Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle?

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
Statistical Analysis
3. Results
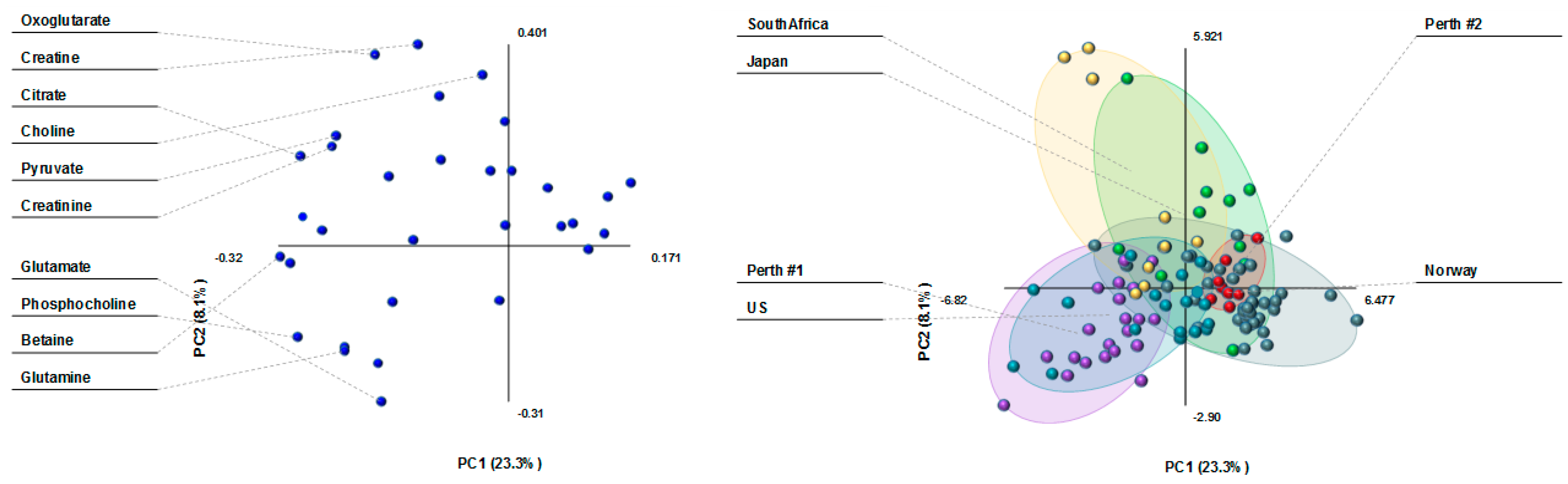
3.1. Milk Metabolite Clusters by Country
3.2. Milk Metabolite Differences in Healthy, Non-Atopic Women
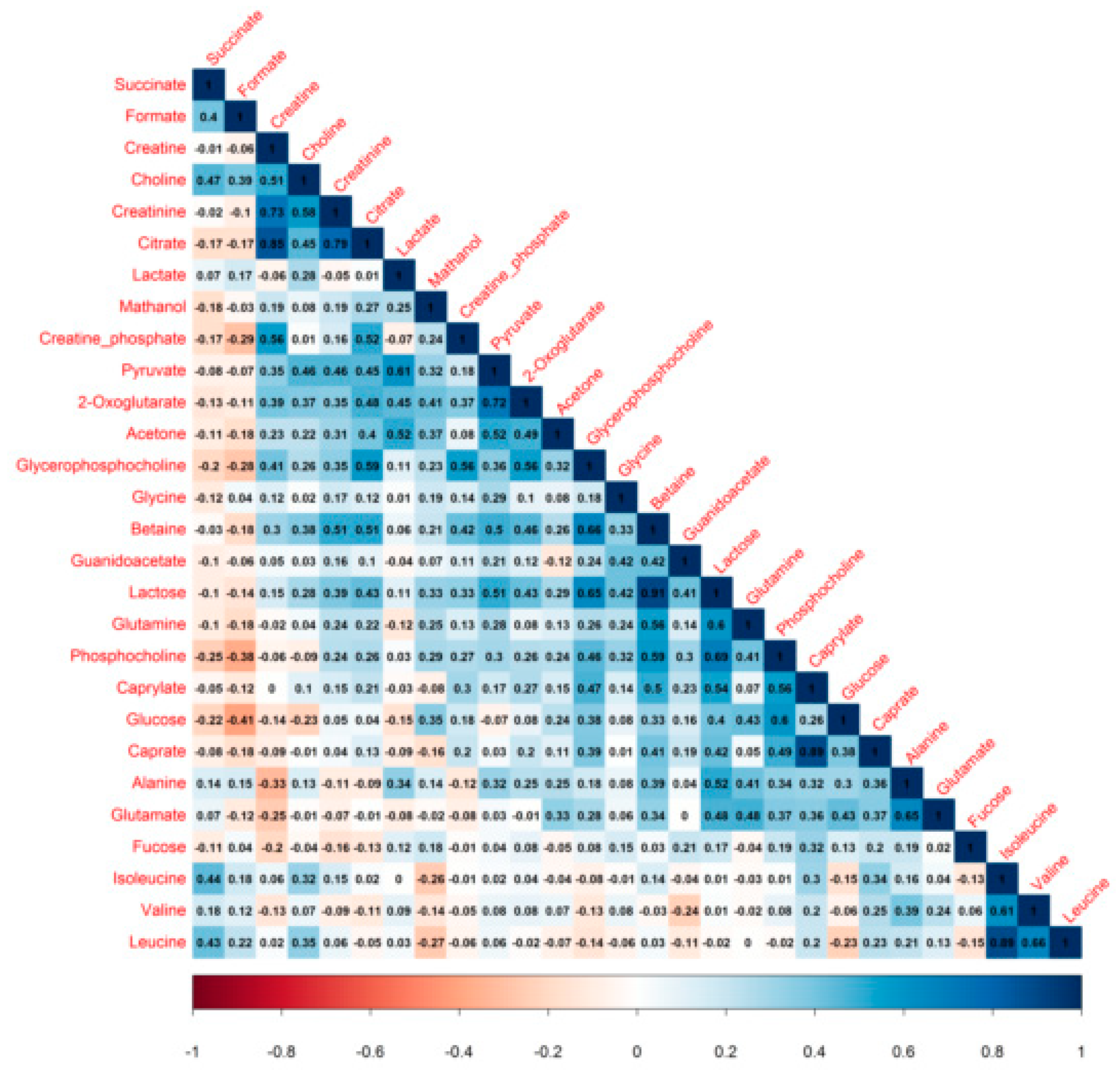
3.3. Milk Metabolite Correlations
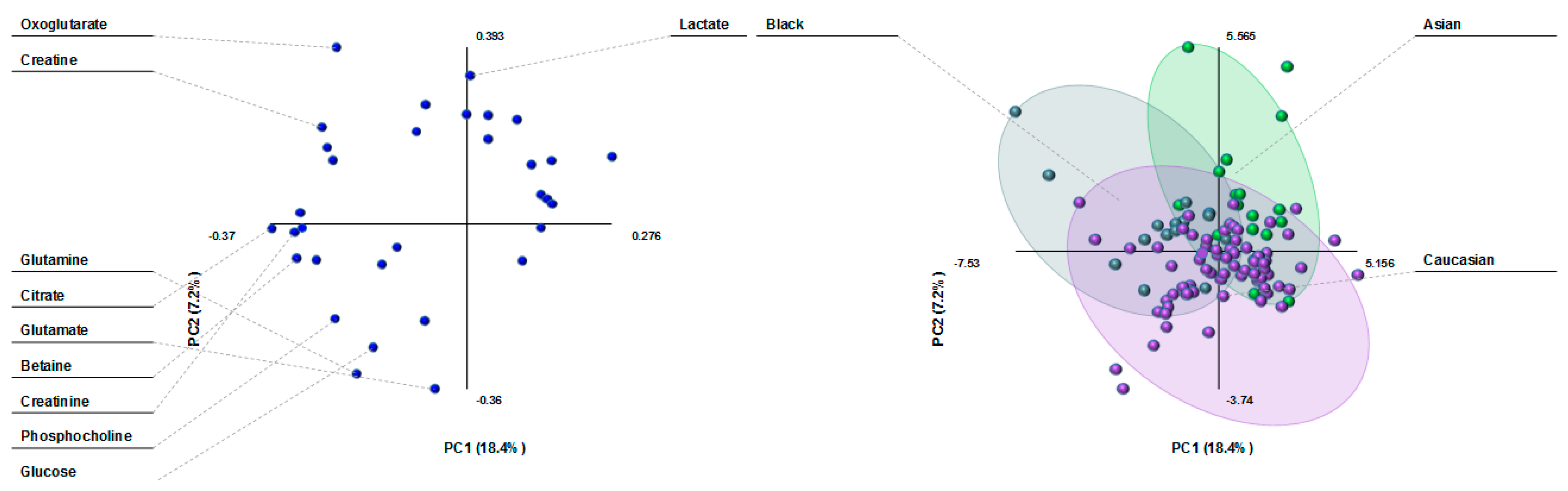
3.4. Milk Metabolite Clusters by Race/Ethnicity
3.5. Milk Metabolite Variation by Ethnicity in Healthy, Non-Atopic Women
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hennet, T.; Borsig, L. Breastfed at tiffany’s. Trends Biochem. Sci. 2016, 41, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Demmelmair, H.; Koletzko, B. Variation of metabolite and hormone contents in human milk. Clin. Perinatol. 2017, 44, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lonnerdal, B.; Slupsky, C.M. The human milk metabolome reveals diverse oligosaccharide profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; du Toit, E.; Kulkarni, A.; Aakko, J.; Linderborg, K.M.; Zhang, Y.; Nicol, M.P.; Isolauri, E.; Yang, B.; Collado, M.C.; et al. Distinct patterns in human milk microbiota and fatty acid profiles across specific geographic locations. Front. Microbiol. 2016, 7, 1619. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, X.; Zhou, B.; Jiang, A.C.; Chai, L. An updated review of worldwide levels of docosahexaenoic and arachidonic acid in human breast milk by region. Public Health Nutr. 2016, 19, 2675–2687. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Adair, L.; Fall, C.; Hallal, P.C.; Martorell, R.; Richter, L.; Sachdev, H.S. Maternal and child undernutrition: Consequences for adult health and human capital. Lancet 2008, 371, 340–357. [Google Scholar] [CrossRef]
