (0.32) | 2.50 | (0.27) | 2.46 | (0.26) | 0.2975 | 0.2921 | 0.0640 |
| C18:1n-7 | Vaccenic | 2.19 | (0.24) | 2.15 | (0.19) | 2.14 | (0.21) | 0.0966 | 0.8250 | 0.0842 |
| C18:1n-9 | Oleic | 5.26 | (0.20) | 5.29 | (0.16) | 5.25 | (0.19) | 0.4828 | 0.1189 | 0.4663 |
| C20:1n-9 | Eicosenoic | 0.86 | (0.24) | 0.76 | (0.23) | 0.80 | (0.26) | 0.0035 | 0.3658 | 0.0366 |
| C22:1n-9 | Erucic | −0.89 | (0.29) | −1.02 | (0.29) | −0.90 | (0.32) | 0.0057 | 0.0133 | 0.6572 |
| C24:1n-9 | Nervonic | −1.62 | (0.95) | −1.60 | (0.57) | −1.38 | (0.56) | 0.2704 | 0.0202 | 0.3101 |
| C15:0 anteiso | Anteisopentadecylic | −0.44 | (0.37) | −0.33 | (0.40) | −0.38 | (0.45) | 0.0221 | 0.5656 | 0.0845 |
| C16:0 iso | Isopalmitic | −0.72 | (0.31) | −0.64 | (0.33) | −0.69 | (0.34) | 0.0728 | 0.3149 | 0.3330 |
| C18:0 iso | −1.89 | (0.24) | −1.86 | (0.23) | −1.84 | (0.24) | 0.1615 | 0.8329 | 0.1142 | |
| C14:1n-5t | Myristelaidic | −4.26 | (0.56) | −4.20 | (0.51) | −4.25 | (0.67) | 0.3483 | 0.3893 | 0.9213 |
| C15:1n-5t | −4.23 | (0.71) | −4.14 | (0.48) | −4.19 | (0.74) | 0.5931 | 0.8921 | 0.5851 | |
| C16:1n-7t | −1.58 | (0.34) | −1.58 | (0.35) | −1.60 | (0.38) | 0.6177 | 0.9027 | 0.7263 | |
| C18:1n-9t | Elaidic | 0.89 | (0.54) | 1.01 | (0.62) | 1.04 | (0.54) | 0.0966 | 0.9439 | 0.0386 |
| C18:2n-6tt | Linolelaidic | −0.90 | (0.67) | −0.73 | (0.50) | −0.97 | (0.88) | 0.0155 | 0.0510 | 0.5908 |
| C18:2n-6 | Linoleic | 4.03 | (0.30) | 4.08 | (0.33) | 4.06 | (0.33) | 0.4612 | 0.6236 | 0.8525 |
| C18:3n-6 | γ-linolenic | −0.42 | (0.39) | -0.61 | (0.37) | −0.93 | (0.44) | 0.0029 | <.0001 | <.0001 |
| C20:2n-6 | Eicosadienoic | 0.36 | (0.26) | 0.19 | (0.28) | 0.24 | (0.29) | <.0001 | 0.1177 | 0.0017 |
| C20:3n-6 | Dihomo-γ-linolenic | 0.66 | (0.27) | 0.38 | (0.26) | 0.31 | (0.29) | <.0001 | 0.1131 | <.0001 |
| C20:4n-6 | Arachidonic | 0.80 | (0.27) | 0.75 | (0.24) | 0.79 | (0.28) | 0.1954 | 0.3847 | 0.6236 |
| C22:2n-6 | Docosadienoic | −2.13 | (0.95) | −2.50 | (1.12) | −2.30 | (0.93) | <.0001 | 0.0667 | 0.0149 |
| C22:4n-6 | Adrenic | −0.97 | (0.65) | −0.99 | (0.29) | −0.85 | (0.41) | 0.0857 | 0.0014 | 0.1321 |
| C22:5n-6 | Osbond | −1.62 | (0.54) | −1.71 | (0.35) | −1.74 | (0.48) | 0.0122 | 0.4954 | 0.0464 |
| C18:3n-3 | α -linoleic | 1.59 | (0.40) | 1.67 | (0.42) | 1.56 | (0.36) | 0.2960 | 0.1237 | 0.7630 |
| C20:3n-3 | Dihomo-α-linoleic | −1.54 | (0.49) | −1.71 | (0.33) | −1.65 | (0.32) | 0.0002 | 0.3684 | 0.0038 |
| C20:4n-3 | −0.74 | (0.37) | −1.10 | (0.35) | −1.32 | (0.43) | <.0001 | 0.0008 | <.0001 | |
