Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors



Abstract
1. Introduction
2. Fat Intake and Metabolic Endotoxemia
2.1. Dysbiosis
2.2. Mechanisms of the Influx of LPS into the Bloodstream
2.3. Kinetics and Activity of LPS
3. Dietary Factors that Decrease Blood LPS Levels
3.1. Probiotics
3.2. Prebiotics
3.3. Polyphenols
3.4. Sulfated Polysaccharide
3.5. Other Dietary Components/Extracts/Foods
3.6. Chinese Medicines
3.7. Dietary Habits
4. Association of Dietary Factor-Induced Reduction of Blood LPS and Modulation of Gut Microbiota
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kiers, D.; Leijte, G.P.; Gerretsen, J.; Zwaag, J.; Kox, M.; Pickkers, P. Comparison of different lots of endotoxin and evaluation of in vivo potency over time in the experimental human endotoxemia model. Innate Immun. 2019, 25, 34–45. [Google Scholar] [CrossRef]
- Benson, S.; Engler, H.; Wegner, A.; Rebernik, L.; Spreitzer, I.; Schedlowski, M.; Elsenbruch, S. What makes you feel sick after inflammation? Predictors of acute and persisting physical sickness symptoms induced by experimental endotoxemia. Clin. Pharmacol. Ther. 2017, 102, 141–151. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Liang, H.; Hussey, S.E.; Sanchez-Avila, A.; Tantiwong, P.; Musi, N. Effect of lipopolysaccharide on inflammation and insulin action in human muscle. PLoS ONE 2013, 8, e63983. [Google Scholar] [CrossRef]
- Pussinen, P.J.; Havulinna, A.S.; Lehto, M.; Sundvall, J.; Salomaa, V. Endotoxemia is associated with an increased risk of incident diabetes. Diabetes Care 2011, 34, 392–397. [Google Scholar] [CrossRef]
- Jin, R.; Willment, A.; Patel, S.S.; Sun, X.; Song, M.; Mannery, Y.O.; Kosters, A.; McClain, C.J.; Vos, M.B. Fructose induced endotoxemia in pediatric nonalcoholic fatty liver disease. Int. J. Hepatol. 2014, 2014, 560620. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Madhulika, A.; Deepika, G.; Rao, G.V.; Reddy, D.N.; Subramanyam, C.; Sasikala, M.; Talukdar, R. Altered intestinal microbiota in patients with chronic pancreatitis: Implications in diabetes and metabolic abnormalities. Sci. Rep. 2017, 7, 43640. [Google Scholar] [CrossRef]
- Zhang, R.; Miller, R.G.; Gascon, R.; Champion, S.; Katz, J.; Lancero, M.; Narvaez, A.; Honrada, R.; Ruvalcaba, D.; McGrath, M.S. Circulating endotoxin and systemic immune activation in sporadic amyotrophic lateral sclerosis (sALS). J. Neuroimmunol. 2009, 206, 121–124. [Google Scholar] [CrossRef]
- Kaliannan, K.; Hamarneh, S.R.; Economopoulos, K.P.; Nasrin Alam, S.; Moaven, O.; Patel, P.; Malo, N.S.; Ray, M.; Abtahi, S.M.; Muhammad, N.; et al. Intestinal alkaline phosphatase prevents metabolic syndrome in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 7003–7008. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Candido, T.L.N.; Bressan, J.; Alfenas, R.d.C.G. Dysbiosis and metabolic endotoxemia induced by high-fat diet. Nutr. Hosp. 2018, 35, 1432–1440. [Google Scholar]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.-L.; Rainteau, D.; Bridonneau, C.; da Costa, G.; van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Gehring, U.; Spithoven, J.; Schmid, S.; Bitter, S.; Braun-Fahrländer, C.; Dalphin, J.-C.; Hyvärinen, A.; Pekkanen, J.; Riedler, J.; Weiland, S.K.; et al. Endotoxin levels in cow’s milk samples from farming and non-farming families - the PASTURE study. Environ. Int. 2008, 34, 1132–1136. [Google Scholar] [CrossRef]
- Lindenberg, F.C.B.; Ellekilde, M.; Thörn, A.C.; Kihl, P.; Larsen, C.S.; Hansen, C.H.F.; Metzdorff, S.B.; Aalbæk, B.; Hansen, A.K. Dietary LPS traces influences disease expression of the diet-induced obese mouse. Res. Vet. Sci. 2019, 123, 195–203. [Google Scholar] [CrossRef]
- Gutsmann, T.; Howe, J.; Zähringer, U.; Garidel, P.; Schromm, A.B.; Koch, M.H.J.; Fujimoto, Y.; Fukase, K.; Moriyon, I.; Martínez-de-Tejada, G.; et al. Structural prerequisites for endotoxic activity in the limulus test as compared to cytokine production in mononuclear cells. Innate Immun. 2010, 16, 39–47. [Google Scholar] [CrossRef]
- Bishop, R.E. Structural biology of membrane-intrinsic beta-barrel enzymes: Sentinels of the bacterial outer membrane. Biochim. Biophys. Acta 2008, 1778, 1881–1896. [Google Scholar] [CrossRef]
- Van Den Berg, C.; de Neeling, A.J.; Schot, C.S.; Hustinx, W.N.M.; Wemer, J.; de Wildt, D.J. Delayed antibiotic-induced lysis of escherichia coli in vitro is correlated with enhancement of LPS release. Scand. J. Infect. Dis. 1992, 24, 619–627. [Google Scholar] [CrossRef]
- Jin, Y.; Wu, Y.; Zeng, Z.; Jin, C.; Wu, S.; Wang, Y.; Fu, Z. From the cover: Exposure to oral antibiotics induces gut microbiota dysbiosis associated with lipid metabolism dysfunction and low-grade inflammation in mice. Toxicol. Sci. 2016, 154, 140–152. [Google Scholar] [CrossRef]
- Radilla-Vázquez, R.B.; Parra-Rojas, I.; Martínez-Hernández, N.E.; Márquez-Sandoval, Y.F.; Illades-Aguiar, B.; Castro-Alarcón, N. Gut microbiota and metabolic endotoxemia in young obese mexican subjects. Obes. Facts 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Laugerette, F.; Vors, C.; Géloën, A.; Chauvin, M.; Soulage, C.; Lambert-Porcheron, S.; Peretti, N.; Alligier, M.; Burcelin, R.; Laville, M.; et al. Emulsified lipids increase endotoxemia: Possible role in early postprandial low-grade inflammation. J. Nutr. Biochem. 2011, 22, 53–59. [Google Scholar] [CrossRef]
- Clemente-Postigo, M.; Queipo-Ortuño, M.I.; Murri, M.; Boto-Ordoñez, M.; Perez-Martinez, P.; Andres-Lacueva, C.; Cardona, F.; Tinahones, F.J. Endotoxin increase after fat overload is related to postprandial hypertriglyceridemia in morbidly obese patients. J. Lipid Res. 2012, 53, 973–978. [Google Scholar] [CrossRef]
- Murakami, Y.; Tanabe, S.; Suzuki, T. High-fat diet-induced intestinal hyperpermeability is associated with increased bile acids in the large intestine of mice. J. Food Sci. 2016, 81, H216–H222. [Google Scholar] [CrossRef]
- Ahmad, R.; Rah, B.; Bastola, D.; Dhawan, P.; Singh, A.B. Obesity-induces organ and tissue specific tight junction restructuring and barrier deregulation by claudin switching. Sci. Rep. 2017, 7, 5125. [Google Scholar] [CrossRef]
- Guo, S.; Nighot, M.; Al-Sadi, R.; Alhmoud, T.; Nighot, P.; Ma, T.Y. Lipopolysaccharide regulation of intestinal tight junction permeability is mediated by TLR4 signal transduction pathway activation of FAK and MyD88. J. Immunol. 2015, 195, 4999–5010. [Google Scholar] [CrossRef]
- Fang, W.; Xue, H.; Chen, X.; Chen, K.; Ling, W. Supplementation with sodium butyrate modulates the composition of the gut microbiota and ameliorates high-fat diet-induced obesity in mice. J. Nutr. 2019, 149, 747–754. [Google Scholar] [CrossRef]
- Blanchard, C.; Moreau, F.; Chevalier, J.; Ayer, A.; Garcon, D.; Arnaud, L.; Pais de Barros, J.-P.; Gautier, T.; Neunlist, M.; Cariou, B.; et al. Sleeve gastrectomy alters intestinal permeability in diet-induced obese mice. Obes. Surg. 2017, 27, 2590–2598. [Google Scholar] [CrossRef]
- González-Quilen, C.; Gil-Cardoso, K.; Ginés, I.; Beltrán-Debón, R.; Pinent, M.; Ardévol, A.; Terra, X.; Blay, M.T. Grape-Seed proanthocyanidins are able to reverse intestinal dysfunction and metabolic endotoxemia induced by a cafeteria diet in wistar rats. Nutrients 2019, 11, 979. [Google Scholar] [CrossRef]
- Anitha, M.; Reichardt, F.; Tabatabavakili, S.; Nezami, B.G.; Chassaing, B.; Mwangi, S.; Vijay-Kumar, M.; Gewirtz, A.; Srinivasan, S. Intestinal dysbiosis contributes to the delayed gastrointestinal transit in high-fat diet fed mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 328–339. [Google Scholar] [CrossRef]
- Reichardt, F.; Chassaing, B.; Nezami, B.G.; Li, G.; Tabatabavakili, S.; Mwangi, S.; Uppal, K.; Liang, B.; Vijay-Kumar, M.; Jones, D.; et al. Western diet induces colonic nitrergic myenteric neuropathy and dysmotility in mice via saturated fatty acid- and lipopolysaccharide-induced TLR4 signalling. J. Physiol. 2017, 595, 1831–1846. [Google Scholar] [CrossRef]