- Adair, L.S.; Fall, C.H.; Osmond, C.; Stein, A.D.; Martorell, R.; Ramirez-Zea, M.; Sachdev, H.S.; Dahly, D.L.; Bas, I.; Norris, S.A.; et al. Associations of linear growth and relative weight gain during early life with adult health and human capital in countries of low and middle income: Findings from five birth cohort studies. Lancet 2013, 382, 525–534. [Google Scholar] [CrossRef]
- Garza, C.; Borghi, E.; Onyango, A.W.; de Onis, M. Parental height and child growth from birth to 2 years in the WHO multicentre growth reference study. Matern. Child Nutr. 2013, 9, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Bahl, R.; Barros, A.J.; Franca, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- Munblit, D.; Peroni, D.G.; Boix-Amoros, A.; Hsu, P.S.; Van’t Land, B.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C.; et al. Human milk and allergic diseases: An unsolved puzzle. Nutrients 2017, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.C.; Totten, S.M.; Huang, J.O.; Nagshbandi, S.; Kirmiz, N.; Garrido, D.A.; Lewis, Z.T.; Wu, L.D.; Smilowitz, J.T.; German, J.B.; et al. Identification of oligosaccharides in feces of breast-fed infants and their correlation with the gut microbial community. Mol. Cell Proteomics 2016, 15, 2987–3002. [Google Scholar] [CrossRef] [PubMed]
- Koleva, P.T.; Bridgman, S.L.; Kozyrskyj, A.L. The infant gut microbiome: Evidence for obesity risk and dietary intervention. Nutrients 2015, 7, 2237–2260. [Google Scholar] [CrossRef] [PubMed]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef] [PubMed]
