Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review


Abstract
:1. Introduction
- Polyphenols have been shown to have a wide range of beneficial effects, of note red wine, rich in flavonoids, phenolic polymers and resveratrol are promising as possible targets for further investigation.
- As well as giving a state-of- the-art review on the mechanisms through which polyphenols exert their beneficial effects focussing on CYP7A1 and bile acid metabolism, critical points that will be of benefit to clinical nutritionists, academic experts in the area of bioactive food compounds, and possible stakeholders have also been raised in this review namely;
- While polyphenols do have effects on bile acid metabolism, it should be born in mind that species differences, time of cull, dose, and length of treatment can also affect the results and as such, may lead to discrepancies between studies. As such, there is a need to standardise animal studies, ensuring that these factors are correctly reported.
- Polyphenols confer their beneficial effects partly through altering the microbiome, thus they could have useful prebiotic-like functions.
- There is a lack of data on the effects conferred by the parent compound versus those of their metabolic products, and future studies should aim to determine the effects of both the parent compounds and their metabolites, with particular emphasis on the gut microbiota.
- Further studies are required in humans to substantiate the mechanisms proposed using animal and cell culture models.
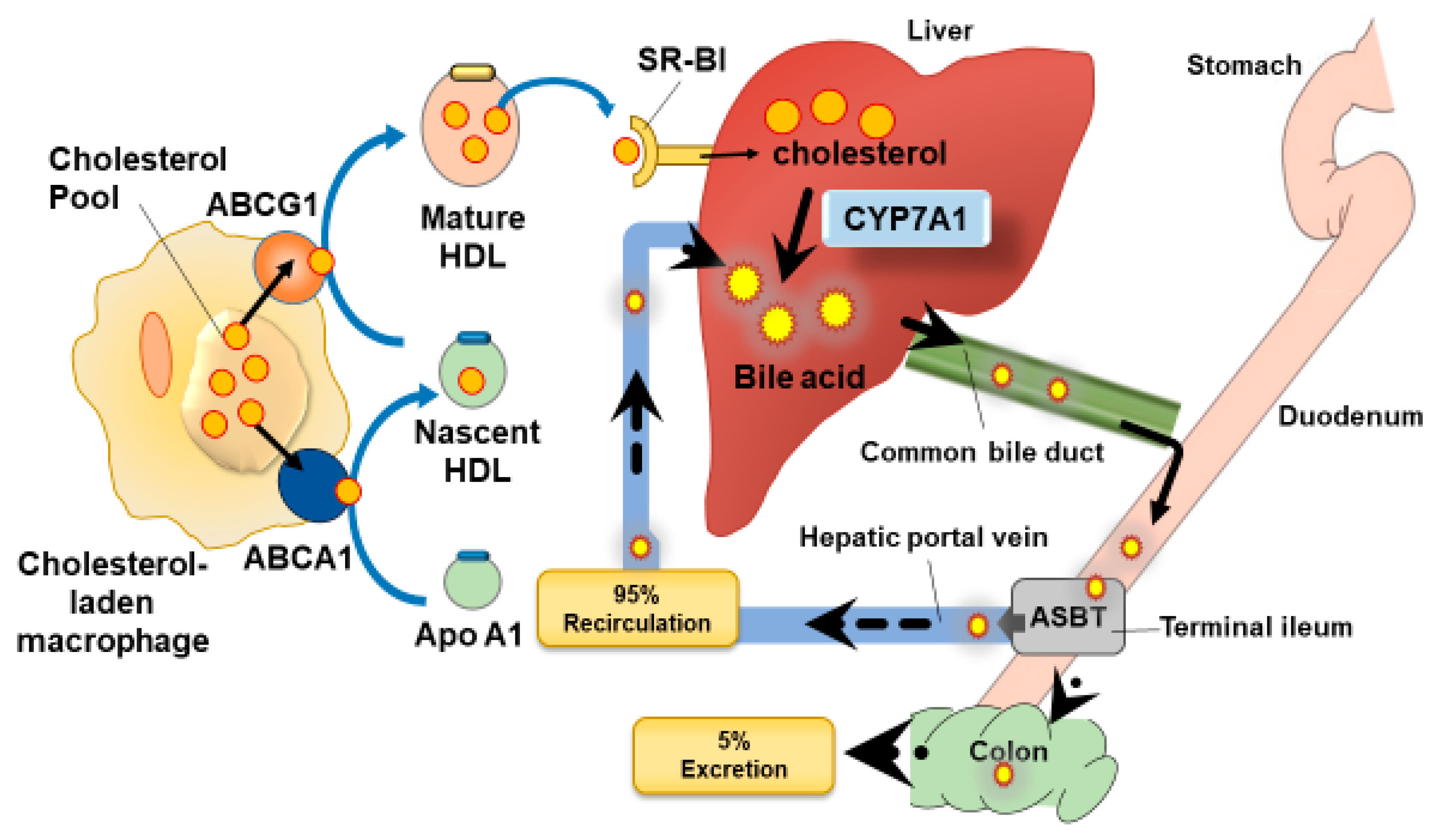
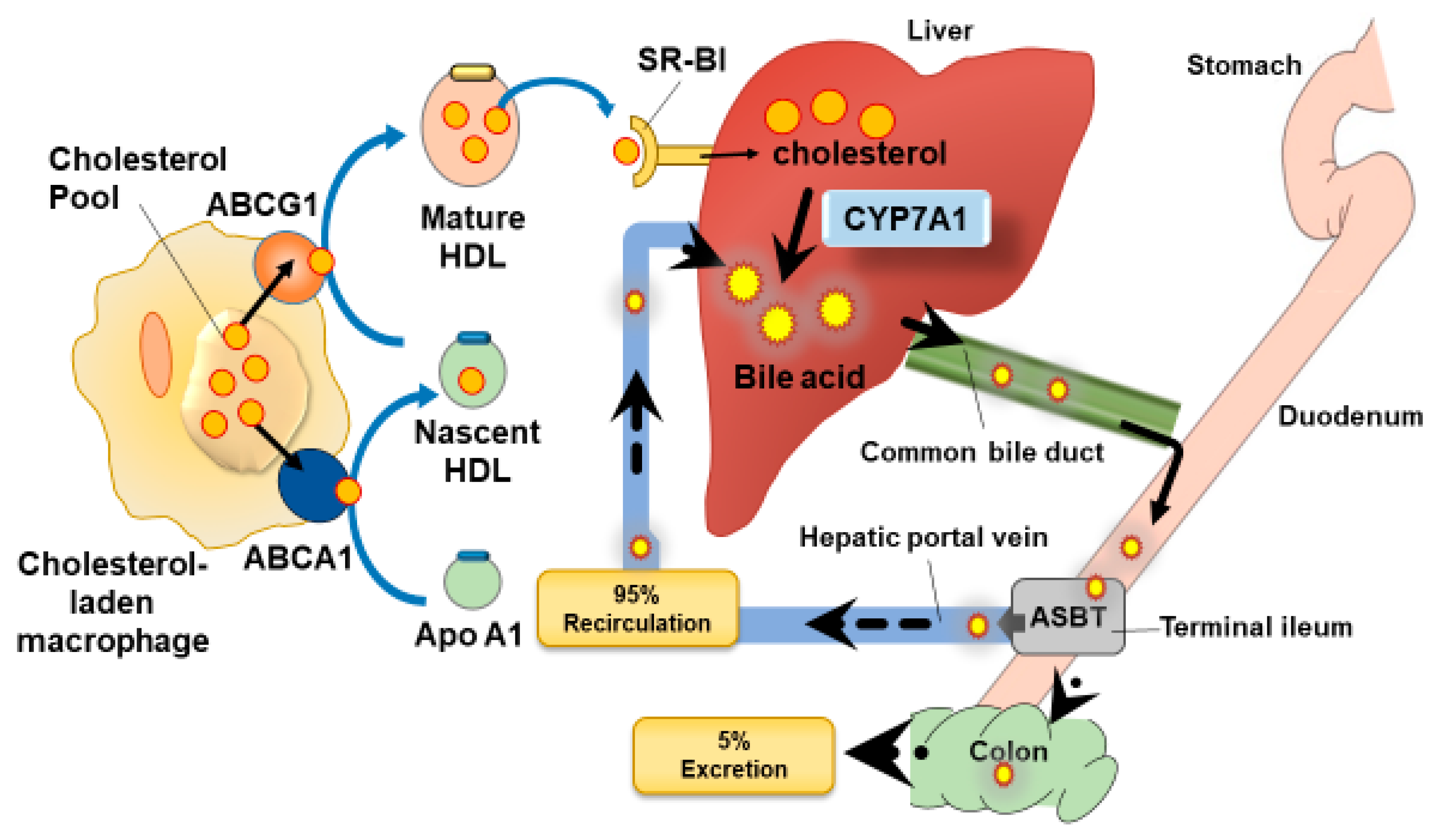
1.1. Reverse Cholesterol Transport
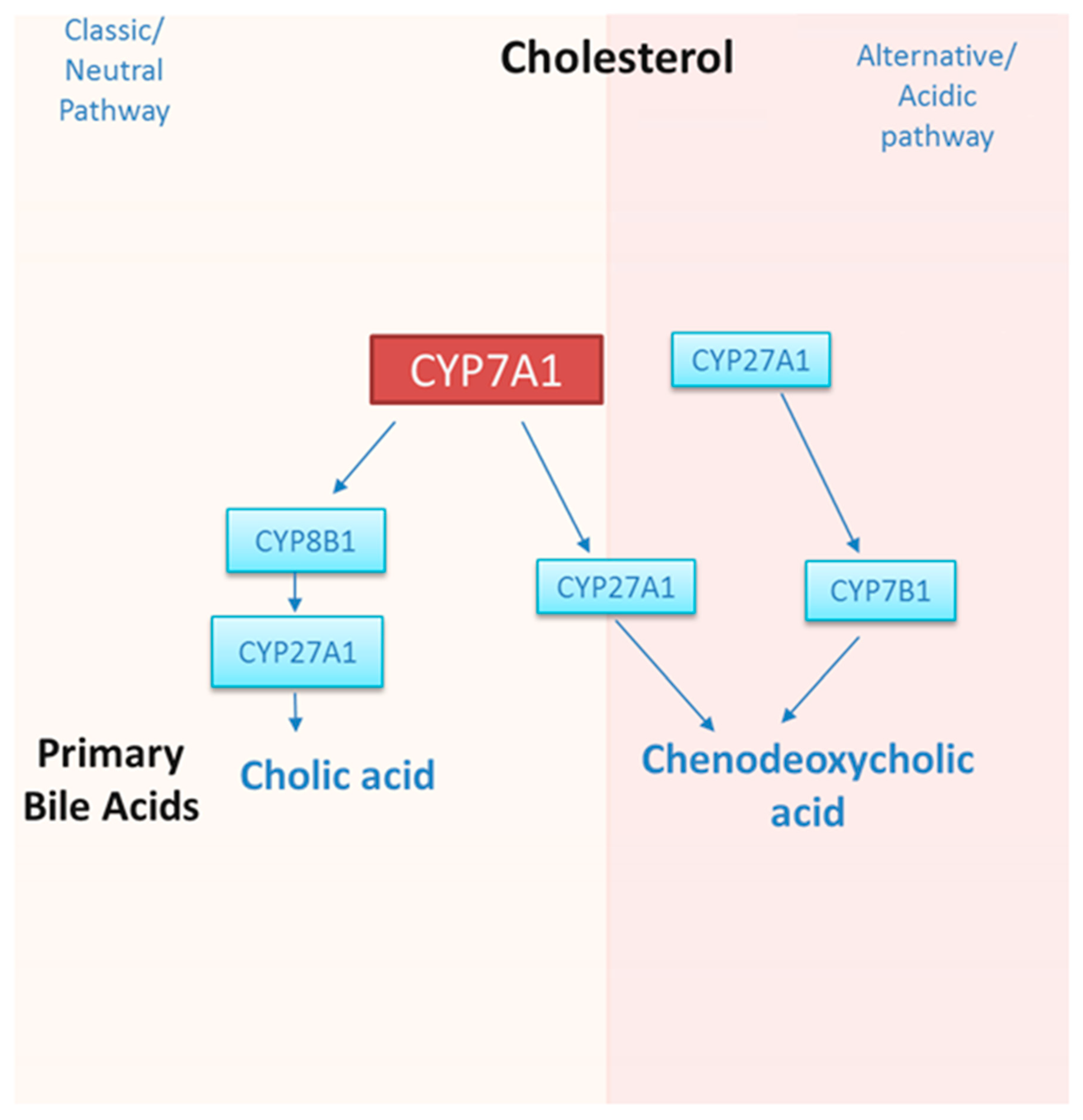
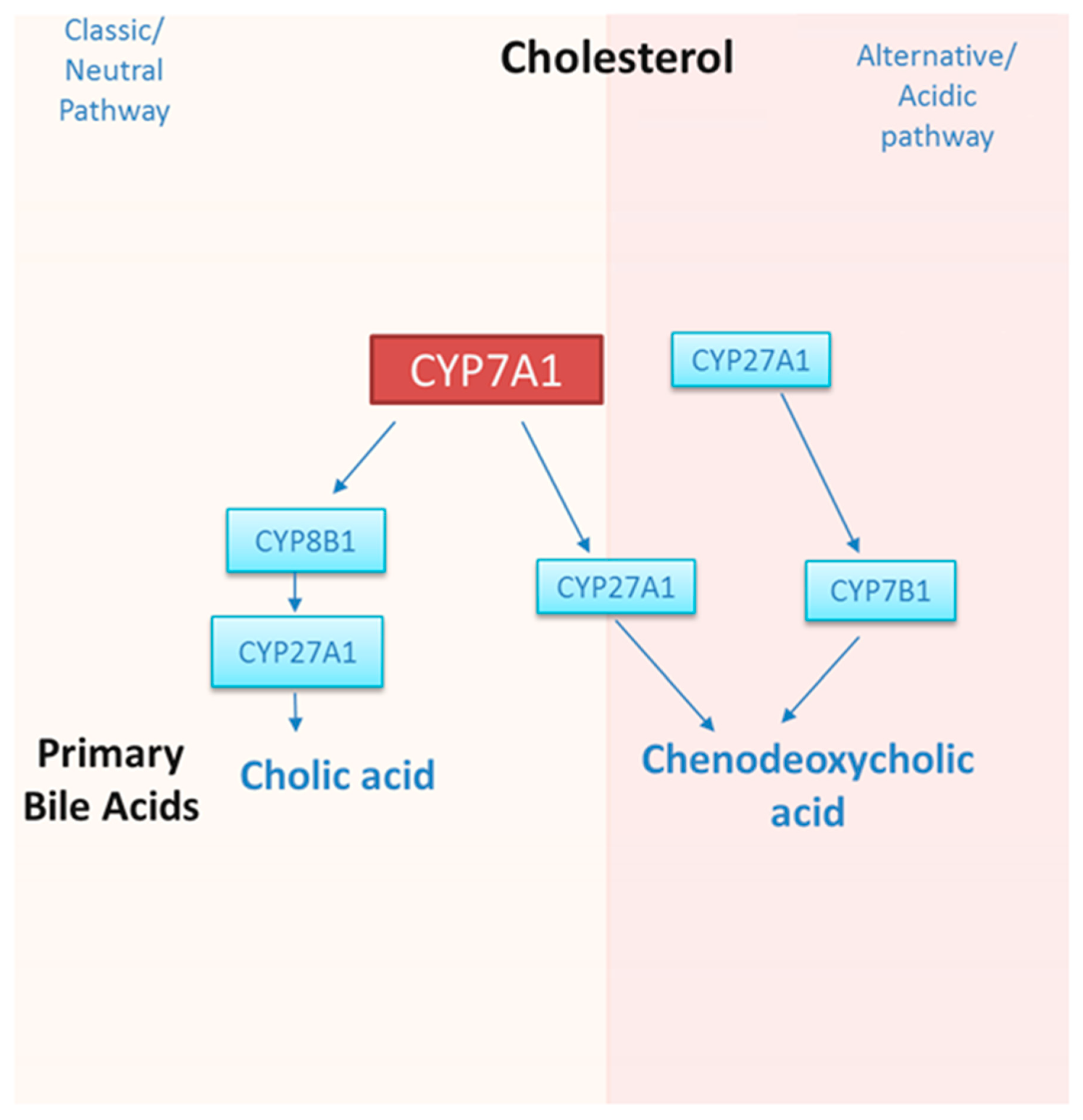
1.2. Regulation of CYP7A1 by Dietary Cholesterol, Circadian Rhythm, Transcription Factors, and microRNAs
2. Current Status of Knowledge: Polyphenols That Regulate Bile Acid Synthesis and CYP7A1
2.1. Flavonoids
2.1.1. Flavanols
2.1.2. Anthocyanins
2.1.3. Flavonols
2.1.4. Flavanones, Flavones, and Isoflavones
2.2. Stillbenes
2.3. Phenolic Acids and Phenolic Polypmers
2.4. Grape Juice, Wine and Grape Seed Extract
3. Polyphenol Mediated Mechanisms of Action
3.1. Control of the Apical Sodium Dependent Bile Acid Transporter (ASBT) by Polyphenols
3.2. Changes in Gut Microbiota Can Increase Bile Acid Excretion: Action of Polyphenols
4. Conclusions
4.1. The Limitations of Existing Models
4.2. The Most Promising Polyphenol Candidates for Future Studies
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BA | bile acid |
| Cyp7a1 | cholesterol 7 alpha-hydroxylase |
| RCT | reverse cholesterol transport |
| HDL | high-density lipoprotein |
| LDL | low-density lipoprotein |
| FXR | farnesoid X receptor |
| BSH | bile salt hydrolase |
| HF | high-fat |
| CDCA | chenodeoxycholic acid |
References
- Gistera, A.; Hansson, G.K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Russell, D.W.; Dietschy, J.M.; Turley, S.D. Marked reduction in bile acid synthesis in cholesterol 7alpha-hydroxylase-deficient mice does not lead to diminished tissue cholesterol turnover or to hypercholesterolemia. J. Lipid Res. 1998, 39, 1833–1843. [Google Scholar] [PubMed]
- Duane, W.C.; Javitt, N.B. 27-hydroxycholesterol: Production rates in normal human subjects. J. Lipid Res. 1999, 40, 1194–1199. [Google Scholar]
- De Aguiar Vallim, T.Q.; Tarling, E.J.; Edwards, P.A. Pleiotropic Roles of Bile Acids in Metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Falany, C.N.; Johnson, M.R.; Barnes, S.; Diasio, R.B. Glycine and taurine conjugation of bile acids by a single enzyme. Molecular cloning and expression of human liver bile acid CoA: Amino acid N-acyltransferase. J. Biol. Chem. 1994, 269, 19375–19379. [Google Scholar] [PubMed]
- Hayashi, H.; Takada, T.; Suzuki, H.; Onuki, R.; Hofmann, A.F.; Sugiyama, Y. Transport by vesicles of glycine- and taurine-conjugated bile salts and taurolithocholate 3-sulfate: A comparison of human BSEP with rat Bsep. Biochim. Biophys. Acta 2005, 1738, 54–62. [Google Scholar] [CrossRef]
- Dawson, P.A.; Lan, T.; Rao, A. Bile acid transporters. J. Lipid Res. 2009, 50, 2340–2357. [Google Scholar] [CrossRef] [Green Version]
- Charlton-Menys, V.; Durrington, P.N. Human cholesterol metabolism and therapeutic molecules. Exp. Physiol. 2008, 93, 27–42. [Google Scholar] [CrossRef]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef]
- Jelinek, D.F.; Andersson, S.; Slaughter, C.A.; Russell, D.W. Cloning and regulation of cholesterol 7 alpha-hydroxylase, the rate-limiting enzyme in bile acid biosynthesis. J. Biol. Chem. 1990, 265, 8190–8197. [Google Scholar] [PubMed]
- Noshiro, M.; Nishimoto, M.; Okuda, K. Rat liver cholesterol 7 alpha-hydroxylase. Pretranslational regulation for circadian rhythm. J. Biol. Chem. 1990, 265, 10036–10041. [Google Scholar] [PubMed]
- Taniguchi, T.; Chen, J.; Cooper, A.D. Regulation of cholesterol 7 alpha-hydroxylase gene expression in Hep-G2 cells. Effect of serum, bile salts, and coordinate and noncoordinate regulation with other sterol-responsive genes. J. Biol. Chem. 1994, 269, 10071–10078. [Google Scholar]
- Andreou, E.R. Analysis of CYP7A1 Gene Regulation in HepG2 Cells by Reverse-Transcriptase Polymerase Chain Reaction; University of Toronto: Toronto, ON, Canada, 1997. [Google Scholar]
- Hirsch, N.; Konstantinov, A.; Anavi, S.; Aronis, A.; Hagay, Z.; Madar, Z.; Tirosh, O. Prolonged feeding with green tea polyphenols exacerbates cholesterol-induced fatty liver disease in mice. Mol. Nutr. Food Res. 2016. [Google Scholar] [CrossRef]
- Tiemann, M.; Han, Z.; Soccio, R.; Bollineni, J.; Shefer, S.; Sehayek, E.; Breslow, J.L. Cholesterol feeding of mice expressing cholesterol 7alpha-hydroxylase increases bile acid pool size despite decreased enzyme activity. Proc. Natl. Acad. Sci. USA 2004, 101, 1846–1851. [Google Scholar] [CrossRef]