- Zhou, X.-L.; Yan, B.; Xiao, Y.; Zhou, Y.; Liu, T. Tartary buckwheat protein prevented dyslipidemia in high-fat diet-fed mice associated with gut microbiota changes. Food Chem. Toxicol. 2018, 119, 296–301. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Ginés, I.; Pinent, M.; Ardévol, A.; Blay, M.; Terra, X. The co-administration of proanthocyanidins and an obesogenic diet prevents the increase in intestinal permeability and metabolic endotoxemia derived to the diet. J. Nutr. Biochem. 2018, 62, 35–42. [Google Scholar] [CrossRef]
- Dey, P.; Sasaki, G.Y.; Wei, P.; Li, J.; Wang, L.; Zhu, J.; McTigue, D.; Yu, Z.; Bruno, R.S. Green tea extract prevents obesity in male mice by alleviating gut dysbiosis in association with improved intestinal barrier function that limits endotoxin translocation and adipose inflammation. J. Nutr. Biochem. 2019, 67, 78–89. [Google Scholar] [CrossRef]
- Vreugdenhil, A.C.E.; Rousseau, C.H.; Hartung, T.; Greve, J.W.M.; van ’t Veer, C.; Buurman, W.A. Lipopolysaccharide (LPS)-binding protein mediates LPS detoxification by chylomicrons. J. Immunol. 2003, 170, 1399–1405. [Google Scholar] [CrossRef]
- Vergès, B.; Duvillard, L.; Lagrost, L.; Vachoux, C.; Garret, C.; Bouyer, K.; Courtney, M.; Pomié, C.; Burcelin, R. Changes in lipoprotein kinetics associated with type 2 diabetes affect the distribution of lipopolysaccharides among lipoproteins. J. Clin. Endocrinol. Metab. 2014, 99, E1245–E1253. [Google Scholar] [CrossRef]
- Levels, J.H.M.; Marquart, J.A.; Abraham, P.R.; van den Ende, A.E.; Molhuizen, H.O.F.; van Deventer, S.J.H.; Meijers, J.C.M. Lipopolysaccharide is transferred from high-density to low-density lipoproteins by lipopolysaccharide-binding protein and phospholipid transfer protein. Infect. Immun. 2005, 73, 2321–2326. [Google Scholar] [CrossRef]
- Emancipator, K.; Csako, G.; Elin, R.J. In vitro inactivation of bacterial endotoxin by human lipoproteins and apolipoproteins. Infect. Immun. 1992, 60, 596–601. [Google Scholar]
- Correa, W.; Brandenburg, K.; Zähringer, U.; Ravuri, K.; Khan, T.; von Wintzingerode, F. Biophysical analysis of lipopolysaccharide formulations for an understanding of the low endotoxin recovery (LER) phenomenon. Int. J. Mol. Sci. 2017, 18, 2737. [Google Scholar] [CrossRef]
- Yao, Z.; Mates, J.M.; Cheplowitz, A.M.; Hammer, L.P.; Maiseyeu, A.; Phillips, G.S.; Wewers, M.D.; Rajaram, M.V.S.; Robinson, J.M.; Anderson, C.L.; et al. Blood-Borne lipopolysaccharide is rapidly eliminated by liver sinusoidal endothelial cells via high-density lipoprotein. J. Immunol. 2016, 197, 2390–2399. [Google Scholar] [CrossRef]
- van Bergenhenegouwen, J.; Kraneveld, A.D.; Rutten, L.; Garssen, J.; Vos, A.P.; Hartog, A. Lipoproteins attenuate TLR2 and TLR4 activation by bacteria and bacterial ligands with differences in affinity and kinetics. BMC Immunol. 2016, 17, 42. [Google Scholar] [CrossRef]
- Wiesner, P.; Choi, S.; Almazan, F.; Benner, C.; Huang, W.; Diehl, C.J.; Gonen, A.; Butler, S.; Witztum, J.L.; Glass, C.K.; et al. Low doses of lipopolysaccharide and minimally oxidized low-density lipoprotein cooperatively activate macrophages via nuclear factor kappa B and activator protein-1: Possible mechanism for acceleration of atherosclerosis by subclinical endotoxemia. Circ. Res. 2010, 107, 56–65. [Google Scholar] [CrossRef]
- Kim, H.I.; Kim, J.-K.; Kim, J.-Y.; Jang, S.E.; Han, M.J.; Kim, D.-H. Lactobacillus plantarum LC27 and Bifidobacterium longum LC67 simultaneously alleviate high-fat diet-induced colitis, endotoxemia, liver steatosis, and obesity in mice. Nutr. Res. 2019, 67, 78–89. [Google Scholar] [CrossRef]
- Shao, B.; Munford, R.S.; Kitchens, R.; Varley, A.W. Hepatic uptake and deacylation of the LPS in bloodborne LPS-lipoprotein complexes. Innate Immun. 2012, 18, 825–833. [Google Scholar] [CrossRef]
- Xie, G.; Wang, X.; Liu, P.; Wei, R.; Chen, W.; Rajani, C.; Hernandez, B.Y.; Alegado, R.; Dong, B.; Li, D.; et al. Distinctly altered gut microbiota in the progression of liver disease. Oncotarget 2016, 7, 19355–19366. [Google Scholar] [CrossRef]
- Dandekar, A.; Qiu, Y.; Kim, H.; Wang, J.; Hou, X.; Zhang, X.; Zheng, Z.; Mendez, R.; Yu, F.-S.; Kumar, A.; et al. Toll-like receptor (TLR) signaling interacts with CREBH to modulate high-density lipoprotein (HDL) in response to bacterial endotoxin. J. Biol. Chem. 2016, 291, 23149–23158. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Leber, B.; Tripolt, N.J.; Blattl, D.; Eder, M.; Wascher, T.C.; Pieber, T.R.; Stauber, R.; Sourij, H.; Oettl, K.; Stadlbauer, V. The influence of probiotic supplementation on gut permeability in patients with metabolic syndrome: An open label, randomized pilot study. Eur. J. Clin. Nutr. 2012, 66, 1110–1115. [Google Scholar] [CrossRef]
- Pei, R.; DiMarco, D.M.; Putt, K.K.; Martin, D.A.; Gu, Q.; Chitchumroonchokchai, C.; White, H.M.; Scarlett, C.O.; Bruno, R.S.; Bolling, B.W. Low-fat yogurt consumption reduces biomarkers of chronic inflammation and inhibits markers of endotoxin exposure in healthy premenopausal women: A randomised controlled trial. Br. J. Nutr. 2017, 118, 1043–1051. [Google Scholar] [CrossRef]
- Pei, R.; DiMarco, D.M.; Putt, K.K.; Martin, D.A.; Chitchumroonchokchai, C.; Bruno, R.S.; Bolling, B.W. Premeal low-fat yogurt consumption reduces postprandial inflammation and markers of endotoxin exposure in healthy premenopausal women in a randomized controlled trial. J. Nutr. 2018, 148, 910–916. [Google Scholar] [CrossRef]
- Chan, Y.K.; Brar, M.S.; Kirjavainen, P.V.; Chen, Y.; Peng, J.; Li, D.; Leung, F.C.; El-Nezami, H. High fat diet induced atherosclerosis is accompanied with low colonic bacterial diversity and altered abundances that correlates with plaque size, plasma A-FABP and cholesterol: A pilot study of high fat diet and its intervention with Lactobacillus rhamno. BMC Microbiol. 2016, 16, 264. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Robles-Sánchez, C.; Abadía-Molina, F.; Morón-Calvente, V.; Sáez-Lara, M.J.; Ruiz-Bravo, A.; Jiménez-Valera, M.; Gil, Á.; Gómez-Llorente, C.; Fontana, L. Adamdec1, Ednrb and Ptgs1/Cox1, inflammation genes upregulated in the intestinal mucosa of obese rats, are downregulated by three probiotic strains. Sci. Rep. 2017, 7, 1939. [Google Scholar] [CrossRef]
- Jang, H.-M.; Han, S.-K.; Kim, J.-K.; Oh, S.-J.; Jang, H.-B.; Kim, D.-H. Lactobacillus sakei alleviates high-fat-diet-induced obesity and anxiety in mice by inducing AMPK activation and SIRT1 expression and inhibiting gut microbiota-mediated NF-κB activation. Mol. Nutr. Food Res. 2019, 63, e1800978. [Google Scholar] [CrossRef]
- Kikuchi, K.; Ben Othman, M.; Sakamoto, K. Sterilized bifidobacteria suppressed fat accumulation and blood glucose level. Biochem. Biophys. Res. Commun. 2018, 501, 1041–1047. [Google Scholar] [CrossRef]
- Xue, L.; He, J.; Gao, N.; Lu, X.; Li, M.; Wu, X.; Liu, Z.; Jin, Y.; Liu, J.; Xu, J.; et al. Probiotics may delay the progression of nonalcoholic fatty liver disease by restoring the gut microbiota structure and improving intestinal endotoxemia. Sci. Rep. 2017, 7, 45176. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Parnell, J.A.; Klancic, T.; Reimer, R.A. Oligofructose decreases serum lipopolysaccharide and plasminogen activator inhibitor-1 in adults with overweight/obesity. Obesity 2017, 25, 510–513. [Google Scholar] [CrossRef]
- Dehghan, P.; Pourghassem Gargari, B.; Asghari Jafar-abadi, M. Oligofructose-enriched inulin improves some inflammatory markers and metabolic endotoxemia in women with type 2 diabetes mellitus: A randomized controlled clinical trial. Nutrition 2014, 30, 418–423. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.D.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.B.; Neyrinck, A.M.; Bindels, L.B.; de Vos, W.M.; Gibson, G.R.; Thissen, J.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2013, 62, 1112–1121. [Google Scholar] [CrossRef]