- Wahlqvist, M.L. Lactose nutrition in lactase nonpersisters. Asia Pac. J. Clin. Nutr. 2015, 24, S21–S25. [Google Scholar] [PubMed]
- Taylor, A.L.; Dunstan, J.A.; Prescott, S.L. Probiotic supplementation for the first 6 months of life fails to reduce the risk of atopic dermatitis and increases the risk of allergen sensitization in high-risk children: A randomized controlled trial. J. Allergy Clin. Immunol. 2007, 119, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Gridneva, Z.; Kugananthan, S.; Hepworth, A.R.; Tie, W.J.; Lai, C.T.; Ward, L.C.; Hartmann, P.E.; Geddes, D.T. Effect of human milk appetite hormones, macronutrients, and infant characteristics on gastric emptying and breastfeeding patterns of term fully breastfed infants. Nutrients 2016, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Wegienka, G.; Havstad, S.; Joseph, C.L.; Zoratti, E.; Ownby, D.; Woodcroft, K.; Johnson, C.C. Racial disparities in allergic outcomes in african americans emerge as early as age 2 years. Clin. Exp. Allergy 2012, 42, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Eggesbo, M.; Thomsen, C.; Jorgensen, J.V.; Becher, G.; Odland, J.O.; Longnecker, M.P. Associations between brominated flame retardants in human milk and thyroid-stimulating hormone (TSH) in neonates. Environ. Res. 2011, 111, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Package “Corrplot”. Available online: https://cran.r-project.org/web/packages/corrplot/corrplot.pdf. (accessed on 23 August 2018).
