Restoration of Muscle Glycogen and Functional Capacity: Role of Post-Exercise Carbohydrate and Protein Co-Ingestion


Abstract
1. Introduction
2. Post-Exercise Recovery
2.1. Glycogen Structure and Localisation
2.2. The Two Phases of Muscle Glycogen Resynthesis
3. Nutrient Intake and Muscle Glycogen Resynthesis
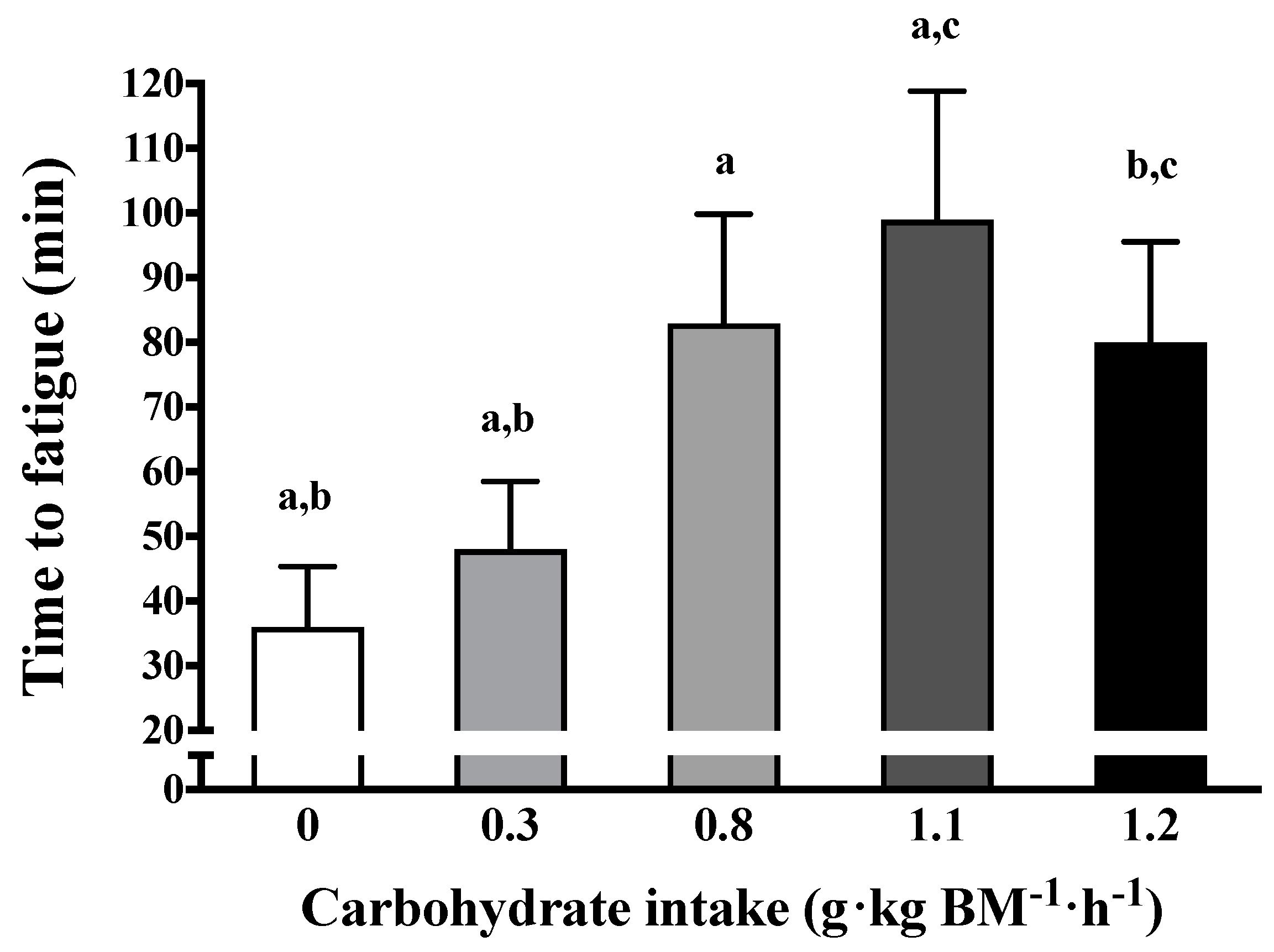
3.1. Amount of Carbohydrate Intake
3.2. Type of Carbohydrate Intake
3.3. Timing of Carbohydrate Intake
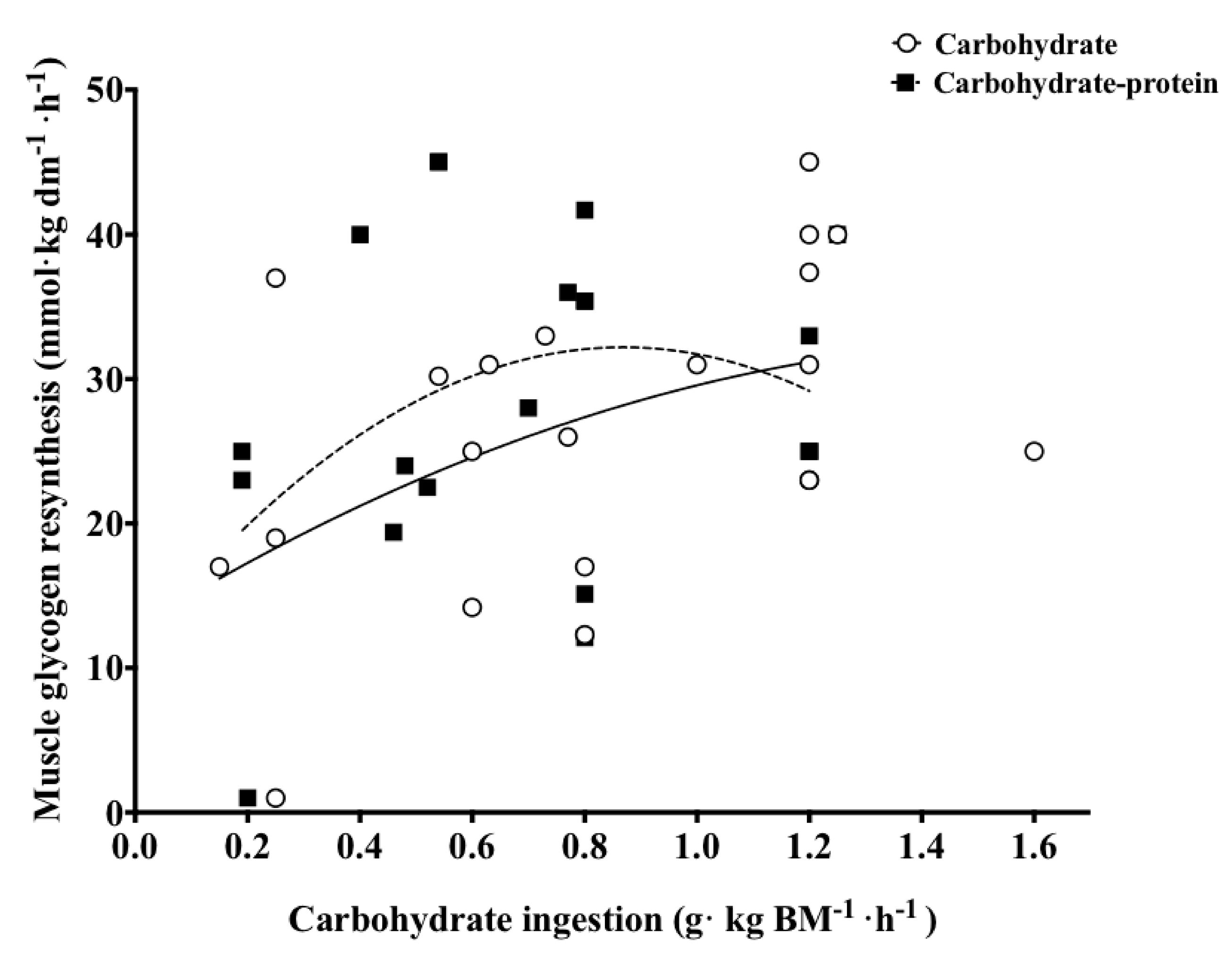
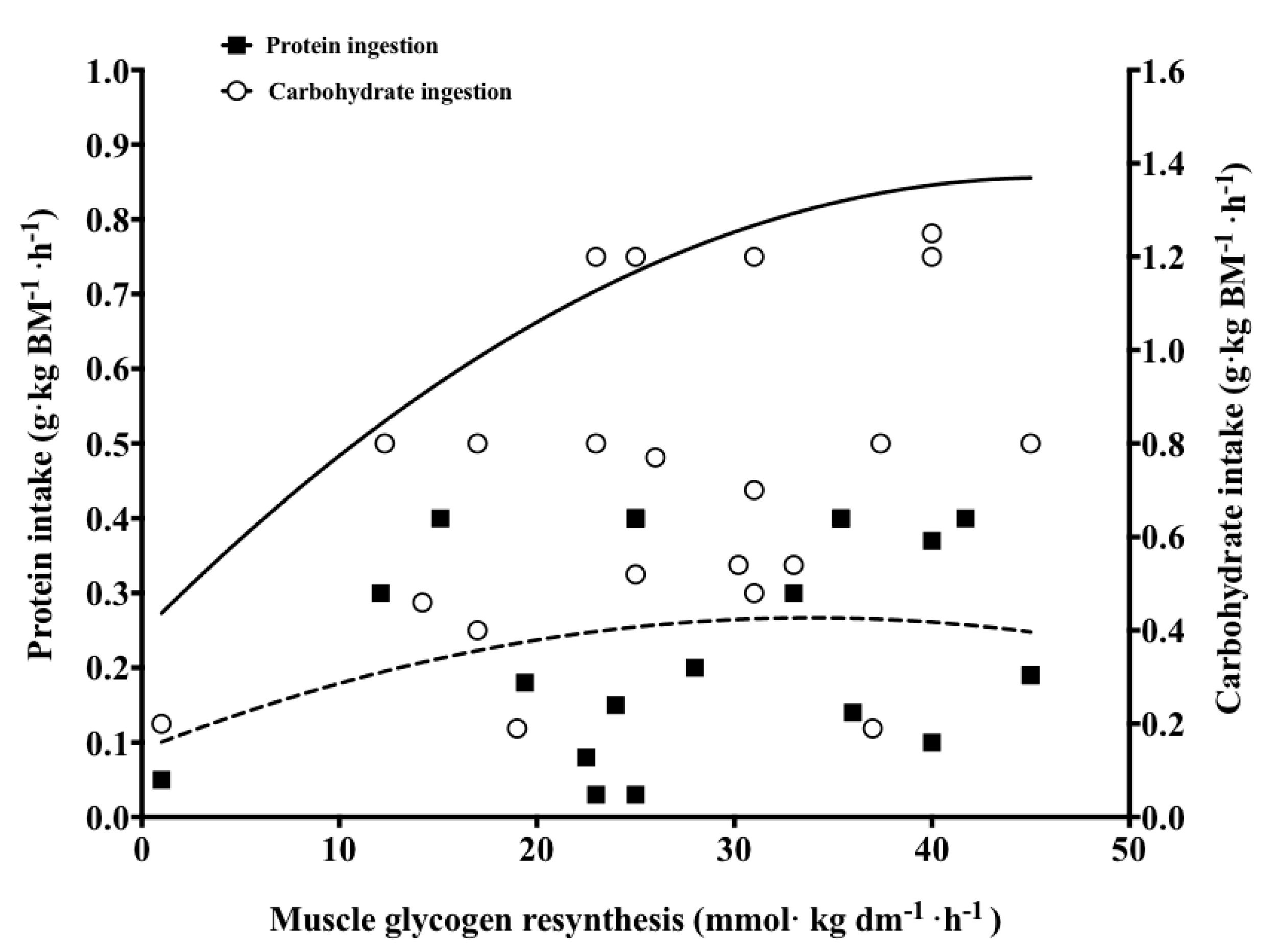
3.4. Protein Co-Ingestion with Carbohydrate
4. Restoration of Exercise Capacity Following Short-Term Recovery
5. Summary
6. Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Reilly, T.; Ekblom, B. The use of recovery methods post-exercise. J. Sports Sci. 2005, 23, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Ament, W.; Verkerke, G.J. Exercise and fatigue. Sports Med. 2009, 39, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Enoka, R.M.; Duchateau, J. Muscle fatigue: What, why and how it influences muscle function. J. Physiol. 2008, 586, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Marino, F.E.; Gard, M.; Drinkwater, E.J. The limits to exercise performance and the future of fatigue research. Br. J. Sports Med. 2011, 45, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S. Fatigue in Sport and Exercise; Routledge: London, UK, 2015. [Google Scholar]
- Alghannam, A.F.; Jedrzejewski, D.; Tweddle, M.; Gribble, H.; Bilzon, J.L.; Betts, J.A. Reliability of time to exhaustion treadmill running as a measure of human endurance capacity. Int. J. Sports Med. 2016, 37, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American college of sports medicine joint position statement. nutrition and athletic performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [PubMed]
- Bergstrom, J.; Hermansen, L.; Hultman, E.; Saltin, B. Diet, muscle glycogen and physical performance. Acta Physiol. 1967, 71, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Coggan, A.R.; Hemmert, M.K.; Ivy, J.L. Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. Appl. Physiol. 1986, 61, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Hermansen, L.; Hultman, E.; Saltin, B. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 1967, 71, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Jeukendrup, A.E.; Wagenmakers, A.J.; Saris, W.H. Fatty acid oxidation is directly regulated by carbohydrate metabolism during exercise. Am. J. Physiol. 1997, 273, E268–E275. [Google Scholar] [CrossRef] [PubMed]
- Karelis, A.D.; Smith, J.W.; Passe, D.H.; Peronnet, F. Carbohydrate administration and exercise performance: What are the potential mechanisms involved? Sports Med. 2010, 40, 747–763. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, K.; Williams, C. Human muscle glycogen metabolism during exercise. Effect of carbohydrate supplementation. Sports Med. 1998, 25, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.; Achten, J.; Jeukendrup, A.E. High oxidation rates from combined carbohydrates ingested during exercise. Med. Sci. Sports Exerc. 2004, 36, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Jentjens, R. Oxidation of carbohydrate feedings during prolonged exercise: Current thoughts, guidelines and directions for future research. Sports Med. 2000, 29, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, O.K.; Williams, C.; Boobis, L.; Greenhaff, P. Carbohydrate ingestion and single muscle fiber glycogen metabolism during prolonged running in men. J. Appl. Physiol. 1996, 81, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.A.; Williams, C. Short-term recovery from prolonged exercise exploring the potential for protein ingestion to accentuate the benefits of carbohydrate supplements. Sports Med. 2010, 40, 941–959. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, K.; Williams, C.; Boobis, L.; Symington, S.; Moorehouse, J.; Garcia-Roves, P.; Nicholas, C. Effect of carbohydrate feeding during recovery from prolonged running on muscle glycogen metabolism during subsequent exercise. Int. J. Sports Med. 2003, 24, 452–458. [Google Scholar] [PubMed]
- Berardi, J.M.; Price, T.B.; Noreen, E.E.; Lemon, P.W.R. Postexercise muscle glycogen recovery enhanced with a carbohydrate-protein supplement. Med. Sci. Sports Exerc. 2006, 38, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.A.; Williams, C.; Boobis, L.; Tsintzas, K. Increased carbohydrate oxidation after ingesting carbohydrate with added protein. Med. Sci. Sports Exerc. 2008, 40, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Alghannam, A.F.; Jedrzejewski, D.; Tweddle, M.G.; Gribble, H.; Bilzon, J.; Thompson, D.; Tsintzas, K.; Betts, J.A. Impact of muscle glycogen availability on the capacity for repeated exercise in man. Med. Sci. Sports Exerc. 2016, 48, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Alghannam, A.F.; Jedrzejewski, D.; Bilzon, J.; Thompson, D.; Tsintzas, K.; Betts, J.A. Influence of post-exercise carbohydrate-protein ingestion on muscle glycogen metabolism in recovery and subsequent running exercise. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.; Williams, C.; Duffy, K.; Gunner, F. The influence of carbohydrate and protein ingestion during recovery from prolonged exercise on subsequent endurance performance. J. Sports Sci. 2007, 25, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Ferguson-Stegall, L.; McCleave, E.L.; Ding, Z.; Doerner, P.G.; Wang, B.; Liao, Y.H.; Kammer, L.; Liu, Y.; Hwang, J.; Dessard, B.M.; et al. Postexercise carbohydrate-protein supplementation improves subsequent exercise performance and intracellular signaling for protein synthesis. J. Strength Cond. Res. 2011, 25, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Mann, R.; Banister, K.; Fox, J.; Morris, P.G.; Macdonald, I.A.; Greenhaff, P.L. Effect of carbohydrate ingestion on glycogen resynthesis in human liver and skeletal muscle, measured by 13C MRS. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E65–E75. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Ortenblad, N. Physiological aspects of the subcellular localization of glycogen in skeletal muscle. Appl. Physiol. Nutr. Metab. 2013, 38, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Nybo, L.; Nielsen, B. Hyperthermia and central fatigue during prolonged exercise in humans. J. Appl. Physiol. 2001, 91, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F. Physical activity as a metabolic stressor. Am. J. Clin. Nutr. 2000, 72, 512S–520S. [Google Scholar] [CrossRef] [PubMed]
- Sawka, M.N.; Noakes, T.D. Does dehydration impair exercise performance? Med. Sci. Sports Exerc. 2007, 39, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, D.; Merimee, T.J.; Maffezzoli, R.; Burgess, J.A. Patterns of hormonal release after glucose, protein, and glucose plus protein. Lancet 1966, 2, 454–456. [Google Scholar] [CrossRef]
- Van Loon, L.J.; Kruijshoop, M.; Verhagen, H.; Saris, W.H.; Wagenmakers, A.J. Ingestion of protein hydrolysate and amino acid-carbohydrate mixtures increases postexercise plasma insulin responses in men. J. Nutr. 2000, 130, 2508–2513. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.; van Loon, L.J.; Mann, C.H.; Wagenmakers, A.J.; Jeukendrup, A.E. Addition of protein and amino acids to carbohydrates does not enhance postexercise muscle glycogen synthesis. J. Appl. Physiol. 2001, 91, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, G.; Shirreffs, S.M.; Calbet, J.A. Muscle glycogen resynthesis during recovery from cycle exercise: No effect of additional protein ingestion. J. Appl. Physiol. 2000, 88, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.; Saris, W.H.; Kruijshoop, M.; Wagenmakers, A.J. Maximizing postexercise muscle glycogen synthesis: Carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures. Am. J. Clin. Nutr. 2000, 72, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Howarth, K.R.; Moreau, N.A.; Phillips, S.M.; Gibala, M.J. Coingestion of protein with carbohydrate during recovery from endurance exercise stimulates skeletal muscle protein synthesis in humans. J. Appl. Physiol. 2009, 106, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Beelen, M.; Kranenburg, J.; Senden, J.M.; Kuipers, H.; van Loon, L.J. Impact of caffeine and protein on postexercise muscle glycogen synthesis. Med. Sci. Sports Exerc. 2012, 44, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Carrithers, J.A.; Williamson, D.L.; Gallagher, P.M.; Godard, M.P.; Schulze, K.E.; Trappe, S.W. Effects of postexercise carbohydrate-protein feedings on muscle glycogen restoration. J. Appl. Physiol. 2000, 88, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L.; Goforth, H.W., Jr.; Damon, B.M.; McCauley, T.R.; Parsons, E.C.; Price, T.B. Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement. J. Appl. Physiol. 2002, 93, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Lunn, W.R.; Pasiakos, S.M.; Colletto, M.R.; Karfonta, K.E.; Carbone, J.W.; Anderson, J.M.; Rodriguez, N.R. Chocolate milk and endurance exercise recovery: Protein balance, glycogen, and performance. Med. Sci. Sports Exerc. 2012, 44, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Morris, P.; Stevenson, E. Improved endurance capacity following chocolate milk consumption compared with 2 commercially available sport drinks. Appl. Physiol. Nutr. Metab. 2009, 34, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.; Jeukendrup, A. Determinants of post-exercise glycogen synthesis during short-term recovery. Sports Med. 2003, 33, 117–144. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.J.; Pagliassotti, M.J.; Hobbs, K.; Hill, J.O. Fuel metabolism in men and women during and after long-duration exercise. J. Appl. Physiol. 1998, 85, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Kiens, B.; Richter, E.A. Utilization of skeletal muscle triacylglycerol during postexercise recovery in humans. Am. J. Physiol. 1998, 275, E332–E337. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed]
- Goforth, H.W., Jr.; Laurent, D.; Prusaczyk, W.K.; Schneider, K.E.; Petersen, K.F.; Shulman, G.I. Effects of depletion exercise and light training on muscle glycogen supercompensation in men. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1304–E1311. [Google Scholar] [CrossRef] [PubMed]
- Melendez, R.; Melendez-Hevia, E.; Canela, E.I. The fractal structure of glycogen: A clever solution to optimize cell metabolism. Biophys. J. 1999, 77, 1327–1332. [Google Scholar] [CrossRef]
- Alonso, M.D.; Lomako, J.; Lomako, W.M.; Whelan, W.J. A new look at the biogenesis of glycogen. FASEB J. 1995, 9, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.E.; Yuan, Z.; Hill, A.K.; Wilson, R.J. The regulation of muscle glycogen: The granule and its proteins. Acta Physiol. 2010, 199, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Jansson, E. Acid soluble and insoluble glycogen in human skeletal muscle. Acta Physiol. 1981, 113, 337–340. [Google Scholar] [CrossRef] [PubMed]
