The Effects of Quassinoid-Rich Eurycoma longifolia Extract on Bone Turnover and Histomorphometry Indices in the Androgen-Deficient Osteoporosis Rat Model

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Blood and Bone Sampling
2.3. Bone Biochemical Markers
2.4. Bone Histomorphometry
2.5. Serum Testosterone, OPG & RANKL
2.6. Statistical Methods
3. Results
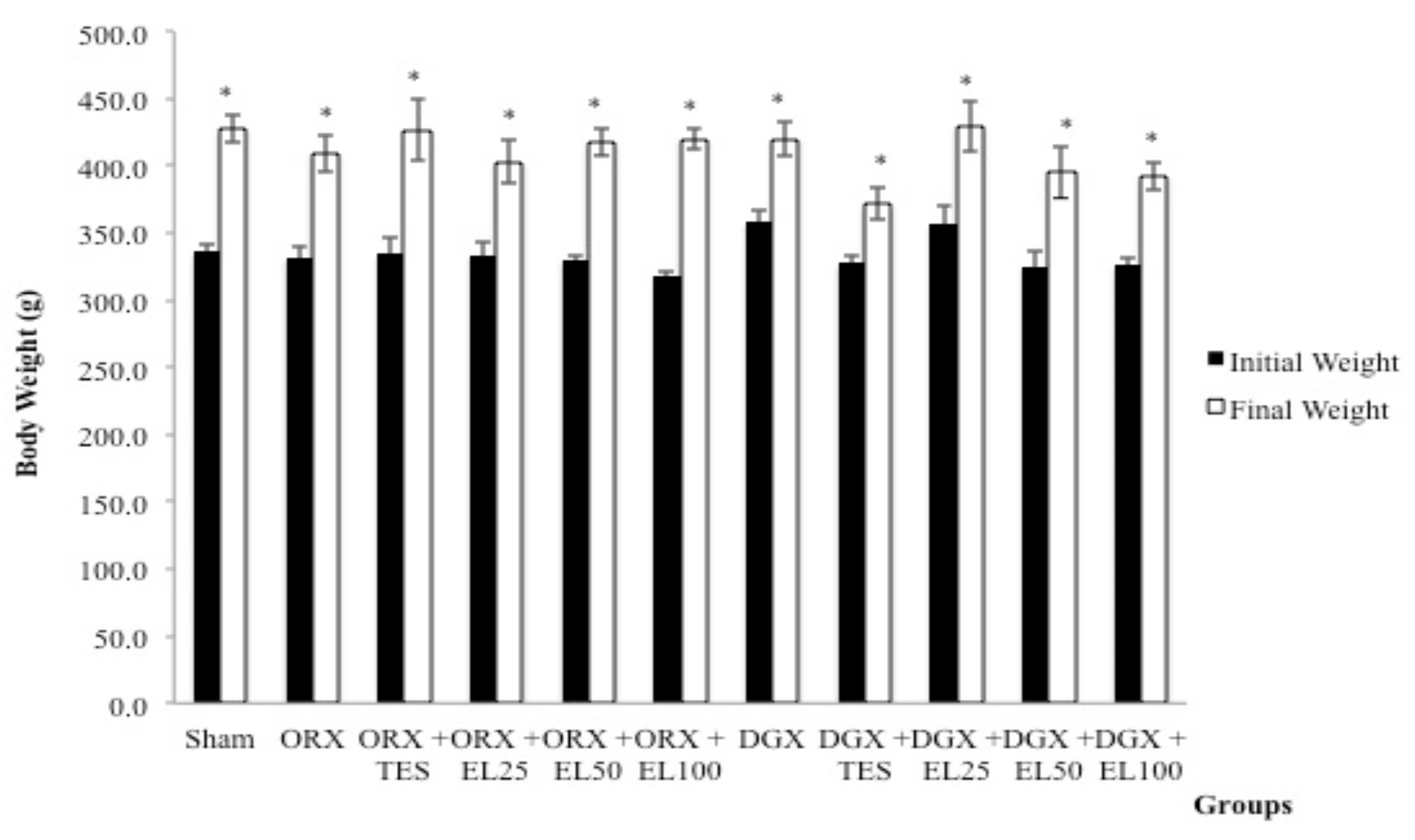
3.1. Body Weight
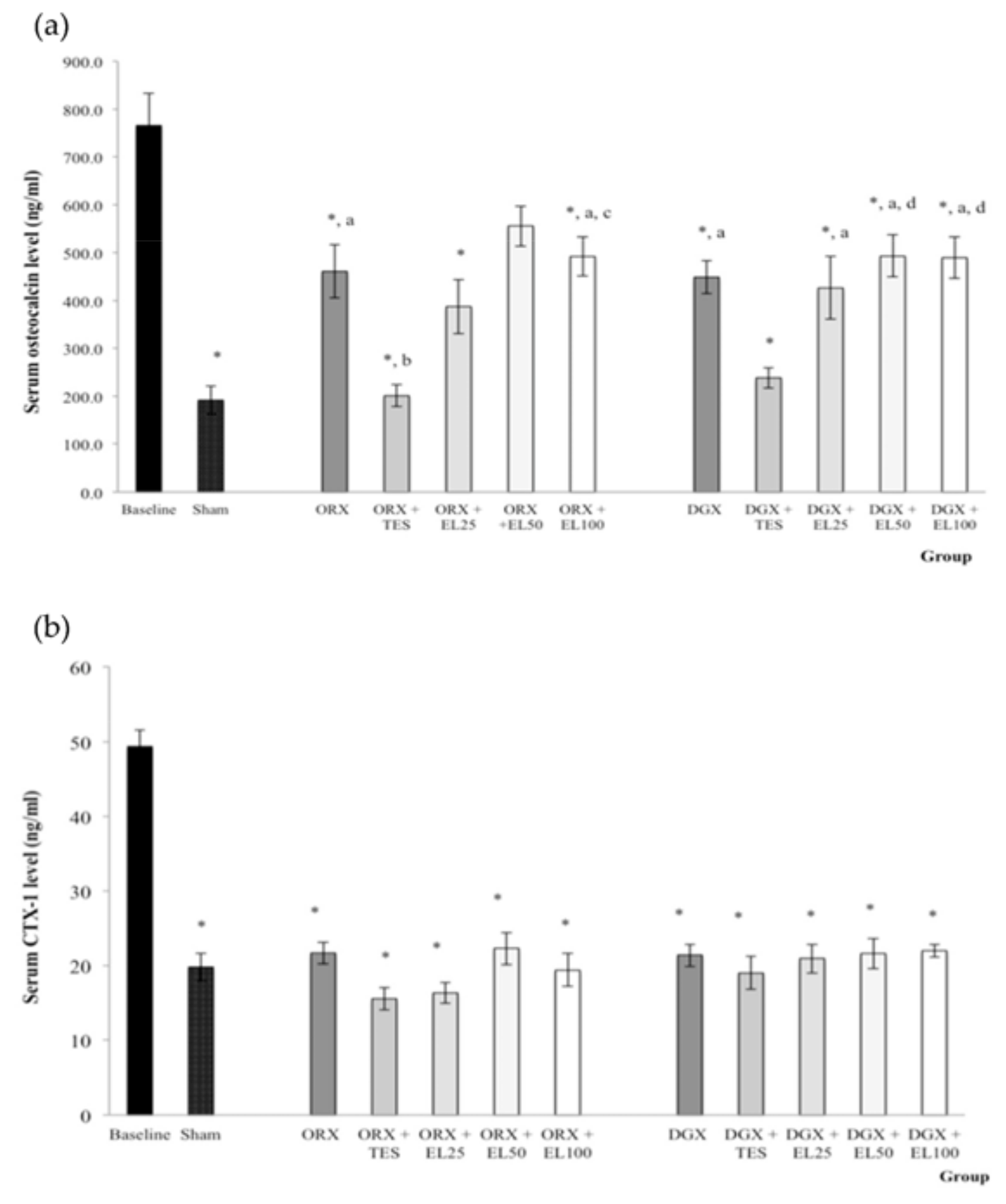
3.2. Bone Biochemical Markers
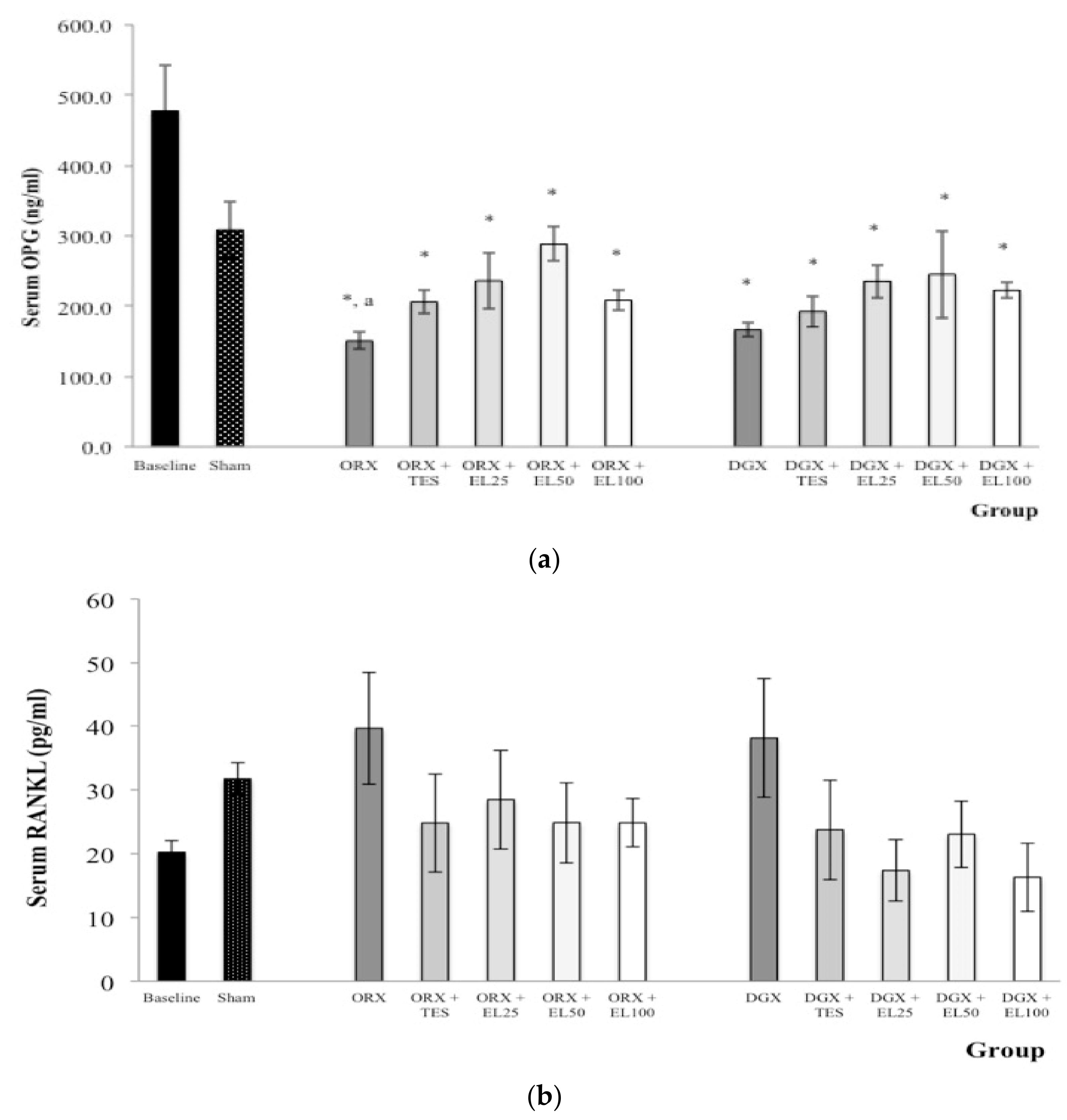
3.3. Serum OPG and RANKL Level
3.4. Serum Testosterone Level
3.5. Static Histomorphometric Parameters
3.6. Structural Histomorphometric Parameters
3.7. Dynamic Histomorphometric Parameters
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Vanderschueren, D.; Laurent, M.R.; Claessens, F.; Gielen, E.; Lagerquist, M.K.; Vandenput, L.; Börjesson, A.E.; Ohlsson, C. Sex steroid actions in male bone. Endocr. Rev. 2014, 35, 906–960. [Google Scholar] [CrossRef] [PubMed]
- Balasch, J. Sex steroids and bone: Current perspectives. Hum. Reprod. Update 2003, 9, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Orwoll, E.S.; Klein, R.F. Osteoporosis in men. Endocr. Rev. 1995, 16, 87–116. [Google Scholar] [CrossRef] [PubMed]
- Kenny, A.M.; Prestwood, K.M.; Marcello, K.M.; Raisz, L.G. Determinants of bone density in healthy older men with low testosterone levels. J. Gerontol. A Biol. Sci. Med. Sci. 2000, 55, M492–M497. [Google Scholar] [CrossRef] [PubMed]
- Blouin, S.; Libouban, H.; Moreau, M.F.; Chappard, D. Orchidectomy models of osteoporosis. In Osteoporosis: Methods and Protocols; Westendorf, J.J., Ed.; Humana Press: New York, NY, USA, 2008; pp. 125–134. ISBN 978-1-58829-828-7. [Google Scholar]
- Erben, R.G.; Eberle, J.; Stahr, K.; Goldberg, M. Androgen deficiency induces high turnover osteopenia in aged male rats: A sequential histomorphometric study. J. Bone Miner. Res. 2000, 15, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Goulding, A.; Gold, E. Flutamide-mediated androgen blockade evokes osteopenia in the female rat. J. Bone Miner. Res. 1993, 8, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Goulding, A.; Gold, E. A new way to induce oestrogen-deficiency osteopaenia in the rat: Comparison of the effects of surgical ovariectomy and administration of the LHRH agonist buserelin on bone resorption and composition. J. Endocr. 