- Spevacek, A.R.; Smilowitz, J.T.; Chin, E.L.; Underwood, M.A.; German, J.B.; Slupsky, C.M. Infant maturity at birth reveals minor differences in the maternal milk metabolome in the first month of lactation. J. Nutr. 2015, 145, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Adelman, A.S.; Rai, D.; Boettcher, J.; Lonnerdal, B. Amino acid profiles in term and preterm human milk through lactation: A systematic review. Nutrients 2013, 5, 4800–4821. [Google Scholar] [CrossRef] [PubMed]
- Prentice, P.; Ong, K.K.; Schoemaker, M.H.; van Tol, E.A.; Vervoort, J.; Hughes, I.A.; Acerini, C.L.; Dunger, D.B. Breast milk nutrient content and infancy growth. Acta Paediatr. 2016, 105, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Mardinoglu, A.; Nielsen, J. Predicting growth of the healthy infant using a genome scale metabolic model. NPJ Syst. Biol. Appl. 2017, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Sachse, D.; Baerug, A.; Sletner, L.; Birkeland, K.I.; Nakstad, B.; Jenum, A.K.; Berg, J.P. Urine nmr metabolomics analysis of breastfeeding biomarkers during and after pregnancy in a large prospective cohort study. Scand. J. Clin. Lab. Investig. 2014, 74, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Zhai, Z.; Ni, X.; Wang, H.; Ji, Y.; Tang, T.; Ren, W.; Long, H.; Deng, B.; Deng, J.; et al. Metabolomic profiles reveal potential factors that correlate with lactation performance in sow milk. Sci. Rep. 2018, 8, 10712. [Google Scholar] [CrossRef] [PubMed]
- Boix-Amoros, A.; Collado, M.C.; Mira, A. Relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 2016, 7, 492. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, A.; Kai-Larsen, Y.; Printz, G.; Yoshio, H.; Alvelius, G.; Lagercrantz, H.; Stromberg, R.; Jornvall, H.; Gudmundsson, G.H.; Agerberth, B. Lactose in human breast milk an inducer of innate immunity with implications for a role in intestinal homeostasis. PLoS ONE 2013, 8, e53876. [Google Scholar] [CrossRef] [PubMed]
- Cocinero, E.J.; Carcabal, P. Carbohydrates. Top. Curr. Chem. 2015, 364, 299–333. [Google Scholar] [PubMed]
- Xi, X.; Kwok, L.Y.; Wang, Y.; Ma, C.; Mi, Z.; Zhang, H. Ultra-performance liquid chromatography-quadrupole-time of flight mass spectrometry ms(e)-based untargeted milk metabolomics in dairy cows with subclinical or clinical mastitis. J. Dairy Sci. 2017, 100, 4884–4896. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, D.; Blomberg, M. Prevalence of postpartum infections: A population-based observational study. Acta. Obstet. Gynecol. Scand. 2014, 93, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Z.; Shi, K.; Wu, X.H.; Xue, M.Y.; Wei, Z.H.; Liu, J.X.; Liu, H.Y. Lactation-related metabolic mechanism investigated based on mammary gland metabolomics and 4 biofluids’ metabolomics relationships in dairy cows. BMC Genom. 2017, 18, 936. [Google Scholar] [CrossRef] [PubMed]
