Combination of Aronia, Red Ginseng, Shiitake Mushroom and Nattokinase Potentiated Insulin Secretion and Reduced Insulin Resistance with Improving Gut Microbiome Dysbiosis in Insulin Deficient Type 2 Diabetic Rats




Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of the Product and Analysis of Ingredients
2.2. Animals and Ethics
2.3. Experimental Design
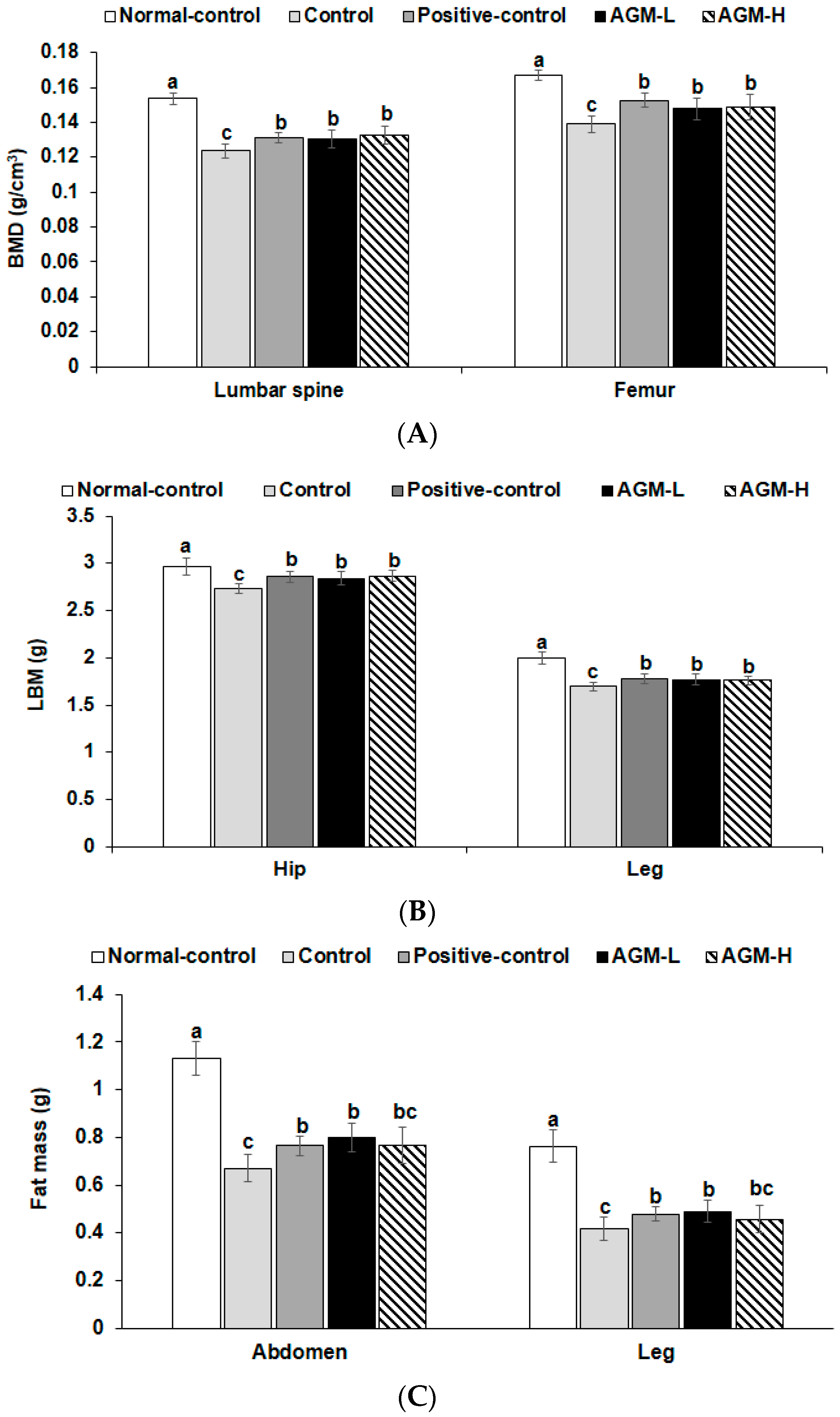
2.4. Body Composition Measurement
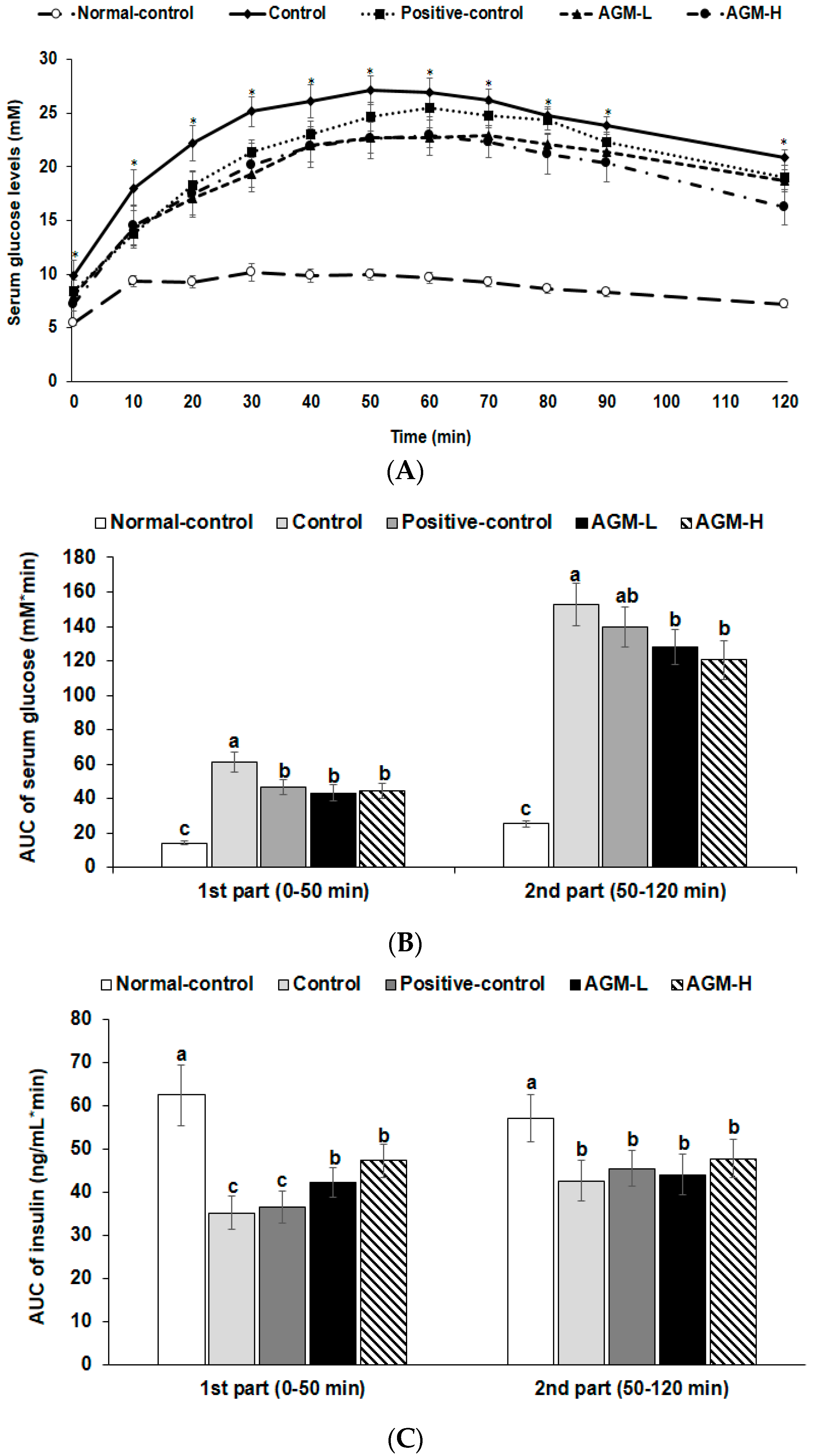
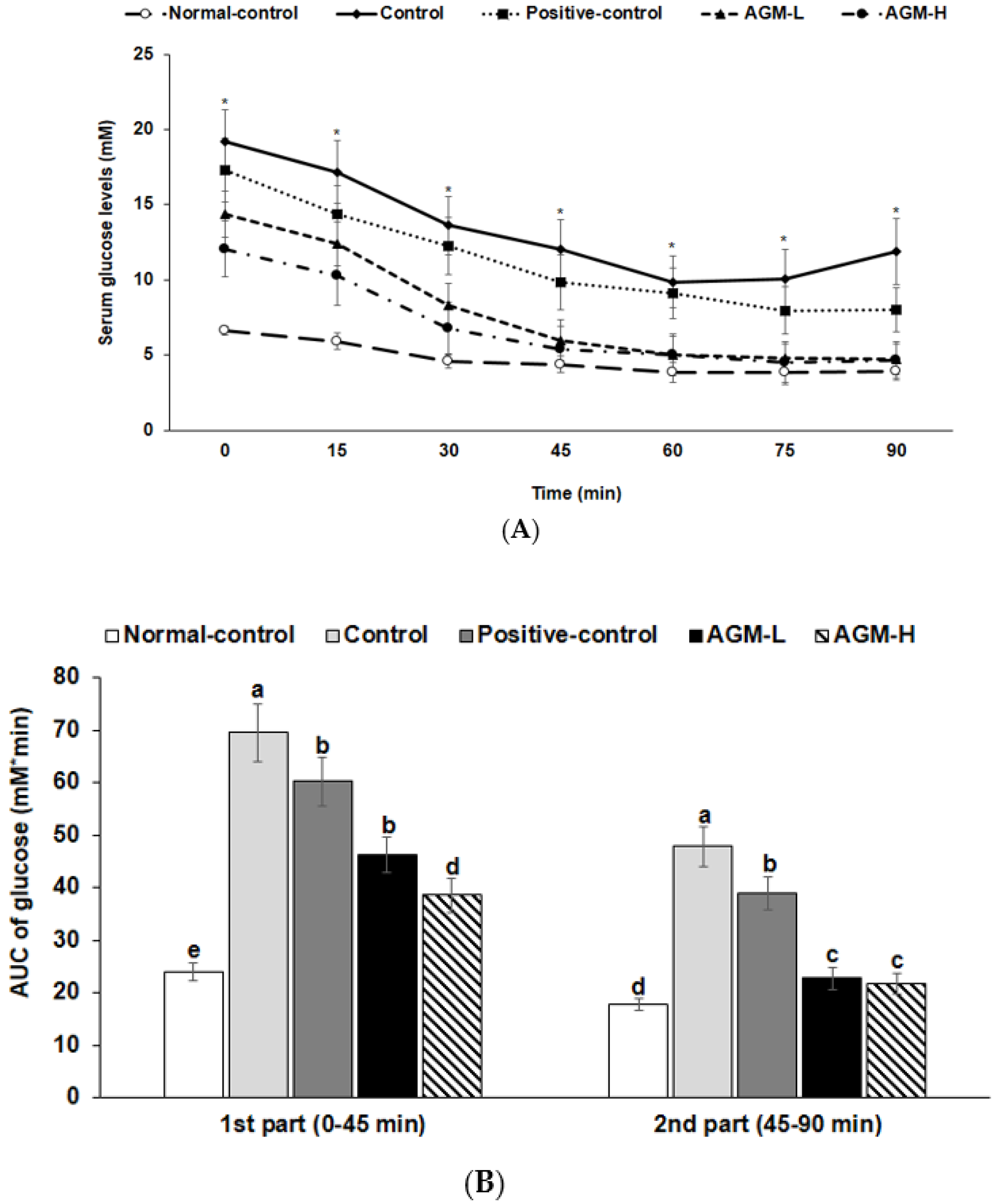
2.5. Glucose Homeostasis
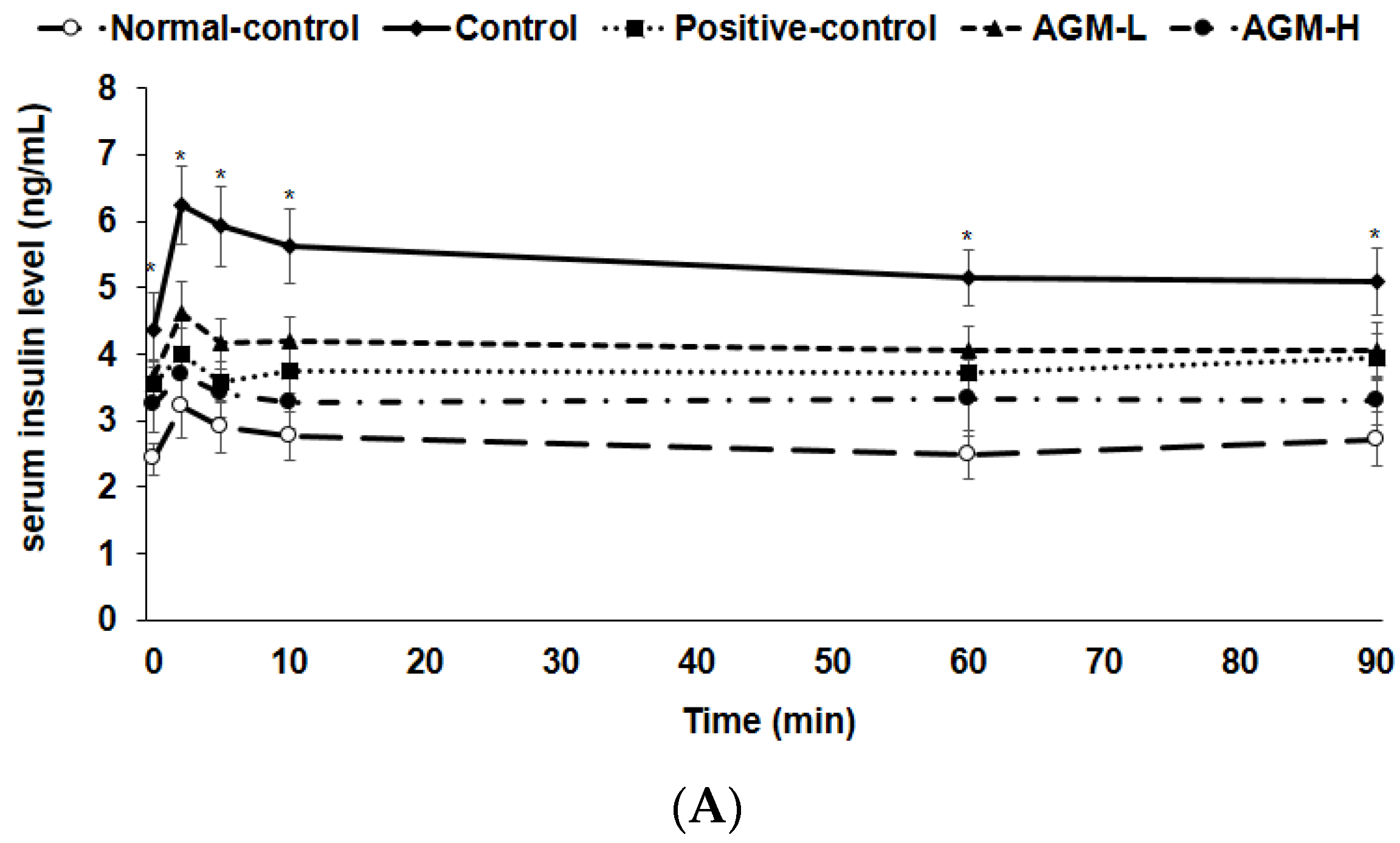
2.6. Hyperglycemic Clamp and Cerebral Blood Flow
2.7. Immunohistochemistry
2.8. Next Generation Sequencing of the Gut Microbiome
2.9. Statistical Analyses
3. Results
3.1. Contents of Anthocyanins and Ginsenoide Rg3
3.2. Body Composition
3.3. Glucose Metabolism
3.4. Insulin Tolerance
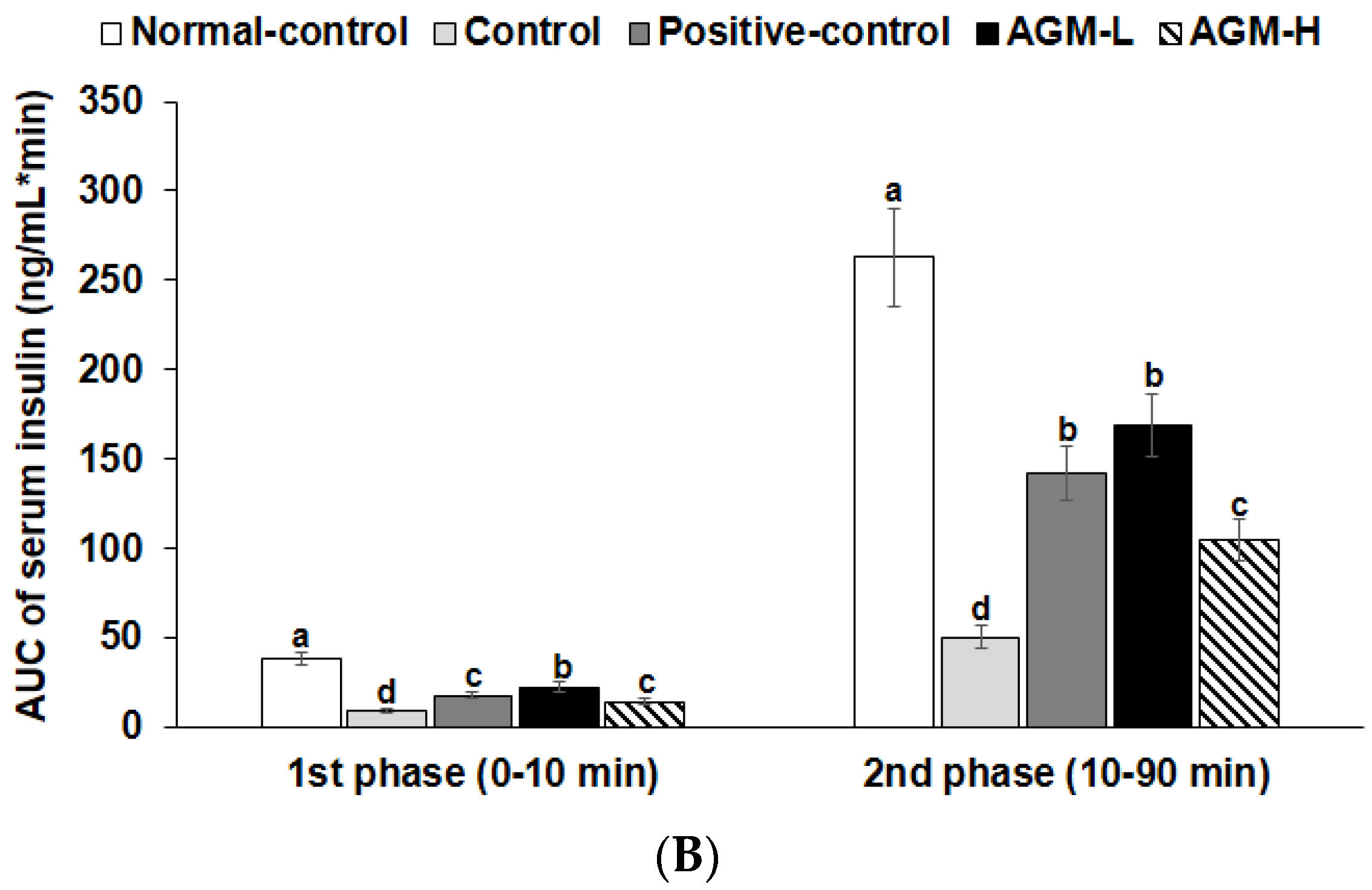
3.5. Hyperglycemic Clamp
3.6. Pancreatic β-cell Mass, Proliferation, and Apoptosis
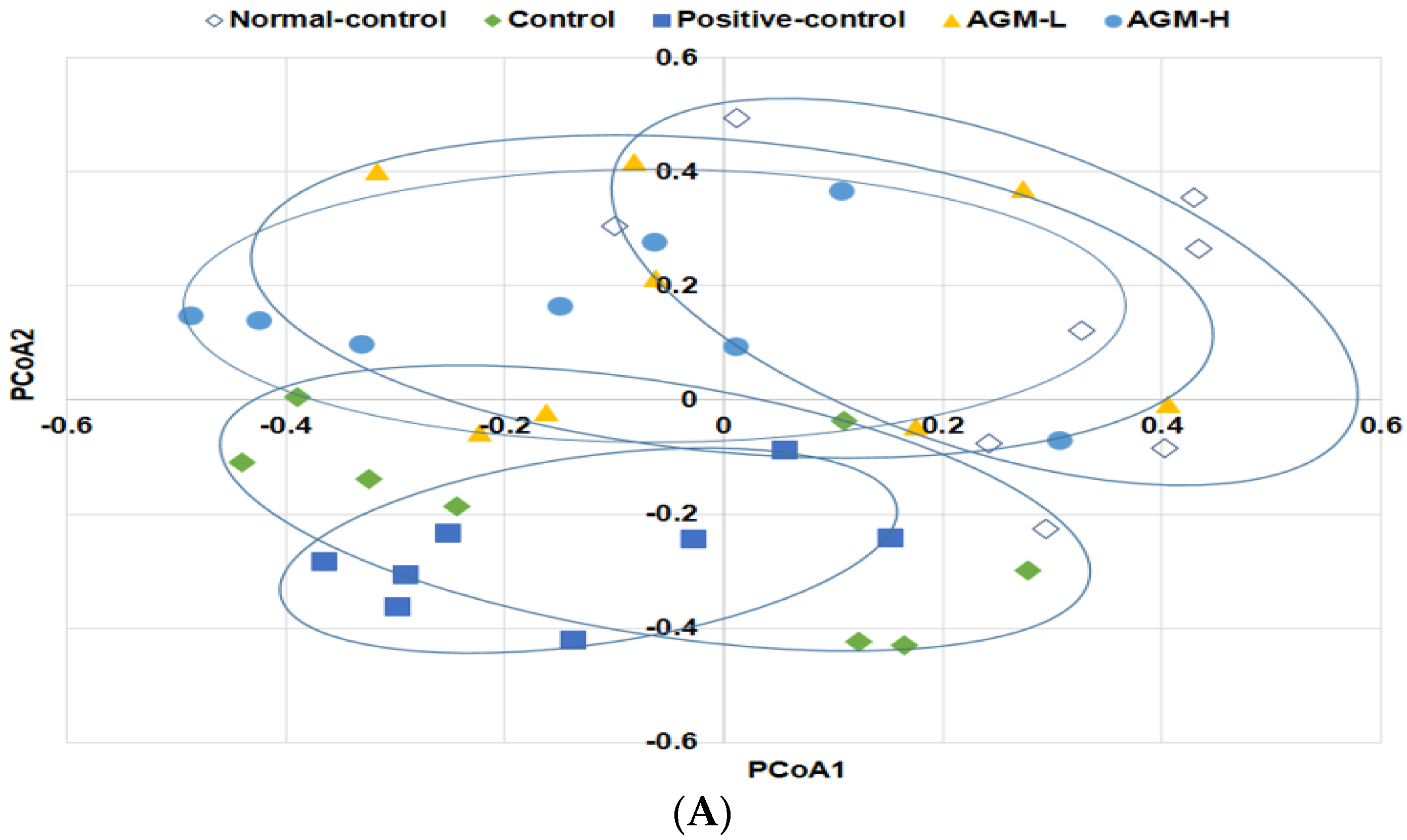
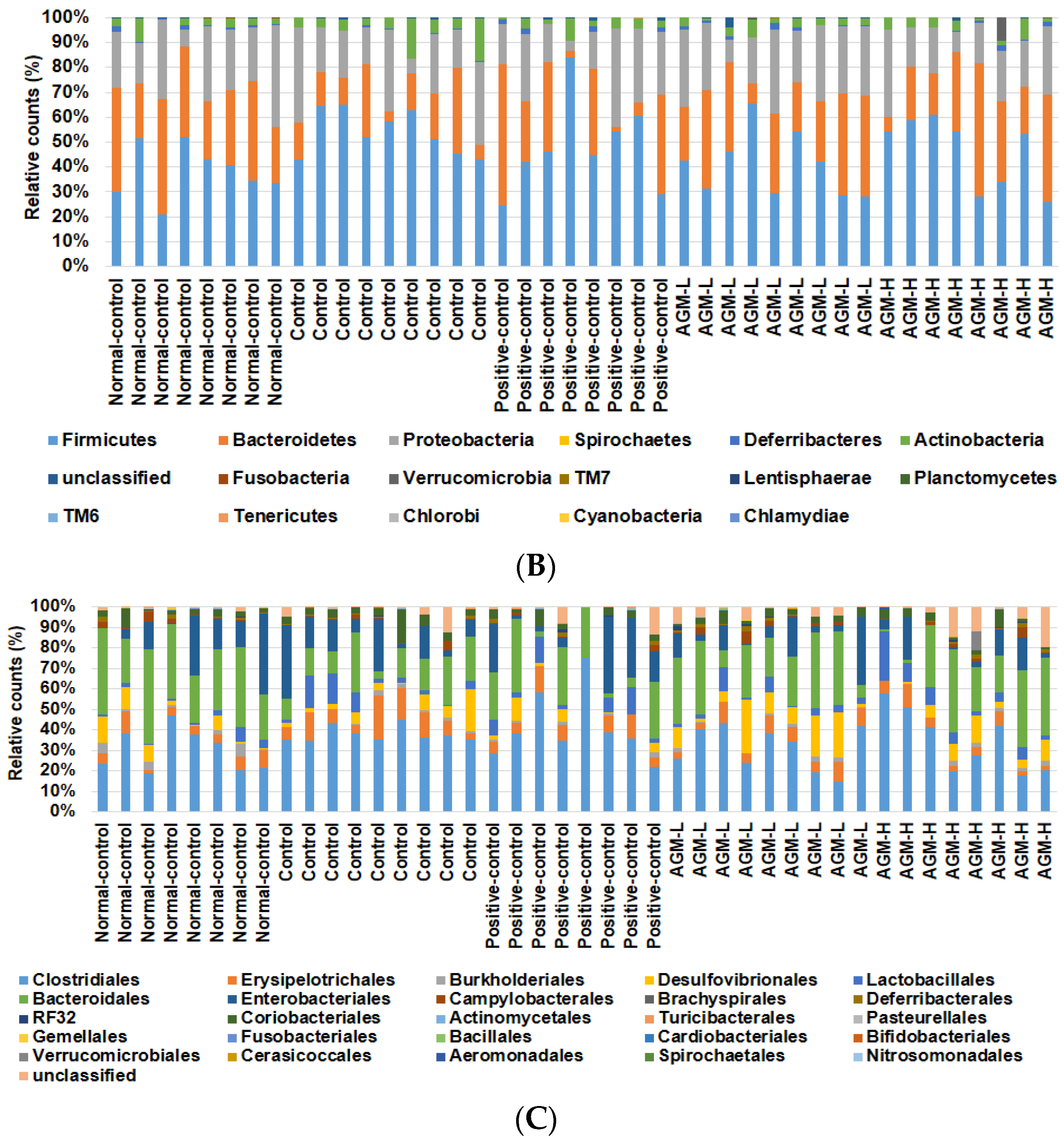
3.7. Gut Microbiome
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yuan, H.; Li, X.; Wan, G.; Sun, L.; Zhu, X.; Che, F.; Yang, Z. Type 2 diabetes epidemic in East Asia: A 35-year systematic trend analysis. Oncotarget 2018, 9, 6718–6727. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, B.C.; Daily, J.W.; Park, S. High genetic risk scores for impaired insulin secretory capacity doubles the risk for type 2 diabetes in asians and is exacerbated by western-type diets. Diabetes Metab. Res. Rev. 2018, 34. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Kim, M.J.; Kwon, D.Y.; Kim, D.S.; Lee, Y.H.; Kim, J.E.; Park, S. Anti-diabetic activities of Gastrodia elata blume water extracts are mediated mainly by potentiating glucose-stimulated insulin secretion and increasing beta-cell mass in non-obese type 2 diabetic animals. Nutrients 2016, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kang, S. Vitamin D deficiency impairs glucose-stimulated insulin secretion and increases insulin resistance by reducing PPAR-gamma expression in nonobese type 2 diabetic rats. J. Nutr. Biochem. 