Consumption of Cherry out of Season Changes White Adipose Tissue Gene Expression and Morphology to a Phenotype Prone to Fat Accumulation



Abstract
1. Introduction
2. Materials and Methods
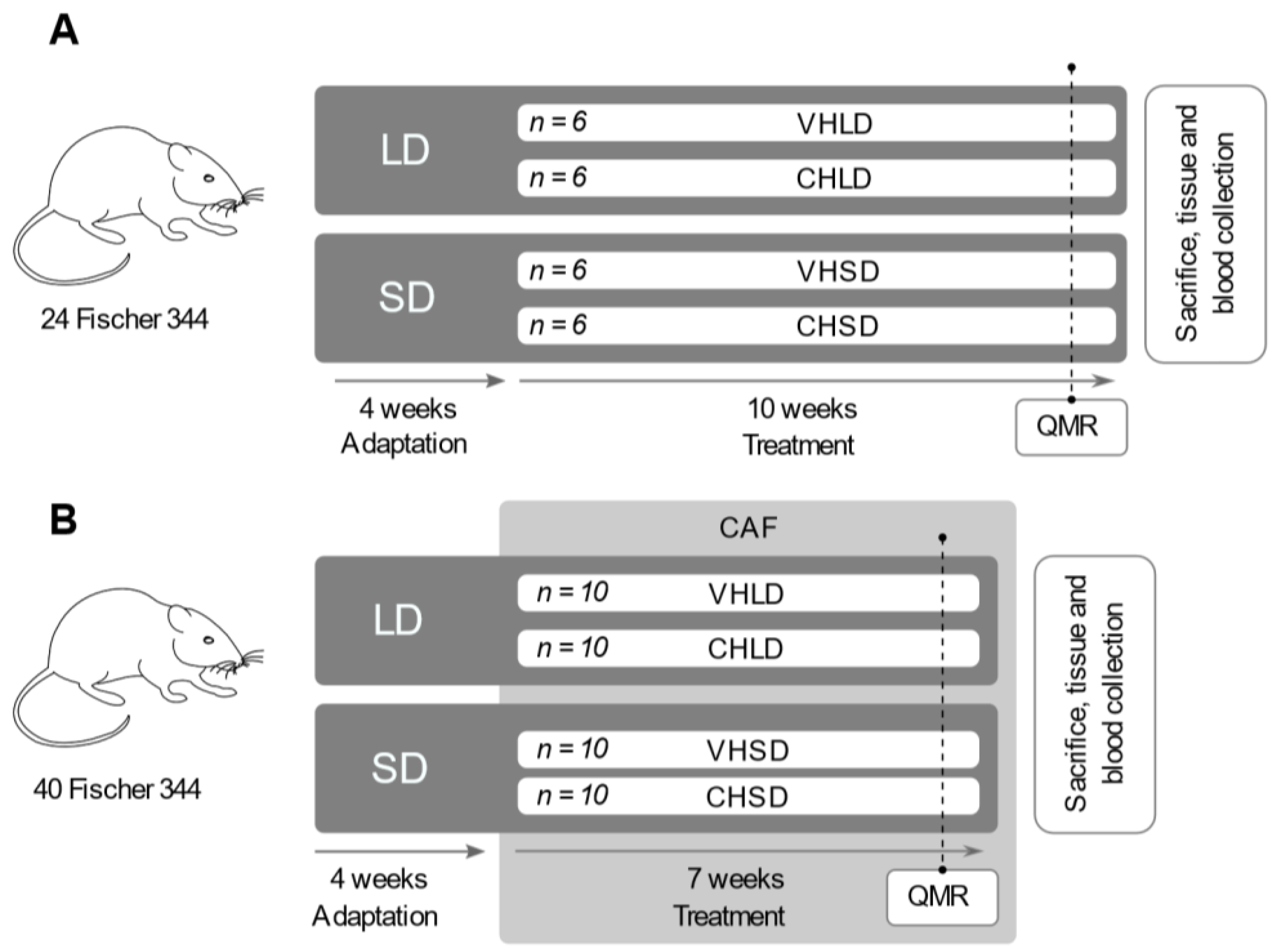
2.1. Treatments
2.2. Animal Experimental Procedure
2.3. Plasma Analysis
2.4. RNA Extraction and Quantification by Real-Time qRT-PCR
2.5. Histology
2.6. Statistical Analysis
3. Results
3.1. Biometric Parameters
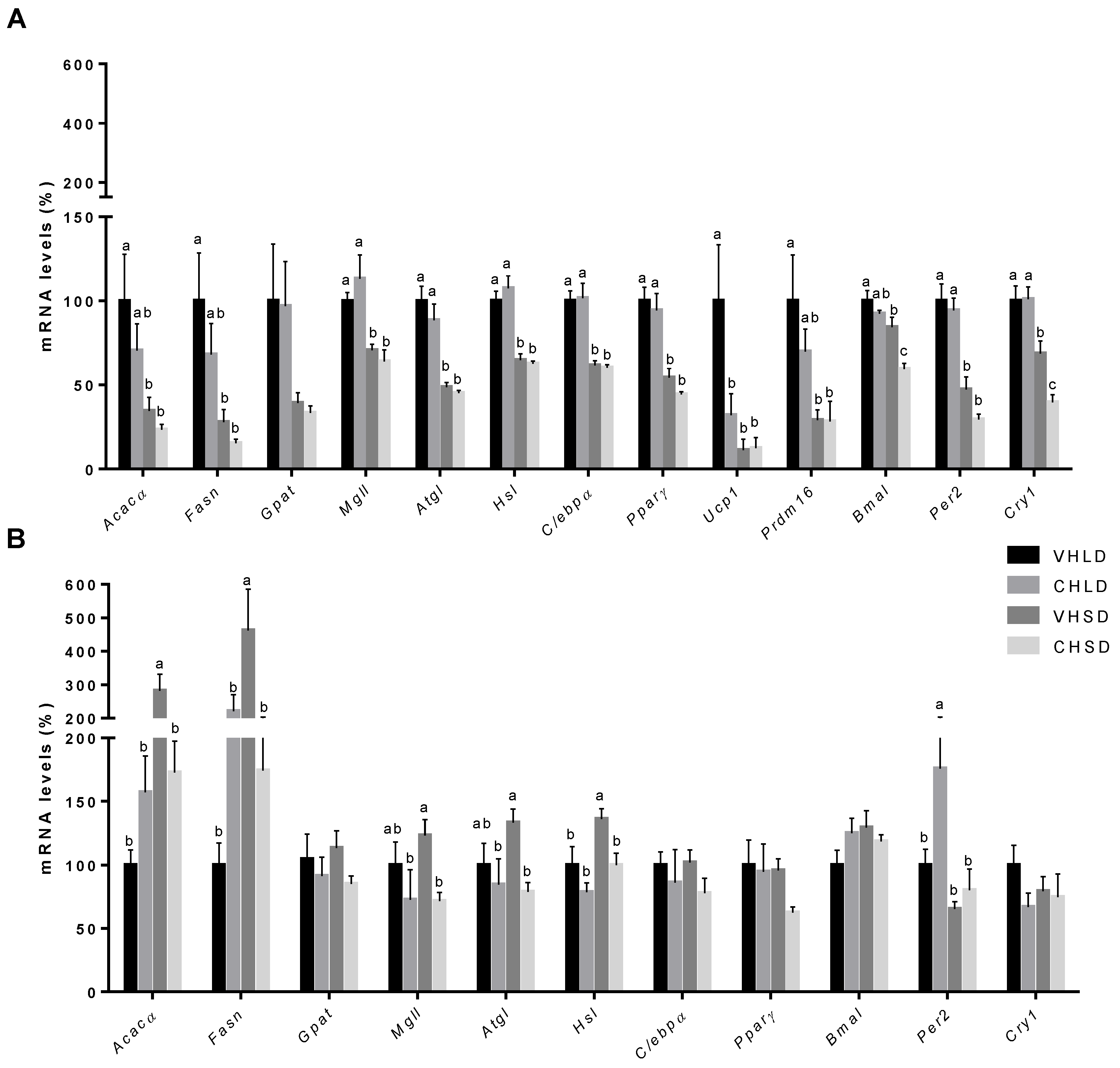
3.2. RWAT Gene Expression
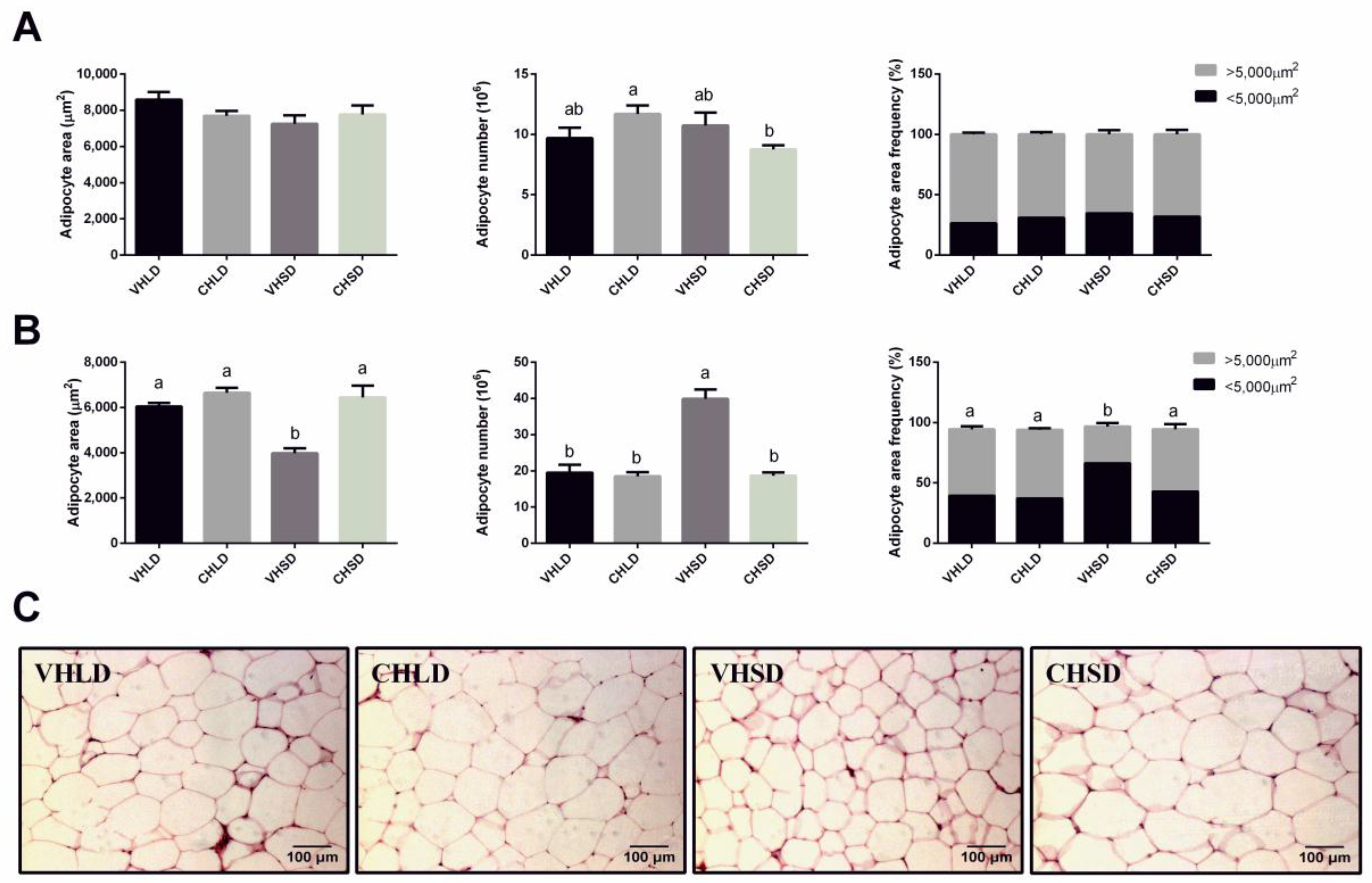
3.3. RWAT Histology
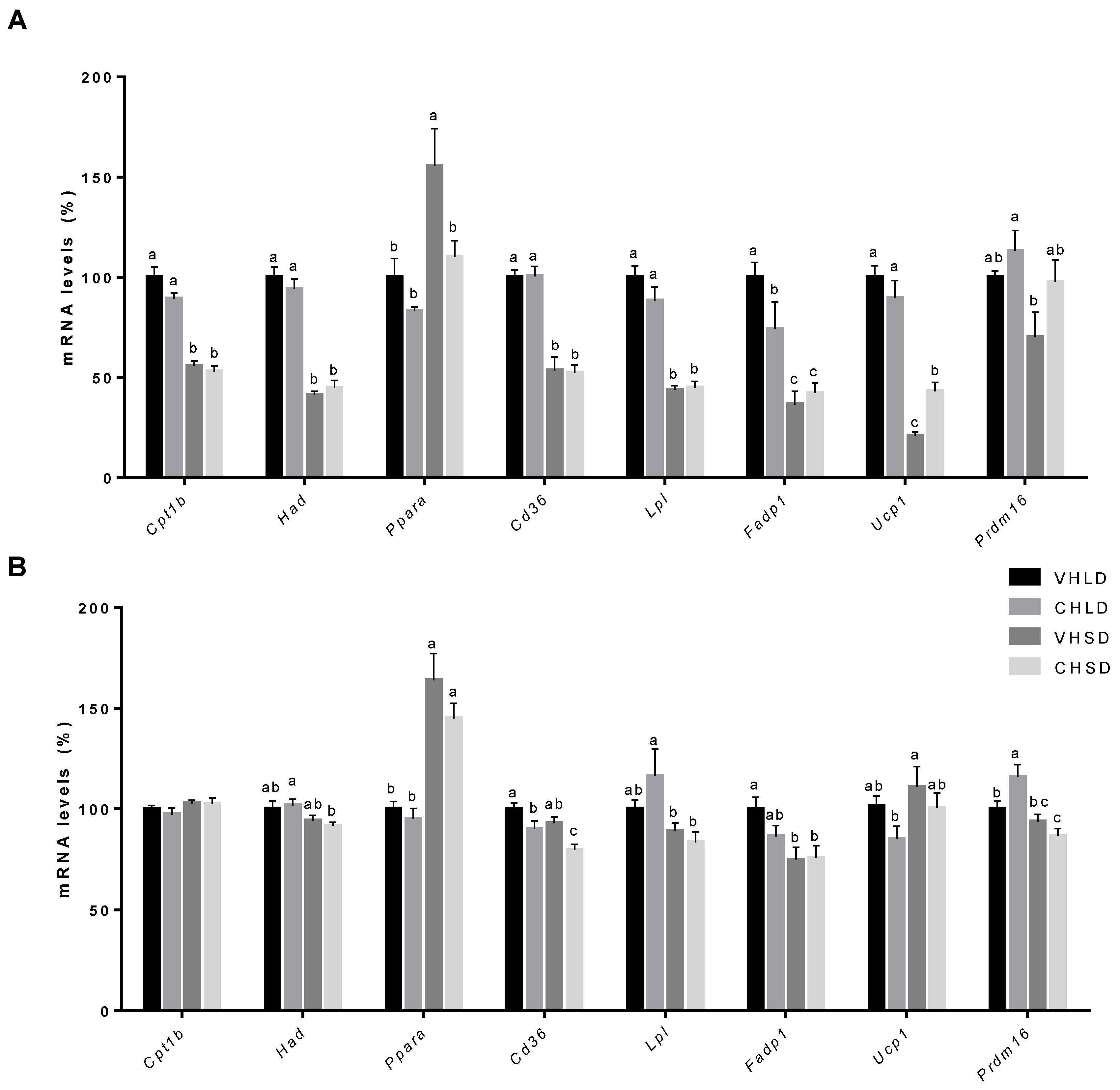
3.4. BAT Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Symonds, M.E.; Sebert, S.; Budge, H. The obesity epidemic: From the environment to epigenetics-not simply a response to dietary manipulation in a thermoneutral environment. Front. Genet. 2011, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Keith, S.W.; Redden, D.T.; Katzmarzyk, P.T.; Boggiano, M.M.; Hanlon, E.C.; Benca, R.M.; Ruden, D.; Pietrobelli, A.; Barger, J.L.; Fontaine, K.R.; et al. Putative contributors to the secular increase in obesity: Exploring the roads less traveled. Int. J. Obes. 2006, 30, 1585–1594. [Google Scholar] [CrossRef] [PubMed]
- Bartness, T.J.; Demas, G.E.; Song, C.K. Seasonal Changes in Adiposity: The Roles of the Photoperiod, Melatonin and Other Hormones, and Sympathetic Nervous System. Exp. Biol. Med. 2002, 227, 363–376. [Google Scholar] [CrossRef]
- Gibert-Ramos, A.; Salvadó, M.J.; Ibars, M.; Crescenti, A. Response to the photoperiod in the white and brown adipose tissues of Fischer 344 rats fed a standard or cafeteria diet. J. Nutr. Biochem. submitted.
- Kanikowska, D.; Sato, M.; Witowski, J. Contribution of daily and seasonal biorhythms to obesity in humans. Int. J. Biometeorol. 2015, 59, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.; Peiser, B. Seasonal variations in health-related human physical activity. Sports Med. 2006, 36, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Küller, R. The influence of light on circarhythms in humans. J. Physiol. Anthropol. Appl. Hum. Sci. 2002, 21, 87–91. [Google Scholar] [CrossRef]
- Goldman, B.D. Mammalian Photoperiodic System: Of Photoperiodic Time Measurement. J. Biol. Rhythm. 2001, 16, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Peek, C.B.; Ramsey, K.M.; Marceva, B.; Bass, J. Nutrient Sensing and the Circadian Clock. Trends Endocrinol. Metab. 2012, 23, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Zucker, I.; Schwartz, W.J. Tracking the seasons: The internal calendars of vertebrates. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 341–361. [Google Scholar] [CrossRef] [PubMed]
