Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes

 ,
, 
 and
and 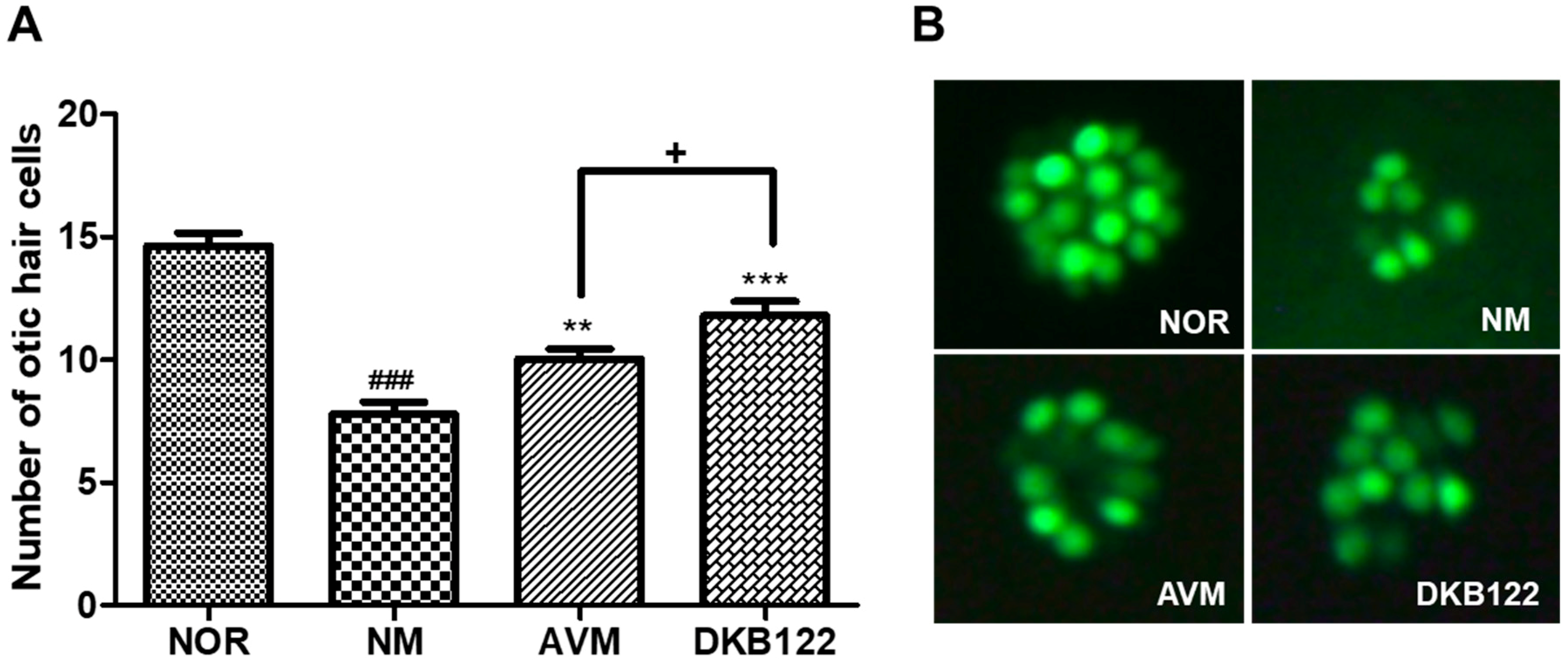{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals
2.3. Ethical Statement
2.4. Neomycin-Induced Ototoxicity in a Zebrafish Model
2.5. The 50% Effective Concentration (EC50)
2.6. Noise-Induced Hearing Loss (NIHL) in Mice
2.7. Auditory Brainstem Response (ABR) Test
2.8. Evaluation of Otoprotective Effects of Cochlear Hair Cells
2.9. Cell Culture
2.10. mRNA Sequencing and Pathway Analysis
2.11. Real Time-qPCR
2.12. Statistical Analysis
3. Results
3.1. Recovery Effect on Neomycin-Induced Otic Hair Cells Damage in Zebrafish
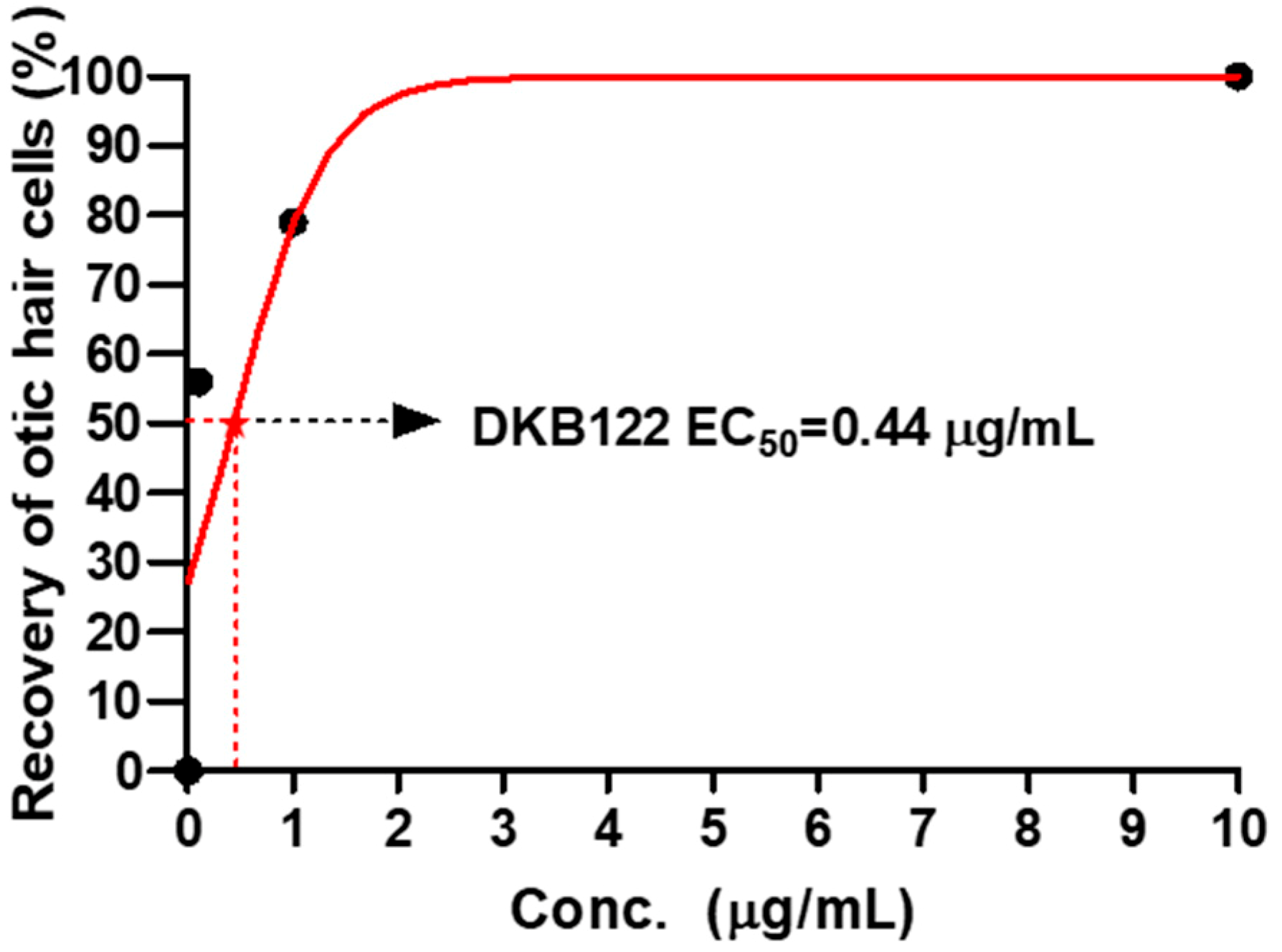
3.2. EC50 Values of DKB122
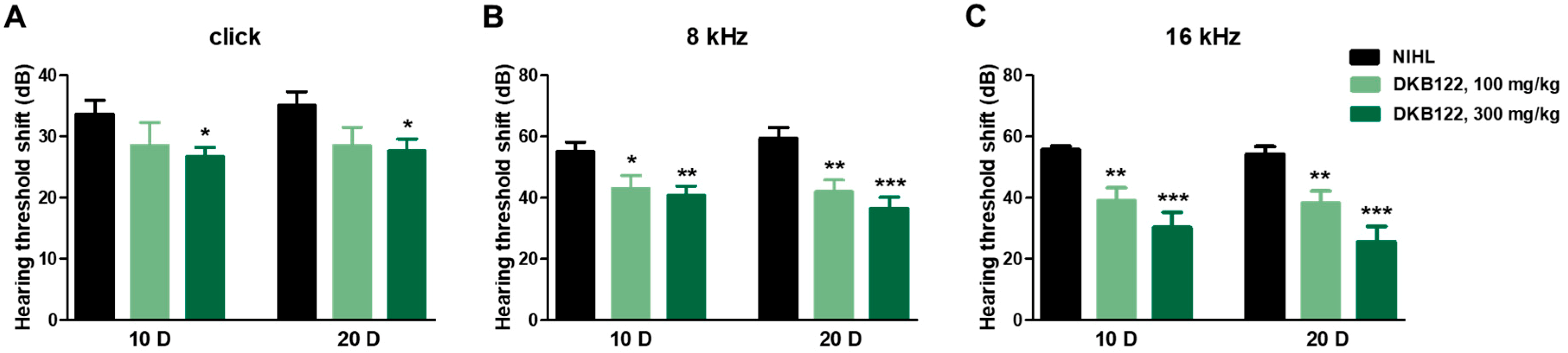
3.3. Auditory Evaluation
3.4. Otoprotective Effects of Cochlear Hair Cells
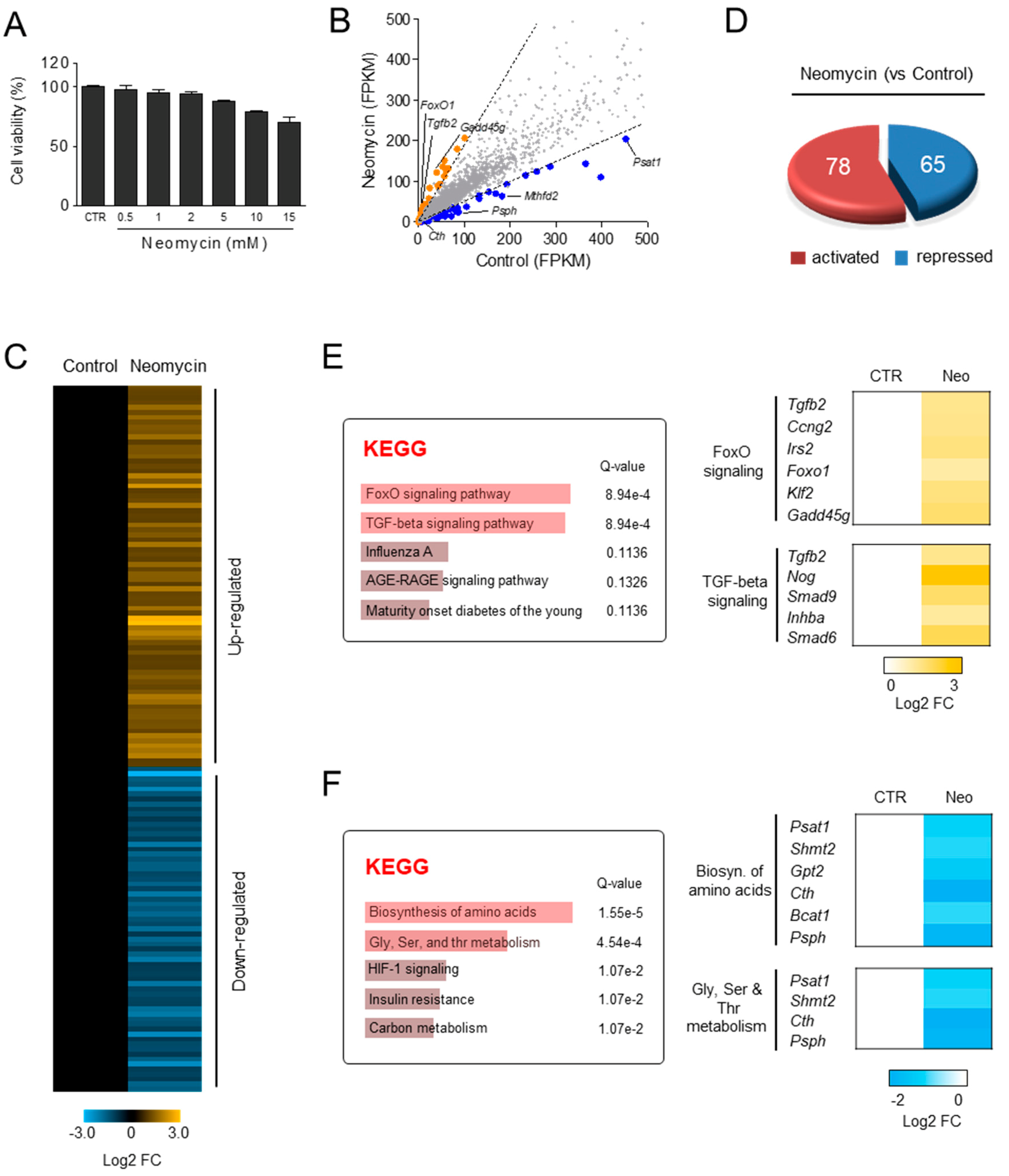
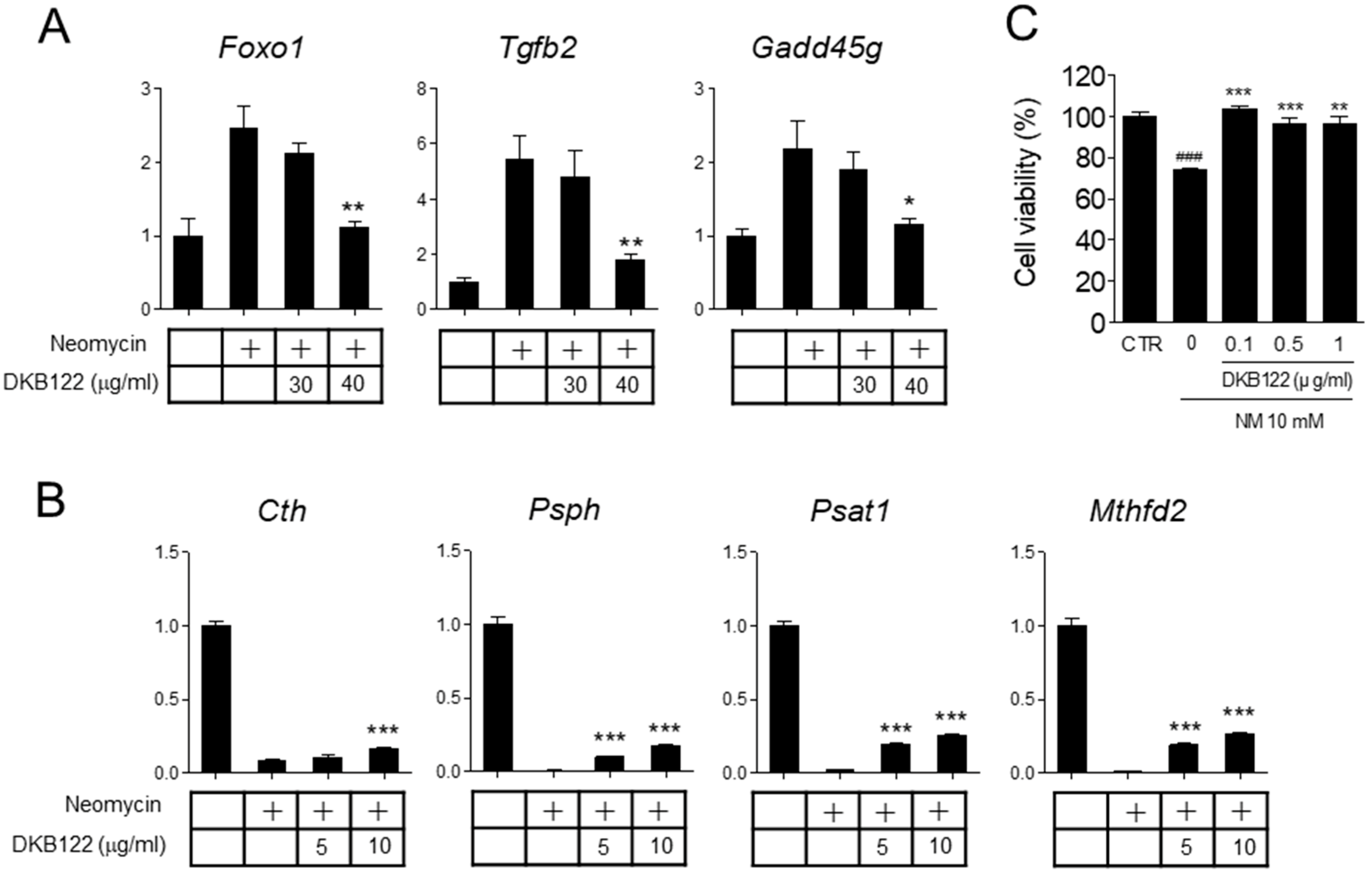
3.5. Differential Gene Expression Induced by Neomycin in HEI-OC1 Cells
3.6. DKB122 Rescues Altered Gene Expression and Reduced Viability by Neomycin in HEI-OC1 Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Basner, M.; Babisch, W.; Davis, A.; Brink, M.; Clark, C.; Janssen, S.; Stansfeld, S. Auditory and non-auditory effects of noise on health. Lancet 2014, 383, 1325–1332. [Google Scholar] [CrossRef]
- Rossini, B.A.A.; Penido, N.O.; Munhoz, M.S.L.; Bogaz, E.A.; Curi, R.S. Sudden Sensorineural Hearing Loss and Autoimmune Systemic Diseases. Int. Arch. Otorhinolaryngol. 2017, 21, 213–223. [Google Scholar] [PubMed]