2016, 27, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kang, S.; Moon, B.R. Fermented soybeans, Chungkookjang, prevent hippocampal cell death and beta-cell apoptosis by decreasing pro-inflammatory cytokines in gerbils with transient artery occlusion. Exp. Biol. Med. 2016, 241, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Wang, Y.F.; Fan, G.W.; Wang, X.Y.; Xu, S.Y.; Zhu, Y. Balancing herbal medicine and functional food for prevention and treatment of cardiometabolic diseases through modulating gut microbiota. Front. Microbiol. 2017, 8, 2146. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Duffy, A. Factors influencing the gut microbiota, inflammation, and type 2 diabetes. J. Nutr. 2017, 147, 1468s–1475s. [Google Scholar] [CrossRef] [PubMed]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ahn, I.S.; Kwon, D.Y.; Ko, B.S.; Jun, W.K. Ginsenosides Rb1 and Rg1 suppress triglyceride accumulation in 3T3-L1 adipocytes and enhance beta-cell insulin secretion and viability in min6 cells via pka-dependent pathways. Biosci. Biotechnol. Biochem. 2008, 72, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, Y.H.; Park, S.K.; Kang, E.S.; Kim, H.J.; Lee, Y.C.; Choi, C.S.; Park, S.E.; Ahn, C.W.; Cha, B.S.; et al. Korean red ginseng (Panax ginseng) improves insulin sensitivity and attenuates the development of diabetes in Otsuka Long-Evans Tokushima fatty rats. Metabolism 2009, 58, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Oh, M.R.; Choi, E.K.; Kim, M.G.; Ha, K.C.; Lee, S.K.; Kim, Y.G.; Park, B.H.; Kim, D.S.; Chae, S.W. An 8-wk, randomized, double-blind, placebo-controlled clinical trial for the antidiabetic effects of hydrolyzed ginseng extract. J. Ginseng Res. 2014, 38, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, K. Regulation of signaling molecules associated with insulin action, insulin secretion and pancreatic beta-cell mass in the hypoglycemic effects of Korean red ginseng in Goto-Kakizaki rats. J. Ethnopharmacol. 2012, 142, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Grotto, D.; Bueno, D.C.; Ramos, G.K.; da Costa, S.R.; Spim, S.R.; Gerenutti, M. Assessment of the safety of the shiitake culinary-medicinal mushroom, lentinus edodes (agaricomycetes), in rats: Biochemical, hematological, and antioxidative parameters. Int. J. Med. Mushrooms 2016, 18, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Alvarsson, M.; Mannheimer, B.; Degerblad, M.; Östenson, C.G. No Effect of high-dose vitamin D treatment on β-cell function, insulin sensitivity, or glucose homeostasis in subjects with abnormal glucose tolerance: A randomized clinical trial. Diabetes Care 2016, 39, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Westein, E.; Hoefer, T.; Calkin, A.C. Thrombosis in diabetes: A shear flow effect? Clin. Sci. 2017, 131, 1245–1260. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Hong, K.; Ito, Y.; Fujii, R.; Kariya, K.; Nishimuro, S. Thrombolytic effect of nattokinase on a chemically induced thrombosis model in rat. Biol. Pharm. Bull. 1995, 18, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.H.; Gheewala, N.M.; O’Keefe, J.O. Dietary strategies for improving post-prandial glucose, lipids, inflammation, and cardiovascular health. J. Am. Coll. Cardiol. 2008, 51, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.A.; Hosokawa, H.; Chen, C.; Leahy, J.L. Mechanism of impaired glucose-potentiated insulin secretion in diabetic 90% pancreatectomy rats. Study using glucagonlike peptide-1 (7-37). J. Clin. Investig. 1996, 97, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Wilson, R.D. Experimentally induced rodent models of type 2 diabetes. Methods Mol. Biol. 2012, 933, 161–174. [Google Scholar] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. Ain-93 purified diets for laboratory rodents: Final report of the american institute of nutrition ad hoc writing committee on the reformulation of the ain-76a rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; da Kim, S.; Kang, S. Gastrodia elata blume water extracts improve insulin resistance by decreasing body fat in diet-induced obese rats: Vanillin and 4-hydroxybenzaldehyde are the bioactive candidates. Eur. J. Nutr. 2011, 50, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.B.; Jang, J.S.; Park, S. Estrogen and exercise may enhance beta-cell function and mass via insulin receptor substrate 2 induction in ovariectomized diabetic rats. Endocrinology 2005, 146, 4786–4794. [Google Scholar] [CrossRef] [PubMed]
- Dobbins, R.L.; Szczepaniak, L.S.; Myhill, J.; Tamura, Y.; Uchino, H.; Giacca, A.; McGarry, J.D. The composition of dietary fat directly influences glucose-stimulated insulin secretion in rats. Diabetes 2002, 51, 1825–1833. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kang, E.S.; Kim, D.B.; Kang, S. Low dose brain estrogen prevents menopausal syndrome while maintaining the diversity of the gut microbiomes in estrogen-deficient rats. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E99–E109. [Google Scholar] [CrossRef] [PubMed]