- Zvonic, S.; Ptitsyn, A.A.; Conrad, S.A.; Scott, L.K.; Floyd, Z.E.; Kilroy, G.; Wu, X.; Goh, B.C.; Mynatt, R.L.; Gimble, J.M. Characterization of peripheral circadian clocks in adipose tissues. Diabetes 2006, 55, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Sumová, A.; Jáč, M.; Sládek, M.; Šauman, I.; Illnerová, H. Clock Gene Daily Profiles and Their Phase Relationship in the Rat Suprachiasmatic Nucleus Are Affected by Photoperiod. J. Biol. Rhythm. 2003, 18, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Engel, L.; Lorenzkowski, V.; Langer, C.; Rohleder, N.; Spessert, R. The photoperiod entrains the molecular clock of the rat pineal. Eur. J. Neurosci. 2005, 21, 2297–2304. [Google Scholar] [CrossRef] [PubMed]
- Zelinski, E.L.; Deibel, S.H.; McDonald, R.J. The trouble with circadian clock dysfunction: Multiple deleterious effects on the brain and body. Neurosci. Biobehav. Rev. 2014, 40, 80–101. [Google Scholar] [CrossRef] [PubMed]
- Wyse, C.A.; Selman, C.; Page, M.M.; Coogan, A.N.; Hazlerigg, D.G. Circadian desynchrony and metabolic dysfunction; did light pollution make us fat? Med. Hypotheses 2011, 77, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Delezie, J.; Challet, E. Interactions between metabolism and circadian clocks: Reciprocal disturbances. Ann. N. Y. Acad. Sci. 2011, 1243, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, J.; Pévet, P.; Challet, E. High-fat feeding alters the clock synchronization to light. J. Physiol. 2008, 586, 5901–5910. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Latre, A.; Baselga-Escudero, L.; Casanova, E.; Arola-Arnal, A.; Salvadó, M.J.; Arola, L.; Bladé, C. Chronic consumption of dietary proanthocyanidins modulates peripheral clocks in healthy and obese rats. J. Nutr. Biochem. 2015, 26, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. What is Xenohormesis? Am. J. Pharmacol. Toxicol. 2008, 3, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Pineli, L.D.L.; Moretti, C.L.; Rodrigues, J.S.; Ferreira, D.B.; Chiarello, M.D. Variations in antioxidant properties of strawberries grown in Brazilian savannah and harvested in different seasons. J. Sci. Food Agric. 