- Shargorodsky, J.; Curhan, S.G.; Curhan, G.C.; Eavey, R. Change in prevalence of hearing loss in US adolescents. JAMA 2010, 304, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Yamasoba, T.; Lin, F.R.; Someya, S.; Kashio, A.; Sakamoto, T.; Kondo, K. Current concepts in age-related hearing loss: Epidemiology and mechanistic pathways. Hearth Res. 2013, 303, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, J.N.; Zhou, H.F. Endothelial progenitor cells in patients with age-related hearing loss. Am. J. Otolaryngol. 2014, 35, 695–698. [Google Scholar] [CrossRef]
- Dreher, M.L.; Davenport, A.J. Hass avocado composition and potential health effects. Crit. Rev. Food Sci. Nutr. 2013, 53, 738–750. [Google Scholar] [CrossRef]
- Krumreich, F.D.; Borges, C.D.; Mendonça, C.R.B.; Jansen-Alves, C.; Zambiazi, R.C. Bioactive compounds and quality parameters of avocado oil obtained by different processes. Food Chem. 2018, 257, 376–381. [Google Scholar] [CrossRef]
- Daiuto, É.R.; Vieites, R.L.; Tremocoldi, M.A.; Vileigas, D.F. Physico chemical stability of avocado product (Persea americana Mill.) storaged under low temperature. Alimentos e Nutrição 2010, 21, 99–107. [Google Scholar]
- Martínez-Padilla, L.P.; Franke, L.; Xu, X.Q.; Juliano, P. Improved extraction of avocado oil by application of sono-physical processes. Ultrason. Sonochem. 2018, 40, 720–726. [Google Scholar] [CrossRef]
- Ortiz-Avila, O.; Esquivel-Martínez, M.; Olmos-Orizaba, B.E.; Saavedra-Molina, A.; Rodriguez-Orozco, A.R.; Cortés-Rojo, C. Avocado oil improves mitochondrial function and decreases oxidative stress in brain of diabetic rats. J. Diabetes Res. 2015, 2015, 485759. [Google Scholar] [CrossRef]
