Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity


Abstract
1. Introduction
2. Human Metabolism of Resveratrol
2.1. RSV is Absorbed and Metabolized in Target Tissues
2.2. The Gut Microbiome Metabolizes RSV and RSV Influences Gut Microbial Composition
2.3. Biotransformation of RSV Limits Plasma Bioavailability
3. Molecular Action of Resveratrol and Metabolites
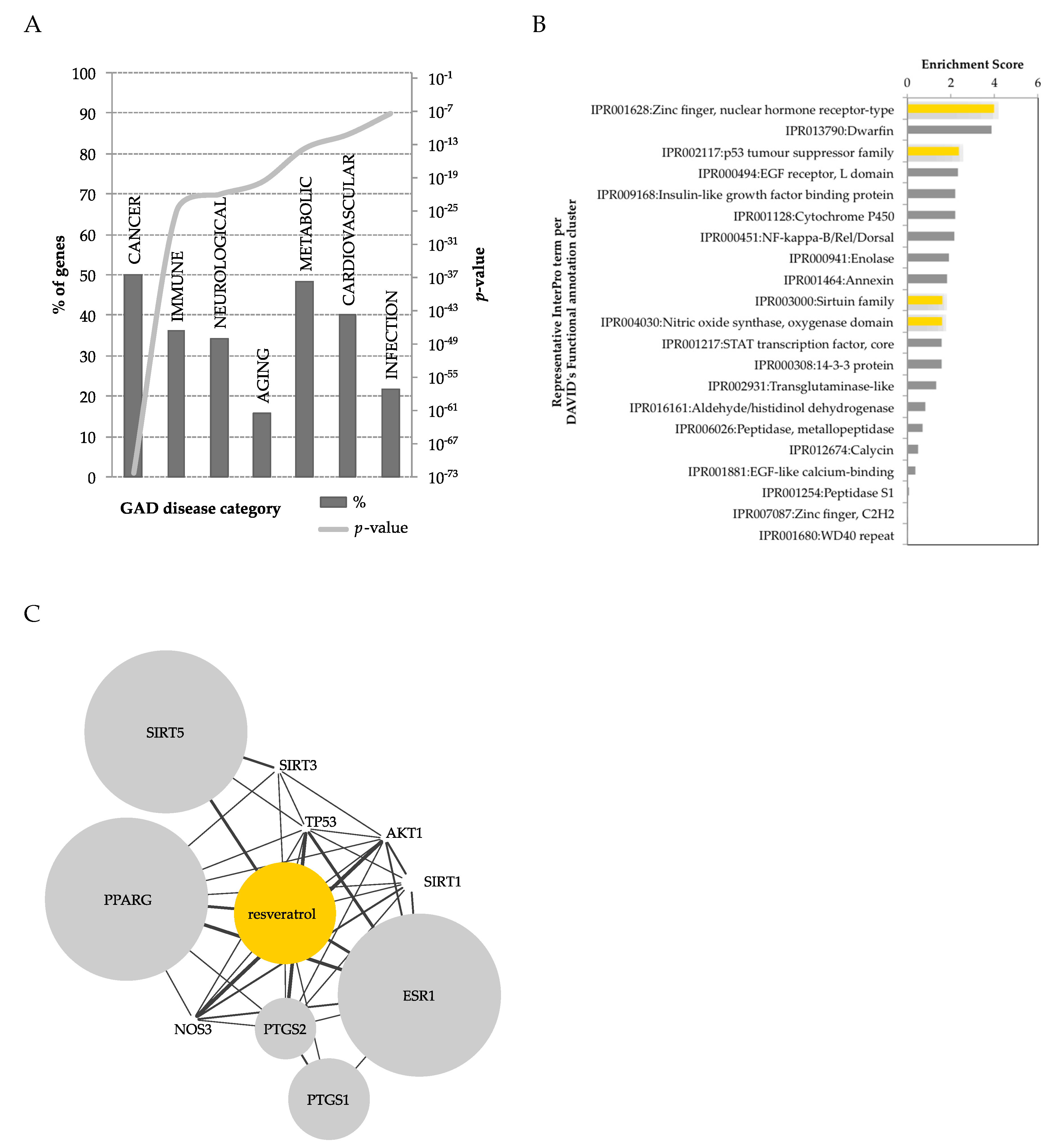
3.1. RSV Modulates a Panoply of Protein Targets
3.2. RSV Increases Energy Expenditure and Vascular Function
3.3. Effect of RSV on Epigenietics
3.4. RSV Influences Redox Metabolism
3.5. RSV Metabolites Exhibit Activity
4. Resveratrol, Metabolic Health and Obesity
5. Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Takaoka, M. The Phenolic Substances of White Hellebore (Veratrum Grandiflorum Hoes. Fil.) I. Nippon Kagaku Kaishi 1939, 60, 1090–1100. [Google Scholar] [CrossRef]
- Siemann, E.H.; Creasy, L.L. Concentration Of The Phytoalexin Resveratrol In Wine. Am. J. Enol. Vitic. 1992, 43, 49–52. [Google Scholar]
- St Leger, A.S.; Cochrane, A.; Moore, F. Factors Associated With Cardiac Mortality In Developed Countries With Particular Reference To The Consumption Of Wine. Lancet 1979, 313, 1017–1020. [Google Scholar] [CrossRef]
- Renaud, S.; De Lorgeril, M. Wine, Alcohol, Platelets, And The French Paradox For Coronary Heart Disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Frankel, E.N.; Waterhouse, A.L.; Kinsella, J.E. Inhibition Of Human Ldl Oxidation By Resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer Chemopreventive Activity Of Resveratrol, A Natural Product Derived From Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Chung, W.J.; Michaluart, P.; Telang, N.; Tanabe, T.; Inoue, H.; Jang, M.; Pezzuto, J.M.; Dannenberg, A.J. Resveratrol Inhibits Cyclooxygenase-2 Transcription And Activity In Phorbol Ester-Treated Human Mammary Epithelial Cells. J. Biol. Chem. 1998, 273, 21875–21882. [Google Scholar] [CrossRef]
- Wallerath, T.; Deckert, G.; Ternes, T.; Anderson, H.; Li, H.; Witte, K.; Förstermann, U. Resveratrol, A Polyphenolic Phytoalexin Present In Red Wine, Enhances Expression And Activity Of Endothelial Nitric Oxide Synthase. Circulation 2002, 106, 1652–1658. [Google Scholar] [CrossRef]
- Park, S.-J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol Ameliorates Aging-Related Metabolic Phenotypes By Inhibiting Camp Phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small Molecule Activators Of Sirtuins Extend Saccharomyces Cerevisiae Lifespan. Nature 2014, 19, 191–196. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol Improves Health And Survival Of Mice On A High-Calorie Diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; Van De Weijer, T.; Goossens, G.H.; Hoeks, J.; Van Der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie Restriction-Like Effects Of 30 Days Of Resveratrol Supplementation On Energy Metabolism And Metabolic Profile In Obese Humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function And Protects Against Metabolic Disease By Activating Sirt1 And Pgc-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential Of Resveratrol: The In Vivo Evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Timmers, S.; De Ligt, M.; Phielix, E.; Van De Weijer, T.; Hansen, J.; Moonen-Kornips, E.; Schaart, G.; Kunz, I.; Hesselink, M.K.C.; Schrauwen-Hinderling, V.B.; et al. Resveratrol As Add-On Therapy In Subjects With Well-Controlled Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2016, 39, 2211–2217. [Google Scholar] [CrossRef] [PubMed]