1989, 121, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Cook, T.; Sheridan, W.P. Development of GnRH antagonists for prostate cancer: New approaches to treatment. Oncologist 2000, 5, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.W.; Small, E.J. Osteoporosis in men treated with androgen deprivation therapy for prostate cancer. J. Urol. 2002, 167, 1952–1956. [Google Scholar] [CrossRef]
- Broqua, P.; Riviere, P.J.-M.; Conn, P.M.; Rivier, J.E.; Aubert, M.L.; Junien, J.-L. Pharmacological profile of a new, potent, and long-acting gonadotropin-releasing hormone antagonist: Degarelix. J. Pharmacol. Exp. Ther. 2002, 301, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Behre, H.M.; Kliesch, S.; Leifke, E.; Link, T.M.; Nieschlag, E. Long-term effect of testosterone therapy on bone mineral density in hypogonadal men. J. Clin. Endocrinol. Metab. 1997, 82, 2386–2390. [Google Scholar] [CrossRef] [PubMed]
- Shimon, I.; Eshed, V.; Doolman, R.; Sela, B.-A.; Karasik, A.; Vered, I. Alendronate for osteoporosis in men with androgen-repleted hypogonadism. Osteoporos. Int. 2005, 16, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Surampudi, P.N.; Wang, C.; Swerdloff, R. Hypogonadism in the aging male diagnosis, potential benefits, and risks of testosterone replacement therapy. Int. J. Endocrinol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Varenna, M.; Bertoldo, F.; Di Monaco, M.; Giusti, A.; Martini, G.; Rossini, M. Safety profile of drugs used in the treatment of osteoporosis: A systematical review of the literature. Reumatism 2013, 65, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Fernaández-Balsells, M.M.; Murad, M.H.; Lane, M.; Lampropulos, J.F.; Albuquerque, F.; Mullan, R.J.; Agrwal, N.; Elamin, M.B.; Gallegos-Orozco, J.F.; Wang, A.T. Adverse effects of testosterone therapy in adult men: A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2010, 95, 2560–2575. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, S.; Cunningham, G.R.; Hayes, F.J.; Matsumoto, A.M.; Snyder, P.J.; Swerdloff, R.S.; Montori, V.M. Testosterone therapy in men with androgen deficiency syndromes: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2010, 95, 2536–2559. [Google Scholar] [CrossRef] [PubMed]