- James, A.P.; Barnes, P.D.; Palmer, T.N.; Fournier, P.A. Proglycogen and macroglycogen: Artifacts of glycogen extraction? Metabolism 2008, 57, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Holmberg, H.C.; Schroder, H.D.; Saltin, B.; Ortenblad, N. Human skeletal muscle glycogen utilization in exhaustive exercise: Role of subcellular localization and fibre type. J. Physiol. 2011, 589, 2871–2885. [Google Scholar] [CrossRef] [PubMed]
- Ortenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Tarnopolsky, M.; Adamo, K.B.; Bourgeois, J.M.; Chorneyko, K.; Graham, T.E. Quantitative assessment of human muscle glycogen granules size and number in subcellular locations during recovery from prolonged exercise. J. Physiol. 2007, 580, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Schroder, H.D.; Rix, C.G.; Ortenblad, N. Distinct effects of subcellular glycogen localization on tetanic relaxation time and endurance in mechanically skinned rat skeletal muscle fibres. J. Physiol. 2009, 587, 3679–3690. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Cheng, A.J.; Ortenblad, N.; Westerblad, H. Subcellular distribution of glycogen and decreased tetanic Ca2+ in fatigued single intact mouse muscle fibres. J. Physiol. 2014, 592, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Gejl, K.D.; Hvid, L.G.; Frandsen, U.; Jensen, K.; Sahlin, K.; Ortenblad, N. Muscle glycogen content modifies SR Ca2+ release rate in elite endurance athletes. Med. Sci. Sports Exerc. 2014, 46, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Ortenblad, N.; Nielsen, J.; Saltin, B.; Holmberg, H.C. Role of glycogen availability in sarcoplasmic reticulum Ca2+ kinetics in human skeletal muscle. J. Physiol. 2011, 589, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Chin, E.R.; Balnave, C.D.; Allen, D.G. Role of intracellular calcium and metabolites in low-frequency fatigue of mouse skeletal muscle. Am. J. Physiol. 1997, 272, C550–C559. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.; Westerblad, H.; Katz, A. Effects of glucose on contractile function, [Ca2+]i, and glycogen in isolated mouse skeletal muscle. Am. J. Physiol. Cell Physiol. 2002, 282, C1306–C1312. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maehlum, S.; Hostmark, A.T.; Hermansen, L. Synthesis of muscle glycogen during recovery after prolonged severe exercise in diabetic and non-diabetic subjects. Scand. J. Clin. Lab. Investig. 1977, 37, 309–316. [Google Scholar] [CrossRef]
- Price, T.B.; Rothman, D.L.; Taylor, R.; Avison, M.J.; Shulman, G.I.; Shulman, R.G. Human muscle glycogen resynthesis after exercise: Insulin- dependent and -independent phases. J. Appl. Physiol. 1994, 76, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Price, T.B.; Perseghin, G.; Duleba, A.; Chen, W.; Chase, J.; Rothman, D.L.; Shulman, R.G.; Shulman, G.I. NMR studies of muscle glycogen synthesis in insulin-resistant offspring of parents with non-insulin-dependent diabetes mellitus immediately after glycogen-depleting exercise. Proc. Natl. Acad. Sci. USA 1996, 93, 5329–5334. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L.; Kuo, C.H. Regulation of GLUT4 protein and glycogen synthase during muscle glycogen synthesis after exercise. Acta Physiol. Scand. 1998, 162, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L.; Lee, M.C.; Brozinick, J.T., Jr.; Reed, M.J. Muscle glycogen storage after different amounts of carbohydrate ingestion. J. Appl. Physiol. 1988, 65, 2018–2023. [Google Scholar] [CrossRef] [PubMed]
- Goodyear, L.J.; Hirshman, M.F.; King, P.A.; Horton, E.D.; Thompson, C.M.; Horton, E.S. Skeletal muscle plasma membrane glucose transport and glucose transporters after exercise. J. Appl. Physiol. 1990, 68, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Danforth, W.H. Glycogen synthetase activity in skeletal muscle. Interconversion of two forms and control of glycogen synthesis. J. Biol. Chem. 1965, 240, 588–593. [Google Scholar] [PubMed]
- Cartee, G.D.; Young, D.A.; Sleeper, M.D.; Zierath, J.; Wallberg-Henriksson, H.; Holloszy, J.O. Prolonged increase in insulin-stimulated glucose transport in muscle after exercise. Am. J. Physiol. 1989, 256, E494–E499. [Google Scholar] [CrossRef] [PubMed]
- Keizer, H.A.; Kuipers, H.; van Kranenburg, G.; Geurten, P. Influence of liquid and solid meals on muscle glycogen resynthesis, plasma fuel hormone response, and maximal physical working capacity. Int. J. Sports Med. 1987, 8, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.C.; Zarrinpashneh, E.; Jensen, J. Additive effect of contraction and insulin on glucose uptake and glycogen synthase in muscle with different glycogen contents. J. Appl. Physiol. 2010, 108, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.S.; Gao, J.; Han, D.H.; Holloszy, J.O.; Nolte, L.A. Activation of amp kinase enhances sensitivity of muscle glucose transport to insulin. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E18–E23. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszewski, J.F.; MacDonald, C.; Nielsen, J.N.; Hellsten, Y.; Hardie, D.G.; Kemp, B.E.; Kiens, B.; Richter, E.A. Regulation of 5′ AMP-activated protein kinase activity and substrate utilization in exercising human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E813–E822. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Kiens, B.; Ivy, J.L. Carbohydrates and fat for training and recovery. J. Sports Sci. 2004, 22, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Beelen, M.; Burke, L.M.; Gibala, M.J.; van Loon, L.J.C. Nutritional strategies to promote postexercise recovery. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Tarnopolsky, M.A.; Bosman, M.; MacDonald, J.R.; Vandeputte, D.; Martin, J.; Roy, B.D. Postexercise protein-carbohydrate and carbohydrate supplements increase muscle glycogen in men and women. J. Appl. Physiol. 1997, 83, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.D.; Tarnopolsky, M.A. Influence of differing macronutrient intakes on muscle glycogen resynthesis after resistance exercise. J. Appl. Physiol. 1998, 84, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.; Gejl, K.D.; Ortenblad, N.; Nielsen, J.L.; Bech, R.D.; Nygaard, T.; Sahlin, K.; Frandsen, U. Carbohydrate restricted recovery from long term endurance exercise does not affect gene responses involved in mitochondrial biogenesis in highly trained athletes. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Philp, A.; Hargreaves, M.; Baar, K. More than a store: Regulatory roles for glycogen in skeletal muscle adaptation to exercise. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1343–E1351. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.D.; Hawley, J.A.; Morton, J.P. Carbohydrate availability and exercise training adaptation: Too much of a good thing? Eur. J. Sport Sci. 2015, 15, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Maehlum, S.; Hermansen, L. Muscle glycogen concentration during recovery after prolonged severe exercise in fasting subjects. Scand. J. Clin. Lab. Investig. 1978, 38, 557–560. [Google Scholar] [CrossRef]
- Blom, P.C.; Hostmark, A.T.; Vaage, O.; Kardel, K.R.; Maehlum, S. Effect of different post-exercise sugar diets on the rate of muscle glycogen synthesis. Med. Sci. Sports Exerc. 1987, 19, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Blom, C.S. Post-exercise glucose uptake and glycogen synthesis in human muscle during oral or IV glucose intake. Eur. J. Appl. Physiol. Occup. Physiol. 1989, 59, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Zachwieja, J.J.; Costill, D.L.; Pascoe, D.D.; Robergs, R.A.; Fink, W.J. Influence of muscle glycogen depletion on the rate of resynthesis. Med. Sci. Sports Exerc. 1991, 23, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Short, A.H.; Hultman, E.; Greenhaff, P.L. Glycogen resynthesis in human muscle fibre types following exercise-induced glycogen depletion. J. Physiol. 1995, 483, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Shearer, J.; Wilson, R.J.; Battram, D.S.; Richter, E.A.; Robinson, D.L.; Bakovic, M.; Graham, T.E. Increases in glycogenin and glycogenin mrna accompany glycogen resynthesis in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E508–E514. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.; Aslesen, R.; Ivy, J.L.; Brors, O. Role of glycogen concentration and epinephrine on glucose uptake in rat epitrochlearis muscle. Am. J. Physiol. 1997, 272, E649–E655. [Google Scholar] [CrossRef] [PubMed]
- Derave, W.; Lund, S.; Holman, G.D.; Wojtaszewski, J.; Pedersen, O.; Richter, E.A. Contraction-stimulated muscle glucose transport and GLUT-4 surface content are dependent on glycogen content. Am. J. Physiol. 1999, 277, E1103–E1110. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; van Hall, G.; Keller, C.; Osada, T.; Schjerling, P.; Pedersen, B.K.; Saltin, B.; Febbraio, M.A. Muscle glycogen content and glucose uptake during exercise in humans: Influence of prior exercise and dietary manipulation. J. Physiol. 2002, 541, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Kiens, B.; Raben, A.B.; Valeur, A.-K.; Richter, E.A. Benefit of dietary simple carbohydrates on the early postexercise muscle glycogen repletion in male athletes. Med. Sci. Sports Exerc. 1990, 22, S89. [Google Scholar]
- Burke, L.M.; Collier, G.R.; Hargreaves, M. Muscle glycogen storage after prolonged exercise: Effect of the glycemic index of carbohydrate feedings. J. Appl. Physiol. 1993, 75, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Wee, S.L.; Williams, C.; Tsintzas, K.; Boobis, L. Ingestion of a high-glycemic index meal increases muscle glycogen storage at rest but augments its utilization during subsequent exercise. J. Appl. Physiol. 2005, 99, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Delarue, J.; Normand, S.; Pachiaudi, C.; Beylot, M.; Lamisse, F.; Riou, J.P. The contribution of naturally labelled 13C fructose to glucose appearance in humans. Diabetologia 1993, 36, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.H.; Hultman, E. Liver and muscle glycogen in man after glucose and fructose infusion. Scand. J. Clin. Lab. Investig. 1974, 33, 5–10. [Google Scholar] [CrossRef]
- Bergstrom, J.; Hultman, E. Synthesis of muscle glycogen in man after glucose and fructose infusion. Acta Med. Scand. 1967, 182, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, A.J.; Houtman, S.; Heerschap, A.; Rehrer, N.J.; van den Boogert, H.J.; Oeseburg, B.; Hopman, M.T. Muscle glycogen recovery after exercise during glucose and fructose intake monitored by 13C-NMR. J. Appl. Physiol. 1996, 81, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Wallis, G.A.; Hulston, C.J.; Mann, C.H.; Roper, H.P.; Tipton, K.D.; Jeukendrup, A.E. Postexercise muscle glycogen synthesis with combined glucose and fructose ingestion. Med. Sci. Sports Exerc. 2008, 40, 1789–1794. [Google Scholar] [CrossRef] [PubMed]