- Walle, T.; Hsieh, F.; Delegge, M.H.; John, E.; Oatis, J.; Walle, U.K. High Absorption But Very Low Bioavailability Of Oral Resveratrol In Humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Geun Shin, Y.; Chow, A.; Li, Y.; Kosmeder, J.W.; Sup Lee, Y.; Hirschelman, W.H.; Pezzuto, J.M.; Mehta, R.G.; Van Breemen, R.B. Human, Rat, And Mouse Metabolism Of Resveratrol. Pharm. Res. 2002, 19, 1907–1914. [Google Scholar] [CrossRef]
- Böhmdorfer, M.; Szakmary, A.; Schiestl, R.; Vaquero, J.; Riha, J.; Brenner, S.; Thalhammer, T.; Szekeres, T.; Jäger, W. Involvement Of Udp-Glucuronosyltransferases And Sulfotransferases In The Excretion And Tissue Distribution Of Resveratrol In Mice. Nutrients 2017, 9, 1347. [Google Scholar] [CrossRef]
- Manach, C.; Milenkovic, D.; Van De Wiele, T.; Rodriguez-Mateos, A.; De Roos, B.; Garcia-Conesa, M.T.; Landberg, R.; Gibney, E.R.; Heinonen, M.; Tomás-Barberán, F.; et al. Addressing The Inter-Individual Variation In Response To Consumption Of Plant Food Bioactives: Towards A Better Understanding Of Their Role In Healthy Aging And Cardiometabolic Risk Reduction. Mol. Nutr. Food Res. 2017, 61, 1600557. [Google Scholar] [CrossRef]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In Vivo And In Vitro Metabolism Of Trans-Resveratrol By Human Gut Microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions Of Gut Microbiota With Dietary Polyphenols And Consequences To Human Health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.K. Intestinal Ugts As Potential Modifiers Of Pharmacokinetics And Biological Responses To Drugs And Xenobiotics. Expert Opin. Drug Metab. Toxicol. 2007, 3, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-H.; Wang, C.-C.; Hou, Y.-H.; Mao, Y.-C.; Yang, Y.-S. Mechanism Of Sulfotransferase Pharmacogenetics In Altered Xenobiotic Metabolism. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1053–1071. [Google Scholar] [CrossRef] [PubMed]
- Lamba, J.K.; Lin, Y.S.; Schuetz, E.G.; Thummel, K.E. Genetic Contribution To Variable Human Cyp3a-Mediated Metabolism. Adv. Drug Deliv. Rev. 2012, 64, 256–269. [Google Scholar] [CrossRef]
- Mena, P.; Del Rio, D. Gold Standards For Realistic (Poly)Phenol Research. J. Agric. Food Chem. 2018, 66, 8221–8223. [Google Scholar] [CrossRef] [PubMed]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; Mcpherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The Global Obesity Pandemic: Shaped By Global Drivers And Local Environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Cypess, A.M.; Kahn, C.R. Cellular Bioenergetics As A Target For Obesity Therapy. Nat. Rev. Drug Discov. 2010, 9, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Gheldof, N.; Moco, S.; Chabert, C.; Teav, T.; Barron, D.; Hager, J. Role Of Sulfotransferases In Resveratrol Metabolism In Human Adipocytes. Mol. Nutr. Food Res. 2017, 61, 1700020. [Google Scholar] [CrossRef]
- Jimenez-Gomez, Y.; Mattison, J.A.; Pearson, K.J.; Martin-Montalvo, A.; Palacios, H.H.; Sossong, A.M.; Ward, T.M.; Younts, C.M.; Lewis, K.; Allard, J.S.; et al. Resveratrol Improves Adipose Insulin Signaling And Reduces The Inflammatory Response In Adipose Tissue Of Rhesus Monkeys On High-Fat, High-Sugar Diet. Cell Metab. 2013, 18, 533–545. [Google Scholar] [CrossRef]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.J.; Crozier, A. Plant Foods And Herbal Sources Of Resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol: Twenty Years Of Growth, Development And Controversy. Biomol. Ther. 2018, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Planas, J.M.; Alfaras, I.; Colom, H.; Juan, M.E. The Bioavailability And Distribution Of Trans-Resveratrol Are Constrained By Abc Transporters. Arch. Biochem. Biophys. 