- Aki, S.; Eskiyurt, N.; Akarirmak, U.; Tuzun, F.; Eryavuz, M.; Alper, S.; Arpacioglu, O.; Atalay, F.; Kavuncu, V.; Kokino, S.; et al. Gastrointestinal side effect profile due to the use of alendronate in the treatment of osteoporosis. Yonsei Med. J. 2003, 44, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.-K.; Yang, K.; Koh, J.; Wong, M.; Chua, S.; Chua, D.; Howe, T. Subtrochanteric insufficiency fractures in patients on alendronate therapy: A caution. Bone Jt. J. 2007, 89, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Karim, A. Tongkat Ali (Eurycoma longifolia Jack): A review on its ethnobotany and pharmacological importance. Fitoterapia 2010, 81, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Liao, J.-W.; Liao, P.-L.; Huang, W.-K.; Tse, L.-S.; Lin, C.-H.; Kang, J.-J.; Cheng, Y.-W. Evaluation of acute 13-week subchronic toxicity and genotoxicity of the powdered root of Tongkat Ali (Eurycoma longifolia Jack). Evid.-Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Ang, H.; Ngai, T. Aphrodisiac evaluation in non-copulator male rats after chronic administration of Eurycoma longifolia Jack. Fundam. Clin. Pharmacol. 2001, 15, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Ang, H.H.; Sim, M.K. Eurycoma longifolia increases sexual motivation in sexually naive male rats. Arch. Pharm. Res. 1998, 21, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Kotirum, S.; Ismail, S.B.; Chaiyakunapruk, N. Efficacy of Tongkat Ali (Eurycoma longifolia) on erectile function improvement: Systematic review and meta-analysis of randomized controlled trials. Complement. Ther. Med. 2015, 23, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Tambi, M.; Imran, M.; Henkel, R. Standardised water-soluble extract of Eurycoma longifolia, Tongkat ali, as testosterone booster for managing men with late-onset hypogonadism? Andrologia 2012, 44, 226–230. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Henkel, R. Phytoandrogenic properties of Eurycoma longifolia as natural alternative to testosterone replacement therapy. Andrologia 2014, 46, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Shuid, A.N.; Abu Bakar, M.F.; Abdul Shukor, T.A.; Muhammad, N.; Mohamed, N.; Soelaiman, I.N. The anti-osteoporotic effect of Eurycoma longifolia in aged orchidectomised rat model. Aging Male 2011, 14, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Abdul Razak, H.S.; Shuid, A.N.; Mohamed, I.N. Combined effects of Eurycoma longifolia and testosterone on androgen-deficient osteoporosis in a male rat model. Evid.-Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Tajul Ariff, A.S.; Soelaiman, I.N.; Pramanik, J.; Shuid, A.N. Effects of Eurycoma longifolia on testosterone level and bone structure in an aged orchidectomised rat model. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Asiah, O.; Nurhanan, M.; Ilham, A.M. Determination of bioactive peptide (4.3 kDa) as an aphrodisiac marker in six Malaysian plants. J. Trop. For. Sci. 2007, 19, 61–63. [Google Scholar]
- Chan, K.-L.; Low, B.-S.; The, C.-H.; Das, P.K. The effect of Eurycoma longifolia on sperm quality of male rats. Nat. Prod. Commun. 2009, 4, 1331–1336. [Google Scholar] [PubMed]
- Low, B.-S.; Das, P.K.; Chan, K.-L. Standardized quassinoid-rich Eurycoma longifolia extract improved spermatogenesis and fertility in male rats via the hypothalamic–pituitary–gonadal axis. J. Ethnopharmacol. 2013, 145, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Low, B.-S.; Choi, S.-B.; Wahab, H.A.; Das, P.K.; Chan, K.-L. Eurycomanone, the major quassinoid in Eurycoma longifolia root extract increases spermatogenesis by inhibiting the activity of phosphodiesterase and aromatase in steroidogenesis. J. Ethnopharmacol. 2013, 149, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, A.M.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R. Bone histomorphometry: Standardization of nomenclature, symbols, and units: Report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 1987, 2, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Michael, H.; Härkönen, P.L.; Väänänen, H.K.; Hentunen, T.A. Estrogen and testosterone use different cellular pathways to inhibit osteoclastogenesis and bone resorption. J. Bone Miner. Res. 2005, 20, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.M.; Bendixen, A.C.; Pathrose, P.; Srivastava, S.; Dienger, K.M.; Shevde, N.K.; Pike, J.W. Androgens suppress osteoclast formation induced by RANKL and macrophage-colony stimulating factor. Endocrinology 2001, 142, 3800–3808. [Google Scholar] [CrossRef] [PubMed]