| C20:5n-3 | Eicosapentaenoic | −1.01 | (0.35) | −1.03 | (0.43) | −1.19 | (0.39) | 0.6142 | 0.0194 | 0.0021 |
| C22:5n-3 | Docosapentanoic | −0.17 | (0.23) | −0.16 | (0.21) | −0.06 | (0.20) | 0.7887 | 0.0030 | 0.0027 |
| C22:6n-3 | Docosahexaenoic | 0.30 | (0.44) | 0.12 | (0.52) | 0.20 | (0.43) | 0.0073 | 0.2025 | 0.1004 |
| Fatty Acid | 6 Weeks (n = 81) | 6 Months (n = 81) | 12 Months (n = 81) | |||
|---|---|---|---|---|---|---|
| LS Means (95% CL) a | Estimate | p Value | Estimate | p Value | ||
| 12 Months Derived | ||||||
| Group 1 | 1.4 | (1.4, 1.4) | 0.037 | 0.4470 | −0.011 | 0.8028 |
| Group 2 | −1.5 | (−1.6, −1.5) | 0.049 | 0.0062 | 0.122 | <0.0001 |
| Group 3 | 1.8 | (1.8, 1.9) | −0.007 | 0.7908 | −0.067 | 0.0029 |
| Group 4 | −1.7 | (−1.8, −1.7) | −0.079 | 0.0073 | −0.044 | 0.0998 |
| SFA | 2.3 | (2.4, 2.4) | 0.038 | 0.1476 | 0.105 | <0.0001 |
| MUFA | 2.3 | (2.2, 2.3) | −0.021 | 0.4191 | −0.080 | 0.0012 |
| Trans-FA | −2 | (−2.1, −1.8) | 0.073 | 0.4245 | 0.090 | 0.2813 |
| BCFAs | −2.9 | (−3.0, −2.8) | 0.041 | 0.5426 | −0.039 | 0.5039 |
| PUFA | 1.2 | (1.1, 1.2) | 0.017 | 0.6632 | −0.034 | 0.2977 |
| ∑n-3 PUFA | −1 | (−1.1, −0.9) | −0.035 | 0.4612 | −0.111 | 0.0125 |
| ∑n-3 LCPUFA | −1.9 | (−2.0, −1.8) | −0.158 | 0.0003 | −0.160 | <0.0001 |
| ∑n-6 PUFA | 1 | (1.0, 1.1) | 0.024 | 0.5694 | −0.022 | 0.5128 |
| ∑n-6 LCPUFA | −1.2 | (−1.3, −1.2) | −0.178 | <.0001 | −0.171 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siziba, L.P.; Lorenz, L.; Stahl, B.; Mank, M.; Marosvölgyi, T.; Decsi, T.; Rothenbacher, D.; Genuneit, J. Changes in Human Milk Fatty Acid Composition during Lactation: The Ulm SPATZ Health Study. Nutrients 2019, 11, 2842. https://doi.org/10.3390/nu11122842
Siziba LP, Lorenz L, Stahl B, Mank M, Marosvölgyi T, Decsi T, Rothenbacher D, Genuneit J. Changes in Human Milk Fatty Acid Composition during Lactation: The Ulm SPATZ Health Study. Nutrients. 2019; 11(12):2842. https://doi.org/10.3390/nu11122842
Chicago/Turabian StyleSiziba, Linda P., Leonie Lorenz, Bernd Stahl, Marko Mank, Tamas Marosvölgyi, Tamas Decsi, Dietrich Rothenbacher, and Jon Genuneit. 2019. "Changes in Human Milk Fatty Acid Composition during Lactation: The Ulm SPATZ Health Study" Nutrients 11, no. 12: 2842. https://doi.org/10.3390/nu11122842
APA StyleSiziba, L. P., Lorenz, L., Stahl, B., Mank, M., Marosvölgyi, T., Decsi, T., Rothenbacher, D., & Genuneit, J. (2019). Changes in Human Milk Fatty Acid Composition during Lactation: The Ulm SPATZ Health Study. Nutrients, 11(12), 2842. https://doi.org/10.3390/nu11122842