- Eggink, H.M.; Oosterman, J.E.; De Goede, P.; De Vries, E.M.; Foppen, E.; Koehorst, M.; Groen, A.K.; Boelen, A.; Romijn, J.A.; La Fleur, S.E.; et al. Complex interaction between circadian rhythm and diet on bile acid homeostasis in male rats. Chronobiol. Int. 2017, 34, 1339–1353. [Google Scholar] [CrossRef]
- Chen, P.P.; Zhang, R.Y.; Mou, L.J.; Li, X.W.; Qin, Y.; Li, X.M. An impaired hepatic clock system effects lipid metabolism in rats with nephropathy. Int. J. Mol. Med. 2018, 42, 2720–2736. [Google Scholar] [CrossRef]
- Story, J.A.; Watterson, J.J.; Matheson, H.B.; Furumoto, E.J. Dietary Fiber and Bile-Acid Metabolism. Adv. Exp. Med. Biol. 1990, 270, 43–48. [Google Scholar]
- Ide, T. Dietary Fiber-Induced Changes in Bile-Acid Conjugation and Taurine Metabolism in Rats. In Food Hydrocolloids; Springer: Boston, MA, USA, 1993; pp. 491–496. [Google Scholar]
- Locket, P.; Gallaher, D. The Effect of Fiber Source and Fat Level on Bile-Acid Metabolism in the Rat. Faseb J. 1988, 2, A862. [Google Scholar]
- Fadden, K.; Hill, M.J.; Owen, R.W. Effect of fibre on bile acid metabolism by human faecal bacteria in batch and continuous culture. Eur. J. Cancer Prev. 1997, 6, 175–194. [Google Scholar]
- Kritchevsky, D. Influence of Dietary Fiber on Bile-Acid Metabolism. Lipids 1978, 13, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Calkin, A.C.; Tontonoz, P. Transcriptional integration of metabolism by the nuclear sterol-activated receptors LXR and FXR. Nat. Rev. Mol. Cell Biol. 2012, 13, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.A.; Lee, D.I.; Kim, H.Y.; Ahn, S.H.; Seong, H.R.; Jung, W.H.; Kim, K.Y.; Kim, S.; Rhee, S.D. Cyp7a1 is continuously increased with disrupted Fxr-mediated feedback inhibition in hypercholesterolemic TALLYHO/Jng mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.; Kimmel, R.; Weinberger, C. Stroup D: Farnesoid X receptor responds to bile acids and represses cholesterol 7alpha-hydroxylase gene (CYP7A1) transcription. J. Biol. Chem. 2000, 275, 10918–10924. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chiang, J.Y. Mechanism of rifampicin and pregnane X receptor inhibition of human cholesterol 7 alpha-hydroxylase gene transcription. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G74–G84. [Google Scholar] [CrossRef]
- Goodwin, B.; Watson, M.A.; Kim, H.; Miao, J.; Kemper, J.K.; Kliewer, S.A. Differential regulation of rat and human CYP7A1 by the nuclear oxysterol receptor liver X receptor-alpha. Mol. Endocrinol. 2003, 17, 386–394. [Google Scholar] [CrossRef]
- Shin, D.J.; Campos, J.A.; Gil, G.; Osborne, T.F. PGC-1alpha activates CYP7A1 and bile acid biosynthesis. J. Biol. Chem. 2003, 278, 50047–50052. [Google Scholar] [CrossRef]
- Kir, S.; Zhang, Y.; Gerard, R.D.; Kliewer, S.A.; Mangelsdorf, D.J. Nuclear receptors HNF4alpha and LRH-1 cooperate in regulating Cyp7a1 in vivo. J. Biol. Chem. 2012, 287, 41334–41341. [Google Scholar] [CrossRef]
- Marrapodi, M.; Chiang, J.Y. Peroxisome proliferator-activated receptor alpha (PPARalpha) and agonist inhibit cholesterol 7alpha-hydroxylase gene (CYP7A1) transcription. J. Lipid Res. 2000, 41, 514–520. [Google Scholar]
- Inoue, Y.; Yu, A.M.; Yim, S.H.; Ma, X.; Krausz, K.W.; Inoue, J.; Xiang, C.C.; Brownstein, M.J.; Eggertsen, G.; Bjorkhem, I.; et al. Regulation of bile acid biosynthesis by hepatocyte nuclear factor 4alpha. J. Lipid Res. 2006, 47, 215–227. [Google Scholar] [CrossRef]
- Patel, D.D.; Knight, B.L.; Soutar, A.K.; Gibbons, G.F.; Wade, D.P. The effect of peroxisome-proliferator-activated receptor-alpha on the activity of the cholesterol 7 alpha-hydroxylase gene. Biochem. J. 2000, 351 Pt 3, 747–753. [Google Scholar] [CrossRef]
- Duan, Y.; Chen, Y.; Hu, W.; Li, X.; Yang, X.; Zhou, X.; Yin, Z.; Kong, D.; Yao, Z.; Hajjar, D.P.; et al. Peroxisome Proliferator-activated receptor gamma activation by ligands and dephosphorylation induces proprotein convertase subtilisin kexin type 9 and low density lipoprotein receptor expression. J. Biol. Chem. 2012, 287, 23667–23677. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Kong, X.; Owsley, E.; Ellis, E.; Strom, S.; Chiang, J.Y. Insulin regulation of cholesterol 7alpha-hydroxylase expression in human hepatocytes: Roles of forkhead box O1 and sterol regulatory element-binding protein 1c. J. Biol. Chem. 2006, 281, 28745–28754. [Google Scholar] [CrossRef] [PubMed]
- Park, W.H.; Pak, Y.K. Insulin-dependent suppression of cholesterol 7alpha-hydroxylase is a possible link between glucose and cholesterol metabolisms. Exp. Mol. Med. 2011, 43, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Le Martelot, G.; Claudel, T.; Gatfield, D.; Schaad, O.; Kornmann, B.; Lo Sasso, G.; Moschetta, A.; Schibler, U. REV-ERBalpha participates in circadian SREBP signaling and bile acid homeostasis. PLoS Biol. 2009, 7, e1000181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, M.; Lu, D.; Wang, S.; Yu, F.; Guo, L.; Wen, S.; Wu, B. REV-ERBalpha Regulates CYP7A1 Through Repression of Liver Receptor Homolog-1. Drug Metab. Dispos. 2018, 46, 248–258. [Google Scholar] [CrossRef]
- Lee, Y.H.; Alberta, J.A.; Gonzalez, F.J.; Waxman, D.J. Multiple, functional DBP sites on the promoter of the cholesterol 7 alpha-hydroxylase P450 gene, CYP7. Proposed role in diurnal regulation of liver gene expression. J. Biol. Chem. 1994, 269, 14681–14689. [Google Scholar]
- Ogawa, A.; Yano, M.; Tsujinaka, T.; Morimoto, T.; Morita, S.; Taniguchi, M.; Shiozaki, H.; Okamoto, K.; Sato, S.; Monden, M. Modulation of circadian expression of D-site binding protein by the schedule of parenteral nutrition in rat liver. Hepatology 1997, 26, 1580–1586. [Google Scholar] [CrossRef]