2012, 92, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Weingart, G.; Schoedl-Hummel, K.; Neumann, N.; Becker, M.; Varmuza, K.; Liebner, F.; Schuhmacher, R.; Forneck, A. Severe drought stress is affecting selected primary metabolites, polyphenols, and volatile metabolites in grapevine leaves (Vitis vinifera cv. Pinot noir). Plant Physiol. Biochem. 2015, 88, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.M. The Fat Controller: Adipocyte Development. PLoS Biol. 2012, 10, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-X.; Zhao, X.-Y.; Lin, J.D. The brown fat secretome: Metabolic functions beyond thermogenesis. Trends Endocrinol. Metab. 2015, 26, 231–237. [Google Scholar] [CrossRef] [PubMed]
- US Department of Agriculture, Agricultural Research Service. N.D.L. USDA Food Composition Databases. Available online: https://ndb.nal.usda.gov/ndb/ (accessed on 8 June 2018).
- Vos, F.; Crespy, V.; Chaffaut, L.; Mennen, L.; Knox, C.; Neveu, V. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, 1–14. [Google Scholar] [CrossRef]
- WHO. Food Based Dietary Guidelines in the WHO European Region Nutrition and Food Security Programme WHO Regional Office for Europe; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- WHO. WHO Calls on Countries to Reduce Sugars Intake Among Adults and Children; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Baselga-Escudero, L.; Pascual-Serrano, A.; Ribas-Latre, A.; Casanova, E.; Salvadó, M.J.; Arola, L.; Arola-Arnal, A.; Bladé, C. Long-term supplementation with a low dose of proanthocyanidins normalized liver miR-33a and miR-122 levels in high-fat diet-induced obese rats. Nutr. Res. 2015, 35, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Smith, M.; Karthikeyan, S.; Jeffers, M.S.; Janik, R.; Thomason, L.A.; Stefanovic, B.; Corbett, D. A physiological characterization of the Cafeteria diet model of metabolic syndrome in the rat. Physiol. Behav. 2016, 167, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Lairon, D.; Lopez-Miranda, J.; Williams, C. Methodology for studying postprandial lipid metabolism. Eur. J. Clin. Nutr. 2007, 61, 1145–1161. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.G.; Poppitt, S.D.; Minihane, A.M. Postprandial lipemia and cardiovascular disease risk: Interrelationships between dietary, physiological and genetic determinants. Atherosclerosis 2012, 220, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N.; Shadid, S.; Hamlani, R.; Humphreys, S.M.; Clark, M.L.; Fielding, B.A.; Boland, O.; Coppack, S.W. Regulation of fatty acid movement in human adipose tissue in the postabsorptive-to-postprandial transition. Am. J. Physiol. Metab. 