Plasma Sulphur-Containing Amino Acids, Physical Exercise and Insulin Sensitivity in Overweight Dysglycemic and Normal Weight Normoglycemic Men

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
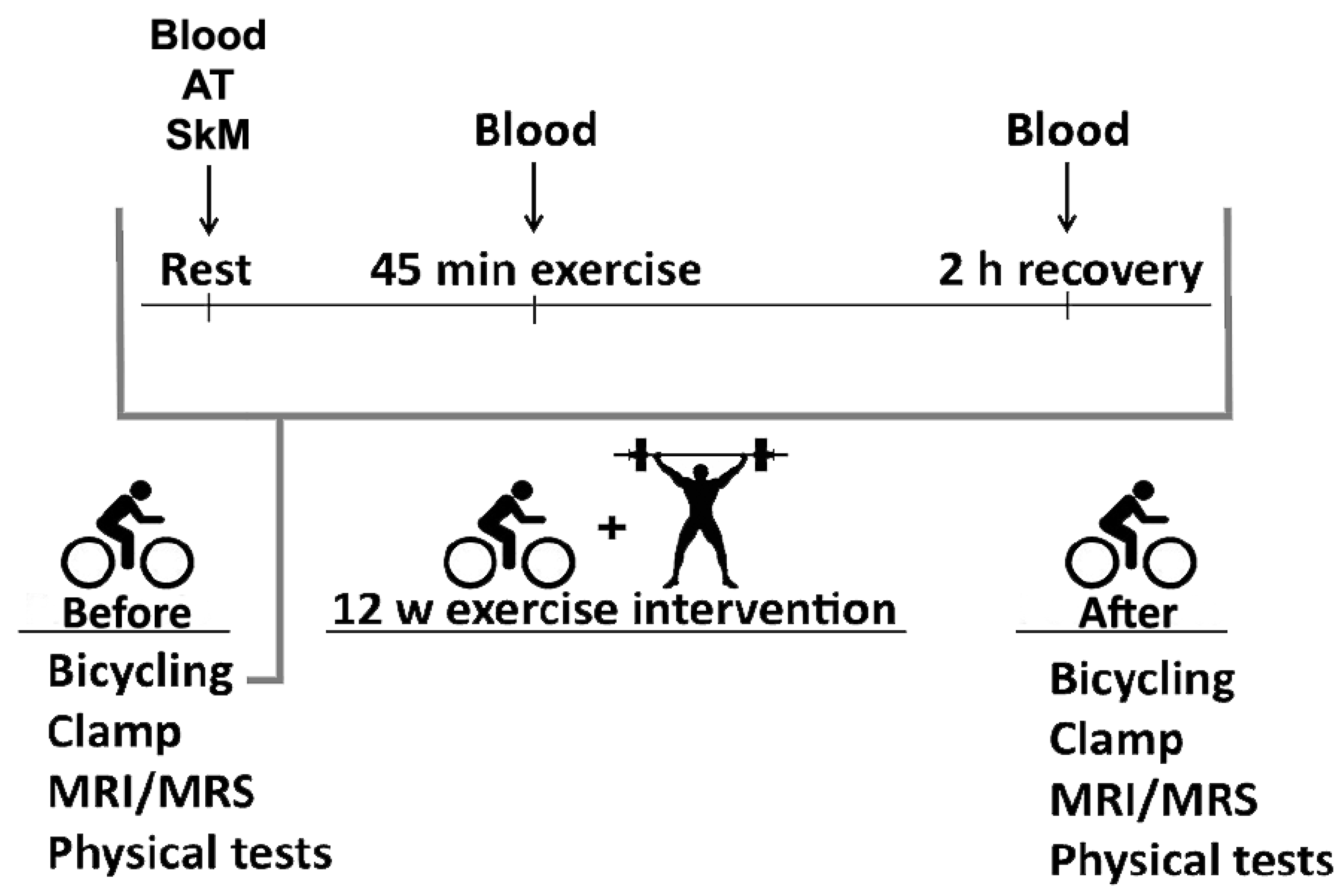
2.1. Study Participants and Experimental Methods
2.2. Diet
2.3. Exercise Intervention
2.3.1. Bicycle Tests
2.3.2. Strength and Endurance Exercise
2.4. Physical Fitness and Insulin Sensitivity
2.4.1. VO2max
2.4.2. Euglycemic Hyperinsulinemic Clamp
2.4.3. Tissue Sampling
2.4.4. Plasma Metabolites
2.4.5. mRNA-Sequencing
2.4.6. Magnetic Resonance Imaging and Spectrometry (MRI/MRS)
2.4.7. Statistics
2.4.8. Study Approval
3. Results
3.1. Subject Characteristics and Responses to 12 Week Exercise Intervention
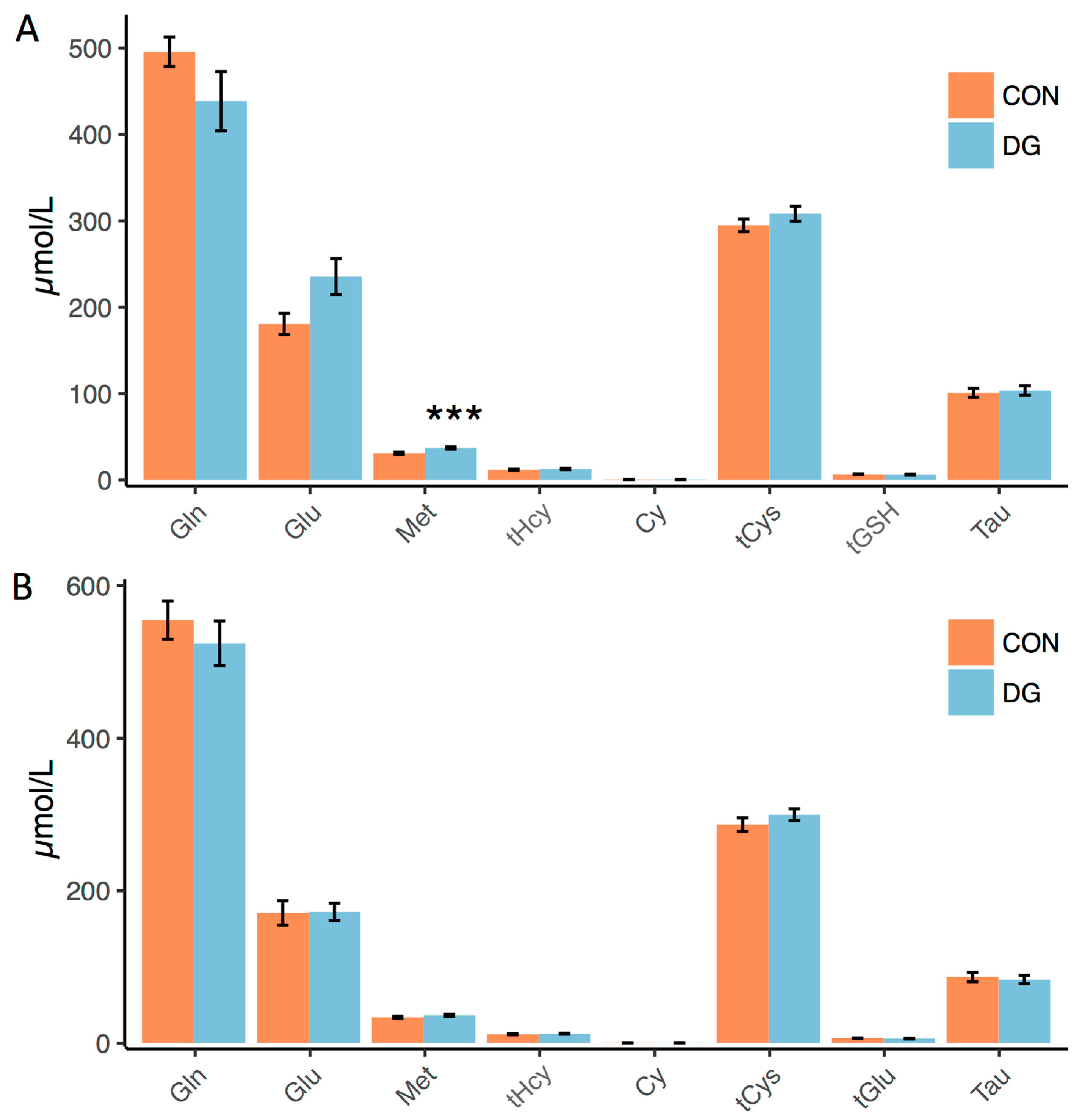
3.2. Plasma Markers of Long-Term Exercise
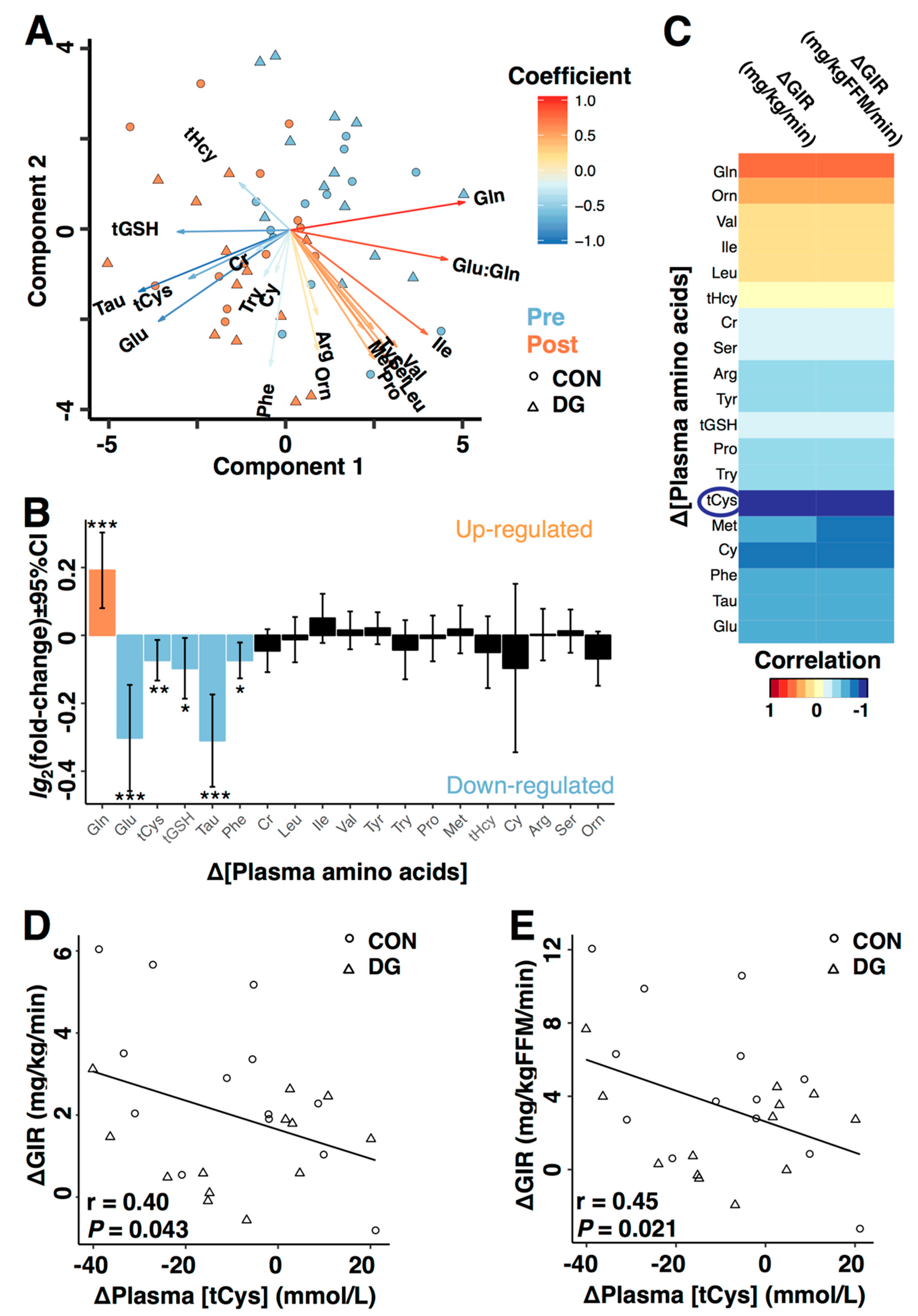
3.3. Plasma Metabolites and Insulin Sensitivity
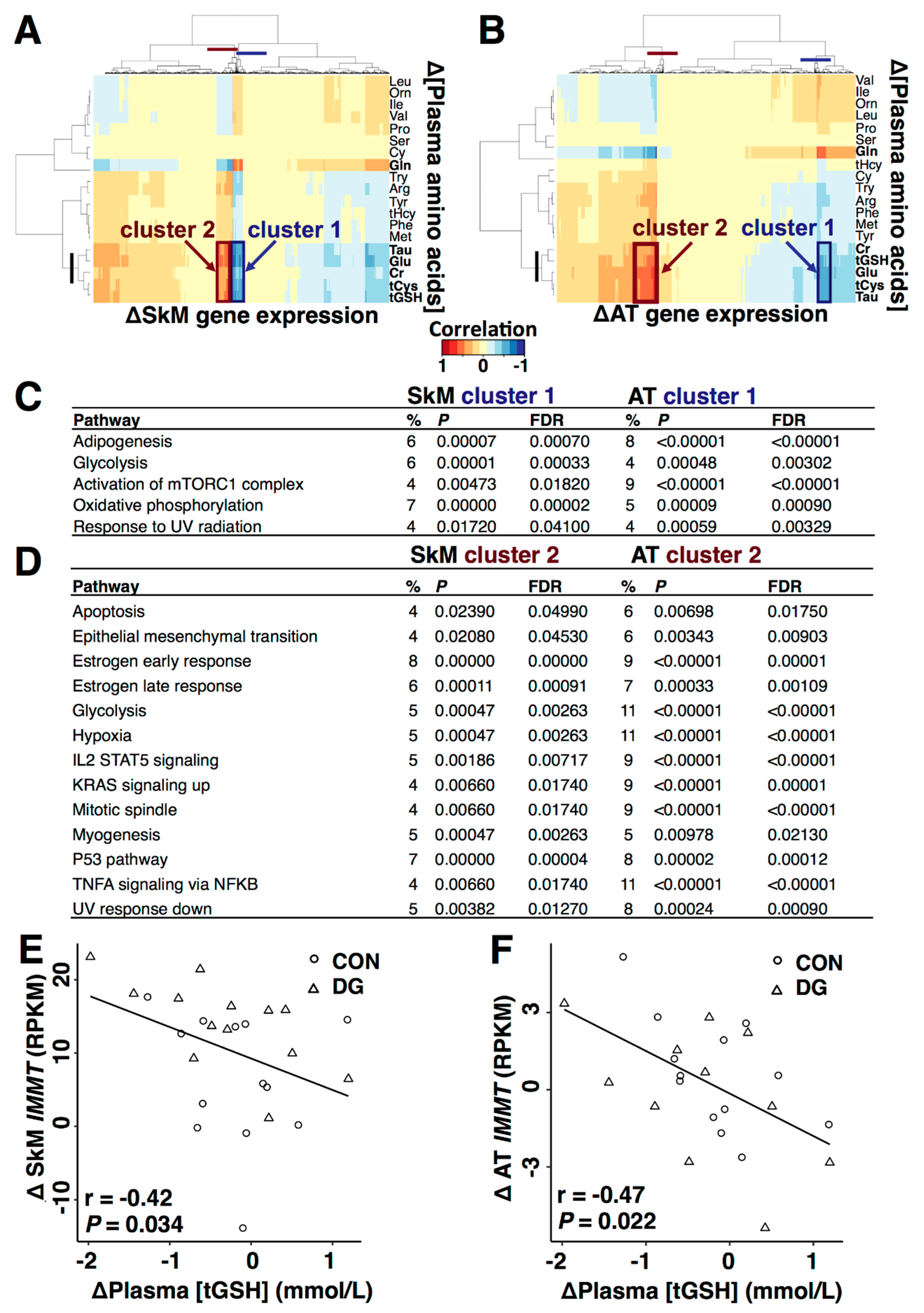
3.4. Plasma Metabolites and Metabolic Gene Expression
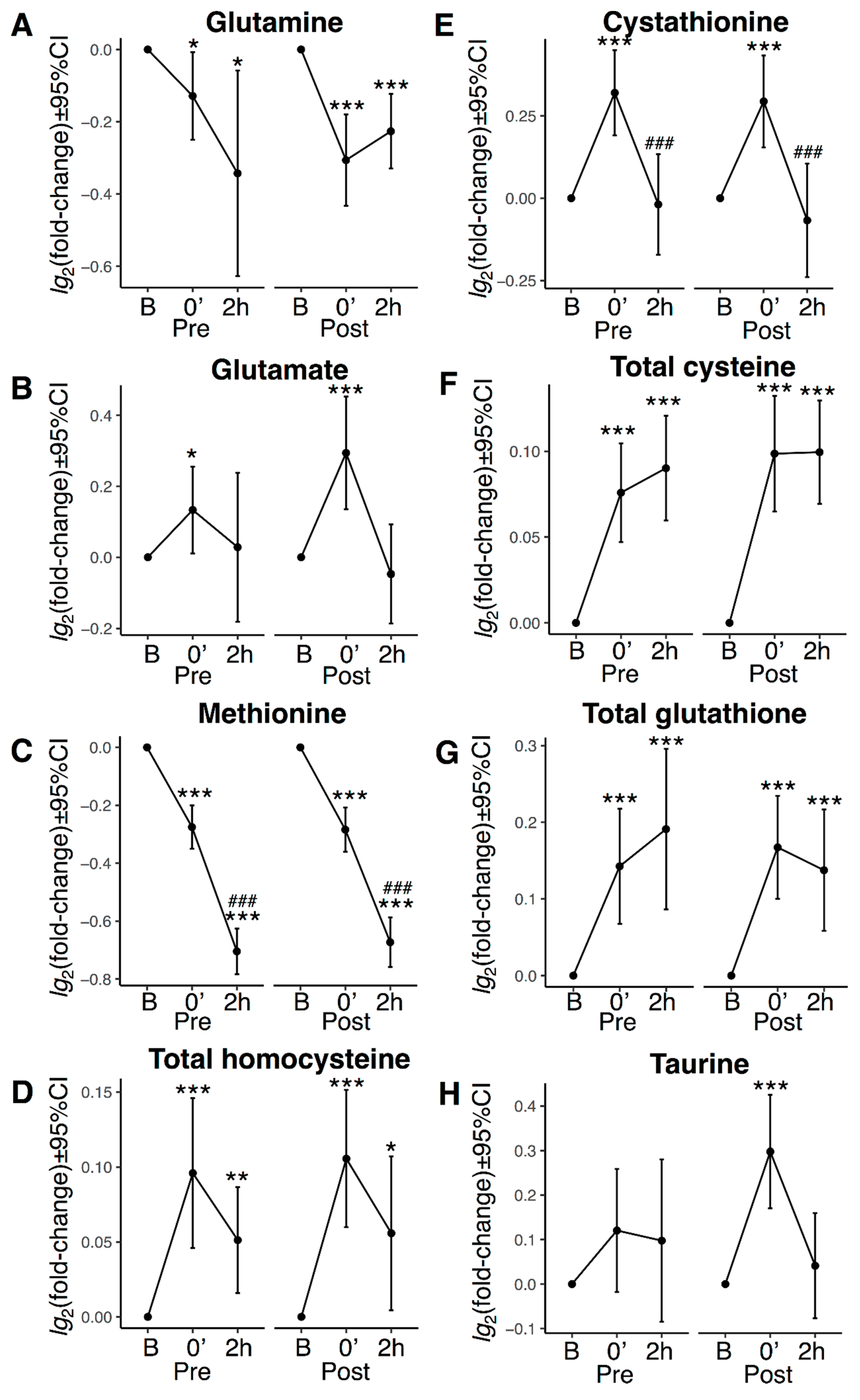
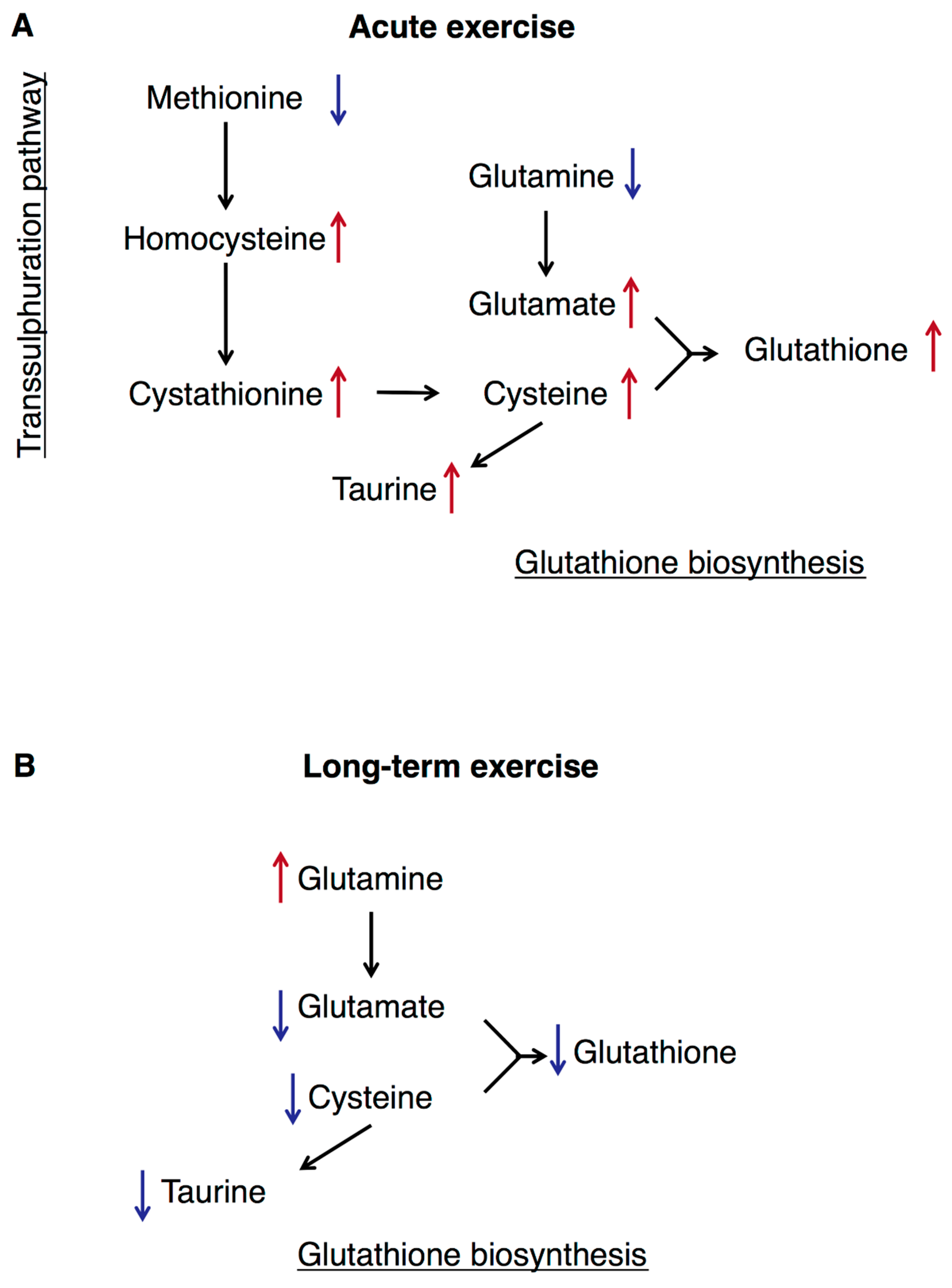
3.5. Transsulphuration, Glutathione Biosynthesis, and Acute Exercise
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Goodyear, L.J.; Kahn, B.B. Exercise, glucose transport, and insulin sensitivity. Ann. Rev. Med. 1998, 49, 235–261. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C.A.; Shangraw, R.E.; Prince, M.J.; Peters, E.J.; Wolfe, R.R. Bed-rest-induced insulin resistance occurs primarily in muscle. Met. Clin. Exp. 1988, 37, 802–806. [Google Scholar] [CrossRef]
- Wilson, P.W.; McGee, D.L.; Kannel, W.B. Obesity, very low density lipoproteins, and glucose intolerance over fourteen years: The Framingham Study. Am. J. Epidemiol. 1981, 114, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Shah, S.H.