- Berard, A.M.; Dumon, M.F.; Darmon, M. Dietary fish oil up-regulates cholesterol 7alpha-hydroxylase mRNA in mouse liver leading to an increase in bile acid and cholesterol excretion. FEBS Lett. 2004, 559, 125–128. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef]
- Li, T.; Francl, J.M.; Boehme, S.; Chiang, J.Y. Regulation of cholesterol and bile acid homeostasis by the cholesterol 7alpha-hydroxylase/steroid response element-binding protein 2/microRNA-33a axis in mice. Hepatology 2013, 58, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Song, K.H.; Li, T.; Owsley, E.; Chiang, J.Y. A putative role of micro RNA in regulation of cholesterol 7alpha-hydroxylase expression in human hepatocytes. J. Lipid Res. 2010, 51, 2223–2233. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.M.; Afonso, M.B.; Simao, A.L.; Carvalho, C.C.; Trindade, A.; Duarte, A.; Borralho, P.M.; Machado, M.V.; Cortez-Pinto, H.; Rodrigues, C.M.; et al. miR-21 ablation and obeticholic acid ameliorate nonalcoholic steatohepatitis in mice. Cell Death Dis. 2017, 8, e2748. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.; Lv, X.; Liu, F. MiRNA-17 encoded by the miR-17-92 cluster increases the potential for steatosis in hepatoma cells by targeting CYP7A1. Cell Mol. Biol. Lett. 2018, 23, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, S.; Nakajima, M.; Kida, K.; Yamaura, Y.; Fukami, T.; Yokoi, T. MicroRNAs regulate human hepatocyte nuclear factor 4alpha, modulating the expression of metabolic enzymes and cell cycle. J. Biol. Chem. 2010, 285, 4415–4422. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Q.; Wang, T.; Zhao, G.; Qian, F.; Huang, J.; Wang, H.; Zhang, X.; Wang, Y. Green tea infusion protects against alcoholic liver injury by attenuating inflammation and regulating the PI3K/Akt/eNOS pathway in C57BL/6 mice. Food Funct. 2017, 8, 3165–3177. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. Polyphenols and bioavailability: An update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Vari, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.J. Antagonism by bioactive polyphenols against inflammation: A systematic view. Inflamm. Allergy Drug Targets 2014, 13, 34–64. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage. EFSA J. 2011. [Google Scholar] [CrossRef]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of Dietary Flavonoids on Reverse Cholesterol Transport, HDL Metabolism, and HDL Function. Adv. Nutr. 2017, 8, 226–239. [Google Scholar] [CrossRef]
- Yang, T.T.; Koo, M.W. Chinese green tea lowers cholesterol level through an increase in fecal lipid excretion. Life Sci. 2000, 66, 411–423. [Google Scholar] [CrossRef]
- Hirsova, P.; Karlasova, G.; Dolezelova, E.; Cermanova, J.; Zagorova, M.; Kadova, Z.; Hroch, M.; Sispera, L.; Tomsik, P.; Lenicek, M.; et al. Cholestatic effect of epigallocatechin gallate in rats is mediated via decreased expression of Mrp2. Toxicology 2013, 303, 9–15. [Google Scholar] [CrossRef]
- Shan, D.; Fang, Y.; Ye, Y.; Liu, J. EGCG reducing the susceptibility to cholesterol gallstone formation through the regulation of inflammation. Biomed. Pharmacother. 2008, 62, 677–683. [Google Scholar] [CrossRef]
- Lee, M.S.; Park, J.Y.; Freake, H.; Kwun, I.S.; Kim, Y. Green tea catechin enhances cholesterol 7alpha-hydroxylase gene expression in HepG2 cells. Br. J. Nutr. 2008, 99, 1182–1185. [Google Scholar] [CrossRef]
- Li, G.; Lin, W.; Araya, J.J.; Chen, T.; Timmermann, B.N.; Guo, G.L. A tea catechin, epigallocatechin-3-gallate, is a unique modulator of the farnesoid X receptor. Toxicol. Appl. Pharmacol. 2012, 258, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Park, E.M.; Kim, E.H.; Chung, I.M. Hypocholesterolemic metabolism of dietary red pericarp glutinous rice rich in phenolic compounds in mice fed a high cholesterol diet. Nutr. Res. Pract. 2014, 8, 632–637. [Google Scholar] [CrossRef] [Green Version]
- Matziouridou, C.; Marungruang, N.; Nguyen, T.D.; Nyman, M.; Fak, F. Lingonberries reduce atherosclerosis in Apoe(-/-) mice in association with altered gut microbiota composition and improved lipid profile. Mol. Nutr. Food Res. 2016, 60, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Maeda, T.; Sarker, M.Z.; Takigawa, S.; Matsuura-Endo, C.; Yamauchi, H.; Mukasa, Y.; Saito, K.; Hashimoto, N.; Noda, T.; et al. Identification of anthocyanins in the sprouts of buckwheat. J. Agric. Food Chem. 2007, 55, 6314–6318. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ayugase, J. Effects of buckwheat sprouts on plasma and hepatic parameters in type 2 diabetic db/db mice. J. Food Sci. 2010, 75, H294–H299. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xia, M.; Gao, S.; Li, D.; Zhang, Y.; Jin, T.; Ling, W. Cyanidin-3-O-beta-glucoside upregulates hepatic cholesterol 7alpha-hydroxylase expression and reduces hypercholesterolemia in mice. Mol. Nutr. Food Res. 2012, 56, 610–621. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, R.; Li, Y.M.; Liang, N.; Zhao, Y.; Zhu, H.; He, Z.; Liu, J.; Hao, W.; Jiao, R.; et al. Cholesterol-Lowering Activity of Tartary Buckwheat Protein. J. Agric. Food Chem. 2017, 65, 1900–1906. [Google Scholar] [CrossRef]
- Huang, J.B.; Feng, S.M.; Liu, A.N.; Dai, Z.Q.; Wang, H.; Reuhl, K.; Lu, W.Y.; Yang, C.S. Green Tea Polyphenol EGCG Alleviates Metabolic Abnormality and Fatty Liver by Decreasing Bile Acid and Lipid Absorption in Mice. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Zhang, M.; Xie, Z.; Gao, W.; Pu, L.; Wei, J.; Guo, C. Quercetin regulates hepatic cholesterol metabolism by promoting cholesterol-to-bile acid conversion and cholesterol efflux in rats. Nutr. Res. 2016, 36, 271–279. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Granados, O.; Torre-Villalvazo, I.; Serna-Saldivar, S.O.; Torres, N.; Palacios-González, B.; Tovar, A.R. Flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats modulate lipid metabolism and biliary cholesterol secretion in C57BL/6 mice. Br. J. Nutr. 2014, 112, 886–899. [Google Scholar] [CrossRef]