1994, 266, E308–E317. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Borén, J.; Dulloo, A.G. De novo lipogenesis in metabolic homeostasis: More friend than foe? Mol. Metab. 2015, 4, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fortuño-Mar, A.; Pasquali, P. Cryobiopsy, Cryoanesthesia, and Cryoanalgesia. In Cryosurgery: A Practical Manual; Pasquali, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 85–91. [Google Scholar]
- Eriksson-Hogling, D.; Andersson, D.P.; Bäckdahl, J.; Hoffstedt, J.; Rössner, S.; Thorell, A.; Arner, E.; Arner, P.; Rydén, M. Adipose tissue morphology predicts improved insulin sensitivity following moderate or pronounced weight loss. Int. J. Obes. 2015, 39, 893. [Google Scholar] [CrossRef] [PubMed]
- Lemonnier, D. Effect of age, sex, and sites on the cellularity of the adipose tissue in mice and rats rendered obese by a high-fat diet. J. Clin. Investig. 1972, 51, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Esquirol, Y.; Bongard, V.; Mabile, L.; Jonnier, B.; Soulat, J.M.; Perret, B. Shift work and metabolic syndrome: Respective impacts of job strain, physical activity, and dietary rhythms. Chronobiol. Int. 2009, 26, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Tavolaro, F.M.; Thomson, L.M.; Ross, A.W.; Morgan, P.J.; Helfer, G. Photoperiodic Effects on Seasonal Physiology, Reproductive Status and Hypothalamic Gene Expression in Young Male F344 Rats. J. Neuroendocrinol. 2015, 27, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Heideman, P.D. Inhibition of Reproductive Maturation and Somatic Growth of Fischer 344 Rats by Photoperiods Shorter than L14:D10 and by Gradually Decreasing Photoperiod. Biol. Reprod. 2000, 63, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Togo, Y.; Otsuka, T.; Goto, M.; Furuse, M.; Yasuo, S. Photoperiod regulates dietary preferences and energy metabolism in young developing Fischer 344 rats but not in same-age Wistar rats. Am. J. Physiol.-Endocrinol. Metab. 2012, 303, E777–E786. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.; Roemmich, J.N.; Claycombe, K.J. Evaluation of markers of beige adipocytes in white adipose tissue of the mouse. Nutr. Metab. 