- Pivonka, P.; Zimak, J.; Smith, D.W.; Gardiner, B.S.; Dunstan, C.R.; Sims, N.A.; Martin, T.J.; Mundy, G.R. Theoretical investigation of the role of the RANK–RANKL–OPG system in bone remodeling. J. Theor. Biol. 2010, 262, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Bu, S.; Lucas, E.; Franklin, M.; Marlow, D.; Brackett, D.; Boldrin, E.; Devareddy, L.; Arjmandi, B.; Smith, B. Comparison of dried plum supplementation and intermittent PTH in restoring bone in osteopenic orchidectomised rats. Osteoporos. Int. 2007, 18, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Saif, A.; Norazlina, M.; Ima-Nirwana, S. Quantification of Bone Histomorphometric Parameters Using the Weibel Technique in Animals. Med. Health 2016, 11, 278–288. [Google Scholar]
- Mehat, M.Z.; Shuid, A.N.; Mohamed, N.; Muhammad, N.; Soelaiman, I.N. Beneficial effects of vitamin E isomer supplementation on static and dynamic bone histomorphometry parameters in normal male rats. J. Bone Miner. Metab. 2010, 28, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.E.; Ellis, R.C. Laboratory Histopathology: A Complete Reference; Elsevier Health Sciences: London, UK, 1994; ISBN 04443049122. [Google Scholar]
- Cohen, A. Chapter 45—Osteoporosis in Premenopausal Women. In Osteoporosis, 4th ed.; Academic Press: San Diego, CA, USA, 2013; pp. 1101–1111. [Google Scholar]
- Soelaiman, I.N.; Ming, W.; Abu Bakar, R.; Hashnan, N.A.; Mohd Ali, H.; Mohamed, N.; Muhammad, N.; Shuid, A.N. Palm tocotrienol supplementation enhanced bone formation in oestrogen-deficient rats. Int. J. Endocrinol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Grimaud, E.; Soubigou, L.; Couillaud, S.; Coipeau, P.; Moreau, A.; Passuti, N.; Gouin, F.; Redini, F.; Heymann, D. Receptor activator of nuclear factor κB ligand (RANKL)/osteoprotegerin (OPG) ratio is increased in severe osteolysis. Am. J. Pathol. 2003, 163, 2021–2031. [Google Scholar] [CrossRef]
- Kawano, H.; Sato, T.; Yamada, T.; Matsumoto, T.; Sekine, K.; Watanabe, T.; Nakamura, T.; Fukuda, T.; Yoshimura, K.; Yoshizawa, T.; et al. Suppressive function of androgen receptor in bone resorption. Proc. Natl. Acad. Sci. USA 2003, 100, 9416–9421. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ominsky, M.S.; Stolina, M.; Warmington, K.S.; Geng, Z.; Niu, Q.-T.; Asuncion, F.J.; Tan, H.