2012, 527, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.; Vitrac, X.; Decendit, A.; Ennamany, R.; Krisa, S.; Mérillon, J.-M. Cellular Uptake And Efflux Of Trans -Piceid And Its Aglycone Trans -Resveratrol On The Apical Membrane Of Human Intestinal Caco-2 Cells. J. Agric. Food Chem. 2005, 53, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W.H. Quantitative Evaluation Of The Expression And Activity Of Five Major Sulfotransferases (Sults) In Human Tissues: The Sult “Pie”. Drug Metab. Dispos. 2009, 37, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Ung, D.; Nagar, S. Variable Sulfation Of Dietary Polyphenols By Recombinant Human Sulfotransferase (Sult) 1a1 Genetic Variants And Sult1e1. Drug Metab. Dispos. 2007, 35, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Miksits, M.; Maier-Salamon, A.; Aust, S.; Thalhammer, T.; Reznicek, G.; Kunert, O.; Haslinger, E.; Szekeres, T.; Jaeger, W. Sulfation Of Resveratrol In Human Liver: Evidence Of A Major Role For The Sulfotransferases Sult1a1 And Sult1e1. Xenobiotica 2005, 35, 1101–1119. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.; Fukami, T.; Yokoi, T.; Nakajima, M. A Comprehensive Review Of Udp-Glucuronosyltransferase And Esterases For Drug Development. Drug Metab. Pharmacokinet. 2015, 30, 30–51. [Google Scholar] [CrossRef]
- Rouleau, M.M.; Audet-Delage, Y.; Desjardins, S.; Rouleau, M.M.; Girard-Bock, C.; Guillemette, C. Endogenous Protein Interactome Of Human Udp-Glucuronosyltransferases Exposed By Untargeted Proteomics. Front. Pharmacol. 2017, 8, 23. [Google Scholar] [CrossRef]
- Dietrich, C.G.; Geier, A.; Oude Elferink, R.P.J. Abc Of Oral Bioavailability: Transporters As Gatekeepers In The Gut. Gut 2003, 52, 1788–1795. [Google Scholar] [CrossRef]
- Maier-Salamon, A.; Böhmdorfer, M.; Riha, J.; Thalhammer, T.; Szekeres, T.; Jaeger, W. Interplay Between Metabolism And Transport Of Resveratrol. Ann. N. Y. Acad. Sci. 2013, 1290, 98–106. [Google Scholar] [CrossRef]
- Vitaglione, P.; Sforza, S.; Galaverna, G.; Ghidini, C.; Caporaso, N.; Vescovi, P.P.; Fogliano, V.; Marchelli, R. Bioavailability Of Trans-Resveratrol From Red Wine In Humans. Mol. Nutr. Food Res. 2005, 49, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical Trials Of Resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Marier, J.-F. Metabolism And Disposition Of Resveratrol In Rats: Extent Of Absorption, Glucuronidation, And Enterohepatic Recirculation Evidenced By A Linked-Rat Model. J. Pharmacol. Exp. Ther. 2002, 302, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Asensi, M.; Medina, I.; Ortega, A.; Carretero, J.; Baño, M.C.; Obrador, E.; Estrela, J.M. Inhibition Of Cancer Growth By Resveratrol Is Related To Its Low Bioavailability. Free Radic. Biol. Med. 2002, 33, 387–398. [Google Scholar] [CrossRef]
- Emília Juan, M.; Buenafuente, J.; Casals, I.; Planas, J.M. Plasmatic Levels Of Trans-Resveratrol In Rats. Food Res. Int. 2002, 35, 195–199. [Google Scholar] [CrossRef]
- Menet, M.-C.; Baron, S.; Taghi, M.; Diestra, R.; Dargère, D.; Laprévote, O.; Nivet-Antoine, V.; Beaudeux, J.-L.; Bédarida, T.; Cottart, C.-H. Distribution Of Trans -Resveratrol And Its Metabolites After Acute Or Sustained Administration In Mouse Heart, Brain, And Liver. Mol. Nutr. Food Res. 2017, 61, 1600686. [Google Scholar] [CrossRef] [PubMed]