2016, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Mele, L.; Bidault, G.; Mena, P.; Crozier, A.; Brighenti, F.; Vidal-Puig, A.; Del Rio, D. Dietary (Poly)phenols, Brown Adipose Tissue Activation, and Energy Expenditure: A Narrative Review. Adv. Nutr. 2017, 8, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Bindels, L.B.; Geurts, L.; Van Hul, M.; Cani, P.D.; Delzenne, N.M. A polyphenolic extract from green tea leaves activates fat browning in high-fat-diet-induced obese mice. J. Nutr. Biochem. 2017, 49, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Arias, N.; Picó, C.; Teresa Macarulla, M.; Oliver, P.; Miranda, J.; Palou, A.; Portillo, M.P. A combination of resveratrol and quercetin induces browning in white adipose tissue of rats fed an obesogenic diet. Obesity 2017, 25, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M.; Rodgers, B.D.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) α1. Int. J. Obes. 2015. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Guo, J.; You, Y.; Yin, M.; Liang, J.; Ren, C.; Zhan, J.; Huang, W. Vanillic acid activates thermogenesis in brown and white adipose tissue. Food Funct. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mandard, S.; Müller, M.; Kersten, S. Peroxisome proliferator-activated receptor a target genes. Cell. Mol. Life Sci. 2004, 61, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Barberá, M.J.; Schlüter, A.; Pedraza, N.; Iglesias, R.; Villarroya, F.; Giralt, M. Peroxisome proliferator-activated receptor α activates transcription of the brown fat uncoupling protein-1 gene. A link between regulation of the thermogenic and lipid oxidation pathways in the brown fat cell. J. Biol. Chem. 2001, 276, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Contreras, A.V.; Torres, N.; Tovar, A.R. Ppar-α as a Key Nutritional and Environmental Sensor for Metabolic Adaptation. Adv. Nutr. 2013, 4, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Nie, B.; Nie, T.; Hui, X.; Gu, P.; Mao, L.; Li, K.; Yuan, R.; Zheng, J.; Wang, H.; Li, K.; et al. Brown Adipogenic Reprogramming Induced by a Small Molecule. Cell Rep. 2017, 18, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.; Houten, S.M.; Huber, A.; Schreiber, V.; Watanabe, M.; Kiss, B.; De Murcia, G.; Auwerx, J.; Ménissier-De Murcia, J. Peroxisome Proliferator-activated Receptor (Ppar)-2 controls adipocyte differentiation and adipose tissue function through the regulation of the activity of the retinoid X receptor/Pparγ heterodimer. J. Biol. Chem. 2007, 282, 37738–37746. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Mccune, L.M.; Kubota, C.; Stendell-hollis, N.R.; Thomson, C.A. Cherries and Health: A Review. Crit. Rev. Food Sci. Nutr. 2010, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.S.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Heyne, A.; Kiesselbach, C.; Sahún, I.; McDonald, J.; Gaiffi, M.; Dierssen, M.; Wolffgramm, J. An animal model of compulsive food-taking behaviour. Addict. Biol. 2009, 14, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Silha, J.V.; Krsek, M.; Skrha, J.V.; Sucharda, P.; Nyomba, B.L.G.; Murphy, L.J. Plasma resistin, adiponectin and leptin levels in lean and obese subjects: Correlations with insulin resistence. Eur. J. Endocrinol. 2003, 149, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Chien, K.L.; Lee, B.C.; Lin, H.J.; Hsu, H.C.; Chen, M.F. Association of fasting and post-prandial hyperglycemia on the risk of cardiovascular and all-cause death among non-diabetic Chinese. Diabetes Res. Clin. Pract. 2009, 83, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Weyer, C.; Foley, J.E.; Bogardus, C.; Tataranni, P.A.; Pratley, R.E. Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts type II diabetes independent of insulin resistance. Diabetologia 2000, 43, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Jernås, M.; Palming, J.; Sjöholm, K.; Jennische, E.; Svensson, P.A.; Gabrielsson, B.G.; Levin, M.; Sjögren, A.; Rudemo, M.; Lystig, T.C.; et al. Separation of human adipocytes by size: Hypertrophic fat cells display distinct gene expression. FASEB J. 2006, 20, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Auguet, T.; Guiu-Jurado, E.; Berlanga, A.; Terra, X.; Martinez, S.; Porras, J.A.; Ceausu, A.; Sabench, F.; Hernandez, M.; Aguilar, C.; et al. Downregulation of lipogenesis and fatty acid oxidation in the subcutaneous adipose tissue of morbidly obese women. Obesity 2014, 22, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Vela, M.E.F.; Torres, N.; Tovar, A.R. White Adipose Tissue as Endocrine Organ and Its Role in Obesity. Arch. Med. Res. 2008, 39, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Hodson, L.; Dennis, A.L.; Neville, M.J.; Humphreys, S.M.; Harnden, K.E.; Micklem, K.J.; Frayn, K.N. Markers of de novo lipogenesis in adipose tissue: Associations with small adipocytes and insulin sensitivity in humans. Diabetologia 2009, 52, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M. Adipose tissue and adipocyte dysregulation. Diabetes Metab. 