- Trommelen, J.; Beelen, M.; Pinckaers, P.J.; Senden, J.M.; Cermak, N.M.; van Loon, L.J. Fructose coingestion does not accelerate postexercise muscle glycogen repletion. Med. Sci. Sports Exerc. 2016, 48, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.J.; Gonzalez, J.T.; Beelen, M.; Cermak, N.M.; Smith, F.E.; Thelwall, P.E.; Taylor, R.; Trenell, M.I.; Stevenson, E.J.; van Loon, L.J. Sucrose ingestion after exhaustive exercise accelerates liver, but not muscle glycogen repletion compared with glucose ingestion in trained athletes. J. Appl. Physiol. 2016, 120, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.T.; Fuchs, C.J.; Betts, J.A.; van Loon, L.J. Glucose plus fructose ingestion for post-exercise recovery-greater than the sum of its parts? Nutrients 2017, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Maunder, E.; Podlogar, T.; Wallis, G.A. Postexercise fructose-maltodextrin ingestion enhances subsequent endurance capacity. Med. Sci. Sports Exerc. 2017. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.J.; Brozinick, J.T., Jr.; Lee, M.C.; Ivy, J.L. Muscle glycogen storage postexercise: Effect of mode of carbohydrate administration. J. Appl. Physiol. 1989, 66, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Wallis, G.A.; Wittekind, A. Is there a specific role for sucrose in sports and exercise performance? Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 571–583. [Google Scholar] [CrossRef]
- Lecoultre, V.; Benoit, R.; Carrel, G.; Schutz, Y.; Millet, G.P.; Tappy, L.; Schneiter, P. Fructose and glucose co-ingestion during prolonged exercise increases lactate and glucose fluxes and oxidation compared with an equimolar intake of glucose. Am. J. Clin. Nutr. 2010, 92, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L. The regulation and synthesis of muscle glycogen by means of nutrient intervention. Encycl. Sports Med. 2013, 19, 113–125. [Google Scholar]
- Levenhagen, D.K.; Gresham, J.D.; Carlson, M.G.; Maron, D.J.; Borel, M.J.; Flakoll, P.J. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E982–E993. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L. Dietary strategies to promote glycogen synthesis after exercise. Can. J. Appl. Physiol. 2001, 26, S236–S245. [Google Scholar] [CrossRef]
- Parkin, J.A.; Carey, M.F.; Martin, I.K.; Stojanovska, L.; Febbraio, M.A. Muscle glycogen storage following prolonged exercise: Effect of timing of ingestion of high glycemic index food. Med. Sci. Sports Exerc. 1997, 29, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.; Jebens, E.; Brennesvik, E.O.; Ruzzin, J.; Soos, M.A.; Engebretsen, E.M.; O’Rahilly, S.; Whitehead, J.P. Muscle glycogen inharmoniously regulates glycogen synthase activity, glucose uptake, and proximal insulin signaling. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E154–E162. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Sherman, W.M.; Fink, W.J.; Maresh, C.; Witten, M.; Miller, J.M. The role of dietary carbohydrates in muscle glycogen resynthesis after strenuous running. Am. J. Clin. Nutr. 1981, 34, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Collier, G.R.; Davis, P.G.; Fricker, P.A.; Sanigorski, A.J.; Hargreaves, M. Muscle glycogen storage after prolonged exercise: Effect of the frequency of carbohydrate feedings. Am. J. Clin. Nutr. 1996, 64, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L. Glycogen resynthesis after exercise: Effect of carbohydrate intake. Int. J. Sports Med. 1998, 19, S142–S145. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.A.; Sherman, W.M.; Strauss, R.L. Effects of eccentric and concentric exercise on muscle glycogen replenishment. J. Appl. Physiol. 1993, 74, 1848–1855. [Google Scholar] [CrossRef] [PubMed]
- Floyd, J.C., Jr.; Fajans, S.S.; Conn, J.W.; Knopf, R.F.; Rull, J. Insulin secretion in response to protein ingestion. J. Clin. Investig. 1966, 45, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Floyd, J.C., Jr.; Fajans, S.S.; Pek, S.; Thiffault, C.A.; Knopf, R.F.; Conn, J.W. Synergistic effect of certain amino acid pairs upon insulin secretion in man. Diabetes 1970, 19, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.T.; Fuchs, C.J.; Smith, F.E.; Thelwall, P.E.; Taylor, R.; Stevenson, E.J.; Trenell, M.I.; Cermak, N.M.; van Loon, L.J. Ingestion of glucose or sucrose prevents liver but not muscle glycogen depletion during prolonged endurance-type exercise in trained cyclists. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E1032–E1039. [Google Scholar] [CrossRef] [PubMed]
- Mace, O.J.; Schindler, M.; Patel, S. The regulation of K- and L -cell activity by GLUT2 and the calcium-sensing receptor casr in rat small intestine. J. Physiol. 2012, 590, 2917–2936. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Crombach, N.; Gijsen, A.P.; Walrand, S.; Fauquant, J.; Kies, A.K.; Lemosquet, S.; Saris, W.H.; Boirie, Y.; van Loon, L.J. Ingestion of a protein hydrolysate is accompanied by an accelerated in vivo digestion and absorption rate when compared with its intact protein. Am. J. Clin. Nutr. 2009, 90, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Morifuji, M.; Ishizaka, M.; Baba, S.; Fukuda, K.; Matsumoto, H.; Koga, J.; Kanegae, M.; Higuchi, M. Comparison of different sources and degrees of hydrolysis of dietary protein: Effect on plasma amino acids, dipeptides, and insulin responses in human subjects. J. Agric. Food Chem. 2010, 58, 8788–8797. [Google Scholar] [CrossRef] [PubMed]