- Kamweru, P.K.; Tindibale, E.L. Vitamin d and vitamin d from ultraviolet-irradiated mushrooms (review). Int. J. Med. Mushrooms 2016, 18, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Reeds, D.N.; Patterson, B.W.; Okunade, A.; Holloszy, J.O.; Polonsky, K.S.; Klein, S. Ginseng and ginsenoside re do not improve beta-cell function or insulin sensitivity in overweight and obese subjects with impaired glucose tolerance or diabetes. Diabetes Care 2011, 34, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Banjari, I.; Misir, A.; Savikin, K.; Jokic, S.; Molnar, M.; De Zoysa, H.K.S.; Waisundara, V.Y. Antidiabetic effects of Aronia melanocarpa and its other therapeutic properties. Front. Nutr. 2017, 4, 53. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Jialal, I. Hyperglycemia induces toll-like receptor activity through increased oxidative stress. Metab. Syndr. Relat. Disord. 2016, 14, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Stitham, J.; Gleim, S.; Di Febbo, C.; Porreca, E.; Fava, C.; Tacconelli, S.; Capone, M.; Evangelista, V.; Levantesi, G.; et al. Glucose and collagen regulate human platelet activity through aldose reductase induction of thromboxane. J. Clin. Investig. 2011, 121, 4462–4476. [Google Scholar] [CrossRef] [PubMed]
- Rugina, D.; Diaconeasa, Z.; Coman, C.; Bunea, A.; Socaciu, C.; Pintea, A. Chokeberry anthocyanin extract as pancreatic beta-cell protectors in two models of induced oxidative stress. Oxid. Med. Cell. Longev. 2015, 2015, 429075. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyanidin-3-O-beta-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Kozuka, M.; Konda, D.; Nakano, Y.; Nakagaki, T.; Ohkubo, I.; Ariga, H. Improvement of blood glucose levels and obesity in mice given aronia juice by inhibition of dipeptidyl peptidase iv and alpha-glucosidase. J. Nutr. Biochem. 2016, 31, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Simeonov, S.B.; Botushanov, N.P.; Karahanian, E.B.; Pavlova, M.B.; Husianitis, H.K.; Troev, D.M. Effects of aronia melanocarpa juice as part of the dietary regimen in patients with diabetes mellitus. Folia Med. 2002, 44, 20–23. [Google Scholar]