2014, 40, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Terada, T.; Okuda, M.; Inui, K. The Diurnal Rhythm of the Intestinal Transporters SGLT1 and PEPT1 Is Regulated by the Feeding Conditions in Rats 1. J. Nutr. 2004, 134, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Gómez-Abellán, P. Timing of food intake and obesity: A novel association. Physiol. Behav. 2014, 134, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Hirao, A.; Tahara, Y.; Kimura, I.; Shibata, S. A Balanced Diet Is Necessary for Proper Entrainment Signals of the Mouse Liver Clock. PLoS ONE 2009, 4, 4–13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VHLD | CHLD | VHSD | CHSD | |
|---|---|---|---|---|
| Weight (g) | 386.5 ± 12.66 | 388 ± 5.11 | 370.33 ± 10.99 | 376.5 ± 13.58 |
| Accumulated caloric intake (kcal) | 504.79 ± 11.67 | 493.3 ± 10.78 | 507.07 ± 9.44 | 498.44 ± 13.38 |
| Fat (g) | 55.64 ± 4.41 a | 58.38 ± 3.11 a | 45.06 ± 1.29 b | 50.41 ± 2.82 ab |
| Lean (g) | 309.74 ± 8.96 | 307.43 ± 4.59 | 295.76 ± 8.19 | 299.16 ± 11.89 |
| Fat (%) | 14.38 ± 0.75 ab | 15.17 ± 0.7 a | 12.52 ± 0.34 b | 13.43 ± 0.57 ab |
| Lean (%) | 80.71 ± 0.67 | 79.95 ± 0.76 | 80.94 ± 1.04 | 79.69 ± 0.62 |
| EWAT (g) | 12.02 ± 0.94 ab | 12.99 ± 0.7 a | 9.65 ± 0.67 b | 9.64 ± 0.71 b |
| MWAT (g) | 8.04 ± 0.85 | 7.95 ± 0.45 | 6.53 ± 0.82 | 6.42 ± 0.65 |
| IWAT (g) | 5.82 ± 0.73 | 6.27 ± 0.3 | 4.52 ± 0.53 | 5.66 ± 1.03 |
| RWAT (g) | 10 ± 0.67 ab | 10.33 ± 0.29 a | 8.52 ± 0.58 b | 8.71 ± 0.47 ab |
| Adiposity Index (%) | 9.22 ± 0.49 ab | 9.67 ± 0.34 a | 7.72 ± 0.47 b | 8.05 ± 0.55 b |
| EWAT (%) | 3.09 ± 0.17 a | 3.34 ± 0.16 a | 2.6 ± 0.14 b | 2.56 ± 0.16 b |
| MWAT (%) | 2.06 ± 0.17 | 2.05 ± 0.11 | 1.74 ± 0.16 | 1.69 ± 0.13 |
| IWAT (%) | 1.5 ± 0.15 | 1.62 ± 0.08 | 1.25 ± 0.14 | 1.49 ± 0.25 |
| RWAT (%) | 2.58 ± 0.11 ab | 2.66 ± 0.07 a | 2.41 ± 0.05 ab | 2.31 ± 0.09 b |
| Glucose (mmol/L) | 136.81 ± 3.48 ab | 133.61 ± 3.49 b | 139.24 ± 3.51 ab | 145.5 ± 2.72 a |
| Triglycerides (mg/dL) | 142.06 ± 7.74 bc | 120.03 ± 10.22 c | 197.9 ± 14.45 a | 181.22 ± 23.03 ab |
| Insulin (ng/mL) | 5.54 ± 0.73 | 6.15 ± 0.96 | 4.04 ± 0.66 | 4.03 ± 0.79 |
| Leptin (ng/mL) | 18.56 ± 0.31 ab | 22.69 ± 2.19 a | 16.59 ± 1.49 b | 16.21 ± 2.08 b |
| VHLD | CHLD | VHSD | CHSD | |
|---|---|---|---|---|
| Weight (g) | 411 ± 7.77 | 422.89 ± 7.14 | 407.1 ± 12.03 | 404.3 ± 9.61 |