- Reitelseder, S.; Agergaard, J.; Doessing, S.; Helmark, I.C.; Lund, P.; Kristensen, N.B.; Frystyk, J.; Flyvbjerg, A.; Schjerling, P.; van Hall, G.; et al. Whey and casein labeled with L-[1-13C] leucine and muscle protein synthesis: Effect of resistance exercise and protein ingestion. Am. J. physiol. Metab. 2011, 300, E231–E242. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.; Saris, W.H.; Verhagen, H.; Wagenmakers, A.J. Plasma insulin responses after ingestion of different amino acid or protein mixtures with carbohydrate. Am. J. Clin. Nutr. 2000, 72, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Di Camillo, B.; Eduati, F.; Nair, S.K.; Avogaro, A.; Toffolo, G.M. Leucine modulates dynamic phosphorylation events in insulin signaling pathway and enhances insulin-dependent glycogen synthesis in human skeletal muscle cells. BMC Cell Biol. 2014, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Morifuji, M.; Aoyama, T.; Nakata, A.; Sambongi, C.; Koga, J.; Kurihara, K.; Kanegae, M.; Suzuki, K.; Higuchi, M. Post-exercise ingestion of different amounts of protein affects plasma insulin concentration in humans. Eur. J. Sport Sci. 2012, 12, 152–160. [Google Scholar] [CrossRef]
- Zawadzki, K.; Yaspelkis, B.B., 3rd; Ivy, J. Carbohydrate-protein complex increases the rate of muscle glycogen storage after exercise. J. Appl. Physiol. 1992, 72, 1854–1859. [Google Scholar] [CrossRef] [PubMed]
- Kaastra, B.; Manders, R.J.; van Breda, E.; Kies, A.; Jeukendrup, A.E.; Keizer, H.A.; Kuipers, H.; van Loon, L.J. Effects of increasing insulin secretion on acute postexercise blood glucose disposal. Med. Sci. Sports Exerc. 2006, 38, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.B.; Raven, P.B.; Fogt, D.L.; Ivy, J.L. Effects of recovery beverages on glycogen restoration and endurance exercise performance. J. Strength Cond. Res. 2003, 17, 12–19. [Google Scholar] [PubMed]
- Morifuji, M.; Kanda, A.; Koga, J.; Kawanaka, K.; Higuchi, M. Post-exercise carbohydrate plus whey protein hydrolysates supplementation increases skeletal muscle glycogen level in rats. Amino Acid. 2010, 38, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Hara, D.; Morrison, P.J.; Ding, Z.; Ivy, J.L. Effect of carbohydrate-protein supplementation postexercise on rat muscle glycogen synthesis and phosphorylation of proteins controlling glucose storage. Metabolism 2011, 60, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Rotman, S.; Slotboom, J.; Kreis, R.; Boesch, C.; Jequier, E. Muscle glycogen recovery after exercise measured by 13C-magnetic resonance spectroscopy in humans: Effect of nutritional solutions. Magn. Reson. Mater. Phys. Biol. Med. 2000, 11, 114–121. [Google Scholar] [CrossRef]
- Detko, E.; O’Hara, J.P.; Thelwall, P.E.; Smith, F.E.; Jakovljevic, D.G.; King, R.F.; Trenell, M.I. Liver and muscle glycogen repletion using 13C magnetic resonance spectroscopy following ingestion of maltodextrin, galactose, protein and amino acids. Br. J. Nutr. 2013, 110, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ding, Z.; Wang, W.; Hwang, J.; Liao, Y.H.; Ivy, J.L. The effect of an amino acid beverage on glucose response and glycogen replenishment after strenuous exercise. Eur. J. Appl. Physiol. 2015, 115, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Cogan, K.E.; Evans, M.; Iuliano, E.; Melvin, A.; Susta, D.; Neff, K.; de Vito, G.; Egan, B. Co-ingestion of protein or a protein hydrolysate with carbohydrate enhances anabolic signaling, but not glycogen resynthesis, following recovery from prolonged aerobic exercise in trained cyclists. Eur. J. Appl. Physiol. 2018, 118, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Price, T.B.; Rothman, D.L.; Shulman, R.G.; Shulman, G.I. Validation of 13C NMR measurement of human skeletal muscle glycogen by direct biochemical assay of needle biopsy samples. Magn. Reson. Med. 1992, 27, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, S.; Asp, S.; Richter, E.A. Decreased muscle GLUT-4 and contraction-induced glucose transport after eccentric contractions. Am. J. Physiol. 1996, 271, R477–R482. [Google Scholar] [CrossRef] [PubMed]
- Asp, S.; Watkinson, A.; Oakes, N.D.; Kraegen, E.W. Prior eccentric contractions impair maximal insulin action on muscle glucose uptake in the conscious rat. J. Appl. Physiol. 1997, 82, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Pascoe, D.D.; Fink, W.J.; Robergs, R.A.; Barr, S.I.; Pearson, D. Impaired muscle glycogen resynthesis after eccentric exercise. J. Appl. Physiol. 1990, 69, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Mochizuki, S.; Sugahara, K.; Yoshizawa, F. Isoleucine, a blood glucose-lowering amino acid, increases glucose uptake in rat skeletal muscle in the absence of increases in amp-activated protein kinase activity. J. Nutr. 2005, 135, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.R.; Liao, Y.H.; Hara, D.; Ding, Z.; Chen, C.Y.; Nelson, J.L.; Ivy, J.L. An amino acid mixture improves glucose tolerance and insulin signaling in sprague-dawley rats. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E752–E760. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, M.; Liao, Y.H.; Nelson, J.L.; Bernard, J.R.; Wang, W.; Ivy, J.L. An amino acid mixture enhances insulin-stimulated glucose uptake in isolated rat epitrochlearis muscle. J. Appl. Physiol. 2011, 111, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Spiller, G.A.; Jensen, C.D.; Pattison, T.S.; Chuck, C.S.; Whittam, J.H.; Scala, J. Effect of protein dose on serum glucose and insulin response to sugars. Am. J. Clin. Nutr. 1987, 46, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Cheung, P.; Stein, T.P.; Kresge, K.; Mozzoli, M. FFA cause hepatic insulin resistance by inhibiting insulin suppression of glycogenolysis. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E12–E19. [Google Scholar] [CrossRef] [PubMed]
