Plant-Based Beverages as Good Sources of Free and Glycosidic Plant Sterols


Abstract
:
1. Introduction
2. Materials and Methods
3. Results
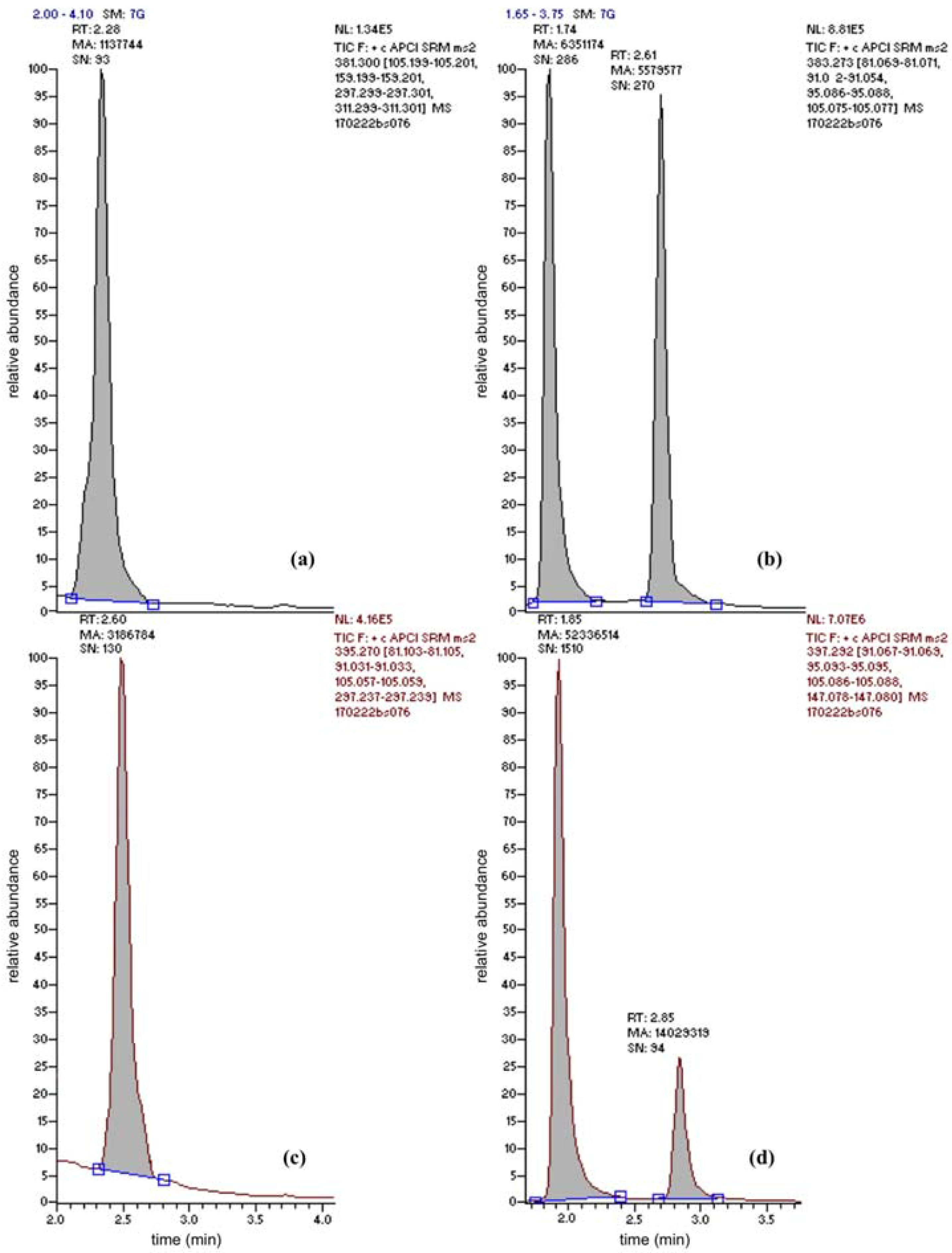
3.1. Data Analysis and Quality Assurance of the Analytical Method: Limits of Detection and Quantification
3.2. Quantification of Cholesterol-Lowering (Conjugated) Plant Sterols and Ergosterol (Provitamin D2) with UPLC-MS/MS
4. Discussion
4.1. Fruit Juices
4.2. Vegetable Juices
4.3. Plant-Based Milk Alternatives
4.4. Gel, Sodas, Teas and Non-Alcoholic Malt-Based Drinks
4.5. Beers
4.6. Other Water-Soluble Glucose-Conjugated Plant Sterols
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bradford, P.G.; Awad, A.B. Phytosterols as anticancer compounds. Mol. Nutr. Food Res. 2007, 51, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Ramprasath, V.R.; Awad, A.B. Role of Phytosterols in Cancer Prevention and Treatment. J. AOAC Int. 2015, 98, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C.; Cont, M.; Camborata, C.; Spinozzi, S.; Montagnani, M.; et al. Antiinflammatory effect of phytosterols in experimental murine colitis model: Prevention, induction, remission study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef] [PubMed]
- Ling, W.H.; Jones, P.J. Dietary phytosterols: A review of metabolism, benefits and side effects. Life Sci. 1995, 57, 195–206. [Google Scholar] [CrossRef]
- Wang, T.; Hicks, K.B.; Moreau, R. Antioxidant activity of phytosterols, oryzanol, and other phytosterol conjugates. J. Am. Oil Chem. Soc. 2002, 79, 1201–1206. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Ramprasath, V.R.; Jones, P.J. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Phillips, K.M.; Ruggio, D.M.; Horst, R.L.; Minor, B.; Simon, R.R.; Feeney, M.J.; Byrdwell, W.C.; Haytowitz, D.B. Vitamin D and Sterol Composition of 10 Types of Mushrooms from Retail Suppliers in the United States. J. Agric. Food Chem. 2011, 59, 7841–7853. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.L. Vitamin D: Two-Volume Set. In Industrial Aspects of Vitamin D; Feldman, D., Pike, J.W., Adamn, J.S., Eds.; Academic Press: Cambridge, MA, USA, 2011; Chapter 6; pp. 73–93. [Google Scholar]