| Accumulated caloric intake (kcal) | 1364.34 ± 37.39 a | 1214.67 ± 58.02 b | 1298.52 ± 48.45 ab | 1198.2 ± 30.34 b |
| Fat (g) | 89.51 ± 3.8 | 86.56 ± 2.23 | 85.84 ± 3.27 | 88.29 ± 5.31 |
| Lean (g) | 291.95 ± 5.14 | 299.07 ± 5.23 | 294.5 ± 8.32 | 296.42 ± 3.82 |
| Fat (%) | 22.03 ± 0.63 | 21.7 ± 0.66 | 21.53 ± 0.79 | 21.82 ± 1.04 |
| Lean (%) | 72.07 ± 0.54 | 72.5 ± 0.62 | 73.68 ± 0.76 | 72.33 ± 1.07 |
| EWAT (g) | 16.23 ± 0.82 | 15.75 ± 0.53 | 14.75 ± 0.72 | 15.61 ± 1.08 |
| MWAT (g) | 9.42 ± 0.61 | 9.25 ± 0.66 | 8.68 ± 0.47 | 8.28 ± 0.69 |
| IWAT (g) | 11.9 ± 1.39 | 11 ± 1.02 | 12.47 ± 1.39 | 10.39 ± 1.25 |
| RWAT (g) | 13.53 ± 0.59 | 13.41 ± 0.26 | 12.87 ± 0.54 | 12.41 ± 0.59 |
| Adiposity Index (%) | 12.23 ± 0.54 | 11.51 ± 0.3 | 11.95 ± 0.42 | 11.5 ± 0.58 |
| EWAT (%) | 3.93 ± 0.13 | 3.76 ± 0.09 | 3.62 ± 0.14 | 3.84 ± 0.21 |
| MWAT (%) | 2.28 ± 0.11 | 2.18 ± 0.13 | 2.15 ± 0.14 | 2.03 ± 0.14 |
| IWAT (%) | 2.87 ± 0.31 | 2.59 ± 0.21 | 3.02 ± 0.27 | 2.57 ± 0.31 |
| RWAT (%) | 3.28 ± 0.1 | 3.18 ± 0.06 | 3.16 ± 0.08 | 3.14 ± 0.06 |
| Glucose (mmol/L) | 182.87 ± 4.65 ab | 180.5 ± 9.23 ab | 164.13 ± 4.91 b | 202.31 ± 15.46 a |
| Triglycerides (mg/dL) | 437.02 ± 30.47 | 421.07 ± 12.89 | 444.19 ± 42.34 | 449.75 ± 48.06 |
| Insulin (ng/mL) | 6.59 ± 0.47 ab | 7.42 ± 0.49 a | 5.82 ± 0.24 b | 7.31 ± 0.59 a |
| Leptin (ng/mL) | 19.89 ± 0.55 | 20.08 ± 1.08 | 21.41 ± 1.33 | 20.9 ± 0.96 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibert-Ramos, A.; Crescenti, A.; Salvadó, M.J. Consumption of Cherry out of Season Changes White Adipose Tissue Gene Expression and Morphology to a Phenotype Prone to Fat Accumulation. Nutrients 2018, 10, 1102. https://doi.org/10.3390/nu10081102
Gibert-Ramos A, Crescenti A, Salvadó MJ. Consumption of Cherry out of Season Changes White Adipose Tissue Gene Expression and Morphology to a Phenotype Prone to Fat Accumulation. Nutrients. 2018; 10(8):1102. https://doi.org/10.3390/nu10081102
Chicago/Turabian StyleGibert-Ramos, Albert, Anna Crescenti, and M. Josepa Salvadó. 2018. "Consumption of Cherry out of Season Changes White Adipose Tissue Gene Expression and Morphology to a Phenotype Prone to Fat Accumulation" Nutrients 10, no. 8: 1102. https://doi.org/10.3390/nu10081102
APA StyleGibert-Ramos, A., Crescenti, A., & Salvadó, M. J. (2018). Consumption of Cherry out of Season Changes White Adipose Tissue Gene Expression and Morphology to a Phenotype Prone to Fat Accumulation. Nutrients, 10(8), 1102. https://doi.org/10.3390/nu10081102