- Pedersen, C.; Gallagher, E.; Horton, F.; Ellis, R.J.; Ijaz, U.Z.; Wu, H.; Jaiyeola, E.; Diribe, O.; Duparc, T.; Cani, P.D.; et al. Host-microbiome interactions in human type 2 diabetes following prebiotic fibre (galacto-oligosaccharide) intake. Br. J. Nutr. 2016, 116, 1869–1877. [Google Scholar] [CrossRef]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.E.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of diet with Galacto-oligosaccharides increases bifidobacteria, but not insulin sensitivity, in obese prediabetic individuals. Gastroenterology 2017, 153, 87–97. [Google Scholar] [CrossRef]
- Morel, F.B.; Dai, Q.; Ni, J.; Thomas, D.; Parnet, P.; Fança-Berthon, P. α-Galacto-oligosaccharides dose-dependently reduce appetite and decrease inflammation in overweight adults. J. Nutr. 2015, 145, 2052–2059. [Google Scholar] [CrossRef]
- Farhangi, M.A.; Javid, A.Z.; Sarmadi, B.; Karimi, P.; Dehghan, P. A randomized controlled trial on the efficacy of resistant dextrin, as functional food, in women with type 2 diabetes: Targeting the hypothalamic-pituitary-adrenal axis and immune system. Clin. Nutr. 2018, 37, 1216–1223. [Google Scholar] [CrossRef]
- Ghanim, H.; Batra, M.; Abuaysheh, S.; Green, K.; Makdissi, A.; Kuhadiya, N.D.; Chaudhuri, A.; Dandona, P. Antiinflammatory and ROS suppressive effects of the addition of fiber to a high-fat high-calorie meal. J. Clin. Endocrinol. Metab. 2017, 102, 858–869. [Google Scholar]
- Kopf, J.C.; Suhr, M.J.; Clarke, J.; Eyun, S.; Riethoven, J.M.; Ramer-Tait, A.E.; Rose, D.J. Role of whole grains versus fruits and vegetables in reducing subclinical inflammation and promoting gastrointestinal health in individuals affected by overweight and obesity: A randomized controlled trial. Nutr. J. 2018, 17, 72. [Google Scholar] [CrossRef]
- de La Serre, C.B.; de Lartigue, G.; Raybould, H.E. Chronic exposure to low dose bacterial lipopolysaccharide inhibits leptin signaling in vagal afferent neurons. Physiol. Behav. 2015, 139, 188–194. [Google Scholar] [CrossRef]
- Afreen, A.; Ahmed, Z.; Anjum, N. Novel nutraceutical compounds. In Therapeutic, Probiotic, and Unconventional Foods; Elsevier: Amsterdam, The Netherlands, 2018; pp. 201–226. [Google Scholar]
- Chiva-Blanch, G.; Badimon, L. Effects of polyphenol intake on metabolic syndrome: Current evidences from human trials. Oxid. Med. Cell. Longev. 2017, 2017, 5812401. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef]
- Clemente-Postigo, M.; Queipo-Ortuño, M.I.; Boto-Ordoñez, M.; Coin-Aragüez, L.; Roca-Rodriguez, M.D.M.; Delgado-Lista, J.; Cardona, F.; Andres-Lacueva, C.; Tinahones, F.J. Effect of acute and chronic red wine consumption on lipopolysaccharide concentrations. Am. J. Clin. Nutr. 2013, 97, 1053–1061. [Google Scholar] [CrossRef]
- Campbell, C.L.; Yu, R.; Li, F.; Zhou, Q.; Chen, D.; Qi, C.; Yin, Y.; Sun, J. Modulation of fat metabolism and gut microbiota by resveratrol on high-fat diet-induced obese mice. Diabetes. Metab. Syndr. Obes. 2019, 12, 97–107. [Google Scholar] [CrossRef]
- Masumoto, S.; Terao, A.; Yamamoto, Y.; Mukai, T.; Miura, T.; Shoji, T. Non-absorbable apple procyanidins prevent obesity associated with gut microbial and metabolomic changes. Sci. Rep. 2016, 6, 31208. [Google Scholar] [CrossRef]
- López, P.; Sánchez, M.; Perez-Cruz, C.; Velázquez-Villegas, L.A.; Syeda, T.; Aguilar-López, M.; Rocha-Viggiano, A.K.; Del Carmen Silva-Lucero, M.; Torre-Villalvazo, I.; Noriega, L.G.; et al. Long-term genistein consumption modifies gut microbiota, improving glucose metabolism, metabolic endotoxemia, and cognitive function in mice fed a high-fat diet. Mol. Nutr. Food Res. 2018, 62, e1800313. [Google Scholar] [CrossRef]
- Kaliannan, K.; Robertson, R.C.; Murphy, K.; Stanton, C.; Kang, C.; Wang, B.; Hao, L.; Bhan, A.K.; Kang, J.X. Estrogen-mediated gut microbiome alterations influence sexual dimorphism in metabolic syndrome in mice. Microbiome 2018, 6, 205. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Kim, J.; Lee, J.H.; Sim, J.H.; Cho, D.; Bae, I.; Lee, H.; Seol, M.A.; Shin, H.M.; Kim, T.-J.; et al. Modulation of gut microbiota and delayed immunosenescence as a result of syringaresinol consumption in middle-aged mice. Sci. Rep. 2016, 6, 39026. [Google Scholar] [CrossRef]
- Liu, A.; Chen, M.; Kumar, R.; Stefanovic-Racic, M.; O’Doherty, R.M.; Ding, Y.; Jahnen-Dechent, W.; Borghesi, L. Bone marrow lympho-myeloid malfunction in obesity requires precursor cell-autonomous TLR4. Nat. Commun. 2018, 9, 708. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wang, L.; Wen, C.; Yang, J.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber and its depolymerized derivative prevent obesity in association with modification of gut microbiota in high-fat diet-fed mice. Mol. Nutr. Food Res. 2018, 62, e1800446. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wu, S.; Wang, L.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber modulates the gut microbiota and its metabolites in normal mice. Int. J. Biol. Macromol. 2018, 120, 502–512. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Xu, Y.; Yang, H.; Wang, J.; Xue, C.; Yan, X.; Su, L. Anti-inflammation effects of fucosylated chondroitin sulphate from acaudina molpadioides by altering gut microbiota in obese mice. Food Funct. 2019, 10, 1736–1746. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, N.; Li, Z.; Wang, X.; Shi, H.; Xue, C.; Li, R.W.; Tang, Q. Chondroitin sulfate disaccharides modified the structure and function of the murine gut microbiome under healthy and stressed conditions. Sci. Rep. 2017, 7, 6783. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, M.; Zhang, P.; Fan, S.; Huang, J.; Yu, S.; Zhang, C.; Li, H. Fucoidan and galactooligosaccharides ameliorate high-fat diet-induced dyslipidemia in rats by modulating the gut microbiota and bile acid metabolism. Nutrition 2019, 65, 50–59. [Google Scholar] [CrossRef]
- Abboud, K.Y.; Reis, S.K.; Martelli, M.E.; Zordão, O.P.; Tannihão, F.; de Souza, A.Z.Z.; Assalin, H.B.; Guadagnini, D.; Rocha, G.Z.; Saad, M.J.A.; et al. Oral glutamine supplementation reduces obesity, pro-inflammatory markers, and improves insulin sensitivity in DIO wistar rats and reduces waist circumference in overweight and obese humans. Nutrients 2019, 11, 536. [Google Scholar] [CrossRef]
- Luthold, R.V.; Fernandes, G.R.; Franco-de-Moraes, A.C.; Folchetti, L.G.D.; Ferreira, S.R.G. Gut microbiota interactions with the immunomodulatory role of vitamin D in normal individuals. Metabolism. 2017, 69, 76–86. [Google Scholar] [CrossRef]
- Ryz, N.R.; Lochner, A.; Bhullar, K.; Ma, C.; Huang, T.; Bhinder, G.; Bosman, E.; Wu, X.; Innis, S.M.; Jacobson, K.; et al. Dietary vitamin D3 deficiency alters intestinal mucosal defense and increases susceptibility to Citrobacter rodentium-induced colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G730–G742. [Google Scholar] [CrossRef]
- Everard, A.; Geurts, L.; Van Roye, M.; Delzenne, N.M.; Cani, P.D. Tetrahydro iso-alpha acids from hops improve glucose homeostasis and reduce body weight gain and metabolic endotoxemia in high-fat diet-fed mice. PLoS ONE 2012, 7, e33858. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.; Zhang, P.; Wang, H.; Zhang, Q.; Yu, S.; Yu, Y. Chronic rhein treatment improves recognition memory in high-fat diet-induced obese male mice. J. Nutr. Biochem. 2016, 36, 42–50. [Google Scholar] [CrossRef]