-L.; Grisanti, M.; Dwyer, D.; et al. Increased RANK ligand in bone marrow of orchiectomized rats and prevention of their bone loss by the RANK ligand inhibitor osteoprotegerin. Bone 2009, 45, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Low, B.S.; Das, P.K.; Chan, K.L. Acute, Reproductive Toxicity and Two-generation Teratology Studies of a Standardized Quassinoid-rich Extract of Eurycoma longifolia Jack in Sprague-Dawley Rats. Phytother. Res. 2014, 28, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Favus, M.J. Bisphosphonates for osteoporosis. New. Engl. J. Med. 2010, 363, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Thu, H.E.; Mohamed, I.N.; Hussain, Z.; Shuid, A.N. Eurycoma longifolia as a potential alternative to testosterone for the treatment of osteoporosis: Exploring time-mannered proliferative, differentiative and morphogenic modulation in osteoblasts. J. Ethnopharmacol. 2017, 195, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Thu, H.E.; Mohamed, I.N.; Hussain, Z.; Mohamed, N.; Shuid, A.N. Eurycoma longifolia, a malaysian medicinal herb, significantly upregulates proliferation and differentiation in pre-osteoblasts (MC3T3-E1): An in vitro model. Int. J. Pharm. Pharm. Sci. 2016, 8, 199–204. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters Measured | Abbreviation |
|---|---|
| Structural Parameters | |
| Trabecular Volume | BV/TV |
| Trabecular Thickness | TbTh |
| Trabecular Number | TbN |
| Trabecular Separation | TbSp |
| Static Parameters | |
| Osteoblast Surface | ObS/BS |
| Osteoclast Surface | OcS/BS |
| Eroded Surface | ES/BS |
| Osteoid Volume | OV/BV |
| Osteoid Surface | OS/BS |
| Dynamic Parameters | |
| Single-labeled Surface | sLS/BS |
| Double-labeled Surface | dLS/BS |
| Mineralizing Surface | MS/BS |
| Mineral Apposition Rate | MAR |
| Bone Formation Rate | BFR/BS |
| Mean (ng/mL) | SEM | |
|---|---|---|
| Baseline | 12.803 | 2.12 |
| Sham | 6.033 * | 0.74 |
| ORX | 0.604 *,a | 0.06 |
| ORX + TES | 19.263 *,a,b | 2.018 |
| ORX + EL25 | 1.132 *,a,b,c | 0.03 |
| ORX + EL50 | 1.042 *,a,c | 0.21 |
| ORX + EL100 | 1.186 *,a,c | 0.083 |
| DGX | 0.594 *,a | 0.02 |
| DGX + TES | 23.218 *,a,d | 1.21 |
| DGX + EL25 | 1.304 *,a,e | 0.12 |
| DGX + EL50 | 1.589 *,a,d,e | 0.14 |
| DGX + EL100 | 1.1314 *,a,d,e | 0.03 |
| ObS/BS (%) | OcS/BS (%) | ES/BS (%) | OS/BS (%) | OV/BV (%) | |