- Andres-Lacueva, C.; Macarulla, M.T.; Rotches-Ribalta, M.; Boto-Ordóñez, M.; Urpi-Sarda, M.; Rodríguez, V.M.; Portillo, M.P. Distribution Of Resveratrol Metabolites In Liver, Adipose Tissue, And Skeletal Muscle In Rats Fed Different Doses Of This Polyphenol. J. Agric. Food Chem. 2012, 60, 4833–4840. [Google Scholar] [CrossRef]
- Alberdi, G.; Rodríguez, V.M.; Miranda, J.; Macarulla, M.T.; Arias, N.; Andrés-Lacueva, C.; Portillo, M.P. Changes In White Adipose Tissue Metabolism Induced By Resveratrol In Rats. Nutr. Metab. 2011, 8, 29. [Google Scholar] [CrossRef]
- Burkon, A.; Somoza, V. Quantification Of Free And Protein-Boundtrans-Resveratrol Metabolites And Identification Oftrans-Resveratrol-C/O-Conjugated Diglucuronides—Two Novel Resveratrol Metabolites In Human Plasma. Mol. Nutr. Food Res. 2008, 52, 549–557. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction Between Phenolics And Gut Microbiota: Role In Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Dao, T.-M.A.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D.J.; Champion, S.; Barthélemy, S.; Barra, Y.; et al. Resveratrol Increases Glucose Induced Glp-1 Secretion In Mice: A Mechanism Which Contributes To The Glycemic Control. PLoS ONE 2011, 6, E20700. [Google Scholar] [CrossRef]
- Larrosa, M.; Gonza, A.; Toti, S.; Joaqui, J.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Cerón, J.J.; Tomás-Barberán, F.; et al. Effect Of A Low Dose Of Dietary Resveratrol On Colon Microbiota, Inflammation And Tissue Damage In A Dss-Induced Colitis Rat Model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Lim, Y.-H. Resveratrol Antibacterial Activity Against Escherichia Coli Is Mediated By Z-Ring Formation Inhibition Via Suppression Of Ftsz Expression. Sci. Rep. 2015, 5, 10029. [Google Scholar] [CrossRef]
- Paulo, L.; Ferreira, S.; Gallardo, E.; Queiroz, J.A.; Domingues, F. Antimicrobial Activity And Effects Of Resveratrol On Human Pathogenic Bacteria. World J. Microbiol. Biotechnol. 2010, 26, 1533–1538. [Google Scholar] [CrossRef]
- Sung, M.M.; Kim, T.T.; Denou, E.; Soltys, C.-L.M.; Hamza, S.M.; Byrne, N.J.; Masson, G.; Park, H.; Wishart, D.S.; Madsen, K.L.; et al. Improved Glucose Homeostasis In Obese Mice Treated With Resveratrol Is Associated With Alterations In The Gut Microbiome. Diabetes 2017, 66, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity Alters Gut Microbial Ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Etxeberria, U.; Arias, N.; Boqué, N.; Macarulla, M.T.; Portillo, M.P.; Martínez, J.A.; Milagro, F.I. Reshaping Faecal Gut Microbiota Composition By The Intake Of. J. Nutr. Biochem. 2015, 26, 1–26. [Google Scholar] [CrossRef]
- Bird, J.K.; Raederstorff, D.; Weber, P.; Steinert, R.E. Cardiovascular And Antiobesity Effects Of Resveratrol Mediated Through The Gut Microbiota. Adv. Nutr. 2017, 8, 839–849. [Google Scholar] [CrossRef]
- Most, J.; Penders, J.; Lucchesi, M.; Goossens, G.H.; Blaak, E.E. Gut Microbiota Composition In Relation To The Metabolic Response To 12-Week Combined Polyphenol Supplementation In Overweight Men And Women. Eur. J. Clin. Nutr. 2017, 71, 1040–1045. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal Microbiota Metabolism Of L-Carnitine, A Nutrient In Red Meat, Promotes Atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Chen, M.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.; Zhang, Q.; Mi, M. Atherosclerosis By Regulating Tmao Synthesis And Bile Acid Metabolism Via Remodeling Of The Gut Microbiota. Mbio 2016, 7, E02210–E02215. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.E.; González-Pons, E.; Planas, J.M. Multidrug Resistance Proteins Restrain The Intestinal Absorption Of Trans-Resveratrol In Rats. J. Nutr. 2010, 140, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Andreadi, C.; Britton, R.G.; Horner-Glister, E.; Karmokar, A.; Sale, S.; Brown, V.A.; Brenner, D.E.; Singh, R.; Steward, W.P.; et al. Sulfate Metabolites Provide An Intracellular Pool For Resveratrol Generation And Induce Autophagy With Senescence. Sci. Transl. Med. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J. Resveratrol Bioavailability And Toxicity In Humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Sergides, C.; Chirilă, M.; Silvestro, L.; Pitta, D.; Pittas, A. Bioavailability And Safety Study Of Resveratrol 500 Mg Tablets In Healthy Male And Female Volunteers. Exp. Ther. Med. 2016, 11, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Novotny, J.A.; Chen, T.-Y.; Terekhov, A.I.; Gebauer, S.K.; Baer, D.J.; Ho, L.; Pasinetti, G.M.; Ferruzzi, M.G. The Effect Of Obesity And Repeated Exposure On Pharmacokinetic Response To Grape Polyphenols In Humans. Mol. Nutr. Food Res. 2017, 61, 1700043. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Haskell-Ramsay, C.F.; Reay, J.L.; Williamson, G.; Dew, T.; Zhang, W.; Kennedy, D.O. The Effects Of Chronic Trans-Resveratrol Supplementation On Aspects Of Cognitive Function, Mood, Sleep, Health And Cerebral Blood Flow In Healthy, Young Humans. Br. J. Nutr. 2015, 114, 1427–1437. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Wightman, E.L.; Reay, J.L.; Lietz, G.; Okello, E.J.; Wilde, A.; Haskell, C.F. Effects Of Resveratrol On Cerebral Blood Flow Variables And Cognitive Performance In Humans: A Double-Blind, Placebo-Controlled, Crossover Investigation. Am. J. Clin. Nutr. 2010, 91, 1590–1597. [Google Scholar] [CrossRef]
- Blanchard, O.L.; Friesenhahn, G.; Javors, M.A.; Smoliga, J.M. Development Of A Lozenge For Oral Transmucosal Delivery Of Trans-Resveratrol In Humans: Proof Of Concept. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Draijer, R.; Van Dorsten, F.A.; Zebregs, Y.E.; Hollebrands, B.; Peters, S.; Duchateau, G.S.; Grün, C.H. Impact Of Proteins On The Uptake, Distribution, And Excretion Of Phenolics In The Human Body. Nutrients 2016, 8, 814. [Google Scholar] [CrossRef]
- Brown, V.A.; Patel, K.R.; Viskaduraki, M.; Crowell, J.A.; Perloff, M.; Booth, T.D.; Vasilinin, G.; Sen, A.; Schinas, A.M.; Piccirilli, G.; et al. Repeat Dose Study Of The Cancer Chemopreventive Agent Resveratrol In Healthy Volunteers: Safety, Pharmacokinetics, And Effect On The Insulin-Like Growth Factor Axis. Cancer Res. 2010, 70, 9003–9011. [Google Scholar] [CrossRef] [PubMed]
- Howells, L.M.; Berry, D.P.; Elliott, P.J.; Jacobson, E.W.; Hoffmann, E.; Hegarty, B.; Brown, K.; Steward, W.P.; Gescher, A.J. Phase I Randomized, Double-Blind Pilot Study Of Micronized Resveratrol (Srt501) In Patients With Hepatic Metastases—Safety, Pharmacokinetics, And Pharmacodynamics. Cancer Prev. Res. 2011, 4, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- La Porte, C.; Voduc, N.; Zhang, G.; Seguin, I.; Tardiff, D.; Singhal, N.; Cameron, D.W. Steady-State Pharmacokinetics And Tolerability Of Trans-Resveratrol 2000mg Twice Daily With Food, Quercetin And Alcohol (Ethanol) In Healthy Human Subjects. Clin. Pharmacokinet. 2010, 49, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.H.X.; Howe, P.R.C.; Buckley, J.D.; Coates, A.M.; Kunz, I.; Berry, N.M. Acute Resveratrol Supplementation Improves Flow-Mediated Dilatation In Overweight/Obese Individuals With Mildly Elevated Blood Pressure. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.S.; Cantó, C. The Molecular Targets Of Resveratrol. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, S.; Holme, A.L. Resveratrol: Its Biologic Targets And Functional Activity. Antioxid. Redox Signal. 2009, 11, 2851–2897. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.G.; Kovoor, C.; Brown, K. Direct Molecular Targets Of Resveratrol: Identifying Key Interactions To Unlock Complex Mechanisms. Ann. N. Y. Acad. Sci. 2015, 1348, 124–133. [Google Scholar] [CrossRef]