- Mei, X.; Zhang, X.; Wang, Z.; Gao, Z.; Liu, G.; Hu, H.; Zou, L.; Li, X. Insulin sensitivity-enhancing activity of phlorizin Is associated with lipopolysaccharide decrease and gut microbiota changes in obese and type 2 diabetes (db/db) mice. J. Agric. Food Chem. 2016, 64, 7502–7511. [Google Scholar] [CrossRef]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut microbiota mediates the protective effects of dietary capsaicin against chronic low-grade Inflammation and associated obesity induced by high-fat diet. MBio 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Guo, X.; Tang, R.; Yang, S.; Lu, Y.; Luo, J.; Liu, Z. Rutin and its combination with Inulin attenuate gut dysbiosis, the inflammatory status and endoplasmic reticulum stress in paneth cells of obese mice induced by high-fat diet. Front. Microbiol. 2018, 9, 2651. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Li, B.; Qiang, Y.; Yuan, T.; Tan, X.; Wang, Z.; Liu, Z.; Liu, X. Lycopene attenuates western-diet-induced cognitive deficits via improving glycolipid metabolism dysfunction and inflammatory responses in gut-liver-brain axis. Int. J. Obes. 2018. [Google Scholar] [CrossRef]
- Nagata, N.; Xu, L.; Kohno, S.; Ushida, Y.; Aoki, Y.; Umeda, R.; Fuke, N.; Zhuge, F.; Ni, Y.; Nagashimada, M.; et al. Glucoraphanin ameliorates obesity and insulin resistance through adipose tissue browning and reduction of metabolic endotoxemia in mice. Diabetes 2017, 66, 1222–1236. [Google Scholar] [CrossRef]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef]
- Anhê, F.F.; Nachbar, R.T.; Varin, T.V.; Trottier, J.; Dudonné, S.; Le Barz, M.; Feutry, P.; Pilon, G.; Barbier, O.; Desjardins, Y.; et al. Treatment with camu camu (Myrciaria dubia) prevents obesity by altering the gut microbiota and increasing energy expenditure in diet-induced obese mice. Gut 2018, 1–12. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, S.; Park, J.Y.; Harvatine, K.; Lambert, J.D. Dietary cocoa reduces metabolic endotoxemia and adipose tissue inflammation in high-fat fed mice. J. Nutr. Biochem. 2014, 25, 439–445. [Google Scholar] [CrossRef]
- Sánchez-Tapia, M.; Aguilar-López, M.; Pérez-Cruz, C.; Pichardo-Ontiveros, E.; Wang, M.; Donovan, S.M.; Tovar, A.R.; Torres, N. Nopal (Opuntia ficus indica) protects from metabolic endotoxemia by modifying gut microbiota in obese rats fed high fat/sucrose diet. Sci. Rep. 2017, 7, 4716. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Tang, R. Changes in mouse gut microbial community in response to the different types of commonly consumed meat. Microorganisms 2019, 7, 76. [Google Scholar] [CrossRef]
- Peng, J.; Leng, J.; Tian, H.; Yang, T.; Fang, Y.; Feng, Q.; Zhao, Y.; Hu, Y.-Y. Geniposide and chlorogenic acid combination ameliorates non-alcoholic steatohepatitis involving the protection on the gut barrier function in mouse induced by high-fat diet. Front. Pharmacol. 2018, 9, 1399. [Google Scholar] [CrossRef]
- Han, L.; Li, T.; Du, M.; Chang, R.; Zhan, B.; Mao, X. Beneficial effects of potentilla discolor bunge water extract on inflammatory cytokines release and gut microbiota in high-fat diet and streptozotocin-induced type 2 diabetic mice. Nutrients 2019, 11, 670. [Google Scholar] [CrossRef]
- Chang, C.; Lin, C.; Lu, C.; Martel, J.; Ko, Y.; Ojcius, D.M.; Tseng, S.; Wu, T.; Chen, Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Dong, Y.; Zhu, T.; Luo, Y.-Y.; Kang, X.-J.; Pang, M.-X.; Li, H.-Z.; Xu, H.; Gu, C.; Pan, S.-H.; et al. Semen hoveniae extract ameliorates alcohol-induced chronic liver damage in rats via modulation of the abnormalities of gut-liver axis. Phytomedicine 2019, 52, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, K.; Deng, Y.; Chen, R.; Liang, S.; Xie, H.; He, Y.; Chen, Y.; Yang, Q. Effects of shenling baizhu powder herbal formula on intestinal microbiota in high-fat diet-induced NAFLD rats. Biomed. Pharmacother. 2018, 102, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Ahola, A.J.; Lassenius, M.I.; Forsblom, C.; Harjutsalo, V.; Lehto, M.; Groop, P.-H. Dietary patterns reflecting healthy food choices are associated with lower serum LPS activity. Sci. Rep. 2017, 7, 6511. [Google Scholar] [CrossRef] [PubMed]
- Amar, J.; Burcelin, R.; Ruidavets, J.B.; Cani, P.D.; Fauvel, J.; Alessi, M.C.; Chamontin, B.; Ferriéres, J. Energy intake is associated with endotoxemia in apparently healthy men. Am. J. Clin. Nutr. 2008, 87, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Ott, B.; Skurk, T.; Hastreiter, L.; Lagkouvardos, I.; Fischer, S.; Büttner, J.; Kellerer, T.; Clavel, T.; Rychlik, M.; Haller, D.; et al. Effect of caloric restriction on gut permeability, inflammation markers, and fecal microbiota in obese women. Sci. Rep. 2017, 7, 11955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, S.; Yang, L.; Huang, P.; Li, W.; Wang, S.; Zhao, G.; Zhang, M.; Pang, X.; Yan, Z.; et al. Structural modulation of gut microbiota in life-long calorie-restricted mice. Nat. Commun. 2013, 4, 2163. [Google Scholar] [CrossRef] [PubMed]
- Fabbiano, S.; Suárez-Zamorano, N.; Chevalier, C.; Lazarević, V.; Kieser, S.; Rigo, D.; Leo, S.; Veyrat-Durebex, C.; Gaïa, N.; Maresca, M.; et al. Functional gut microbiota remodeling contributes to the caloric restriction-induced metabolic improvements. Cell Metab. 2018, 28, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, M.; Vacante, M.; Antic, T.; Giordano, M.; Chisari, G.; Acquaviva, R.; Mastrojeni, S.; Malaguarnera, G.; Mistretta, A.; Li Volti, G.; et al. Bifidobacterium longum with fructo-oligosaccharides in patients with non alcoholic steatohepatitis. Dig. Dis. Sci. 2012, 57, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Röytiö, H.; Mokkala, K.; Vahlberg, T.; Laitinen, K. Dietary intake of fat and fibre according to reference values relates to higher gut microbiota richness in overweight pregnant women. Br. J. Nutr. 2017, 118, 343–352. [Google Scholar] [CrossRef]
- Rios, J.L.; Bomhof, M.R.; Reimer, R.A.; Hart, D.A.; Collins, K.H.; Herzog, W. Protective effect of prebiotic and exercise intervention on knee health in a rat model of diet-induced obesity. Sci. Rep. 2019, 9, 3893. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhang, L.; Xue, J.; Yang, X.; Dong, X.; Sha, L.; Lei, H.; Zhang, X.; Zhu, L.; Wang, Z.; et al. Dietary inulin alleviates diverse stages of type 2 diabetes mellitus via anti-inflammation and modulating gut microbiota in db/db mice. Food Funct. 2019, 10, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Van Hée, V.F.; Piront, N.; De Backer, F.; Toussaint, O.; Cani, P.D.; Delzenne, N.M. Wheat-derived arabinoxylan oligosaccharides with prebiotic effect increase satietogenic gut peptides and reduce metabolic endotoxemia in diet-induced obese mice. Nutr. Diabetes 2012, 2, e28. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, K.R.; Liu, S.X.L.; Tian, R.; Kushnir, A.; Turner, J.R.; Li, H.-L.; Chou, P.M.; Weber, C.R.; De Plaen, I.G. Bifidobacteria stabilize claudins at tight junctions and prevent intestinal barrier dysfunction in mouse necrotizing enterocolitis. Am. J. Pathol. 2013, 182, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, W.; Zuo, L.; Zhu, W.; Wang, B.; Li, Q.; Li, J. Bifidobacteria may be beneficial to intestinal microbiota and reduction of bacterial translocation in mice following ischaemia and reperfusion injury. Br. J. Nutr. 2013, 109, 1990–1998. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-Chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell. Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef]
- Yildiz, H.M.; Carlson, T.L.; Goldstein, A.M.; Carrier, R.L. Mucus barriers to microparticles and microbes are altered in hirschsprung’s disease. Macromol. Biosci. 2015, 15, 712–718. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, L.; Dou, X.; Wang, C.; Zhang, W.; Gao, K.; Liu, J.; Wang, H. Lactobacillus reuteri ZJ617 maintains intestinal integrity via regulating tight junction, autophagy and apoptosis in mice challenged with lipopolysaccharide. Oncotarget 2017, 8, 77489–77499. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, J.; Quan, G.; Wang, X.; Yang, L.; Zhong, L. Lactobacillus acidophilus ATCC 4356 prevents atherosclerosis via inhibition of intestinal cholesterol absorption in apolipoprotein E-knockout mice. Appl. Environ. Microbiol. 2014, 80, 7496–7504. [Google Scholar] [CrossRef]