- Daniel, R.S.; Devi, K.S.; Augusti, K.T.; Sudhakaran Nair, C.R. Mechanism of action of antiatherogenic and related effects of Ficus bengalensis Linn. flavonoids in experimental animals. Indian J. Exp. Biol. 2003, 41, 296–303. [Google Scholar]
- Mathew, B.C.; Yoseph, B.A.; Dessale, T.; Daniel, R.S.; Alemayehu, A.; Campbell, I.W.; Augusti, K.T. Hypolipidaemic effect of Leucodelphinidin derivative from Ficus bengalensis Linn on Cholesterol fed rats. Res. J. Chem. Sci. 2012, 2, 54–60. [Google Scholar]
- Hoang, M.H.; Jia, Y.; Mok, B.; Jun, H.J.; Hwang, K.Y.; Lee, S.J. Kaempferol ameliorates symptoms of metabolic syndrome by regulating activities of liver X receptor-beta. J. Nutr. Biochem. 2015, 26, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Goldwasser, J.; Cohen, P.Y.; Yang, E.; Balaguer, P.; Yarmush, M.L.; Nahmias, Y. Transcriptional Regulation of Human and Rat Hepatic Lipid Metabolism by the Grapefruit Flavonoid Naringenin: Role of PPARα, PPARγ and LXRα. PLoS ONE 2010, 5, e12399. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Wang, C.; Peng, J.; Li, W.; Jin, Y.; Liu, Q.; Meng, Q.; Liu, K.; Sun, H. Naringin regulates cholesterol homeostasis and inhibits inflammation via modulating NF-kappaB and ERK signaling pathways in vitro. Die Pharm. 2016, 71, 101–108. [Google Scholar]
- Lin, H.R.; Chou, T.H.; Huang, D.W.; Chen, I.S. Cryptochinones from Cryptocarya chinensis act as farnesoid X receptor agonists. Bioorg. Med. Chem. Lett. 2014, 24, 4181–4186. [Google Scholar] [CrossRef] [PubMed]
- de Almeida Jackix, E.; Monteiro, E.B.; Raposo, H.F.; Vanzela, E.C.; Amaya-Farfan, J. Taioba (Xanthosoma sagittifolium) leaves: Nutrient composition and physiological effects on healthy rats. J. Food Sci. 2013, 78, H1929–H1934. [Google Scholar] [CrossRef] [PubMed]
- Picerno, P.; Mencherini, T.; Lauro, M.R.; Barbato, F.; Aquino, R. Phenolic Constituents and Antioxidant Properties of Xanthosoma violaceum Leaves. J. Agric. Food Chem. 2003, 51, 6423–6428. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Ding, J.; Zhao, H.; Liu, C.M. Puerarin attenuates carbon tetrachloride-induced liver oxidative stress and hyperlipidaemia in mouse by JNK/c-Jun/CYP7A1 pathway. Basic Clin. Pharmacol. Toxicol. 2014, 115, 389–395. [Google Scholar] [CrossRef]
- Yan, L.P.; Chan, S.W.; Chan, A.S.; Chen, S.L.; Ma, X.J.; Xu, H.X. Puerarin decreases serum total cholesterol and enhances thoracic aorta endothelial nitric oxide synthase expression in diet-induced hypercholesterolemic rats. Life Sci. 2006, 79, 324–330. [Google Scholar] [CrossRef]
- Chung, M.J.; Sung, N.-J.; Park, C.-S.; Kweon, D.-K.; Mantovani, A.; Moon, T.-W.; Lee, S.-J.; Park, K.-H. Antioxidative and hypocholesterolemic activities of water-soluble puerarin glycosides in HepG2 cells and in C57 BL/6J mice. Eur. J. Pharmacol. 2008, 578, 159–170. [Google Scholar] [CrossRef]
- Feng, D.; Sun, J.G.; Sun, R.B.; Ou-Yang, B.C.; Yao, L.; Aa, J.Y.; Zhou, F.; Zhang, J.W.; Zhang, J.; Wang, G.J. Isoflavones and phytosterols contained in Xuezhikang capsules modulate cholesterol homeostasis in high-fat diet mice. Acta Pharmacol. Sin. 2015, 36, 1462–1472. [Google Scholar] [CrossRef] [Green Version]
- Mvondo, M.A.; Njamen, D.; Kretzschmar, G.; Imma Bader, M.; Tanee Fomum, S.; Wandji, J.; Vollmer, G. Alpinumisoflavone and abyssinone V 4’-methylether derived from Erythrina lysistemon (Fabaceae) promote HDL-cholesterol synthesis and prevent cholesterol gallstone formation in ovariectomized rats. J. Pharm. pharmacol. 2015, 67, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.D.; Wei, X.J.; Zhang, C.X.; Zhong, Y.; Lu, L.Z.; Grossmann, R.; Zhao, R.Q. The effect of equol injection in ovo on lipid metabolism and hepatic lipogenic gene expression in broilers. Anim. Int. J. Anim. Biosci. 2012, 6, 1444–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Harada, T.; Takagi, N.; Tsuzuki, K.; Sugawara, M.; Fukuda, M. Effects of lactic acid-fermented soymilk on lipid metabolism-related gene expression in rat liver. Biosci. Biotechnol. Biochem. 2012, 76, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bartley, G.E.; Mitchell, C.R.; Zhang, H.; Yokoyama, W. Lower weight gain and hepatic lipid content in hamsters fed high fat diets supplemented with white rice protein, brown rice protein, soy protein, and their hydrolysates. J. Agric. Food Chem. 2011, 59, 10927–10933. [Google Scholar] [CrossRef] [PubMed]
- Laurent, T.; Okuda, Y.; Chijimatsu, T.; Umeki, M.; Kobayashi, S.; Kataoka, Y.; Tatsuguchi, I.; Mochizuki, S.; Oda, H. Freshwater Clam Extract Ameliorates Triglyceride and Cholesterol Metabolism through the Expression of Genes Involved in Hepatic Lipogenesis and Cholesterol Degradation in Rats. Evid.-Based Complement. Altern. Med. eCAM 2013, 2013, 830684. [Google Scholar] [CrossRef]
- Kroon, P.A.; Iyer, A.; Chunduri, P.; Chan, V.; Brown, L. The cardiovascular nutrapharmacology of resveratrol: Pharmacokinetics, molecular mechanisms and therapeutic potential. Curr. Med. Chem. 2010, 17, 2442–2455. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, E.; Ma, L.; Zhai, P. Dietary resveratrol increases the expression of hepatic 7alpha-hydroxylase and ameliorates hypercholesterolemia in high-fat fed C57BL/6J mice. Lipids Health Dis. 2012, 11, 56. [Google Scholar] [CrossRef]