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.; Slentz, C.A.; Tanner, C.J.; Kuchibhatla, M.; Houmard, J.A.; Newgard, C.B.; et al. Relationships between circulating metabolic intermediates and insulin action in overweight to obese, inactive men and women. Diabetes Care 2009, 32, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Met. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ni, Y.; Ma, X.; Bao, Y.; Liu, J.; Huang, F.; Hu, C.; Xie, G.; Zhao, A.; Jia, W.; et al. Branched-chain and aromatic amino acid profiles and diabetes risk in Chinese populations. Sci. Rep. 2016, 6, 20594. [Google Scholar] [CrossRef] [PubMed]
- Knebel, B.; Strassburger, K.; Szendroedi, J.; Kotzka, J.; Scheer, M.; Nowotny, B.; Mussig, K.; Lehr, S.; Pacini, G.; Finner, H.; et al. Specific Metabolic Profiles and Their Relationship to Insulin Resistance in Recent-Onset Type 1 and Type 2 Diabetes. J. Clin. Endocrinol. Met. 2016, 101, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S1640S. [Google Scholar] [CrossRef]
- El-Khairy, L.; Ueland, P.M.; Nygard, O.; Refsum, H.; Vollset, S.E. Lifestyle and cardiovascular disease risk factors as determinants of total cysteine in plasma: The Hordaland Homocysteine Study. Am. J. Clin. Nutr. 1999, 70, 1016–1024. [Google Scholar] [CrossRef]
- Elshorbagy, A.K.; Nurk, E.; Gjesdal, C.G.; Tell, G.S.; Ueland, P.M.; Nygard, O.; Tverdal, A.; Vollset, S.E.; Refsum, H. Homocysteine, cysteine, and body composition in the Hordaland Homocysteine Study: Does cysteine link amino acid and lipid metabolism? Am. J. Clin. Nutr. 2008, 88, 738–746. [Google Scholar] [CrossRef]
- Elshorbagy, A.K.; Refsum, H.; Smith, A.D.; Graham, I.M. The association of plasma cysteine and gamma-glutamyltransferase with BMI and obesity. Obesity (Silver Spring) 2009, 17, 1435–1440. [Google Scholar] [CrossRef]