- Shishtar, E.; Sievenpiper, J.L.; Djedovic, V.; Cozma, A.I.; Ha, V.; Jayalath, V.H.; Jenkins, D.J.; Meija, S.B.; de Souza, R.J.; Jovanovski, E.; et al. The effect of ginseng (the Genus panax) on glycemic control: A systematic review and meta-analysis of randomized controlled clinical trials. PLoS ONE 2014, 9, e107391. [Google Scholar] [CrossRef] [PubMed]
- Maschio, B.H.; Gentil, B.C.; Caetano, E.L.A.; Rodrigues, L.S.; Laurino, L.F.; Spim, S.R.V.; Jozala, A.F.; Dos Santos, C.A.; Grotto, D.; Gerenutti, M. Characterization of the effects of the shiitake culinary-medicinal mushroom, lentinus edodes (agaricomycetes), on severe gestational diabetes mellitus in rats. Int. J. Med. Mushrooms 2017, 19, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.Y.; Kang, S.; Hua, C.S.; Ting, Z.; Park, S. Synbiotic effects of beta-glucans from cauliflower mushroom and Lactobacillus fermentum on metabolic changes and gut microbiome in estrogen-deficient rats. Genes Nutr. 2017, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.K.; Park, S.; Kim, Y. Age- and gender-specific associations between low serum 25-hydroxyvitamin d level and type 2 diabetes in the Korean general population: Analysis of 2008–2009 Korean national health and nutrition examination survey data. Asia Pac. J. Clin. Nutr. 2012, 21, 536–546. [Google Scholar] [PubMed]
- Park, S.; Ham, J.O.; Lee, B.K. A positive association of vitamin d deficiency and sarcopenia in 50 year old women, but not men. Clin. Nutr. 2014, 33, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Lee, O.H.; Kim, K.J.; Lee, B.Y. Ginsenoside Rg1 promotes glucose uptake through activated AMPK pathway in insulin-resistant muscle cells. Phytother. Res. 2012, 26, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.S.; Yoon, M. Korean red ginseng (Panax ginseng) inhibits obesity and improves lipid metabolism in high fat diet-fed castrated mice. J. Ethnopharmacol. 2018, 210, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Sikora, J.; Broncel, M.; Markowicz, M.; Chalubinski, M.; Wojdan, K.; Mikiciuk-Olasik, E. Short-term supplementation with Aronia melanocarpa extract improves platelet aggregation, clotting, and fibrinolysis in patients with metabolic syndrome. Eur. J. Nutr. 2012, 51, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Lian, S.; Qian, X.; Wang, N.; Huang, H.; Yao, J.; Tang, K.; Chen, L.; Li, L.; Lin, W.; et al. Association between bone mineral density and pancreatic beta-cell function in elderly men and postmenopausal women. J. Endocr. Soc. 2017, 1, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Buranasinsup, S.; Bunyaratavej, N. The Intriguing Correlation between Undercarboxylated Osteocalcin and Vitamin D. J. Med. Assoc. Thail. 