- Shao, D.; Wang, Y.; Huang, Q.; Shi, J.; Yang, H.; Pan, Z.; Jin, M.; Zhao, H.; Xu, X. Cholesterol-Lowering Effects and Mechanisms in View of Bile Acid Pathway of Resveratrol and Resveratrol Glucuronides. J. Food Sci. 2016. [Google Scholar] [CrossRef]
- Wang, T.; Zhou, Z.X.; Sun, L.X.; Li, X.; Xu, Z.M.; Chen, M.; Zhao, G.L.; Jiang, Z.Z.; Zhang, L.Y. Resveratrol effectively attenuates alpha-naphthyl-isothiocyanate-induced acute cholestasis and liver injury through choleretic and anti-inflammatory mechanisms. Acta Pharmacol. Sin. 2014, 35, 1527–1536. [Google Scholar] [CrossRef]
- Miura, D.; Miura, Y.; Yagasaki, K. Hypolipidemic action of dietary resveratrol, a phytoalexin in grapes and red wine, in hepatoma-bearing rats. Life Sci. 2003, 73, 1393–1400. [Google Scholar] [CrossRef]
- Marier, J.F.; Vachon, P.; Gritsas, A.; Zhang, J.; Moreau, J.P.; Ducharme, M.P. Metabolism and disposition of resveratrol in rats: Extent of absorption, glucuronidation, and enterohepatic recirculation evidenced by a linked-rat model. J. Pharmacol. Exp. Ther. 2002, 302, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef]
- Kemper, J.K.; Xiao, Z.; Ponugoti, B.; Miao, J.; Fang, S.; Kanamaluru, D.; Tsang, S.; Wu, S.Y.; Chiang, C.M.; Veenstra, T.D. FXR acetylation is normally dynamically regulated by p300 and SIRT1 but constitutively elevated in metabolic disease states. Cell Metab. 2009, 10, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.D.; Zhang, Q.Y.; Mi, M.T. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. MBio 2016, 7, e02210–e02215. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yamagami, T. Antihypercholesterolemic effect of Chinese black tea extract in human subjects with borderline hypercholesterolemia. Nutr. Res. 2008, 28, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Wang, C.F.; Shen, S.M.; Wang, G.L.; Liu, P.; Liu, Z.M.; Wang, Y.Y.; Du, S.S.; Liu, Z.L.; Deng, Z.W. Antioxidant phenolic compounds from Pu-erh tea. Molecules 2012, 17, 14037–14045. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yamagami, T. Efficacy and safety of Chinese black tea (Pu-Ehr) extract in healthy and hypercholesterolemic subjects. Ann. Nutr. Metab. 2008, 53, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yamagami, T. Extract of black tea (pu-ehr) inhibits postprandial rise in serum cholesterol in mice, and with long term use reduces serum cholesterol and low density lipoprotein levels and renal fat weight in rats. Phytother. Res. PTR 2008, 22, 1275–1281. [Google Scholar] [CrossRef]
- Tan, Z.; Luo, M.; Yang, J.; Cheng, Y.; Huang, J.; Lu, C.; Song, D.; Ye, M.; Dai, M.; Gonzalez, F.J.; et al. Chlorogenic acid inhibits cholestatic liver injury induced by alpha-naphthylisothiocyanate: Involvement of STAT3 and NFkappaB signalling regulation. J. Pharm. Pharmacol. 2016, 68, 1203–1213. [Google Scholar] [CrossRef]
- Spiller, G.A.; Story, J.A.; Furumoto, E.J.; Chezem, J.C.; Spiller, M. Effect of tartaric acid and dietary fibre from sun-dried raisins on colonic function and on bile acid and volatile fatty acid excretion in healthy adults. Br. J. Nutr. 2003, 90, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Spiller, G.A.; Story, J.A.; Lodics, T.A.; Pollack, M.; Monyan, S.; Butterfield, G.; Spiller, M. Effect of sun-dried raisins on bile acid excretion, intestinal transit time, and fecal weight: A dose-response study. J. Med. Food 2003, 6, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Carughi, A. Polyphenol content and health benefits of raisins. Nutr. Res. 2010, 30, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Tebib, K.; Besancon, P.; Rouanet, J.M. Dietary grape seed tannins affect lipoproteins, lipoprotein lipases and tissue lipids in rats fed hypercholesterolemic diets. J. Nutr. 1994, 124, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Camire, M.E.; Dougherty, M.P. Raisin dietary fiber composition and in vitro bile acid binding. J. Agric. Food Chem. 2003, 51, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Ngamukote, S.; Makynen, K.; Thilawech, T.; Adisakwattana, S. Cholesterol-lowering activity of the major polyphenols in grape seed. Molecules 2011, 16, 5054–5061. [Google Scholar] [CrossRef]
- Yao, R.; Yasuoka, A.; Kamei, A.; Ushiama, S.; Kitagawa, Y.; Rogi, T.; Shibata, H.; Abe, K.; Misaka, T. Nuclear receptor-mediated alleviation of alcoholic fatty liver by polyphenols contained in alcoholic beverages. PLoS ONE 2014, 9, e87142. [Google Scholar] [CrossRef]
- Jiao, R.; Zhang, Z.; Yu, H.; Huang, Y.; Chen, Z.Y. Hypocholesterolemic activity of grape seed proanthocyanidin is mediated by enhancement of bile acid excretion and up-regulation of CYP7A1. J. Nutr. Biochem. 2010, 21, 1134–1139. [Google Scholar] [CrossRef]
- Downing, L.E.; Edgar, D.; Ellison, P.A.; Ricketts, M.L. Mechanistic insight into nuclear receptor-mediated regulation of bile acid metabolism and lipid homeostasis by grape seed procyanidin extract (GSPE). Cell Biochem. Funct. 2017, 35, 12–32. [Google Scholar] [CrossRef]
- Del Bas, J.M.; Fernandez-Larrea, J.; Blay, M.; Ardevol, A.; Salvado, M.J.; Arola, L.; Blade, C. Grape seed procyanidins improve atherosclerotic risk index and induce liver CYP7A1 and SHP expression in healthy rats. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2005, 19, 479–481. [Google Scholar] [CrossRef]
- Sembries, S.; Dongowski, G.; Mehrlander, K.; Will, F.; Dietrich, H. Physiological effects of extraction juices from apple, grape, and red beet pomaces in rats. J. Agric. Food Chem. 2006, 54, 10269–10280. [Google Scholar] [CrossRef]