- Vang, O. Resveratrol: Challenges In Analyzing Its Biological Effects. Ann. N. Y. Acad. Sci. 2015, 1348, 161–170. [Google Scholar] [CrossRef]
- Dolinsky, V.W.; Dyck, J.R.B. Calorie Restriction And Resveratrol In Cardiovascular Health And Disease. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 1477–1489. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Larrosa, M.; González-Sarrías, A.; Tomás-Barberán, F.; García-Conesa, M.; Espín, J. Resveratrol And Clinical Trials: The Crossroad From In Vitro Studies To Human Evidence. Curr. Pharm. Des. 2013, 19, 6064–6093. [Google Scholar] [CrossRef]
- Lacroix, S.; Klicic Badoux, J.; Scott-Boyer, M.-P.; Parolo, S.; Matone, A.; Priami, C.; Morine, M.J.; Kaput, J.; Moco, S. A Computationally Driven Analysis Of The Polyphenol-Protein Interactome. Sci. Rep. 2018, 8, 2232. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. David Bioinformatics Resources: Expanded Annotation Database And Novel Algorithms To Better Extract Biology From Large Gene Lists. Nucleic Acids Res. 2007, 35, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.G.; Barnes, K.C.; Bright, T.J.; Wang, S.A. The Genetic Association Database. Nat. Genet. 2004, 36, 431–432. [Google Scholar] [CrossRef]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.-Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. Interpro In 2017—Beyond Protein Family And Domain Annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Von Mering, C.; Campillos, M.; Jensen, L.J.; Bork, P. Stitch: Interaction Networks Of Chemicals And Proteins. Nucleic Acids Res. 2008, 36, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins As Regulators Of Metabolism And Healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Xia, N.; Förstermann, U.; Li, H. Effects Of Resveratrol On Enos In The Endothelium And The Perivascular Adipose Tissue. Ann. N. Y. Acad. Sci. 2017, 1403, 132–141. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; Von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. Stitch 5: Augmenting Protein-Chemical Interaction Networks With Tissue And Affinity Data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef]
- Portela, A.; Esteller, M. Epigenetic Modifications And Human Disease. Nat. Biotechnol. 2010, 28, 1057. [Google Scholar] [CrossRef]
- Kemper, J.K.; Xiao, Z.; Ponugoti, B.; Miao, J.; Fang, S.; Kanamaluru, D.; Tsang, S.; Wu, S.; Chiang, C.; Veenstra, T.D. Fxr Acetylation Is Normally Dynamically Regulated By P300 And Sirt1 But Constitutively Elevated In Metabolic Disease States. Cell Metab. 2009, 10, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Keylor, M.H.; Matsuura, B.S.; Stephenson, C.R.J. Chemistry And Biology Of Resveratrol-Derived Natural Products. Chem. Rev. 2015, 115, 8976–9027. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 Regulatory Network Provides An Interface Between Redox And Intermediary Metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Erlank, H.; Elmann, A.; Kohen, R.; Kanner, J. Polyphenols Activate Nrf2 In Astrocytes Via H2o2, Semiquinones, And Quinones. Free Radic. Biol. Med. 2011, 51, 2319–2327. [Google Scholar] [CrossRef]