- Kalinec, G.M.; Park, C.; Thein, P.; Kalinec, F. Working with Auditory HEI-OC1 Cells. J. Vis. Exp. 2016, 115, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Korrapati, S.; Roux, I.; Glowatzki, E.; Doetzlhofer, A. Notch signaling limits supporting cell plasticity in the hair cell-damaged early postnatal murine cochlea. PLoS ONE 2013, 8, e73276. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Song, Y.; Wang, R.; Gao, S.S.; Clifton, W.; Raphael, P.; Chao, S.I.; Pereira, F.A.; Groves, A.K.; Oghalai, J.S. Prestin regulation and function in residual outer hair cells after noise-induced hearing loss. PLoS ONE 2013, 8, e82602. [Google Scholar] [CrossRef]
- Park, S.Y.; Park, J.M.; Back, S.A.; Yeo, S.W.; Park, S.N. Functional Significance of Medial Olivocochlear System Morphology in the Mouse Cochlea. Clin. Exp. Otorhinolaryngol. 2017, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, X.; Sun, M.; Xu, T.; Wang, A. FoxO3a plays a key role in the protective effects of pomegranate peel extract against amikacin-induced ototoxicity. Int. J. Mol. Med. 2017, 40, 175–181. [Google Scholar] [CrossRef]
- Murillo-Cuesta, S.; Rodríguez-de la Rosa, L.; Contreras, J.; Celaya, A.M.; Camarero, G.; Rivera, T.; Varela-Nieto, I. Transforming growth factor β1 inhibition protects from noise-induced hearing loss. Front. Aging Neurosci. 2015, 7, 32. [Google Scholar]
- Yang, Y.; Wu, J.; Cai, J.; He, Z.; Yuan, J.; Zhu, X.; Li, Y.; Li, M.; Guan, H. PSAT1 regulates cyclin D1 degradation and sustains proliferation of non-small cell lung cancer cells. Int. J. Cancer 2015, 136, E39–E50. [Google Scholar] [CrossRef]