- Heidker, R.M.; Caiozzi, G.C.; Ricketts, M.L. Dietary procyanidins selectively modulate intestinal farnesoid X receptor-regulated gene expression to alter enterohepatic bile acid recirculation: Elucidation of a novel mechanism to reduce triglyceridemia. Mol. Nutr. Food Res. 2016, 60, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.J.; Cho, Y.; Moon, J.; Jeon, H.J.; Lee, S.M.; Chung, J.H. Hypocholesterolemic effect of daily fisetin supplementation in high fat fed Sprague-Dawley rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 57, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Del Bas, J.M.; Crescenti, A.; Arola-Arnal, A.; Oms-Oliu, G.; Arola, L.; Caimari, A. Intake of grape procyanidins during gestation and lactation impairs reverse cholesterol transport and increases atherogenic risk indexes in adult offspring. J. Nutr. Biochem. 2015, 26, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Oike, H.; Kobori, M. Resveratrol regulates circadian clock genes in Rat-1 fibroblast cells. Biosci. Biotechnol. Biochem. 2008, 72, 3038–3040. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Portillo, M.P.; Madrid, J.A.; Arias, N.; Macarulla, M.T.; Garaulet, M. Effects of resveratrol on changes induced by high-fat feeding on clock genes in rats. Br. J. Nutr. 2013, 110, 1421–1428. [Google Scholar] [CrossRef] [Green Version]
- Baselga-Escudero, L.; Blade, C.; Ribas-Latre, A.; Casanova, E.; Salvado, M.J.; Arola, L.; Arola-Arnal, A. Grape seed proanthocyanidins repress the hepatic lipid regulators miR-33 and miR-122 in rats. Mol. Nutr. Food Res. 2012, 56, 1636–1646. [Google Scholar] [CrossRef]
- Baselga-Escudero, L.; Blade, C.; Ribas-Latre, A.; Casanova, E.; Suarez, M.; Torres, J.L.; Salvado, M.J.; Arola, L.; Arola-Arnal, A. Resveratrol and EGCG bind directly and distinctively to miR-33a and miR-122 and modulate divergently their levels in hepatic cells. Nucleic Acids Res. 2014, 42, 882–892. [Google Scholar] [CrossRef]
- Jahanafrooz, Z.; Motamed, N.; Bakhshandeh, B. Effects of miR-21 downregulation and silibinin treatment in breast cancer cell lines. Cytotechnology 2017. [Google Scholar] [CrossRef]
- Su, X.; Zhang, J.; Wang, H.; Xu, J.; He, J.; Liu, L.; Zhang, T.; Chen, R.; Kang, J. Phenolic Acid Profiling, Antioxidant, and Anti-Inflammatory Activities, and miRNA Regulation in the Polyphenols of 16 Blueberry Samples from China. Molecules 2017, 22, 312. [Google Scholar] [CrossRef]
- Milenkovic, D.; Deval, C.; Gouranton, E.; Landrier, J.F.; Scalbert, A.; Morand, C.; Mazur, A. Modulation of miRNA expression by dietary polyphenols in apoE deficient mice: A new mechanism of the action of polyphenols. PLoS ONE 2012, 7, e29837. [Google Scholar] [CrossRef]
- Shimoyama, M.D.P.J.; Hayman, G.T.; Laulederkind, S.J.; Liu, W.; Nigam, R.; Petri, V.; Smith, J.R.; Tutaj, M.; Wang, S.J.; Worthey, E.; et al. The Rat Genome Database 2015: Genomic, phenotypic and environmental variations and disease. Nucleic Acids Res. 2014, 43, D743–D750. [Google Scholar] [CrossRef] [PubMed]
- Arola-Arnal, A.; Blade, C. Proanthocyanidins modulate microRNA expression in human HepG2 cells. PLoS ONE 2011, 6, e25982. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Kosters, A.; Mells, J.E.; Zhang, W.; Setchell, K.D.; Amanso, A.M.; Wynn, G.M.; Xu, T.; Keller, B.T.; Yin, H.; et al. Inhibition of ileal bile acid uptake protects against nonalcoholic fatty liver disease in high-fat diet-fed mice. Sci. Transl. Med. 2016, 8, 357ra122. [Google Scholar] [CrossRef] [PubMed]
- Bhat, B.G.; Rapp, S.R.; Beaudry, J.A.; Napawan, N.; Butteiger, D.N.; Hall, K.A.; Null, C.L.; Luo, Y.; Keller, B.T. Inhibition of ileal bile acid transport and reduced atherosclerosis in apoE-/-mice by SC-435. J. Lipid Res. 2003, 44, 1614–1621. [Google Scholar] [CrossRef]
- Huff, M.W.; Telford, D.E.; Edwards, J.Y.; Burnett, J.R.; Barrett, P.H.R.; Rapp, S.R.; Napawan, N.; Keller, B.T. Inhibition of the Apical Sodium-Dependent Bile Acid Transporter Reduces LDL Cholesterol and ApoB by Enhanced Plasma Clearance of LDL ApoB. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1884–1891. [Google Scholar] [CrossRef]
- Thomas, C.; Landrier, J.F.; Gaillard, D.; Grober, J.; Monnot, M.C.; Athias, A.; Besnard, P. Cholesterol dependent downregulation of mouse and human apical sodium dependent bile acid transporter (ASBT) gene expression: Molecular mechanism and physiological consequences. Gut 2006, 55, 1321–1331. [Google Scholar] [CrossRef]
- Alrefai, W.A.; Sarwar, Z.; Tyagi, S.; Saksena, S.; Dudeja, P.K.; Gill, R.K. Cholesterol modulates human intestinal sodium-dependent bile acid transporter. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G978–G985. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Shneider, B.L.; Shefer, S.; Nguyen, L.B.; Batta, A.K.; Tint, G.S.; Arrese, M.; Thevananther, S.; Ma, L.; Stengelin, S.; et al. Ileal bile acid transport regulates bile acid pool, synthesis, and plasma cholesterol levels differently in cholesterol-fed rats and rabbits. J. Lipid Res. 2000, 41, 298–304. [Google Scholar]
- Torchia, E.C.; Cheema, S.K.; Agellon, L.B. Coordinate regulation of bile acid biosynthetic and recovery pathways. Biochem. Biophys. Res. Commun. 1996, 225, 128–133. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Guo, G.L.; Klaassen, C.D. Diurnal variations of mouse plasma and hepatic bile acid concentrations as well as expression of biosynthetic enzymes and transporters. PLoS ONE 2011, 6, e16683. [Google Scholar] [CrossRef]
- Odermatt, A.; Da Cunha, T.; Penno, C.A.; Chandsawangbhuwana, C.; Reichert, C.; Wolf, A.; Dong, M.; Baker, M.E. Hepatic reduction of the secondary bile acid 7-oxolithocholic acid is mediated by 11beta-hydroxysteroid dehydrogenase 1. Biochem. J. 2011, 436, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.M.; Marchesi, J.R. Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 13580–13585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayin, S.I.; Wahlstrom, A.; Felin, J.; Jantti, S.; Marschall, H.U.; Bamberg, K.; Angelin, B.; Hyotylainen, T.; Oresic, M.; Backhed, F. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Wostmann, B.S. Intestinal bile acids and cholesterol absorption in the germfree rat. J. Nutr. 1973, 103, 982–990. [Google Scholar] [CrossRef]