- Dorokhov, Y.L.; Shindyapina, A.V.; Sheshukova, E.V.; Komarova, T.V. Metabolic methanol: Molecular pathways and physiological roles. Physiol. Rev. 2015, 95, 603–644. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.M.; da Costa, K.A.; Galanko, J.; Sha, W.; Stephenson, B.; Vick, J.; Zeisel, S.H. Choline intake and genetic polymorphisms influence choline metabolite concentrations in human breast milk and plasma. Am. J. Clin. Nutr. 2010, 92, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Balzer, M.S.; Gross, M.M.; Lichtinghagen, R.; Haller, H.; Schmitt, R. Got milk? Breastfeeding and milk analysis of a mother on chronic hemodialysis. PLoS ONE 2015, 10, e0143340. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.E.; Rezamand, P.; Williams, J.E.; Price, W.; Chahine, M.; McGuire, M.A. Effects of dietary betaine on milk yield and milk composition of mid-lactation holstein dairy cows. J. Dairy Sci. 2012, 95, 6557–6562. [Google Scholar] [CrossRef] [PubMed]
- Manso, H.E.; Filho, H.C.; de Carvalho, L.E.; Kutschenko, M.; Nogueira, E.T.; Watford, M. Glutamine and glutamate supplementation raise milk glutamine concentrations in lactating gilts. J. Anim. Sci. Biotechnol. 2012, 3, 2. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sun, H.; Xue, M.; Wang, D.; Guan, L.L.; Liu, J. Serum metabolome profiling revealed potential biomarkers for milk protein yield in dairy cows. J. Proteomics 2018, 184, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Dangat, K.; Upadhyay, D.; Kilari, A.; Sharma, U.; Kemse, N.; Mehendale, S.; Lalwani, S.; Wagh, G.; Joshi, S.; Jagannathan, N.R. Altered breast milk components in preeclampsia; an in-vitro proton nmr spectroscopy study. Clin. Chim. Acta 2016, 463, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.E.; Carrothers, J.M.; Lackey, K.A.; Beatty, N.F.; York, M.A.; Brooker, S.L.; Shafii, B.; Price, W.J.; Settles, M.L.; McGuire, M.A.; et al. Human milk microbial community structure is relatively stable and related to variations in macronutrient and micronutrient intakes in healthy lactating women. J. Nutr. 2017, 147, 1739–1748. [Google Scholar] [PubMed]
- Munblit, D.; Treneva, M.; Peroni, D.G.; Colicino, S.; Chow, L.; Dissanayeke, S.; Abrol, P.; Sheth, S.; Pampura, A.; Boner, A.L.; et al. Colostrum and mature human milk of women from London, Moscow, and Verona: Determinants of immune composition. Nutrients 2016, 8, e695. [Google Scholar] [CrossRef] [PubMed]
- Budree, S.; Goddard, E.; Brittain, K.; Cader, S.; Myer, L.; Zar, H.J. Infant feeding practices in a south African birth cohort-a longitudinal study. Matern. Child Nutr. 2017, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Osgood-Zimmerman, A.; Millear, A.I.; Stubbs, R.W.; Shields, C.; Pickering, B.V.; Earl, L.; Graetz, N.; Kinyoki, D.K.; Ray, S.E.; Bhatt, S.; et al. Mapping child growth failure in Africa between 2000 and 2015. Nature 2018, 555, 41–47. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEAN (SD) | |||||
|---|---|---|---|---|---|