- Ahlborg, B.; Bergstrom, J.; Ekelund, L.; Hultman, E. Muscle glycogen and muscle electrolytes during prolonged physical exercise. Acta Physiol. Scand. 1967, 70, 129–142. [Google Scholar] [CrossRef]
- Nicholas, C.W.; Green, P.A.; Hawkins, R.D.; Williams, C. Carbohydrate intake and recovery of intermittent running capacity. Int. J. Sport Nutr. 1997, 7, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Flynn, M.G.; Kirwan, J.P.; Houmard, J.A.; Mitchell, J.B.; Thomas, R.; Park, S.H. Effects of repeated days of intensified training on muscle glycogen and swimming performance. Med. Sci. Sports Exerc. 1988, 20, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Yeo, W.K.; Paton, C.D.; Garnham, A.P.; Burke, L.M.; Carey, A.L.; Hawley, J.A. Skeletal muscle adaptation and performance responses to once a day versus twice every second day endurance training regimens. J. Appl. Physiol. 2008, 105, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Fallowfield, J.L.; Williams, C.; Singh, R. The influence of ingesting a carbohydrate-electrolyte beverage during 4 hours of recovery on subsequent endurance capacity. Int. J. Sport Nutr. 1995, 5, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Bilzon, J.L.; Allsopp, A.J.; Williams, C. Short-term recovery from prolonged constant pace running in a warm environment: The effectiveness of a carbohydrate-electrolyte solution. Eur. J. Appl. Physiol. 2000, 82, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.W.; Nio, A.Q.X.; Ang, W.H.; Law, L.Y.L.; Lim, C.L. Effects of ingesting a sports drink during exercise and recovery on subsequent endurance capacity. Eur. J. Sport Sci. 2011, 11, 77–86. [Google Scholar] [CrossRef]
- Taylor, C.; Higham, D.; Close, G.L.; Morton, J.P. The effect of adding caffeine to postexercise carbohydrate feeding on subsequent high-intensity interval-running capacity compared with carbohydrate alone. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Nybo, L. CNS fatigue provoked by prolonged exercise in the heat. Front. Biosci. 2010, 2, 779–792. [Google Scholar] [CrossRef]
- Wong, S.H.; Williams, C.; Adams, N. Effects of ingesting a large volume of carbohydrate-electrolyte solution on rehydration during recovery and subsequent exercise capacity. Int. J. Sport Nutr. Exerc. Metab. 2000, 10, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Fallowfield, J.L.; Williams, C. The influence of a high carbohydrate intake during recovery from prolonged, constant-pace running. Int. J. Sport Nutr. 1997, 7, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Siu, P.M.; Wong, S.H.; Morris, J.G.; Lam, C.W.; Chung, P.K.; Chung, S. Effect of frequency of carbohydrate feedings on recovery and subsequent endurance run. Med. Sci. Sports Exerc. 2004, 36, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.H.; Williams, C. Influence of different amounts of carbohydrate on endurance running capacity following short term recovery. Int. J. Sports Med. 2000, 21, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Schabort, E.J.; Hawley, J.A. Reliability of power in physical performance tests. Sports Med. 2001, 31, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Millard-Stafford, M.; Warren, G.L.; Thomas, L.M.; Doyle, J.A.; Snow, T.; Hitchcock, K. Recovery from run training: Efficacy of a carbohydrate-protein beverage? Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.A.; Stevenson, E.; Williams, C.; Sheppard, C.; Grey, E.; Griffin, J. Recovery of endurance running capacity: Effect of carbohydrate-protein mixtures. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 590–609. [Google Scholar] [CrossRef] [PubMed]
- Karp, J.R.; Johnston, J.D.; Tecklenburg, S.; Mickleborough, T.D.; Fly, A.D.; Stager, J.M. Chocolate milk as a post-exercise recovery aid. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.L.; Coburn, J.W.; Beam, W.C.; Brown, L.E. Effects of isocaloric carbohydrate vs. carbohydrate-protein supplements on cycling time to exhaustion. J. Strength Cond. Res. 2012, 26, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, D.J.; Lessard, S.J.; Coffey, V.G.; Churchley, E.G.; Wootton, A.M.; Ng, T.; Watt, M.J.; Hawley, J.A. High rates of muscle glycogen resynthesis after exhaustive exercise when carbohydrate is coingested with caffeine. J. Appl. Physiol. 2008, 105, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Berardi, J.M.; Noreen, E.E.; Lemon, P.W. Recovery from a cycling time trial is enhanced with carbohydrate-protein supplementation vs. isoenergetic carbohydrate supplementation. J. Int. Soc. Sports Nutr. 2008, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Wagenmakers, A.J. Muscle amino acid metabolism at rest and during exercise: Role in human physiology and metabolism. Exerc. Sport Sci. Rev. 1998, 26, 287–314. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; Gonzalez-Alonso, J.; Saltin, B. Dissociation between muscle tricarboxylic acid cycle pool size and aerobic energy provision during prolonged exercise in humans. J. Physiol. 