- Kellogg, T.F.; Wostmann, B.S. Fecal neutral steroids and bile acids from germfree rats. J. Lipid Res. 1969, 10, 495–503. [Google Scholar]
- Madsen, D.; Beaver, M.; Chang, L.; Bruckner-Kardoss, E.; Wostmann, B. Analysis of bile acids in conventional and germfree rats. J. Lipid Res. 1976, 17, 107–111. [Google Scholar]
- Li, F.; Jiang, C.; Krausz, K.W.; Li, Y.; Albert, I.; Hao, H.; Fabre, K.M.; Mitchell, J.B.; Patterson, A.D.; Gonzalez, F.J. Microbiome remodelling leads to inhibition of intestinal farnesoid X receptor signalling and decreased obesity. Nat. Commun. 2013, 4, 2384. [Google Scholar] [CrossRef]
- Jeun, J.; Kim, S.; Cho, S.Y.; Jun, H.J.; Park, H.J.; Seo, J.G.; Chung, M.J.; Lee, S.J. Hypocholesterolemic effects of Lactobacillus plantarum KCTC3928 by increased bile acid excretion in C57BL/6 mice. Nutrition 2010, 26, 321–330. [Google Scholar] [CrossRef]
- Jiang, C.; Xie, C.; Lv, Y.; Li, J.; Krausz, K.W.; Shi, J.; Brocker, C.N.; Desai, D.; Amin, S.G.; Bisson, W.H.; et al. Intestine-selective farnesoid X receptor inhibition improves obesity-related metabolic dysfunction. Nat. Commun. 2015, 6, 10166. [Google Scholar] [CrossRef]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br. J. Nutr. 2012, 107, 1505–1513. [Google Scholar] [CrossRef]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Cholesterol lowering and inhibition of sterol absorption by Lactobacillus reuteri NCIMB 30242: A randomized controlled trial. Eur. J. Clin. Nutr. 2012, 66, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; De Boever, P.; Verstraete, W. Cholesterol lowering in pigs through enhanced bacterial bile salt hydrolase activity. Br. J. Nutr. 1998, 79, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaihara, A.; Yoshii, K.; Tsumura, Y.; Ishimitsu, S.; Tonogai, Y. Effects of the Oral Administration of Green Tea Polyphenol and Tannic Acid on Serum and Hepatic Lipid Contents and Fecal Steroid Excretion in Rats. J. Health Sci. 2001, 47, 107–117. [Google Scholar] [CrossRef]
- Han, Y.; Haraguchi, T.; Iwanaga, S.; Tomotake, H.; Okazaki, Y.; Mineo, S.; Moriyama, A.; Inoue, J.; Kato, N. Consumption of some polyphenols reduces fecal deoxycholic acid and lithocholic acid, the secondary bile acids of risk factors of colon cancer. J. Agric. Food Chem. 2009, 57, 8587–8590. [Google Scholar] [CrossRef]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef]
- Queipo-Ortuno, M.I.; Boto-Ordonez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andres-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. Fems Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef]
- Dolara, P.; Luceri, C.; Filippo, C.D.; Femia, A.P.; Giovannelli, L.; Caderni, G.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Cresci, A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 591, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Caderni, G.; Remy, S.; Cheynier, V.; Morozzi, G.; Dolara, P. Effect of complex polyphenols on colon carcinogenesis. Eur. J. Nutr. 1999, 38, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-week consumption of a wild blueberry powder drink increases bifidobacteria in the human gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, J.; Tokutake, S.; Kikuchi, M.; Kubota, Y.; Konishi, H.; Mitsuoka, T. Effect of Proanthocyanidin-Rich Extract from Grape Seeds on Human Fecal Flora and Fecal Odor. Microb. Ecol. Health Dis. 2001, 13, 25–31. [Google Scholar] [CrossRef]
- Larrosa, M.; Yanez-Gascon, M.J.; Selma, M.V.; Gonzalez-Sarrias, A.; Toti, S.; Ceron, J.J.; Tomas-Barberan, F.; Dolara, P.; Espin, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef]
- Le Roy, C.I.; Wells, P.M.; Si, J.; Raes, J.; Bell, J.T.; Spector, T.D. Red Wine Consumption Associated With Increased Gut Microbiota alpha-diversity in 3 Independent Cohorts. Gastroenterology 2019. [Google Scholar] [CrossRef]
- Fotschki, B.; Juskiewicz, J.; Jurgonski, A.; Rigby, N.; Sojka, M.; Kolodziejczyk, K.; Mackie, A.; Zdunczyk, Z. Raspberry pomace alters cecal microbial activity and reduces secondary bile acids in rats fed a high-fat diet. J. Nutr. Biochem. 2017, 46, 13–20. [Google Scholar] [CrossRef]
- Kim, H.; Kim, D.H.; Seo, K.H.; Chon, J.W.; Nah, S.Y.; Bartley, G.E.; Arvik, T.; Lipson, R.; Yokoyama, W. Modulation of the intestinal microbiota is associated with lower plasma cholesterol and weight gain in hamsters fed chardonnay grape seed flour. J. Agric. Food Chem. 2015, 63, 1460–1467. [Google Scholar] [CrossRef]
- Ren, K.; Jiang, T.; Zhao, G.J. Quercetin induces the selective uptake of HDL-cholesterol via promoting SR-BI expression and the activation of the PPARgamma/LXRalpha pathway. Food Funct. 2018, 9, 624–635. [Google Scholar] [CrossRef]
- Hou, D.X.; Luo, D.; Tanigawa, S.; Hashimoto, F.; Uto, T.; Masuzaki, S.; Fujii, M.; Sakata, Y. Prodelphinidin B-4 3′-O-gallate, a tea polyphenol, is involved in the inhibition of COX-2 and iNOS via the downregulation of TAK1-NIF-kappa B pathway. Biochem. Pharmacol. 2007, 74, 742–751. [Google Scholar] [CrossRef]