- Ambalam, P.; Kondepudi, K.K.; Balusupati, P.; Nilsson, I.; Wadström, T.; Ljungh, Å. Prebiotic preferences of human lactobacilli strains in co-culture with bifidobacteria and antimicrobial activity against Clostridium difficile. J. Appl. Microbiol. 2015, 119, 1672–1682. [Google Scholar] [CrossRef]
- Sims, I.M.; Ryan, J.L.J.; Kim, S.H. In vitro fermentation of prebiotic oligosaccharides by Bifidobacterium lactis HN019 and Lactobacillus spp. Anaerobe 2014, 25, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Amirdivani, S.; Baba, A.S.H. Green tea yogurt: Major phenolic compounds and microbial growth. J. Food Sci. Technol. 2015, 52, 4652–4660. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Xie, Y.; Peng, C.; Liu, Y.; Wang, H. A novel antidiabetic food produced via solid-state fermentation of tartary buckwheat by L. plantarum TK9 and L. paracasei TK1501. Food Technol. Biotechnol. 2018, 56, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-Y.; Liao, N.-B.; Zhang, Y.; Ye, X.-Q.; Li, S.; Hu, Y.-Q.; Liu, D.-H.; Linhardt, R.J.; Wang, X.; Chen, S.-G. In vitro fermentation behaviors of fucosylated chondroitin sulfate from pearsonothuria graeffei by human gut microflora. Int. J. Biol. Macromol. 2017, 102, 1195–1201. [Google Scholar] [CrossRef]
- Medina, D.A.; Pinto, F.; Ovalle, A.; Thomson, P.; Garrido, D. Prebiotics mediate microbial interactions in a consortium of the infant gut microbiome. Int. J. Mol. Sci. 2017, 18, 2095. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef]
- Tan, H.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Novel strains of Bacteroides fragilis and Bacteroides ovatus alleviate the LPS-induced inflammation in mice. Appl. Microbiol. Biotechnol. 2019, 103, 2353–2365. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef]
- Cartmell, A.; Lowe, E.C.; Baslé, A.; Firbank, S.J.; Ndeh, D.A.; Murray, H.; Terrapon, N.; Lombard, V.; Henrissat, B.; Turnbull, J.E.; et al. How members of the human gut microbiota overcome the sulfation problem posed by glycosaminoglycans. Proc. Natl. Acad. Sci. USA 2017, 114, 7037–7042. [Google Scholar] [CrossRef]
- Salyers, A.A.; O’Brien, M.; Kotarski, S.F. Utilization of chondroitin sulfate by Bacteroides thetaiotaomicron growing in carbohydrate-limited continuous culture. J. Bacteriol. 1982, 150, 1008–1015. [Google Scholar] [PubMed]
- Earley, H.; Lennon, G.; Balfe, A.; Kilcoyne, M.; Clyne, M.; Joshi, L.; Carrington, S.; Martin, S.T.; Coffey, J.C.; Winter, D.C.; et al. A preliminary study examining the binding capacity of Akkermansia muciniphila and desulfovibrio spp., to colonic mucin in health and ulcerative colitis. PLoS ONE 2015, 10, e0135280. [Google Scholar] [CrossRef] [PubMed]
- Ottman, N.; Davids, M.; Suarez-Diez, M.; Boeren, S.; Schaap, P.J.; Martins Dos Santos, V.A.P.; Smidt, H.; Belzer, C.; de Vos, W.M. Genome-scale model and omics analysis of metabolic capacities of Akkermansia muciniphila reveal a preferential mucin-degrading lifestyle. Appl. Environ. Microbiol. 2017, 83, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Van Herreweghen, F.; Van den Abbeele, P.; De Mulder, T.; De Weirdt, R.; Geirnaert, A.; Hernandez-Sanabria, E.; Vilchez-Vargas, R.; Jauregui, R.; Pieper, D.H.; Belzer, C.; et al. In vitro colonisation of the distal colon by Akkermansia muciniphila is largely mucin and pH dependent. Benef. Microbes 2017, 8, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Chia, L.W.; Hornung, B.V.H.; Aalvink, S.; Schaap, P.J.; de Vos, W.M.; Knol, J.; Belzer, C. Deciphering the trophic interaction between Akkermansia muciniphila and the butyrogenic gut commensal Anaerostipes caccae using a metatranscriptomic approach. Antonie Van Leeuwenhoek 2018, 111, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Choi, Y.; Kim, D.-K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.-S.; Jee, Y.-K.; Gho, Y.S.; et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Romo-Vaquero, M.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal simulation model TWIN-SHIME shows differences between human Urolithin-Metabotypes in gut microbiota composition, Pomegranate Polyphenol Metabolism, and Transport along the Intestinal Tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef]
- Mirande, C.; Kadlecikova, E.; Matulova, M.; Capek, P.; Bernalier-Donadille, A.; Forano, E.; Béra-Maillet, C. Dietary fibre degradation and fermentation by two xylanolytic bacteria Bacteroides xylanisolvens XB1A and Roseburia intestinalis XB6B4 from the human intestine. J. Appl. Microbiol. 2010, 109, 451–460. [Google Scholar]
- Patterson, A.M.; Mulder, I.E.; Travis, A.J.; Lan, A.; Cerf-Bensussan, N.; Gaboriau-Routhiau, V.; Garden, K.; Logan, E.; Delday, M.I.; Coutts, A.G.P.; et al. Human gut symbiont roseburia hominis promotes and regulates innate immunity. Front. Immunol. 2017, 8, 1166. [Google Scholar] [CrossRef] [PubMed]
- Falony, G.; Vlachou, A.; Verbrugghe, K.; De Vuyst, L. Cross-feeding between Bifidobacterium longum BB536 and acetate-converting, butyrate-producing colon bacteria during growth on oligofructose. Appl. Environ. Microbiol. 2006, 72, 7835–7841. [Google Scholar] [CrossRef] [PubMed]
- Chilton, P.M.; Hadel, D.M.; To, T.T.; Mitchell, T.C.; Darveau, R.P. Adjuvant activity of naturally occurring monophosphoryl lipopolysaccharide preparations from mucosa-associated bacteria. Infect. Immun. 2013, 81, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Franke, T.; Deppenmeier, U. Physiology and central carbon metabolism of the gut bacterium Prevotella copri. Mol. Microbiol. 2018, 109, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Spiga, L.; Winter, M.G.; Furtado de Carvalho, T.; Zhu, W.; Hughes, E.R.; Gillis, C.C.; Behrendt, C.L.; Kim, J.; Chessa, D.; Andrews-Polymenis, H.L.; et al. An oxidative central metabolism enables salmonella to utilize microbiota-derived succinate. Cell Host Microbe 2017, 22, 291–301. [Google Scholar] [CrossRef]
- Ferreyra, J.A.; Wu, K.J.; Hryckowian, A.J.; Bouley, D.M.; Weimer, B.C.; Sonnenburg, J.L. Gut microbiota-produced succinate promotes C. difficile infection after antibiotic treatment or motility disturbance. Cell Host Microbe 2014, 16, 770–777. [Google Scholar] [CrossRef]
- Macias-Ceja, D.C.; Ortiz-Masiá, D.; Salvador, P.; Gisbert-Ferrándiz, L.; Hernández, C.; Hausmann, M.; Rogler, G.; Esplugues, J.V.; Hinojosa, J.; Alós, R.; et al. Succinate receptor mediates intestinal inflammation and fibrosis. Mucosal Immunol. 2019, 12, 178–187. [Google Scholar] [CrossRef]
- Leite, A.Z.; Rodrigues, N.d.C.; Gonzaga, M.I.; Paiolo, J.C.C.; de Souza, C.A.; Stefanutto, N.A.V.; Omori, W.P.; Pinheiro, D.G.; Brisotti, J.L.; Matheucci Junior, E.; et al. Detection of increased plasma Interleukin-6 levels and prevalence of prevotella copri and bacteroides vulgatus in the feces of type 2 diabetes Patients. Front. Immunol. 2017, 8, 1107. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef]
- Gronbach, K.; Flade, I.; Holst, O.; Lindner, B.; Ruscheweyh, H.J.; Wittmann, A.; Menz, S.; Schwiertz, A.; Adam, P.; Stecher, B.; et al. Endotoxicity of lipopolysaccharide as a determinant of T-cell-mediated colitis induction in mice. Gastroenterology 2014, 146, 765–775. [Google Scholar] [CrossRef]