- Elshorbagy, A.K. Body composition in gene knockouts of sulfur amino acid-metabolizing enzymes. Mamm. Genome 2014. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Church, C.; Valdivia-Garcia, M.; Smith, A.D.; Refsum, H.; Cox, R. Dietary cystine level affects metabolic rate and glycaemic control in adult mice. J. Nutr. Biochem. 2012, 23, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Refsum, H.; Butte, N. The Association of Cysteine with Obesity, Inflammatory Cytokines and Insulin Resistance in Hispanic Children and Adolescents. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Glutathione synthesis. BBA-Gen Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Mosharov, E.; Cranford, M.R.; Banerjee, R. The quantitatively important relationship between homocysteine metabolism and glutathione synthesis by the transsulfuration pathway and its regulation by redox changes. Biochemistry 2000, 39, 13005–13011. [Google Scholar] [CrossRef] [PubMed]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [CrossRef]
- Helmrich, S.P.; Ragland, D.R.; Leung, R.W.; Paffenbarger, R.S., Jr. Physical activity and reduced occurrence of non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1991, 325, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Soman, V.R.; Koivisto, V.A.; Deibert, D.; Felig, P.; DeFronzo, R.A. Increased insulin sensitivity and insulin binding to monocytes after physical training. N. Engl. J. Med. 1979, 301, 1200–1204. [Google Scholar] [CrossRef]
- Borghouts, L.B.; Keizer, H.A. Exercise and insulin sensitivity: A review. Int. J. Sport. Med. 2000, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sigal, R.J.; Kenny, G.P.; Boule, N.G.; Wells, G.A.; Prud’homme, D.; Fortier, M.; Reid, R.D.; Tulloch, H.; Coyle, D.; Phillips, P.; et al. Effects of aerobic training, resistance training, or both on glycemic control in type 2 diabetes: A randomized trial. Ann. Int. Med. 2007, 147, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Cuff, D.J.; Meneilly, G.S.; Martin, A.; Ignaszewski, A.; Tildesley, H.D.; Frohlich, J.J. Effective exercise modality to reduce insulin resistance in women with type 2 diabetes. Diabetes Care 2003, 26, 2977–2982. [Google Scholar] [CrossRef] [PubMed]