|---|---|---|---|---|---|
| Baseline | 8.77 ± 0.52 | 1.52 ± 0.36 | 5.43 ± 0.50 | 2.68 ± 0.39 | 3.07 ± 0.32 |
| Sham | 12.53 ± 0.75 | 2.96 ± 0.36 | 4.98 ± 0.57 | 5.02 ± 0.32 * | 6.12 ± 0.41 * |
| ORX | 9.74 ± 0.81 | 7.24 ± 0.44 *,a | 4.13 ± 0.54 | 4.73 ± 0.31 | 5.94 ± 0.59 * |
| ORX + TES | 11.17 ± 0.34 | 5.23 ± 0.24 *,a | 4.81 ± 0.39 | 4.81 ± 0.57 * | 6.27 ± 0.78 * |
| ORX + EL25 | 12.93 ± 0.77 | 4.11 ± 0.59 | 5.04 ± 0.47 | 5.74 ± 0.47 * | 7.37 ± 0.62 * |
| ORX + EL50 | 15.72 ± 1.87 * | 2.87 ± 0.28 b,c | 3.70 ± 0.39 | 5.81 ± 0.69 * | 6.97 ± 0.66 * |
| ORX + EL100 | 12.40 ± 0.89 | 3.49 ± 0.42 b | 4.55 ± 0.41 | 5.63 ± 0.45 * | 6.90 ± 0.82 * |
| DGX | 12.16 ± 0.51 | 4.67 ± 0.23 *,a | 5.98 ± 0.81 | 4.60 ± 0.21 | 5.32 ± 0.40 |
| DGX + TES | 19.36 ± 1.20 *,a,d | 3.98 ± 0.35 * | 4.42 ± 0.59 | 5.58 ± 0.30 * | 7.75 ± 0.64 * |
| DGX + EL25 | 17.95 ± 0.94 *,a,d | 2.38 ± 0.20 d | 4.76 ± 0.51 | 4.55 ± 0.27 | 6.12 ± 0.41 * |
| DGX + EL50 | 16.71 ± 1.45 * | 2.55 ± 0.32 d | 3.63 ± 0.53 | 4.56 ± 0.59 | 5.93 ± 0.55 * |
| DGX + EL100 | 15.46 ± 1.19 * | 2.91 ± 0.40 | 3.54 ± 0.40 | 4.59 ± 0.48 | 5.84 ± 0.40 * |
| BV/TV | TbTh | TbN | TbSp | |
|---|---|---|---|---|
| Baseline | 42.06 ± 2.06 | 139.86 ± 8.68 | 0.0030 ± 0.0001 | 193.62 ± 9.13 |
| Sham | 38.29 ± 2.90 | 167.09 ± 11.80 | 0.0029 ± 0.0002 | 282.06 ± 16.77 |
| ORX | 24.45 ± 0.73 *,a | 114.37 ± 6.91 | 0.0021 ± 0.0001 *,a | 377.25 ± 9.78 *,a |
| ORX + TES | 38.63 ± 2.07 b | 149.65 ± 13.99 | 0.0026 ± 0.0002 | 237.97 ± 17.89 b |
| ORX + EL25 | 31.55 ± 1.59 * | 123.32 ± 3.71 | 0.0025 ± 0.0002 | 276.44 ± 25.99 |
| ORX + EL50 | 28.96 ± 1.29 * | 136.82 ± 7.62 | 0.0020 ± 0.0001 *,a | 317.14 ± 16.59 * |
| ORX + EL100 | 32.36 ± 1.75 * | 128.18 ± 5.83 | 0.0028 ± 0.0001 | 281.18 ± 15.56 b |
| DGX | 19.53 ± 0.94 *,a | 98.96 ± 5.72 | 0.0018 ± 0.0001 *,a | 419.25 ± 16.67 *,a |
| DGX + TES | 33.08 ± 1.88 *,d | 132.31 ± 20.49 | 0.0029 ± 0.0002 d | 212.10 ± 8.55 d |
| DGX + EL25 | 30.01 ± 1.83 *,d | 125.96 ± 13.59 | 0.0025 ± 0.0002 d | 243.18 ± 11.13 d |
| DGX + EL50 | 29.76 ± 0.78 * | 100.47 ± 7.44 | 0.0029 ± 0.0001 d | 240.97 ± 12.53 d |
| DGX + EL100 | 33.56 ± 3.00 d | 107.95 ± 9.24 | 0.0029 ± 0.0002 d | 228.23 ± 10.78 d |
| sLS/BS | dLS/BS | MAR | MS/BS | BFR/BS | |
|---|---|---|---|---|---|