- Crestani, M.; Galli, G.; Chiang, J.Y. Genomic cloning, sequencing, and analysis of the hamster cholesterol 7 alpha-hydroxylase gene (CYP7). Arch. Biochem. Biophys. 1993, 306, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.I.; Karaoglu, D.; Haro, D.; Barillas, C.; Bashirzadeh, R.; Gil, G. Cholesterol and bile acids regulate cholesterol 7 alpha-hydroxylase expression at the transcriptional level in culture and in transgenic mice. Mol. Cell. Biol. 1994, 14, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Gatmaitan, Z.; Jefferson, D.M.; Ruiz-Opazo, N.; Biempica, L.; Arias, I.M.; Dudas, G.; Leinwand, L.A.; Reid, L.M. Regulation of growth and differentiation of a rat hepatoma cell line by the synergistic interactions of hormones and collagenous substrata. J. Cell Biol. 1983, 97, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Enat, R.; Jefferson, D.M.; Ruiz-Opazo, N.; Gatmaitan, Z.; Leinwand, L.A.; Reid, L.M. Hepatocyte proliferation in vitro: Its dependence on the use of serum-free hormonally defined medium and substrata of extracellular matrix. Proc. Natl. Acad. Sci. USA 1984, 81, 1411–1415. [Google Scholar] [CrossRef]
- Davalos, A.; Fernandez-Hernando, C.; Cerrato, F.; Martinez-Botas, J.; Gomez-Coronado, D.; Gomez-Cordoves, C.; Lasuncion, M.A. Red grape juice polyphenols alter cholesterol homeostasis and increase LDL-receptor activity in human cells in vitro. J. Nutr. 2006, 136, 1766–1773. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
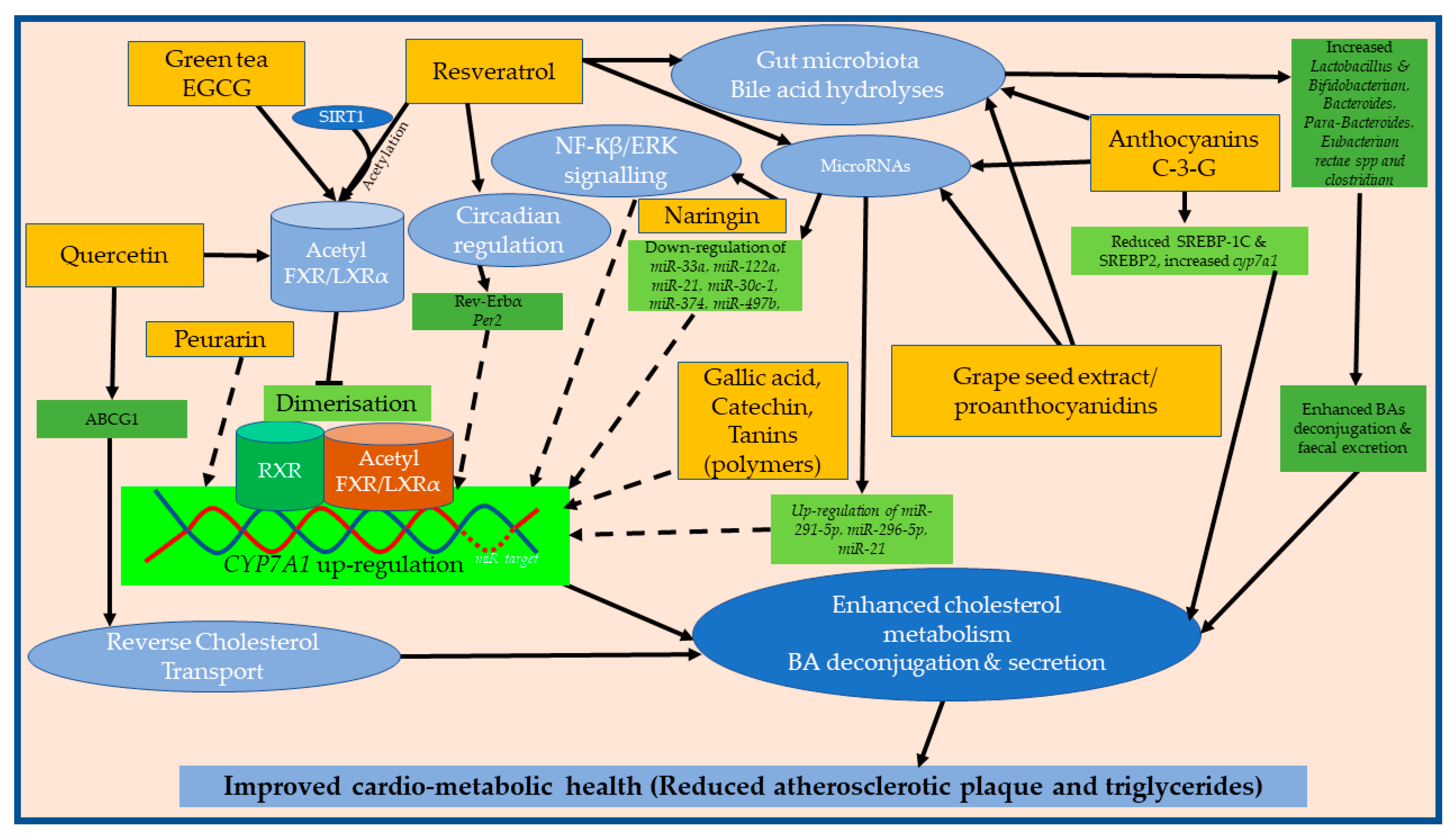{kind=link}
| Factor | Humans/Human Cell Lines | Rodents | Ref. |
|---|---|---|---|
| Farnesoid X receptor, retinoic acid receptor and small heterodimer partner (FXR-RXR and SHP) | FXR is highly expressed in both the liver and ileum tissue. Ligand binding to FXR allows translocation from the cytoplasm to the nucleus to bind RXR at FXR-response elements. FXR is not able to bind to CYP7A1 but can bind to the promoter of the SHP gene. SHP represses CYP7A1 gene expression by binding to human α-fetoprotein transcription factor (FTF). FXR-RXR complex also binds illeal FGF19 (humans) or FGFR4 (mice) translocates to the liver, activates FGR4 which inhibits c CYP7A1 transcription. | The same as humans, however, FTF is called liver receptor homolog-1 (LRH-1) in mice. | [24,25,26] |
| Pregnane X receptor (PXR) | PXR activation by specific bile acids such as lithocholic acid (LCA) leads to the repression of bile acid synthesis by binding and inactivating the transcription factor, hepatocyte nuclear factor 4 alpha (HNF4α) so that it can no-longer bind to its transcriptional co-activator, proliferator-activated receptor γ co-activator 1-α (PGC1α) to induce CYP7A1 transcription. | [27] | |
| Liver X receptor α (LXRα) | LXRα cannot bind to the human CYP7A1 promoter due to an alteration of the DR4 motif in the BARE-I sequence. Therefore, in humans LXRα does not play a role in the regulation of CYP7A1 gene expression. | Unlike in humans, LXRα can directly bind to the Cyp7a1 promoter to upregulate expression. | [28] |
| Hepatocyte nuclear factor 4 alpha/Peroxisome proliferator-activated receptor γ co-activator 1-α (HNF4α /PGC1α) | HNF4α is a transcription factor that upregulates CYP7A1 by directly binding to its promoter along with the trans-activator PGC-1α. | Same as humans. | [29,30,31,32] |
| Peroxisome Proliferator Activated Receptor Alpha (PPARα) | In vitro PPARα over-expression in human liver cells has been shown to reduce CYP7A1gene expression. However, when activators of PPARα where added to non-over-expressing cells a moderate amount of inhibition was observed. | PPARα knock out mice did not show altered Cyp7a1 levels. | [31,33] |
| Peroxisome Proliferator Activated Receptor gamma (PPARγ) | PPARγ activation induced CYP7A1 expression in HepG2 cells | [34] | |
| Forkhead box protein O1 (FoxO1) | FoxO1 is an in-direct suppressor of CYP7A1, there is no binding site for FoxO1 on the human CYP7A11 promoter. FoxO1 inhibits CYP7A1 by inhibiting expression of HNF-4α and PGC-1α TFs. | FoxO1 has the opposite function in mice and upregulates Cyp7a1 directly by binding to its promoter. | [35,36] |
| Nuclear receptor subfamily 1, group D, member 1 (NR1D1 or Rev-Erba) | NR | Competes for the promoter of the clock gene Bmal1 and mediate the circadian regulation of Cyp7a1 | [37,38]. |
| Per1 and Per2 | NR | Genetic ablation in mice disrupts normal BA control, increases serum BA and in parallel reduces dbp and Cyp7a1 expression in both rats and mice | [39,40]. |
| D site albumin promoter binding protein (DBP) | Gain-of-function studies have shown that DBP serves as a circadian activator of CYP7A1 transcription. | Cyp7a1 peaks after dark | [37,41,42] |
| Enhancer binding protein C/EBPβ-LAP | Binds cyp7a1 promoter site at DBP | [39] | |
| miR-33 | Located in the intron sequence of SREBP and regulates Cyp7a1 expression possibly synergistically to control hepatic cholesterol metabolism and BA synthesis | [43,44,45,46] | |
| miR-144-3p and miR-99a-3p | Target CYP7A1 and other non-alcoholic fatty liver disease related genes | [43] | |
| miR-122 and miR-422 | Cyp7a1 also has recognition sequences for miR-122 and miR-422 in its 3′-UTR. A synthetic miR-122 mimic inhibits CYP7A1expression in vitro and miR-122 inhibition has shown the opposite effect | [44] | |
| miR-24 and miR-34 | Indirectly decrease CYP7A1 by decreasing HNF4α transcription factor | [47] | |
| miR-17 | Leads a reduction in CYP7A1 mRNA expression | Reduces Cyp7a1 mRNA expression accompanied by hepatic steatosis | [30]. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients 2019, 11, 2588. https://doi.org/10.3390/nu11112588
Chambers KF, Day PE, Aboufarrag HT, Kroon PA. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients. 2019; 11(11):2588. https://doi.org/10.3390/nu11112588
Chicago/Turabian StyleChambers, Karen F., Priscilla E. Day, Hassan T. Aboufarrag, and Paul A. Kroon. 2019. "Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review" Nutrients 11, no. 11: 2588. https://doi.org/10.3390/nu11112588
APA StyleChambers, K. F., Day, P. E., Aboufarrag, H. T., & Kroon, P. A. (2019). Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients, 11(11), 2588. https://doi.org/10.3390/nu11112588