- Gao, S.; Ge, A.; Xu, S.; You, Z.; Ning, S.; Zhao, Y.; Pang, D. PSAT1 is regulated by ATF4 and enhances cell proliferation via the GSK3β/β-catenin/cyclin D1 signaling pathway in ER-negative breast cancer. J. Exp. Clin. Cancer Res. 2017, 36, 179. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Z.; Li, X.; Liu, B.; Liu, M.; Liu, L.; Chen, S.; Ren, M.; Wang, Y.; Yu, M.; et al. SHMT2 Desuccinylation by SIRT5 Drives Cancer Cell Proliferation. Cancer Res. 2018, 78, 372–386. [Google Scholar] [CrossRef]
- Sen, S.; Kawahara, B.; Mahata, S.K.; Tsai, R.; Yoon, A.; Hwang, L.; Hu-Moore, K.; Villanueva, C.; Vajihuddin, A.; Parameshwar, P.; et al. Cystathionine: A novel oncometabolite in human breast cancer. Arch. Biochem. Biophys. 2016, 604, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Bachelor, M.A.; Lu, Y.; Owens, D.M. l-3-Phosphoserine phosphatase (PSPH) regulates cutaneous squamous cell carcinoma proliferation independent of L-serine biosynthesis. J. Dermatol. Sci. 2011, 63, 164–172. [Google Scholar] [CrossRef]
- Gopinath, B.; Flood, V.M.; Rochtchina, E.; McMahon, C.M.; Mitchell, P. Consumption of omega-3 fatty acids and fish and risk of age-related hearing loss. Am. J. Clin. Nutr. 2010, 92, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vega, R.; Partearroyo, T.; Vallecillo, N.; Varela-Moreiras, G.; Pajares, M.A.; Varela-Nieto, I. Long-term omega-3 fatty acid supplementation prevents expression changes in cochlear homocysteine metabolism and ameliorates progressive hearing loss in C57BL/6J mice. J. Nutr. Biochem. 