| Baseline | 8.75 ± 0.68 | 9.50 ± 0.41 | 1.39 ± 0.09 | 40.29 ± 3.67 | 54.0 ± 7.55 |
| Sham | 6.29 ± 0.93 | 15.11 ± 1.31 * | 1.35 ± 0.06 | 67.95 ± 9.06 | 73.39 ± 12.85 |
| ORX | 15.71 ± 1.21 *,a | 6.33 ± 0.84 a | 0.76 ± 0.03 *,a | 37.92 ± 5.74 | 40.59 ± 7.99 |
| ORX + TES | 8.18 ± 0.65 b | 10.99 ± 0.99 b | 1.28 ± 0.05 b | 48.23 ± 4.95 | 58.99 ± 8.45 |
| ORX + EL25 | 6.94 ± 0.68 b | 13.80 ± 0.77 b | 1.05 ± 0.09 | 49.22 ± 9.33 | 55.35 ± 10.84 |
| ORX + EL50 | 8.43 ± 1.24 | 11.95 ± 0.57 b | 1.33 ± 0.10 b | 49.47 ± 9.77 | 57.0 ± 16.27 |
| ORX + EL100 | 6.56 ± 0.44 b | 13.07 ± 0.89 b | 1.23 ± 0.10 b | 42.31 ± 4.56 | 51.37 ± 6.93 |
| DGX | 16.94 ± 1.04 *,a | 6.66 ± 0.73 a | 0.90 ± 0.05 *,a | 51.12 ± 0.77 | 52.76 ± 3.72 |
| DGX + TES | 10.75 ± 1.66 | 13.01 ± 0.93 d | 1.26 ± 0.13 | 54.59 ± 7.85 | 68.37 ± 9.26 |
| DGX + EL25 | 9.64 ± 0.93 c | 11.83 ± 0.89 d | 1.35 ± 0.09 d | 51.04 ± 5.61 | 69.20 ± 7.91 |
| DGX + EL50 | 7.75 ± 0.83 c | 11.11 ± 1.04 d | 1.32 ± 0.09 | 51.59 ± 4.36 | 62.94 ± 13.70 |
| DGX + EL100 | 6.22 ± 0.79 c | 11.67 ± 0.79 d | 1.22 ± 0.08 | 50.28 ± 6.83 | 59.28 ± 14.34 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayusman, P.A.; Mohamed, I.N.; Alias, E.; Mohamed, N.; Shuid, A.N. The Effects of Quassinoid-Rich Eurycoma longifolia Extract on Bone Turnover and Histomorphometry Indices in the Androgen-Deficient Osteoporosis Rat Model. Nutrients 2018, 10, 799. https://doi.org/10.3390/nu10070799
Jayusman PA, Mohamed IN, Alias E, Mohamed N, Shuid AN. The Effects of Quassinoid-Rich Eurycoma longifolia Extract on Bone Turnover and Histomorphometry Indices in the Androgen-Deficient Osteoporosis Rat Model. Nutrients. 2018; 10(7):799. https://doi.org/10.3390/nu10070799
Chicago/Turabian StyleJayusman, Putri Ayu, Isa Naina Mohamed, Ekram Alias, Norazlina Mohamed, and Ahmad Nazrun Shuid. 2018. "The Effects of Quassinoid-Rich Eurycoma longifolia Extract on Bone Turnover and Histomorphometry Indices in the Androgen-Deficient Osteoporosis Rat Model" Nutrients 10, no. 7: 799. https://doi.org/10.3390/nu10070799
APA StyleJayusman, P. A., Mohamed, I. N., Alias, E., Mohamed, N., & Shuid, A. N. (2018). The Effects of Quassinoid-Rich Eurycoma longifolia Extract on Bone Turnover and Histomorphometry Indices in the Androgen-Deficient Osteoporosis Rat Model. Nutrients, 10(7), 799. https://doi.org/10.3390/nu10070799