- Tarnopolsky, M. Protein requirements for endurance athletes. Nutrition 2004, 20, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Medelli, J.; Lounana, J.; Hill, D. Variation in plasma amino acid concentrations during a cycling competition. J. Sport. Med. Phys. Fit. 2003, 43, 236–242. [Google Scholar]
- Deminice, R.; Ribeiro, D.F.; Frajacomo, F.T. The Effects of Acute Exercise and Exercise Training on Plasma Homocysteine: A Meta-Analysis. PLoS ONE 2016, 11, e0151653. [Google Scholar] [CrossRef] [PubMed]
- Langleite, T.M.; Jensen, J.; Norheim, F.; Gulseth, H.L.; Tangen, D.S.; Kolnes, K.J.; Heck, A.; Storas, T.; Grothe, G.; Dahl, M.A.; et al. Insulin sensitivity, body composition and adipose depots following 12 w combined endurance and strength training in dysglycemic and normoglycemic sedentary men. Arch. Physiol. Biochem. 2016, 122, 167–179. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Lillegaard, I.T.; Karlsen, A.; Blomhoff, R.; Drevon, C.A.; Andersen, L.F. Evaluation of energy and dietary intake estimates from a food frequency questionnaire using independent energy expenditure measurement and weighed food records. Nutr. J. 2010, 9, 37. [Google Scholar] [CrossRef]
- Nes, M.; Frost Andersen, L.; Solvoll, K.; Sandstad, B.; Hustvedt, B.E.; Lovo, A.; Drevon, C.A. Accuracy of a quantitative food frequency questionnaire applied in elderly Norwegian women. Eur. J. Clin. Nutr. 1992, 46, 809–821. [Google Scholar]
- Antoniades, C.; Shirodaria, C.; Leeson, P.; Baarholm, O.A.; Van-Assche, T.; Cunnington, C.; Pillai, R.; Ratnatunga, C.; Tousoulis, D.; Stefanadis, C.; et al. MTHFR 677 C>T Polymorphism reveals functional importance for 5-methyltetrahydrofolate, not homocysteine, in regulation of vascular redox state and endothelial function in human atherosclerosis. Circulation 2009, 119, 2507–2515. [Google Scholar] [CrossRef]