2015, 26, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.O.; Dorantes, L.; Galíndez, J.; Guzmán, R.I. Effect of different extraction methods on fatty acids, volatile compounds, and physical and chemical properties of avocado (Persea americana Mill.) oil. J. Agric. Food Chem. 2003, 51, 2216–2221. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.J.; Venkatesan, T.K. Aqueous isopropyl alcohol for extraction of free fatty acids from oils. J. Am. Oil Chem. Soc. 1989, 66, 783–787. [Google Scholar] [CrossRef]
- Pal, P.K.; Bhattacharyya, D.K.; Ghosh, S. Isopropanol fractionation of butter oil and characteristics of fractions. J. Am. Oil Chem. Soc. 2000, 77, 1215–1218. [Google Scholar] [CrossRef]
- Stawicki, T.M.; Esterberg, R.; Hailey, D.W.; Raible, D.W.; Rubel, E.W. Using the zebrafish lateral line to uncover novel mechanisms of action and prevention in drug-induced hair cell death. Front. Cell Neurosci. 2015, 9, 46. [Google Scholar] [CrossRef]
- Castañeda, R.; Natarajan, S.; Jeong, S.Y.; Hong, B.N.; Kang, T.H. Traditional oriental medicine for sensorineural hearing loss: Can ethnopharmacology contribute to potential drug discovery? J. Ethnopharmacol. 2018, 231, 409–428. [Google Scholar] [CrossRef]
- Yu, X.; Liu, W.; Fan, Z.; Qian, F.; Zhang, D.; Han, Y.; Xu, L.; Sun, G.; Qi, J.; Zhang, S.; et al. c-Myb knockdown increases the neomycin-induced damage to hair-cell-like HEI-OC1 cells in vitro. Sci. Rep. 2017, 7, 41094. [Google Scholar] [CrossRef]
- Partearroyo, T.; Vallecillo, N.; Pajares, M.A.; Varela-Moreiras, G.; Varela-Nieto, I. Cochlear Homocysteine Metabolism at the Crossroad of Nutrition and Sensorineural Hearing Loss. Front. Mol. Neurosci. 2017, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.A.; Teh, B.M.; Friedland, P. Bilateral sudden sensorineural hearing loss: Review. J. Laryngol. Otol. 2014, 128, S8–S15. [Google Scholar] [CrossRef] [PubMed]