- Marion, S.; Studer, N.; Desharnais, L.; Menin, L.; Escrig, S.; Meibom, A.; Hapfelmeier, S.; Bernier-Latmani, R. In vitro and in vivo characterization of Clostridium scindens bile acid transformations. Gut Microbes 2019, 10, 481–503. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Dong, W.; Wang, S.; Zhang, Y.; Liu, T.; Xie, R.; Wang, B.; Cao, H. Deoxycholic acid disrupts the intestinal mucosal barrier and promotes intestinal tumorigenesis. Food Funct. 2018, 9, 5588–5597. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Sánchez-Patán, F.; Monagas, M.; Walton, G.E.; Gibson, G.R.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. In vitro fermentation of grape seed flavan-3-ol fractions by human faecal microbiota: Changes in microbial groups and phenolic metabolites. FEMS Microbiol. Ecol. 2013, 83, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Dávila-Aviña, J.; Castillo, S.L.; Heredia, N.; Vázquez-Alvarado, R.; García, S. Antibacterial and antioxidant activities in extracts of fully grown cladodes of 8 cultivars of cactus pear. J. Food Sci. 2014, 79, M659–M664. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Benefiel, A.C.; Gaskins, H.R. Contributions of the microbial hydrogen economy to colonic homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Rowan, F.; Docherty, N.G.; Murphy, M.; Murphy, B.; Calvin Coffey, J.; O’Connell, P.R. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis. Colon Rectum 2010, 53, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Huang, K.; Liu, Y.; Yang, Y.; Tang, H.; Liu, X.; Wang, C.; Chen, H.; Xiong, Y.; Zhang, J.; et al. Modulation of intestinal microbiota by glycyrrhizic acid prevents high-fat diet-enhanced pre-metastatic niche formation and metastasis. Mucosal Immunol. 2019, 12, 945–957. [Google Scholar] [CrossRef]
- Marquet, P.; Duncan, S.H.; Chassard, C.; Bernalier-Donadille, A.; Flint, H.J. Lactate has the potential to promote hydrogen sulphide formation in the human colon. FEMS Microbiol. Lett. 2009, 299, 128–134. [Google Scholar] [CrossRef]
- Ritz, N.L.; Lin, D.M.; Wilson, M.R.; Barton, L.L.; Lin, H.C. Sulfate-reducing bacteria slow intestinal transit in a bismuth-reversible fashion in mice. Neurogastroenterol. Motil. 2017, 29, e12907. [Google Scholar] [CrossRef]
- Tierens, K.F.; Thomma, B.P.; Brouwer, M.; Schmidt, J.; Kistner, K.; Porzel, A.; Mauch-Mani, B.; Cammue, B.P.; Broekaert, W.F. Study of the role of antimicrobial glucosinolate-derived isothiocyanates in resistance of Arabidopsis to microbial pathogens. Plant Physiol. 2001, 125, 1688–1699. [Google Scholar] [CrossRef]
- Andersson, M.X.; Nilsson, A.K.; Johansson, O.N.; Boztaş, G.; Adolfsson, L.E.; Pinosa, F.; Petit, C.G.; Aronsson, H.; Mackey, D.; Tör, M.; et al. Involvement of the electrophilic isothiocyanate sulforaphane in Arabidopsis local defense responses. Plant Physiol. 2015, 167, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, R.J.; van Dam, N.M.; van Loon, J.J.A. Role of glucosinolates in insect-plant relationships and multitrophic interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.-G.; Chen, T.-Y.; Fahey, J.W.; Talalay, P. Keap1-nrf2 signaling: A target for cancer prevention by sulforaphane. Top. Curr. Chem. 2013, 329, 163–177. [Google Scholar] [PubMed]
- Yoshida, K.; Ushida, Y.; Ishijima, T.; Suganuma, H.; Inakuma, T.; Yajima, N.; Abe, K.; Nakai, Y. Broccoli sprout extract induces detoxification-related gene expression and attenuates acute liver injury. World J. Gastroenterol. 2015, 21, 10091–10103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Ushida, Y.; Suganuma, H.; et al. Prophylactic effects of sulforaphane on depression-like behavior and dendritic changes in mice after inflammation. J. Nutr. Biochem. 2017, 39, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shen, Y.; Zhu, Y.; Mupunga, J.; Zou, L.; Liu, C.; Liu, S.; Mao, J. Broccoli ingestion increases the glucosinolate hydrolysis activity of microbiota in the mouse gut. Int. J. Food Sci. Nutr. 2019, 70, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Thimmulappa, R.K.; Mai, K.H.; Srisuma, S.; Kensler, T.W.; Yamamoto, M.; Biswal, S. Identification of Nrf2-regulated genes induced by the chemopreventive agent sulforaphane by oligonucleotide microarray. Cancer Res. 2002, 62, 5196–5203. [Google Scholar]
- Aires, A.; Mota, V.R.; Saavedra, M.J.; Rosa, E.A.S.; Bennett, R.N. The antimicrobial effects of glucosinolates and their respective enzymatic hydrolysis products on bacteria isolated from the human intestinal tract. J. Appl. Microbiol. 2009, 106, 2086–2095. [Google Scholar] [CrossRef]
- Glymenaki, M.; Singh, G.; Brass, A.; Warhurst, G.; McBain, A.J.; Else, K.J.; Cruickshank, S.M. Compositional changes in the gut mucus microbiota precede the onset of colitis-induced inflammation. Inflamm. Bowel Dis. 2017, 23, 912–922. [Google Scholar] [CrossRef]



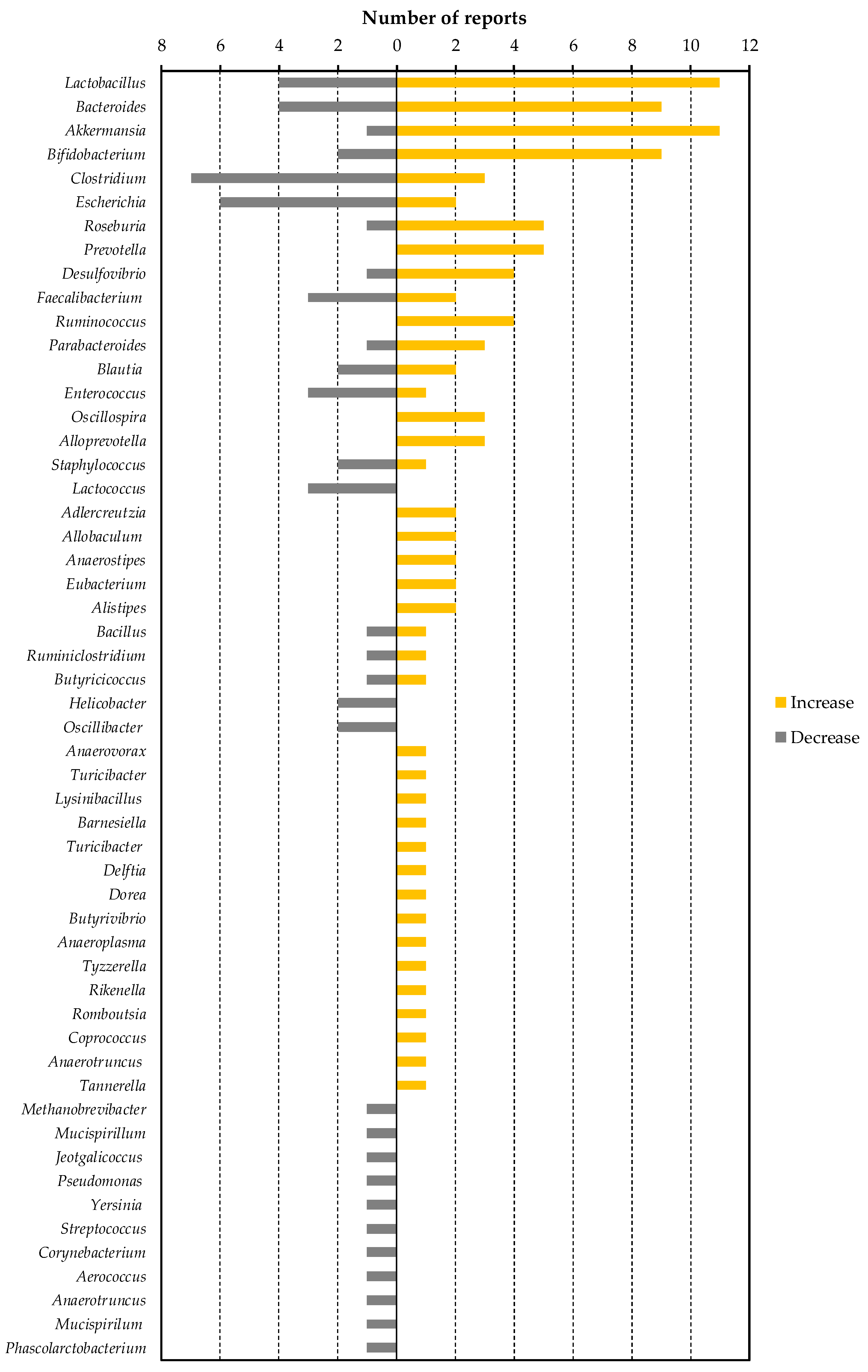{kind=link}
{kind=link}
| Category | Dietary Factor | Dose | Consumption Period | Subject | LPS | LBP | Gut Microbes with Significant Changes in Proportion ** | |
|---|---|---|---|---|---|---|---|---|
| Increase | Decrease | |||||||
| Probiotics/ Prebiotics | Yakult light (Lactobacillus casei Shirota 1 × 108 CFU/mL) [48] | 195 mL | 3 months | Metabolic syndrome | ND | ↑ | — | — |
| Low-fat yogurt [49] | 339 g | 9 weeks | Healthy subject or Obesity | → | → | — | — | |
| Low-fat yogurt [50] | 226 g | Premeal | Healthy subject or Obesity (postprandial endotoxemia was assessed) | → | → | — | — | |
| Oligofructose [57] | 21 g | 12 weeks | Overweight/ Obesity | ↓ | — | — | — | |
| Oligofructose- enriched inulin [58] | 10 g | 8 weeks | Type 2 diabetes | ↓ | — | — | — | |
| Inulin + Oligofructose [59] | 8 g 8 g | 3 months | Obesity | → | — | Bifidobacterium, Faecalibacterium prausnitzii | Bacteroides intestinalis, Bacteroides vulgatus, Propionibacterium | |
| Galacto- oligosaccharide [60] | 5.5 g | 12 weeks | Type 2 diabetes | → | → | none | none | |
| Galacto- oligosaccharide [61] | 15 g | 12 weeks | Overweight/ Obesity | — | → | Bifidobacterium spp. | none | |
| α-Galacto- oligosaccharide [62] | 6–18 g | 14 days | Overweight | ↓ | — | Bifidobacteria | none | |
| Resistant dextrin [63] | 10 g | 8 weeks | Type 2 diabetes | ↓ | — | — | — | |
| Insoluble dietary fiber [from Fiber One Original cereal (General mills)] [64] | 30 g | With high-fat, high-calorie meal | Healthy subject (postprandial endotoxemia was assessed) | ↓* | — | — | — | |
| Whole grains [65] | 3 servings | 6 weeks | Overweight/ Obesity | — | ↓ | none | none | |
| Probiotics/ Prebiotics | Bifidobacterium longum + Oligofructose + Life style modification [110] | — | 24 weeks | Non-alcoholic steatohepatitis | ↓ | — | — | — |
| Polyphenol | Resveratrol + Polyphenol [72] | 100 mg 75 mg | 10 minutes before intake of high-fat high-carbohydrate meal | Healthy subjects (postprandial endotoxemia was assessed) | — | ↓ * | — | — |
| Red wine [73] | 272 mL | With high-fat meal | Healthy subjects (postprandial endotoxemia was assessed) | → | → | — | — | |
| Dietary habits | Fruits + Vegetables [65] | 3 servings | 6 weeks | Overweight/ Obesity | — | ↓ | α-diversity (No significant change in bacterial genera was found) | none |
| Caloric restriction [107] | 800 kcal | 4 weeks | Obesity | — | ↓ | Anaerostipes hadrus, Blautia sp., Ruminococcus faecis, Bifidodbacterium sp. | Agathobacter rectalis | |
| Others | Glutamine [85] | 30 g | 14 weeks | Overweight/ Obesity | ↓ | — | — | — |
| Subject | Number of Subject | Correlation of Dietary Factor and Gut Microbe * | Correlation of Blood LPS and Gut Microbe | Correlation of Blood LPS and Dietary Factor | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Over- weight pregnant women [111] | 88 | P | Dietary fiber vs. | diversity, richness, Firmicutes in unidentified family of order Clostridiales, Barnciellaceae family belonging to the phylum Bacteroidetes | P | none | P | none | ||
| Vitamin A, β-Carotene vs. | Firmicutes | |||||||||
| N | Fat vs. | diversity, richness, Barnsiellaceae | N | none | N | none | ||||
| Healthy subjects [86] | 150 | N | 25-Hydroxy vitamin D vs. | Coprococcus, Bifdobacterium | N | LPS vs. | Faecalibacterium | N | LPS vs. | 25-Hydroxy vitamin D |
| Type 1 diabetes [105] | 668 | — | — | — | — | — | — | N | LPS vs. | Dietary pattern; “Fish”(frequently eat fish dishes), “Healthy snack” (frequently eat fruits, berries, fresh vegetable, yoghurt, low-fat cheese, and do not drink much soft drinks), “Modern”(frequently eat poultry, pasta, rice, meat dishes, fried and grilled foods, and fresh vegetables) |
| Category | Dietary Factor | Dose | Administration Period | Model | LPS | LBP | Significant Change in Gut Microbiota |
|---|---|---|---|---|---|---|---|