- Seyyedebrahimi, S.; Khodabandehloo, H.; Nasli Esfahani, E.; Meshkani, R. The Effects Of Resveratrol On Markers Of Oxidative Stress In Patients With Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Acta Diabetol. 2018, 55, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Calamini, B.; Ratia, K.; Malkowski, M.G.; Cuendet, M.; Pezzuto, J.M.; Santarsiero, B.D.; Mesecar, A.D. Pleiotropic Mechanisms Facilitated By Resveratrol And Its Metabolites. Biochem. J. 2010, 429, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.L.; Ding, D.J.; Yan, W.J.; Li, R.R.; Dai, F.; Wang, Q.; Yu, S.S.; Li, Y.; Jin, X.L.; Zhou, B. Influence Of Glucuronidation And Reduction Modifications Of Resveratrol On Its Biological Activities. Chembiochem 2013, 14, 1094–1104. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Hu, F.B. The Epidemiology Of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Sethi, J.K.; Vidal-Puig, A.J. Thematic Review Series: Adipocyte Biology. Adipose Tissue Function And Plasticity Orchestrate Nutritional Adaptation. J. Lipid Res. 2007, 48, 1253–1262. [Google Scholar] [CrossRef]
- Maclean, P.S.; Higgins, J.A.; Giles, E.D.; Sherk, V.D.; Jackman, M.R. The Role For Adipose Tissue In Weight Regain After Weight Loss. Obes. Rev. 2015, 16, 45–54. [Google Scholar] [CrossRef]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, M. Resveratrol: Anti-Obesity Mechanisms Of Action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef]
- Korsholm, A.; Kjær, T.; Ornstrup, M.; Pedersen, S. Comprehensive Metabolomic Analysis In Blood, Urine, Fat, And Muscle In Men With Metabolic Syndrome: A Randomized, Placebo-Controlled Clinical Trial On The Effects Of Resveratrol After Four Months’ Treatment. Int. J. Mol. Sci. 2017, 18, 554. [Google Scholar] [CrossRef]
- Konings, E.; Timmers, S.; Boekschoten, M.V.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Müller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E.E. The Effects Of 30 Days Resveratrol Supplementation On Adipose Tissue Morphology And Gene Expression Patterns In Obese Men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef]
- Liu, K.; Zhou, R.; Wang, B.; Mi, M. Effect Of Resveratrol On Glucose Control And Insulin Sensitivity: A Meta-Analysis Of 11 Randomized Controlled Trials. Am. J. Clin. Nutr. 2014, 99, 1510–1519. [Google Scholar] [CrossRef]
- Moco, S.; Bino, R.J.; De Vos, R.C.H.; Vervoort, J. Metabolomics Technologies And Metabolite Identification. Trends Analaytical Chem. 2007, 26, 1694–1703. [Google Scholar] [CrossRef]
- Naranjo Pinta, M.; Montoliu, I.; Aura, A.-M.; Seppänen-Laakso, T.; Barron, D.; Moco, S. In Vitro Gut Metabolism Of [U-13C]-Quinic Acid, The Other Hydrolysis Product Of Chlorogenic Acid. Mol. Nutr. Food Res. 2018, 1800396. [Google Scholar] [CrossRef]




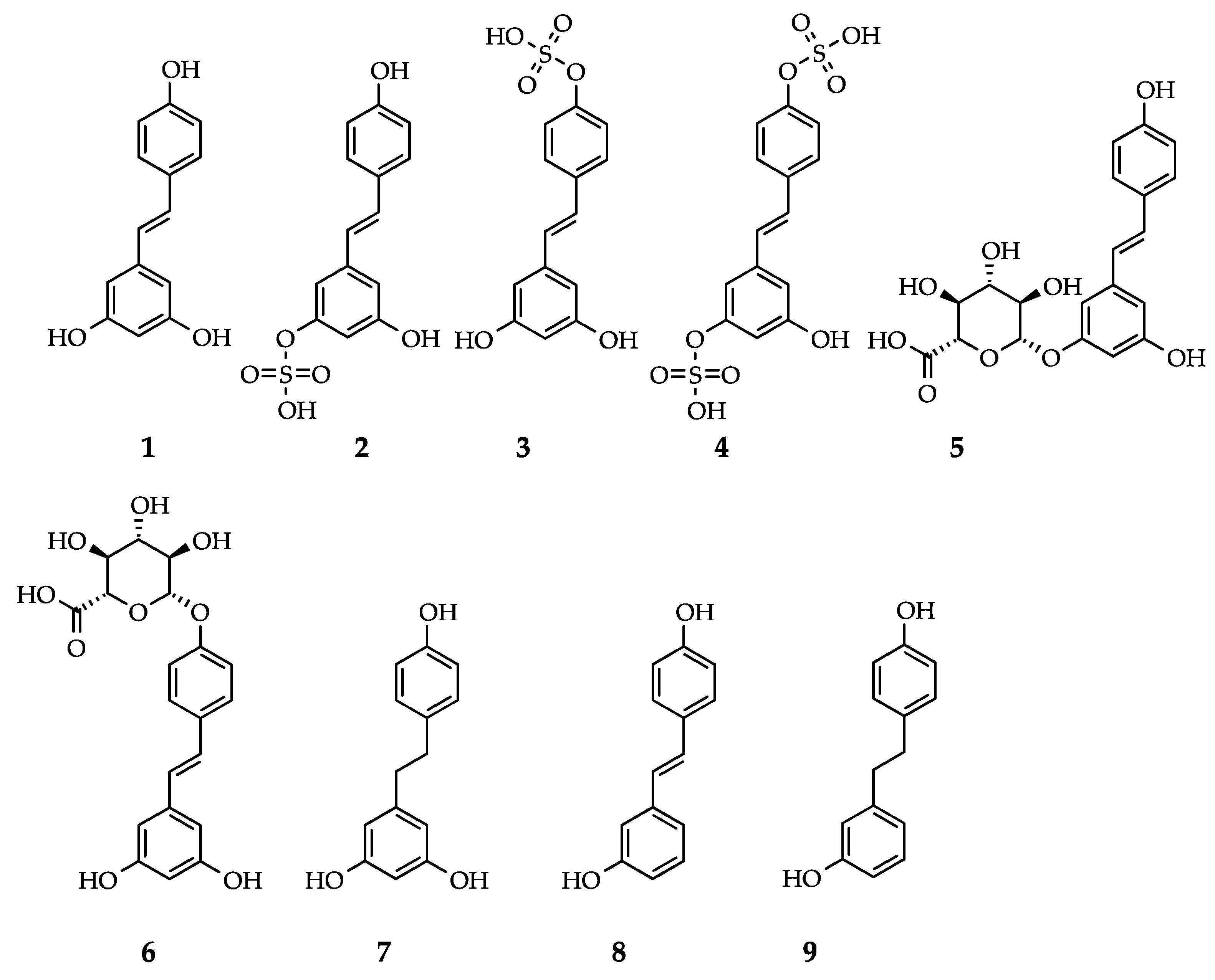{kind=link}
{kind=link}
{kind=link}
| Metabolite | Species and Tissue or Biofluid [Reference] |
|---|---|
| trans-resveratrol | Human: serum [41], plasma [15,16,42], urine [16,20] Rat: plasma [43,44,45], liver [44], lung [44], brain [44], kidney [44] Mouse: plasma [18,44,46], liver [18,44,46], lung [18,44], brain [44,46], kidney [18,44], heart [18,46], stomach [18], duodenum [18], intestine [18], muscle [18], spleen [18], thymus [18], urine [18], feces [18] |
| trans-resveratrol-4’-O-glucuronide | Human: serum [41], plasma [42], urine [42] Mouse: plasma [46] |