| Metabolites/Countries | South Africa | Japan | Norway | US | Perth #2 |
| 2-Oxoglutarate | 79.6 (25.6) | 79.1 (49.2) ## | 30.5 (31.2) *** | 39.0 (12.7) * | 47.5 (19.9) |
| Acetone | 13.2 (6.4) | 19.8 (15.7) | 10.8 (7.6) | 11.4 (3.3) | 13.3 (7.5) |
| Alanine | 161.0 (93.5) | 233.6 (96.8) | 208.1 (84.5) | 240.1 (91.9) | 203.2 (48.4) |
| Betaine | 668.1 (201.8) | 355.3 (367.1) | 180.4 (318.8) *** | 408.4 (567.8) | 58.8 (6.6) *** |
| Caprate | 108.6 (124.7) | 193.1 (216.3) | 121.5 (120.3) | 60.0 (90.6) | 114.0 (42.9) |
| Caprylate | 157.1 (137.2) | 357.6 (497.0) | 131.6 (207.1) | 92.7 (132.5) | 82.7 (47.1) |
| Choline | 322.1 (194.9) | 192.0 (128.3) | 184.1 (163.7) | 133.0 (77.0) | 138.5 (71.1) |
| Citrate | 5614 (3920) | 2830 (1313) * | 2718 (1361) *** | 3642 (1272) | 2856 (836.1) * |
| Creatine | 274.7 (342.9) | 60.3 (28.4) * | 63.9 (26.9) *** | 56.8 (23.4) * | 66.5 (19.0) * |
| Creatine phosphate | 49.6 (54.3) | 45.2 (31.1) | 20.6 (13.1) * | 42.0 (12.7) | 23.9 (7.6) |
| Creatinine | 108.8 (82.7) | 42.9 (15.4) ** | 43.7 (15.6) *** | 57.3 (22.9) * | 45.0 (5.7) ** |
| Formate | 83.4 (79.4) | 112.2 (127.8) | 902.0 (1695.2) | 91.9 (55.7) | 127.1 (24.9) |
| Fucose | 278.5 (373.5) | 570.9 (482.9) | 330.5 (252.5) | 344.4 (216.4) | 382.3 (277.5) |
| Glucose | 1347 (852.9) | 1563 (1327) | 926.4 (747.8) | 1697 (413.5) | 1699 (935.9) |
| Glutamate | 858.9 (491.3) | 1296 (565.8) | 1467 (835.2) | 1573 (830.2) | 1554 (369.4) |
| Glutamine | 282.0 (150.9) | 103.0 (88.1) | 101.9 (119.0) | 514.2 (618.2) ### | 207.6 (178.2) |
| Glycine | 2796 (759.9) | 3135 (1373) | 2512 (1191) | 1641 (1777) | 1919 (949.5) |
| Guanidoacetate | 4771 (1048) | 3777 (1747) | 3373 (2944) | 3435 (1770) | 1622 (286.5) * |
| Isoleucine | 20.8 (12.2) | 24.1 (25.9) | 24.2 (21.1) | 10.9 (2.6) | 12.0 (4.0) |
| Lactate | 213.7 (63.1) | 3215 (5561) **## | 443.5 (693.6) | 137.8 (102.0) | 125.7 (41.5) |
| Lactose | 189,874 (34,234) | 139,161 (83,333) | 101,351 (84,404) * | 161,160 (123,638) | 83,524 (23,509) * |
| Leucine | 33.7 (21.1) | 49.9 (61.2) | 66.2 (76.2) | 30.9 (8.9) | 34.0 (7.8) |
| Methanol | 78.0 (17.6) | 97.7 (49.2) ### | 46.1 (19.3) *** | 89.2 (6.7) ### | 72.0 (6.8) # |
| Phosphocholine | 636.9 (328.6) | 784.7 (470.6) | 463.7 (326.7) | 698.3 (231.2) | 488.4 (145.6) |
| Pyruvate | 53.3 (29.5) | 68.6 (69.8) ## | 20.0 (26.3) * | 31.7 (39.7) | 6.3 (2.5) * |
| Succinate | 48.3 (67.0) | 47.9 (25.9) | 210.8 (515.7) | 21.2 (9.6) | 49.3 (15.7) |
| Valine | 35.3 (11.9) | 63.9 (33.5) | 65.6 (32.7) * | 50.2 (21.1) | 57.4 (17.6) |
| Glycerophosphocholine | 797.6 (439.3) | 645.0 (393.0) | 414.9 (346.0) * | 589.2 (239.7) | 517.4 (118.5) |
| # metabolite differences with South Africa | na | 4 | 11 | 3 | 7 |
| # metabolite differences with Norway | - | 4 | na | 2 | 1 |
| MEAN (SD) | |||
|---|---|---|---|
| Metabolites/Countries | Black | Caucasian | Asian |
| 2-Oxoglutarate | 75.4 (31.1) | 37.6 (29.2) ** | 55.3 (47.1) |
| Acetone | 12.9 (5.3) | 11.9 (7.4) | 14.2 (13.0) |