2002, 545, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, R. Exercise, nutrition and the brain. Sports Med. 2014, 44, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, E.; Hassmen, P.; Ek, S.; Ekblom, B.; Newsholme, E.A. Influence of ingesting a solution of branched-chain amino acids on perceived exertion during exercise. Acta Physiol. Scand. 1997, 159, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Alghannam, A.F. Carbohydrate-protein ingestion improves subsequent running capacity towards the end of a football-specific intermittent exercise. Appl. Physiol. Nutr. Metab. 2011, 36, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, G.; Raaymakers, J.S.; Saris, W.H.; Wagenmakers, A.J. Ingestion of branched-chain amino acids and tryptophan during sustained exercise in man: Failure to affect performance. J. Physiol. 1995, 486, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.; MacLean, D.A.; Kiens, B.; Christensen, D. Effects of glucose, glucose plus branched-chain amino acids, or placebo on bike performance over 100 km. J. Appl. Physiol. 1996, 81, 2644–2650. [Google Scholar] [CrossRef] [PubMed]
- Morifuji, M.; Kanda, A.; Koga, J.; Kawanaka, K.; Higuchi, M. Preexercise ingestion of carbohydrate plus whey protein hydrolysates attenuates skeletal muscle glycogen depletion during exercise in rats. Nutrition 2011, 27, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Marmy-Conus, N.; Fabris, S.; Proietto, J.; Hargreaves, M. Preexercise glucose ingestion and glucose kinetics during exercise. J. Appl. Physiol. 1996, 81, 853–857. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Study | Carbohydrate Intake (g·kg BM−1·h−1) | Recovery Time (h) | Timing of Ingestion (min) | Mode of Exercise | Repeated Exercise | Repeated Exercise Bout (min) |
|---|---|---|---|---|---|---|
| Fallowfield et al. [146] | 0, 0.5 | 4 | Immediately post-exercise, 120 | Running | Time to exhaustion at 70% VO2max | 40, 62 * |
| Fallowfield & Williams [152] | 0.5, 1.5 | 4 | Immediately post-exercise, 120 | Running | Time to exhaustion at 70% VO2max | 59, 58 |
| Wong & Williams [154] | 0.15, 0.53 | 4 | 30, 60, 90, 120, 150 | Running | Time to exhaustion at 70% VO2max | 65, 57 |
| Wong et al. [151] | 0, 0.9 | 4 | 30, 60, 90, 120, 150, 180 | Running | Time to exhaustion at 70% VO2max | 45, 69 * |
| Bilzon et al. [147] | 0, 0.43 | 4 | Immediately post-exercise, 60, 120, 180 | Running | Time to exhaustion at 60% VO2max | 45, 61 * |
| Casey et al. [25] | 0, 0.25 G, 0.25 S | 4 | Immediately post-exercise | Cycling | Time to exhaustion at 70% VO2max | 35, 40 G, 46 S |
| Betts et al. [23] | 0.8, 1.1 | 4 | Immediately post-exercise, 30, 60, 90, 120, 150, 180, 210 | Running | Time to exhaustion at 70% VO2max | 84, 100 * |
| Alghannam et al. [21] | 0.3, 1.2 | 4 | Immediately post-exercise, 30, 60, 90, 120, 150, 180, 210 | Running | Time to exhaustion at 70% VO2max | 48, 80 * |
| Study | Carbohydrate Intake (g·kg BM−1·h−1) | Protein Intake (g·kg BM−1·h−1) | Recovery Time (h) | Timing of Ingestion (min) | Mode of Exercise | Repeated Exercise | Repeated Exercise Bout (min) |
|---|---|---|---|---|---|---|---|
| Williams et al. [126] | 0.15, 0.4 | 0, 0.1 | 4 | Immediately post-exercise, 120 | Running | Time to exhaustion at 85% VO2max | 20, 31 * |
| Millard-Stafford et al. [156] | 0.6, 1.0, 0.8 | 0, 0, 0.2 | 2 | Immediately post-exercise, 60 | Running | Time to exhaustion at 90% VO2max | 6, 6, 5 |
| Betts et al. [157] | 0.8, 0.8, 1.2, 1.2 | 0, 0.1, 0, 0.2 | 4 | Immediately post-exercise, 30, 60, 90, 120, 150, 180, 210 | Running | Time to exhaustion at 85% VO2max | 18, 20, 15, 18 |
| Karp et al. [158] | 0.2, 0.5, 0.5 † | 0, 0.13, 0.13 † | 4 | Immediately post-exercise, 120 | Cycling | Time to exhaustion at 70% VO2max | 41 *, 29, 40 †,* |
| Betts et al. [23] | 0.8, 1.1, 0.8 | 0, 0, 0.3 | 4 | Immediately post-exercise, 30, 60, 90, 120, 150, 180, 210 | Running | Time to exhaustion at 70% VO2max | 84, 91 *, 100 * |
| Thomas et al. [41] | 0.2, 0.5, 0.4 † | 0, 0.13, 0.10 † | 4 | Immediately post-exercise, 120 | Cycling | Time to exhaustion at 70% VO2max | 23, 21, 32 †,* |
| Lunn et al. [40] | 0.25, 0.20 † | 0, 0.05 † | 3 | Immediately post-exercise | Running | Time to exhaustion at incline achieved at VO2peak test | 3, 4 †,* |
| Richardson et al. [159] | 1.5, 1.2 | 0, 0.3 | 3 | Immediately post-exercise, 30, 60, 90, 120 | Cycling | Time to exhaustion at 75% VO2max | 25, 24 |
| Alghannam et al. [22] | 1.2,0.8 | 0, 0.4 | 4 | Immediately post-exercise, 30, 60, 90, 120, 150, 180, 210 | Running | Time to exhaustion at 70% VO2max | 51, 49 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alghannam, A.F.; Gonzalez, J.T.; Betts, J.A. Restoration of Muscle Glycogen and Functional Capacity: Role of Post-Exercise Carbohydrate and Protein Co-Ingestion. Nutrients 2018, 10, 253. https://doi.org/10.3390/nu10020253
Alghannam AF, Gonzalez JT, Betts JA. Restoration of Muscle Glycogen and Functional Capacity: Role of Post-Exercise Carbohydrate and Protein Co-Ingestion. Nutrients. 2018; 10(2):253. https://doi.org/10.3390/nu10020253
Chicago/Turabian StyleAlghannam, Abdullah F., Javier T. Gonzalez, and James A. Betts. 2018. "Restoration of Muscle Glycogen and Functional Capacity: Role of Post-Exercise Carbohydrate and Protein Co-Ingestion" Nutrients 10, no. 2: 253. https://doi.org/10.3390/nu10020253
APA StyleAlghannam, A. F., Gonzalez, J. T., & Betts, J. A. (2018). Restoration of Muscle Glycogen and Functional Capacity: Role of Post-Exercise Carbohydrate and Protein Co-Ingestion. Nutrients, 10(2), 253. https://doi.org/10.3390/nu10020253