- Layman, W.S.; Zuo, J. Preventing ototoxic hearing loss by inhibiting histone deacetylases. Cell Death Dis. 2015, 6, e1882. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Cao, Y.; Jiang, Y.; Wei, Y.; Liu, H. Melatonin protects mouse granulosa cells against oxidative damage by inhibiting FOXO1-mediated autophagy: Implication of an antioxidation-independent mechanism. Redox. Biol. 2018, 18, 138–157. [Google Scholar] [CrossRef] [PubMed]
- Schuster, N.; Krieglstein, K. Mechanisms of TGF-β-mediated apoptosis. Cell Tissue Res. 2002, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gu, H.; Chang, J.; Wu, J.; Wang, D.; Chen, S.; Yang, X.; Qian, B. MicroRNA-383 regulates the apoptosis of tumor cells through targeting Gadd45g. PLoS ONE 2014, 9, e110472. [Google Scholar] [CrossRef] [PubMed]
- Fischel-Ghodsian, N. Genetic factors in aminoglycoside toxicity. Pharmacogenomics 2005, 6, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, R.; Wang, Q.; Yan, Q.; Deng, J.H.; Han, D.; Bai, Y.; Young, W.Y.; Guan, M.X. Maternally inherited aminoglycoside-induced and nonsyndromic deafness is associated with the novel C1494T mutation in the mitochondrial 12S rRNA gene in a large Chinese family. Am. J. Hum. Genet. 2004, 74, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Prezant, T.R.; Agapian, J.V.; Bohlman, M.C.; Bu, X.; Oztas, S.; Qiu, W.Q.; Arnos, K.S.; Cortopassi, G.A.; Jaber, L.; Rotter, J.I.; et al. Mitochondrial ribosomal RNA mutation associated with both antibiotic-induced and non-syndromic deafness. Nat. Genet. 