2015, 98, S16–S20. [Google Scholar]
- Sabek, O.M.; Nishimoto, S.K.; Fraga, D.; Tejpal, N.; Ricordi, C.; Gaber, A.O. Osteocalcin Effect on Human β-Cells Mass and Function. Endocrinology 2015, 156, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Kover, K.; Yan, Y.; Tong, P.Y.; Watkins, D.; Li, X.; Tasch, J.; Hager, M.; Clements, M.; Moore, W.V. Osteocalcin protects pancreatic beta cell function and survival under high glucose conditions. Biochem. Biophys. Res. Commun. 2015, 462, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Gower, B.A.; Pollock, N.K.; Casazza, K.; Clemens, T.L.; Goree, L.L.; Granger, W.M. Associations of total and undercarboxylated osteocalcin with peripheral and hepatic insulin sensitivity and β-cell function in overweight adults. J. Clin. Endocrinol. Metab. 2013, 98, E1173–E1180. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Yu, J.; Choi, H.; An, J.H.; Kim, S.W.; Park, K.S.; Jang, H.C.; Kim, S.Y.; Shin, C.S. Vitamin K2 supplementation improves insulin sensitivity via osteocalcin metabolism: A placebo-controlled trial. Diabetes Care 2011, 34, e147. [Google Scholar] [CrossRef] [PubMed]
- Lecka-Czernik, B. Diabetes, bone and glucose-lowering agents: Basic biology. Diabetologia 2017, 60, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, M.F.; Su, Y.P.; Jiang, H.M.; You, X.J.; Yang, Y.J.; Zhang, H.L. Ginsenoside Re reduces insulin resistance through activation of PPAR-gamma pathway and inhibition of TNF-alpha production. J. Ethnopharmacol. 2013, 147, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Mollah, M.L.; Kim, G.S.; Moon, H.K.; Chung, S.K.; Cheon, Y.P.; Kim, J.K.; Kim, K.S. Antiobesity effects of wild ginseng (Panax ginseng C.A. Meyer) mediated by PPAR-gamma, GLUT4 and LPL in ob/ob mice. Phytother. Res. 2009, 23, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Pedersen, H.K.; et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Hiel, S.; Delzenne, N.M. Metformin: Old friend, new ways of action-implication of the gut microbiome? Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 294–301. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contents (mg/g Powder) | |