| Alanine | 164.4 (87.9) | 210.6 (82.1) | 222.3 (90.9) |
| Betaine | 581.1 (283.6) | 204.9 (352.7) * | 354.3 (390.1) |
| Caprate | 77.2 (87.8) | 109.8 (112.3) | 185.1 (168.3) |
| Caprylate | 134.7 (136.2) | 118.1 (184.1) | 266.9 (375.3) |
| Choline | 285.1 (167.0) | 170.2 (154.4) | 218.7 (145.5) |
| Citrate | 4892 (3687) | 3123 (1833) | 2776 (1061) |
| Creatine | 229.4 (326.5) | 77.6 (101.1) * | 63.2 (28.5) * |
| Creatine phosphate | 54.0 (56.2) | 24.1 (13.7) ** | 35.9 (26.8) |
| Creatinine | 89.7 (68.2) | 51.6 (37.0) * | 44.2 (15.0) * |
| Formate | 86.5 (79.3) | 582.6 (1363) | 534.5 (1350) |
| Fucose | 328.0 (376.9) | 306.0 (236.6) # | 556.4 (391.5) |
| Glucose | 1264 (672.2) | 1263 (866.0) | 1148 (1083) |
| Glutamate | 1021 (614.5) | 1478 (791.6) | 1272 (536.3) |
| Glutamine | 252.4 (122.0) | 204.7 (310.6) | 89.5 (79.2) |
| Glycine | 2646 (1051) | 2360 (1307) | 2596 (1169) |
| Guanidoacetate | 4405 (1378) | 2912 (2538) | 4544 (1916) |
| Isoleucine | 17.3 (11.5) | 20.5 (18.4) | 23.4 (21.8) |
| Lactate | 198.2 (82.2) | 339.5 (600.5) # | 1918 (4213) |
| Lactose | 178,365 (50,934) | 110,060 (86,787) | 131,284 (85,509) |
| Leucine | 31.5 (14.8) | 52.1 (56.4) | 64.0 (87.0) |
| Methanol | 80.4 (15.7) | 59.3 (23.6) | 71.0 (46.6) |
| Phosphocholine | 584.6 (305.1) | 521.9 (304.1) | 642.5 (435.6) |
| Pyruvate | 46.5 (25.3) | 21.4 (29.6) | 47.2 (58.8) |
| Succinate | 36.1 (53.0) | 152.7 (446.7) | 100.8 (124.4) |
| Valine | 34.6 (12.0) | 61.1 (26.2) * | 63.9 (40.5) |
| Glycerophosphocholine | 846.4 (424.6) | 467.7 (319.9) ** | 518.9 (327.7) |
| # metabolite differences with Black race | na | 7 | 2 |
| # metabolite differences with Asian race | 3 | na | 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gay, M.C.L.; Koleva, P.T.; Slupsky, C.M.; Toit, E.D.; Eggesbo, M.; Johnson, C.C.; Wegienka, G.; Shimojo, N.; Campbell, D.E.; Prescott, S.L.; et al. Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients 2018, 10, 1151. https://doi.org/10.3390/nu10091151
Gay MCL, Koleva PT, Slupsky CM, Toit ED, Eggesbo M, Johnson CC, Wegienka G, Shimojo N, Campbell DE, Prescott SL, et al. Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients. 2018; 10(9):1151. https://doi.org/10.3390/nu10091151
Chicago/Turabian StyleGay, Melvin C. L., Petya T. Koleva, Carolyn M. Slupsky, Elloise Du Toit, Merete Eggesbo, Christine C. Johnson, Ganesa Wegienka, Naoki Shimojo, Dianne E. Campbell, Susan L. Prescott, and et al. 2018. "Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle?" Nutrients 10, no. 9: 1151. https://doi.org/10.3390/nu10091151
APA StyleGay, M. C. L., Koleva, P. T., Slupsky, C. M., Toit, E. D., Eggesbo, M., Johnson, C. C., Wegienka, G., Shimojo, N., Campbell, D. E., Prescott, S. L., Munblit, D., Geddes, D. T., Kozyrskyj, A. L., & InVIVO LactoActive Study Investigators. (2018). Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients, 10(9), 1151. https://doi.org/10.3390/nu10091151