| trans-resveratrol-3-O-glucuronide | Human: serum [41], plasma [42], urine [16,42] Rat: plasma [43], liver [47], adipose tissue [47,48], skeletal muscle [47] Mouse: plasma [18,46], liver [18,46], lung [18], brain [46], kidney [18], heart [18,46], stomach [18], duodenum [18], intestine [18], muscle [18], spleen [18], thymus [18], urine [18], feces [18] |
| trans-resveratrol-diglucuronide | Human: plasma [49], urine [49] Mouse: plasma [46], liver [46] |
| trans-resveratrol-3-O-sulfate | Human: plasma [42], plasma [16], urine [16] Rat: adipose tissue [47,48] Mouse: plasma [18,46], liver [18,46], lung [18], brain [46], kidney [18], heart [18,46], stomach [18], duodenum [18], intestine [18], muscle [18], spleen [18], thymus [18], urine [18], feces [18] |
| trans-resveratrol-4’-O-sulfate | Human: plasma [42], plasma [16], urine [16,42] Rat: liver [47], adipose tissue [47,48] |
| cis-resveratrol-3-O-sulfate | Rat: adipose tissue [47,48] |
| trans-resveratrol-3,4’-disulfate | Human: plasma [42] Rat: adipose tissue [48] Mouse: plasma [18], liver [18], lung [18], kidney [18], heart [18], stomach [18], duodenum [18], intestine [18], muscle [18], urine [18], feces [18] |
| trans-resveratrol-glucuronide-sulfate | Mouse: plasma [46], liver [46] |
| dihydroresveratrol | Human: urine [20], plasma [15] Rat: liver [47], skeletal muscle [47] |
| dihydroresveratrol-glucuronide | Human: urine [16] Rat: liver [47] Mouse: plasma [46], liver [46] |
| dihydroresveratrol-sulfate | Human: urine [16] Rat: liver [47], adipose tissue [47] Mouse: plasma [46], liver [46] |
| dihydroresveratrol-glucuronide-sulfate | Mouse: plasma [46] |
| 3,4’-dihydroxy-trans-stilbene | Human: urine [20] |
| lunularin | Human: urine [20] |
| Number of Participants, Characteristics | Dose (mg) | Administration | Peak Plasma Concentration (ng/mL) | Reference |
|---|---|---|---|---|
| 15, healthy | 500 | Tablet | 71.18 | [65] |
| 6, low BMI 6, high BMI | 2125 | Tablet and drink | 634.32 498.56 | [66] |
| 7, healthy | 500 | Capsule 1 | 1598 | [67] |
| 8-9/dose, healthy | 250 500 | Capsule | 5.65 14.4 | [68] |
| 2, healthy | 146 | Lozenge | 328.5 | [69] |
| Number of Participants, Characteristics, Study Type | Dose (mg/day) | Days | Administration | Peak Plasma Concentration (ng/mL) | Reference |
|---|---|---|---|---|---|
| 6, low BMI 6, high BMI | 2125 | 11 | Tablet and drink | 903.0 245.0 | [66] |
| 35, healthy males, cross-over study | 800 | 5 | Capsule Dairy drink Soy drink Protein-free drink | Capsule: 0.56 Dairy drink: 0.61 Soy drink: 0.58 Protein free drink: 0.70 | [70] |
| 7, healthy | 500 | 28 | Capsule 1 | 2967.25 | [67] |
| 40, healthy, repeated sequential dosing | 500 1000 2500 5000 | 29 | Caplet | 43.8 141 331 967 | [71] |
| 6, patients with hepatic metastases, randomized double-blind clinical trial | 5000 | 14 | Micronized resveratrol mixed in liquid | 1942 | [72] |
| 8, healthy subjects | 2000 | 7 | Capsule | 1274 | [73] |
| 19, overweight or obese, randomized, double-blind, placebo-controlled, crossover intervention | 30 90 270 | 6 | Capsule | 181.31 532.00 1232.16 | [74] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. https://doi.org/10.3390/nu11010143
Springer M, Moco S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients. 2019; 11(1):143. https://doi.org/10.3390/nu11010143
Chicago/Turabian StyleSpringer, Margherita, and Sofia Moco. 2019. "Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity" Nutrients 11, no. 1: 143. https://doi.org/10.3390/nu11010143
APA StyleSpringer, M., & Moco, S. (2019). Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients, 11(1), 143. https://doi.org/10.3390/nu11010143