|---|---|
| C3-Galactoside | 6.22 |
| C3-Glucoside | 0.33 |
| C3-Arabinoside | 1.53 |
| Total Anthocyanin | 8.08 |
| Ginsenoside Rg3 | 2.5 |
| β-glucan | 244 |
| Normal-Control (n = 10) | Control (n = 10) | Positive-Control (n = 10) | AGM-L (n = 10) | AGM-H (n = 10) | |
|---|---|---|---|---|---|
| Body weight gain for 10 week (g) | 283 ± 10.5 a | 144 ± 10.7 b | 169 ± 11.8 b | 156 ± 19.1 b | 155 ± 13.0 b |
| Food intake (g/day) | 14.4 ± 1.0 a | 15.1 ± 0.8 a | 15.0 ± 1.0 a | 13.4 ± 1.5 b | 12.3 ± 1.5 b |
| Food efficiency | 0.30 ± 0.01 a | 0.15 ± 0.01 d | 0.18 ± 0.12 c | 0.19 ± 0.03 c | 0.22 ± 0.02 b |
| Epididymal fat pads (g) | 6.7 ± 0.7 a | 3.0 ± 0.4 c | 2.9 ± 0.3 c | 3.8 ± 0.5 b | 3.3 ± 0.4 c |
| Retroperitoneal fat mass (g) | 8.1 ± 0.8 a | 3.6 ± 0.5 c | 3.8 ± 0.5 c | 4.3 ± 0.6 b,c | 5.0 ± 0.7 b |
| Visceral fat (g) | 14.8 ± 1.6 a | 6.7 ± 0.9 c | 6.7 ± 0.8 c | 8.1 ± 1.0 b | 8.3 ± 1.0 b |
| Normal-Control (n = 10) | Control (n = 10) | Positive-Control (n = 10) | AGM-L (n = 10) | AGM-H (n = 10) | |
|---|---|---|---|---|---|
| Serum glucose at fasting state (mM) | 5.4 ± 0.5 d | 9.8 ± 0.6 a | 8.4 ± 0.6 b | 8.1 ± 0.7 b | 7.2 ± 0.5 c |
| Serum insulin at fasting state (ng/mL) | 3.78 ± 0.36 a | 2.83 ± 0.32 c | 3.55 ± 0.35 a,b | 3.27 ± 0.34 b | 3.23 ± 0.36 b |
| HOMA-IR | 5.4 ± 0.6 d | 9.8 ± 1.0 a | 8.4 ± 0.9 b | 8.1 ± 0.8 b | 7.2 ± 0.8 c |
| Urinary glucose | - | ++++ | +++ | ++ | + |
| Mean cerebral blood flow (mm/s) | 657 ± 45 a | 405 ± 42 c | 424 ± 45 c | 643 ± 65 a | 571 ± 67 b |
| Normal-Control (n = 10) | Control (n = 10) | Positive-Control (n = 10) | AGM-L (n = 10) | AGM-H (n = 10) | |
|---|---|---|---|---|---|
| Serum glucose levels at 60 min (mM) | 9.8 ± 0.9 c | 17.7 ± 1.4 a | 14.6 ± 1.7 b | 15.1 ± 1.6 b | 15.1 ± 1.1 b |
| Serum glucose levels at 90 min (mM) | 9.9 ± 0.9 c | 18.3 ± 1.7 a | 15.6 ± 1.7 b | 15.2 ± 1.3 b | 14.6 ± 1.2 b |
| Serum insulin levels at 2 min (ng/mL) | 6.2 ± 0.6 a | 3.2 ± 0.5 d | 4.0 ± 0.4 c | 4.6 ± 0.5 b | 3.7 ± 0.4 c |
| Serum insulin levels at 60 min (ng/mL) | 5.1 ± 0.4 a | 2.5 ± 0.4 d | 3.7 ± 0.3 b,c | 4.1 ± 0.5 b | 3.3 ± 0.6 c |
| Glucose infusion rates (umol/kg bw/min) | 59.4 ± 3.9 a | 26.1 ± 2.8 d | 35.6 ± 3.9 c | 32.8 ± 4.4 c | 43.9 ± 3.9 b |