- Lee, S.; Norheim, F.; Gulseth, H.L.; Langleite, T.M.; Aker, A.; Gundersen, T.E.; Holen, T.; Birkeland, K.I.; Drevon, C.A. Skeletal muscle phosphatidylcholine and phosphatidylethanolamine respond to exercise and influence insulin sensitivity in men. Sci. Rep. 2018, 8, 6531. [Google Scholar] [CrossRef] [PubMed]
- Liquet, B.; Le Cao, K.A.; Hocini, H.; Thiebaut, R. A novel approach for biomarker selection and the integration of repeated measures experiments from two assays. BMC Bioinform. 2012, 13, 325. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, J.A.; van Velzen, E.J.J.; Hoefsloot, H.C.J.; Smilde, A.K. Multivariate paired data analysis: Multilevel PLSDA versus OPLSDA. Metabolomics 2010, 6, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Bushel, P.R.; Wolfinger, R.D.; Gibson, G. Simultaneous clustering of gene expression data with clinical chemistry and pathological evaluations reveals phenotypic prototypes. BMC Syst. Biol. 2007, 1, 15. [Google Scholar] [CrossRef]
- Le Cao, K.A.; Rossouw, D.; Robert-Granie, C.; Besse, P. A sparse PLS for variable selection when integrating omics data. Stat. Appl. Genet. Mol. Biol. 2008, 7, Article–35. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdottir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Subramanian, A.; Kuehn, H.; Gould, J.; Tamayo, P.; Mesirov, J.P. GSEA-P: A desktop application for Gene Set Enrichment Analysis. Bioinformatics 2007, 23, 3251–3253. [Google Scholar] [CrossRef]
- Benjamini, Y. Discovering the false discovery rate. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2010, 72, 405–416. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Elshorbagy, A.K.; Smith, A.D.; Kozich, V.; Refsum, H. Cysteine and obesity. Obesity (Silver Spring) 2012, 20, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H.; Dominy, J.E., Jr.; Lee, J.I.; Coloso, R.M. Mammalian cysteine metabolism: New insights into regulation of cysteine metabolism. J. Nutr. 2006, 136, 1652S–1659S. [Google Scholar] [CrossRef] [PubMed]