1993, 4, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.P.; Katz, J.; Fanning, K.D.; Harris, K.A.; Nicholas, B.D.; Lacy, M.; Pagana, J.; Agris, P.F.; Shin, J.B. A novel role of cytosolic protein synthesis inhibition in aminoglycoside ototoxicity. J. Neurosci. 2013, 33, 3079–3093. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, Y.H.; Rodriguez, I.; Jeong, S.Y.; Pham, T.N.M.; Nuankaew, W.; Kim, Y.H.; Castañeda, R.; Jeong, S.Y.; Park, M.S.; Lee, K.W.; et al. Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes. Nutrients 2019, 11, 113. https://doi.org/10.3390/nu11010113
Nam YH, Rodriguez I, Jeong SY, Pham TNM, Nuankaew W, Kim YH, Castañeda R, Jeong SY, Park MS, Lee KW, et al. Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes. Nutrients. 2019; 11(1):113. https://doi.org/10.3390/nu11010113
Chicago/Turabian StyleNam, Youn Hee, Isabel Rodriguez, Seo Yeon Jeong, Thu Nguyen Minh Pham, Wanlapa Nuankaew, Yun Hee Kim, Rodrigo Castañeda, Seo Yule Jeong, Min Seon Park, Kye Wan Lee, and et al. 2019. "Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes" Nutrients 11, no. 1: 113. https://doi.org/10.3390/nu11010113
APA StyleNam, Y. H., Rodriguez, I., Jeong, S. Y., Pham, T. N. M., Nuankaew, W., Kim, Y. H., Castañeda, R., Jeong, S. Y., Park, M. S., Lee, K. W., Lee, J. S., Kim, D. H., Park, Y. H., Kim, S. H., Moon, I. S., Choung, S.-Y., Hong, B. N., Jeong, K. W., & Kang, T. H. (2019). Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes. Nutrients, 11(1), 113. https://doi.org/10.3390/nu11010113