| Insulin sensitivity at hyperglycemic state (µmol glucose min−1 100 g−1 per µmol insulin/L) | 33.3 ± 3.9 a | 19.8 ± 2.4 d | 25.8 ± 2.9 c | 23.4 ± 2.6 c | 28.9 ± 3.3 b |
| Normal-Control (n = 5) | Control (n = 5) | Positive-Control (n = 5) | AGM-L (n = 5) | AGM-H (n = 5) | |
|---|---|---|---|---|---|
| β-cell area (%) | 5.5 ± 0.7 c | 6.3 ± 0.8 b | 6.8 ± 0.8 a,b | 6.9 ± 0.8 a,b | 7.6 ± 0.9 a |
| Individual β-cell size (μm2) | 185 ± 23 c | 239 ± 26 a | 209 ± 23 b | 206 ± 25 b | 189 ± 22 b,c |
| Absolute β-cell mass (mg) | 33.4 ± 2.9 a | 17.9 ± 1.9 d | 22.4 ± 2.6 c | 23.7 ± 2.9 c | 28.8 ± 3.5 b |
| BrdU+ cells (% BrdU+ cells of islets) | 0.72 ± 0.09 c | 0.84 ± 0.09 b | 0.89 ± 0.11 b | 0.90 ± 0.12 b | 1.04 ± 0.12 a |
| Apoptosis (% apoptotic bodies of islets) | 0.64 ± 0.07 a,b | 0.70 ± 0.09 a | 0.66 ± 0.07 a,b | 0.65 ± 0.07 a,b | 0.59 ± 0.07 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.J.; Kim, M.J.; Kwon, D.Y.; Kim, D.S.; Zhang, T.; Ha, C.; Park, S. Combination of Aronia, Red Ginseng, Shiitake Mushroom and Nattokinase Potentiated Insulin Secretion and Reduced Insulin Resistance with Improving Gut Microbiome Dysbiosis in Insulin Deficient Type 2 Diabetic Rats. Nutrients 2018, 10, 948. https://doi.org/10.3390/nu10070948
Yang HJ, Kim MJ, Kwon DY, Kim DS, Zhang T, Ha C, Park S. Combination of Aronia, Red Ginseng, Shiitake Mushroom and Nattokinase Potentiated Insulin Secretion and Reduced Insulin Resistance with Improving Gut Microbiome Dysbiosis in Insulin Deficient Type 2 Diabetic Rats. Nutrients. 2018; 10(7):948. https://doi.org/10.3390/nu10070948
Chicago/Turabian StyleYang, Hye Jeong, Min Jung Kim, Dae Young Kwon, Da Sol Kim, Ting Zhang, Chulgyu Ha, and Sunmin Park. 2018. "Combination of Aronia, Red Ginseng, Shiitake Mushroom and Nattokinase Potentiated Insulin Secretion and Reduced Insulin Resistance with Improving Gut Microbiome Dysbiosis in Insulin Deficient Type 2 Diabetic Rats" Nutrients 10, no. 7: 948. https://doi.org/10.3390/nu10070948
APA StyleYang, H. J., Kim, M. J., Kwon, D. Y., Kim, D. S., Zhang, T., Ha, C., & Park, S. (2018). Combination of Aronia, Red Ginseng, Shiitake Mushroom and Nattokinase Potentiated Insulin Secretion and Reduced Insulin Resistance with Improving Gut Microbiome Dysbiosis in Insulin Deficient Type 2 Diabetic Rats. Nutrients, 10(7), 948. https://doi.org/10.3390/nu10070948