| Probiotics/ Prebiotics | Lactobacillus rhamnosus GG [51] | 1 × 108 CFU/day | 12 weeks | HFD-fed ApoE KO mouse | ↓ | — | no |
| Lactobacillus rhamnosus CNCM I-4036 [52] | 1 × 1010 CFU/day | 30 days | Chow diet-fed Zucker-Lepfa/fa rat | — | ↓ | — | |
| Lactobacillus sakei OK67 +/− Lactobacillus sakei PK16 [53] | 1 × 109 CFU/day 1 × 109 CFU/day | 4 weeks | HFD-fed C57BL/6 mouse | ↓ | — | yes | |
| Bifidobacterium longum BR-108 (sterilized) [54] | 200, 400 mg/kg/day | 4 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Bifidobacterium infantis + Lactobacillus acidophilus + Bacillus cereus [55] | 0.5 × 106 CFU/day 0.5 × 106 CFU/day 0.5 × 105 CFU/day | 12 weeks | HFHSD-fed SD rat | ↓ | — | yes | |
| Lactobacillus plantarum LC27 +/− Bifidobacterium longum LC67 [43] | 1 × 109 CFU/day each (or 0.75 × 109 (LC27) + 0.25×109 (LC67) CFU/day in mix) | 4 weeks | HFD-fed C57BL/6 mouse | ↓ | — | yes | |
| Oligofructose [112] | 10% (mixed in diet) | 12 weeks | HFHSD-fed SD rat | ↓ | — | yes | |
| Galacto- oligosaccharide [84] | 800 mg/kg/day | 8 weeks | HFD-fed SD rat | ↓ | — | yes | |
| Inulin [113] | 5% (intragastric administration, sample volume was not described) | 6 weeks | standardized diet (kcal %: 10% fat, 20% protein, and 70% carbo- hydrate; 3.85 kcal g−1)-fed db/db mouse | ↓ | — | yes | |
| Wheat-derived arabinoxylan [114] | 7.5% (mixed in diet) | 8 weeks | HFD-fed C57BL/6J mouse | ↓ | — | — | |
| Polyphenols | Grape seed proanthocyanidin [33] | 500 mg/kg/day | 10 days (prophylactic) or 17 weeks (with cafeteria diet) | Cafeteria diet (high-fat/high carbohydrate diet)-fed Wistar rat | ↓ | — | — |
| Grape-seed proanthocyanidin [29] | 100, 500 mg/kg/day | 2 weeks | Cafeteria diet (high saturated-fat/high refined-carbohydrate diet)-fed Wistar rat | ↓ | — | — | |
| Resveratrol [74] | 50, 75, 100 mg/kg/day | 16 weeks | HFD-fed C57BL/6 mouse | ↓ | ↓ | yes | |
| Apple-derived polymeric procyanidins [75] | 0.5% (administration route was not described) | 20 weeks | HFHSD-fed C57BL/6J mouse | ↓ | — | yes | |
| Genistein [76] | 0.2% (mixed in diet) | 6 months | HFD-fed C57BL/6 mouse | ↓ | — | yes | |
| Isoflavone [77] | 0.1% (mixed in diet) | 5 weeks | HFD-fed C57BL/6 mouse | ↓ | ↓ | yes | |
| Syringaresinol [78] | 50 mg/kg/day | 10 weeks | 40-week-old C57BL/6 mouse | — | ↓ | yes | |
| Sulfated polysaccharide | Sea cucumber-derived sulfated polysaccharide [80] | 300 mg/kg/day | 8 weeks | HFD-fed BALB/c mouse | — | ↓ | yes |
| Sea cucumber-derived sulfated polysaccharide [81] | 300 mg/kg/day | 42 days | Chow-fed BALB/c mouse | — | ↓ | yes | |
| Acaudina molpadioides-derived fucosylated chondroitin sulfate [82] | 80 mg/kg/day | 10 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Chicken-derived chondroitin sulfate [83] | 150 mg/kg/day | 16 days | Exhaustive exercise stress model BALB/c mouse | ↓ | — | yes | |
| Fucoidan [84] | 100 mg/kg/day | 8 weeks | HFD-fed SD rat | ↓ | — | yes | |
| Other dietary components | Tetrahydro iso-alpha acid (included in hops) [88] | 0.1% (mixed in diet) | 8 weeks | HFD-fed C57BL/6J mouse | ↓ | — | — |
| Rhein (included in rhubarb) [89] | 120 mg/kg/day | 6 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Phlorizin (included in apple) [90] | 20 mg/kg/day | 10 weeks | Chow-fed db/db mouse | ↓ | — | yes | |
| Capsaicin [91] | 0.01% (mixed in diet) | 12 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Rutin [92] | 0.64% (mixed in diet) | 20 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Lycopene [93] | 0.03% (mixed in diet) | 10 weeks | HFD and fructose-fed C57BL/6 J mouse | ↓ | — | — | |
| Other extracts/dietary components | Broccoli sprout extract [94] | 2.2% (mixed in diet) | 14 weeks | HFD-fed C57BL/6JSlc mouse | ↓ | ↓ | yes |
| Camu camu extract [96] | 200 mg/kg/day | 8 weeks | HFHSD-fed C57BL/6J mouse | ↓ | — | yes | |
| Other extracts/dietary components | Cranberry extract [95] | 200 mg/kg/day | 8 weeks | HFHSD-fed C57BL/6J mouse | ↓ | — | yes |
| Green tea extract [34] | 2% (mixed in diet) | 8 weeks | HFD-fed C57BL/6J mouse | ↓ | — | yes | |
| Tartary buckwheat protein [32] | 23.5% (mixed in diet) | 6 weeks | HFD-fed C57BL/6 mouse | ↓ | — | yes | |
| Foods | Cocoa [97] | 8% (mixed in diet) | 18 weeks | HFD-fed C57BL/6J mouse | ↓ | — | — |
| Nopal [98] | 5% of dietary fiber was replaced with those of nopal-derived (mixed in diet) | 1 month | HFHSD-fed Wistar rat | ↓ | — | yes | |
| Steamed fish meat [99] | Ad libitum (9:00–12:00 and 18:00–21:00) | 8 weeks | Chow-fed C57BL/6 mouse | — | ↓ | yes | |
| Chinese medicines | Geniposide + Chlorogenic acid [100] | 90 mg/kg/day 1.34 mg/kg/day | 4 weeks | HFD-fed C57BL/6 mouse | — | ↓ | — |
| Potentilla discolor Bunge water extract [101] | 400 mg/kg/day | 8 weeks | HFD-fed, streptozotocin- injected C57BL/6J mouse | ↓ | ↓ | yes | |
| Ganoderma lucidum mycelium water extract [102] | 2–8 mg/day | 8 weeks | HFD-fed C57BL/6NCrlBltw mouse | ↓ | — | yes | |
| Semen hoveniae extract [103] | 300, 600 mg/kg/day | 8 weeks | Alcohol-containing Lieber-DeCarli diet-fed SD rat (Alcoholic liver disorder model) | ↓ | — | yes | |
| Shenling Baizhu powder [104] | 30 g/kg/day | 16 weeks | HFD-fed SD rat | ↓ | — | yes | |
| Caloric restriction | 30% caloric restriction [108] | — | 62–141 weeks | HFD, LFD-fed C57BL/6J mouse | — | ↓ | yes |
| 40% caloric restriction [109] | — | 30 days | Chow-fed C57BL/6J mouse | ↓ | ↓ | yes |
| Category | Dietary Factor | Sample | Method | Gut Microbe with Significant Changes in Proportion * | |
|---|---|---|---|---|---|
| Increase | Decrease | ||||
| Probiotics/ Prebiotics | Lactobacillus sakei OK67 +/− Lactobacillus sakei PK16 [53] | Feces | PCR, NGS | OTU (O67), Ace (O67), Chao1 (O67), Shanon (O67) | Simpson (O67), Proteobacteria, Firmicutes, Firmicutes/Bacteroidetes, Proteobacteria/Bacteroidetes |
| Bifidobacterium longum BR-108 (sterilized) [54] | Cecal contents | PCR | Bifidobacterium spp., Lactobacillus spp. | Firmicutes | |
| Bifidobacterium infantis + Lactobacillus acidophilus + Bacillus cereus [55] | Feces | PCR | Bifidobacteria, Lactobacillus, Bacteroides, Bifidobacteria/Escherichia coli | Escherichia coli, Enterococcus | |
| Lactobacillus plantarum LC27 +/− Bifidobacterium longum LC67 [43] | Feces | PCR | Actinobacteria (LC67, LC27 + LC67) | Firmicutes, Bacteroidetes, δ/γ-Proteobacteria, Deferribacteres (LC67, LC27+LC67), Firmicutes/Bacteroidetes, Proteobacteria/Bacteroidetes | |
| Oligofructose [112] | Cecal contents | PCR | Bacteroides/Prevotella, Bifidobacterium, Lactobacillus, Roseburia | Clostridium leptum (cluster IV), Clostridium cluster I, Clostridium cluster XI, Methanobrevibacter, Akkemansia muciniphila, Faecalibacterium prausnitzii | |
| Galacto-oligosaccharide [84] | Cecal contents | NGS | Verrucomicrobia, Akkermansia, Ruminococcus, Blautia, Bacteroidetes, Proteobacteria, Adlercreutzia, Staphylococcus, Prevotella, Oscillospira, Lactobacillus, Desulfovibrio | Firmicutes, Actinobacteria, Clostridium, Bacillus | |
| Inulin [113] | Feces | NGS | Bacteroidetes, Cyanobacteria, Bacteroides | Firmicutes, Deferribacteres, Tenericutes, Ruminiclostridium_6, Mucispirillum | |
| Polyphenols | Resveratrol [74] | Cecal contents | NGS | Deferribacteraceae | none (In this study, population of Desulfovibrionaceae in the high-fat diet + intervention group was at the same level with normal chow group, but there was no significant reduction from high-fat diet group.) |
| Apple-derived polymeric procyanidins [75] | Cecal contents | NGS | Bacteroidetes, Verrucomicrobia, Adlerceitzia, Roseburia, S24-7, Bacteroids, Anaerovorax, rc4-4, Akkermansia | Firmicutes, Firmicutes/Bacteroidetes, Clostridium, Lachnospiraceae, Bifidobacterium | |
| Polyphenols | Genistein [76] | Feces | NGS | Firmicutes, Verrucomicrobia, Prevotellaceae, Verrucomicrobia, Prevotella, Akkermansia, Faecalibacterium, Prevotella copri, Prevotella stercorea, Akkermansia muciniphila | Bacteroidetes, Bacteroidaceae, Bacteroides, Bacteroides acidifaciens, Bacteroides uniformis |