- Olsen, T.; Ovrebo, B.; Turner, C.; Bastani, N.E.; Refsum, H.; Vinknes, K.J. Combining Dietary Sulfur Amino Acid Restriction with Polyunsaturated Fatty Acid Intake in Humans: A Randomized Controlled Pilot Trial. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.; Jerneren, F.; Basta, M.; Basta, C.; Turner, C.; Khaled, M.; Refsum, H. Amino acid changes during transition to a vegan diet supplemented with fish in healthy humans. Eur. J. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Aasheim, E.T.; Elshorbagy, A.K.; My Diep, L.; Sovik, T.T.; Mala, T.; Valdivia-Garcia, M.; Olbers, T.; Bohmer, T.; Birkeland, K.I.; Refsum, H. Effect of bariatric surgery on sulphur amino acids and glutamate. Brit. J. Nutr. 2011. [Google Scholar] [CrossRef] [PubMed]
- Do, K.Q.; Mattenberger, M.; Streit, P.; Cuenod, M. In vitro release of endogenous excitatory sulfur-containing amino acids from various rat brain regions. J. Neurochem. 1986, 46, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Fisher-Wellman, K.H.; Neufer, P.D. Linking mitochondrial bioenergetics to insulin resistance via redox biology. Trends Endocrinol. Metab. 2012, 23, 142–153. [Google Scholar] [CrossRef]
- Goodpaster, B.H. Mitochondrial Deficiency Is Associated with Insulin Resistance. Diabetes 2013, 62, 1032. [Google Scholar] [CrossRef]
- Ushmorov, A.; Hack, V.; Droge, W. Differential reconstitution of mitochondrial respiratory chain activity and plasma redox state by cysteine and ornithine in a model of cancer cachexia. Cancer Res. 1999, 59, 3527–3534. [Google Scholar]
- Gazit, V.; Ben-Abraham, R.; Vofsi, O.; Katz, Y. L-cysteine increases glucose uptake in mouse soleus muscle and SH-SY5Y cells. Metab. Brain Dis. 2003, 18, 221–231. [Google Scholar] [CrossRef]
- John, G.B.; Shang, Y.; Li, L.; Renken, C.; Mannella, C.A.; Selker, J.M.L.; Rangell, L.; Bennett, M.J.; Zha, J. The Mitochondrial Inner Membrane Protein Mitofilin Controls Cristae Morphology. Mol. Biol. Cell 2005, 16, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Skovierova, H.; Vidomanova, E.; Mahmood, S.; Sopkova, J.; Drgova, A.; Cervenova, T.; Halasova, E.; Lehotsky, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Chung, H.Y.; Goto, S. Systemic adaptation to oxidative challenge induced by regular exercise. Free Radic. Biol. Med. 2008, 44, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen consumption and usage during physical exercise: The balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline | Post | %-Change | ||||
|---|---|---|---|---|---|---|
| Control (n = 13) | DG (n = 13) | Control (n = 13) | DG (n = 13) | Control (n = 13) | DG (n = 13) | |
| Body composition | ||||||
| Weight (kg) | 78.5 (8.2) | 95.4 (10.2) * | 78.3 (8.2) | 93.7 (9.7) * | −0.3 (2.1) | −1.7 (2.4) † |
| BMI (kg/m2) | 23.5 (2.0) | 29.0 (2.4) * | 23.5 (1.8) | 28.6 (2.3) * | 0.0 (2.0) | −1.2 (4.5) |
| FFM volume (kg) a | 34.9 (3.5) | 37.7 (5.0) | 37.1 (3.5) | 39.6 (5.1) | 6.4 (3.8) † | 5.3 (2.7) † |
| SAT (kg) a | 10.3 (2.7) | 18.0 (4.2) * | 9.7 (1.9) | 16.6 (3.7) * | −6.6 (9.2) † | −7.3 (6.0) † |
| IAAT (kg) a | 4.0 (2.0) | 8.8 (2.6) * | 3.2 (1.6) | 7.2 (2.8) * | −16.9 (15.1) † | −19.4 (10.8) † |
| Hepatic fat (AU) b | 2.8 (2.2) | 9.1 (5.9) * | 2.2 (2.4) | 6.5 (4.2) * | −23.3 (50.7) † | −27.4 (15.8) † |
| Thigh muscle area (AU) a | 20344.0 (2932.1) | 23993.5 (3071.7) * | 22233.2 (2572.5) | 25619.8 (2877.4) * | 9.7 (4.7) † | 7.1 (6.7) † |
| Physical fitness | ||||||
| VO2max (mL/kg/min) | 44.1 (4.4) | 37.1 (4.9) * | 49.8 (5.1) | 41.9 (5.0) * | 13.2 (9.7) † | 13.3 (7.7) † |
| Chest press (kg) | 65.6 (16.8) | 68.7 (13.7) | 77.1 (20.2) | 77.3 (12.7) | 18.4 (8.7) † | 13.6 (8.4) † |
| Pull down (kg) | 68.8 (9.3) | 75.6 (15.1) | 79.8 (9.6) | 85.2 (13.8) | 18.3 (10.1) † | 13.7 (7.3) † |
| Leg press (kg) | 199.6 (36.9) | 248.7 (30.3) * | 218.1 (37.9) | 278.3 (27.8) * | 9.8 (7.6) † | 12.5 (8.4) † |
| Glucose metabolism | ||||||
| HbA1c (mmol/mol) | 33 (4) | 37 (4) * | N.A. | N.A. | N.A. | N.A. |
| HbA1c (%) | 5.2 (0.2) | 5.5 (0.4) * | N.A. | N.A. | N.A. | N.A. |