| Isoflavone [77] | Feces | NGS | α-diversity, Actinobacteria, Verrucomicrobia, Bifidobacterium/Enterobacteriaceae, Akkermansia | Proteobacteria | |
| Syringaresinol [78] | Cecal contents | NGS | Firmicutes/Bacteroidetes, Firmicutes, Lactobacillus, Lactobacillus animalis, Lactobacillus johnsonii, Lactobacillus reuteri, Lactobacillus intestinalis, Bifidobacterium pseudolongum | Shannon diversity indices, Jeotgalicoccus nanhaiensis, Staphylococcus lentus, Bacteroidaceae (EF098405_s), Bacteroides vulgatus, Akkermansia muciniphila | |
| Sulfated polysaccharide | Sea cucumber-derived sulfated polysaccharide [80] | Feces | NGS | bacterial diversity, Verrucomicrobia (depolymerized sulfated polysaccharide), Bacteroides, Alloprevotella, Ruminiclostridium_9, Butyricicoccus, Akkermansia | Proteobacteria, Escherichia-Shigella (polymerized sulfated polysaccharide), Pseudomonas (depolymerized sulfated polysaccharide), Yersinia (depolymerized sulfated polysaccharide), (In this study, decrease of Desulfovibrio with the intervention of sulfated polysaccharide to high-fat diet-fed mouse was shown as heatmap, but significance of difference was not described.) |
| Sea cucumber-derived sulfated polysaccharide [81] | Feces | NGS | Proteobacteria (polymerized sulfated polysaccharide), Bacteroides (polymerized sulfated polysaccharide), Allobaculum (depolymerized sulfated polysaccharide), Alloprevotella, Roseburia, Turicibacter, Desulfovibrio | Enterococcus, Streptococcus, Escherichia-Shigella, Lactobacillus | |
| Acaudina molpadioides-derived fucosylated chondroitin sulfate [82] | Feces | PCR, NGS | Bacteroidetes, Lactobacillus, Actinobacteria, Faecalibacterium prausnitzii, Deferribacteres, Bacteroidales, Bifidobacteriales, Lachnospiraceae NK4A136 group, Bacteroides, Bacteroides acidifaciens, Bifidobacterium choerinum | Firmicutes, Escherichia coli, Clostridiales, Bacilli, Lactobacillales, Clostridia Clostridiales, Firmicutes Clostridiales, Lactococcus, Clostridium ruminantium | |
| Chicken-derived chondroitin sulfate [83] | Feces | NGS | Bacteroidetes, Bacteroides acidifaciens, family S24-7, Lysinibacillus boronitolerans | Firmicutes, β-Proteobacteria | |
| Fucoidan [84] | Cecal contents | NGS | Proteobacteria, Verrucomicrobia, Enterobacter, Bacteroidetes, Bacillus, Ruminococcus, Adlercreutzia, Prevotella, Oscillospira, Desulfovibrio, | Firmicutes, Actinobacteria, Clostridium, Corynebacterium, Staphylococcus, Lactobacillus, Aerococcus | |
| Other dietary components | Rhein (included in rhubarb) [89] | Cecal contents | PCR | Bacteroides/Prevotella, Desulfovibrio | Bifidobacterium, Lactobacillus |
| Phlorizin (included in apple) [90] | Feces | PCR, DGGE | Akkermansia muciniphila, Prevotella | none | |
| Capsaicin [91] | Cecal contents | NGS | Ruminococcaceae, Lachnospiraceae | family S24_7 | |
| Rutin [92] | Small intestinal contents | NGS | Bacteroidales_S24-7 group, Bacteroidaceae, Porphyromonadaceae, Rikenellaceae, Desulfovibrionaceae | Firmicutes, Firmicutes/Bacteroidetes, Deferribacteraceae, Lachnospiraceae | |
| Other extracts/dietary components | Broccoli sprout extract [94] | Cecal contents | NGS | none | Proteobacteria, Desulfovibrionaceae |
| Camu camu extract [96] | Feces | NGS | microbial richness, Bifidobacterium, Barnesiella, Barnesiella spp., Turicibacter spp., Akkermansia muciniphila, Delftia, Roseburia, Anaerostipes, unclassified genera within the families Christensenellaceae, unclassified genera within the families Erysipelotrichaceae | Firmicutes/Bacteroidetes, Lactobacillus, Anaerotruncus, Parabacteroides | |
| Cranberry extract [95] | Feces | PCR, NGS | Akkermansia | none | |
| Green tea extract [34] | Cecal contents | NGS | Shannon index, Chao1 richness, Bacteroidetes, Actinobacteria, Verrucomicrobia, Bacteroidales, Bifidobacteriales, Verrucomicrobiales, Turicibacterales. RF39, Coriobacteriales, Bifidobacterium, Blautia, Dorea, Lactobacillus, Ruminococcus, Akkermansia, Butyrivibrio, Akkermansia muciniphila, Ruminococcus gnavus, Bifidobacterium pseudolongum, Bifidobacterium adolescentis | Firmicutes, Firmicutes/Bacteroidetes, Clostridiales, SMB53 | |
| Tartary buckwheat protein [32] | Feces | PCR | Bifidobacterium, Lactobacillus, Enterococcus, Clostridium | Escherichia coli, Bacaeroides | |
| Foods | Nopal [98] | Feces | NGS | α-diversity, Anaeroplasma, Prevotella, Ruminucoccus, Bacteroides fragilis, Ruminococcus bromii, Rumminococcus flavefaciens, Lactobacillus reuteri, Akkermansia muciniphila | Firmicutes/Bacteroidetes, Faecalibacterium, Clostridium, Butyricicoccus, Bacteroides acidifaciens, Blautia producta, Faecalibacterium prausnitzii, Butyricicoccus pullicaecorum, Clostridium citroniae |
| Steamed fish meat [99] | Feces | NGS | Proteobacteria, Firmicutes, Ruminococcaceae, Oscillospira, Clostridium, Escherichia | Shannon index, Bacteroidetes, S24-7 | |
| Chinese medicines | Potentilla discolor Bunge water extract [101] | Feces | NGS | Bacteroidetes, Bacteroidales_S24-7_group, norank_f_Bacteroidales_S24-7_group, Parabacteroides, Eubacterium_nodatum_group, norank_f_Rhodospirillaceae, Tyzzerella, Rikenella, Alistipes, Lachnospiraceae_NK4A136_group, norank_f_Ruminococcaceae, Romboutsia, Coriobacteriaceae_UCG_002, Bacteroides, Allobaculum, Coprococcus_3, norank_f_Christensenellaceae | Proteobacteria, Helicobacteraceae, Helicobacter |
| Ganoderma lucidum mycelium water extract [102] | Cecal contents | NGS | Parabacteroides goldsteinii, Bacteroides spp., Anaerotruncus colihominis, Roseburia hominis, Clostridium methylpentosum (Clostridium IV), Clostridium XIVa, Clostridium XVIII, Eubacterium coprostanoligenes | Firmicutes/Bacteroidetes, Proteobacteria, Mucispirilum shaedleri, Escherichia fergusonii, Enterococcus spp., Lactococcus lactis, Clostridium lactatifermentans (Clostridium XIVb), Oscillibacter valericigenes | |
| Semen hoveniae extract [103] | Feces | NGS | Shannon index, Verrucomicrobia, Bacteroidetes, Parabacteroides, Alloprevotella, Alistipes, Lactobacillus, Akkermansia | Proteobacteria, Firmicutes/Bacteroidetes, Oscillibacter, Helicobacter | |
| Chinese medicines | Shenling Baizhu powder [104] | Feces | NGS | Shannon index, Actinobacteria, Cyanobacteria, Anaerostipes, Bifidobacterium | Firmicutes/Bacteroidetes, Blautia, Roseburia, Phascolarctobacterium, Desulfovibrio (Significance of difference was not described) |
| Caloric restriction | 30% caloric restriction [108] | Feces | NGS | (low-fat diet vs. low-fat diet with caloric restriction) Lactobacillus, OTU45 (in Lactobacillus), Bifidobacterium, [increased by caloric restriction with both of low-fat diet or high-fat diet] OTU119, OTU155, OTU267 (in Tannerella) | (low-fat diet vs. low-fat diet with caloric restriction) Streptococcaceae, TM7, OTU469 (in Desulfovibrionaceae) [decreased by caloric restriction with both of low-fat diet or high-fat diet] OTU65 (in Lactococcus), OTU366 (in Bacteroidales), OTU37 (in Peptostreptococcaceae), |
| 40% caloric restriction [109] | Feces | NGS | Lactobacillaceae, Erysipelotichaceae, Bacteroidaceae, Verrucomicrobiaceae | Firmicutes | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuke, N.; Nagata, N.; Suganuma, H.; Ota, T. Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors. Nutrients 2019, 11, 2277. https://doi.org/10.3390/nu11102277
Fuke N, Nagata N, Suganuma H, Ota T. Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors. Nutrients. 2019; 11(10):2277. https://doi.org/10.3390/nu11102277
Chicago/Turabian StyleFuke, Nobuo, Naoto Nagata, Hiroyuki Suganuma, and Tsuguhito Ota. 2019. "Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors" Nutrients 11, no. 10: 2277. https://doi.org/10.3390/nu11102277
APA StyleFuke, N., Nagata, N., Suganuma, H., & Ota, T. (2019). Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors. Nutrients, 11(10), 2277. https://doi.org/10.3390/nu11102277