| F-glucose (mmol/L) | 5.4 (0.5) | 5.9 (0.3) * | 5.5 (0.5) | 5.9 (0.2) * | 3.1 (4.5) † | 1.8 (6.8) |
| F-C-Peptide (pmol/L) | 588.0 (117.8) | 932.8 (248.9) * | 5.5 (0.5) | 5.9 (0.2) * | 7.3 (23.8) | 12.3 (45.3) |
| F-Insulin (pmol/L) | 38.5 (18.6) | 65.3 (27.1) * | 617.5 (124.3) | 976.6 (196.9) * | 15.1 (49.2) | 27.6 (66.2) |
| FFA (mmol/L) | 0.3 (0.1) | 0.2 (0.1) | 38.8 (12.0) | 77.0 (31.4) * | −21.7 (31.1) † | 16.0 (53.1) |
| GIR (mg/kg/min) | 7.6 (1.6) | 4.2 (1.8) * | 0.2 (0.0) | 0.2 (0.1) * | 37.8 (30.1) † | 44.4 (58.8) † |
| GIR (mg/kgFFM/min) | 18.5 (3.4) | 11.5 (5.2) * | 10.4 (2.6) | 5.4 (1.8) * | 32.4 (30.9) † | 39.0 (63.5) † |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Olsen, T.; Vinknes, K.J.; Refsum, H.; Gulseth, H.L.; Birkeland, K.I.; Drevon, C.A. Plasma Sulphur-Containing Amino Acids, Physical Exercise and Insulin Sensitivity in Overweight Dysglycemic and Normal Weight Normoglycemic Men. Nutrients 2019, 11, 10. https://doi.org/10.3390/nu11010010
Lee S, Olsen T, Vinknes KJ, Refsum H, Gulseth HL, Birkeland KI, Drevon CA. Plasma Sulphur-Containing Amino Acids, Physical Exercise and Insulin Sensitivity in Overweight Dysglycemic and Normal Weight Normoglycemic Men. Nutrients. 2019; 11(1):10. https://doi.org/10.3390/nu11010010
Chicago/Turabian StyleLee, Sindre, Thomas Olsen, Kathrine J. Vinknes, Helga Refsum, Hanne L. Gulseth, Kåre I. Birkeland, and Christian A. Drevon. 2019. "Plasma Sulphur-Containing Amino Acids, Physical Exercise and Insulin Sensitivity in Overweight Dysglycemic and Normal Weight Normoglycemic Men" Nutrients 11, no. 1: 10. https://doi.org/10.3390/nu11010010
APA StyleLee, S., Olsen, T., Vinknes, K. J., Refsum, H., Gulseth, H. L., Birkeland, K. I., & Drevon, C. A. (2019). Plasma Sulphur-Containing Amino Acids, Physical Exercise and Insulin Sensitivity in Overweight Dysglycemic and Normal Weight Normoglycemic Men. Nutrients, 11(1), 10. https://doi.org/10.3390/nu11010010