- Brufau, G.; Canela, M.A.; Rafecas, M. Phytosterols: Physiologic and metabolic aspects related to cholesterol-lowering properties. Nutr. Res. 2008, 28, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E.A.; Duchateau, G.S.M.J.E.; Lin, Y.; Ntanios, F.Y. Proposed mechanisms of cholesterol-lowering action of plant sterols. Eur. J. Lipid Sci. Technol. 2003, 105, 171–185. [Google Scholar] [CrossRef]
- Higdon, J.; Drake, V.; Higdon, J. An Evidence-Based Approach to Phytochemicals and Other Dietary Factors, 2nd ed.; Thieme: Stuttgart, Germany, 2013; p. 304. [Google Scholar]
- Kritchevsky, D.; Chen, S.C. Phytosterols—Health benefits and potential concerns: A review. Nutr. Res. 2005, 25, 413–428. [Google Scholar] [CrossRef]
- Abumweis, S.S.; Barake, R.; Jones, P.J. Plant sterols/stanols as cholesterol lowering agents: A meta-analysis of randomized controlled trials. Food Nutr. Res. 2008, 52. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Cabral, J.M.S. Phytosterols: Applications and recovery methods. Bioresour. Technol. 2007, 98, 2335–2350. [Google Scholar] [CrossRef] [PubMed]
- Quilez, J.; Garcia-Lorda, P.; Salas-Salvado, J. Potential uses and benefits of phytosterols in diet: Present situation and future directions. Clin. Nutr. 2003, 22, 343–351. [Google Scholar] [CrossRef]
- Laitinen, K.; Gylling, H.; Kaipiainen, L.; Nissinen, M.J.; Simonen, P. Cholesterol lowering efficacy of plant stanol ester in a new type of product matrix, a chewable dietary supplement. J. Funct. Foods 2017, 30, 119–124. [Google Scholar] [CrossRef]
- Lin, X.B.; Ma, L.; Racette, S.B.; Spearie, C.L.A.; Ostlund, R.E., Jr. Phytosterol glycosides reduce cholesterol absorption in humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G931–G935. [Google Scholar] [CrossRef] [PubMed]
- Racette, S.B.; Spearie, C.A.; Phillips, K.M.; Lin, X.; Ma, L.; Ostlund, R.E., Jr. Phytosterol-Deficient and High-Phytosterol Diets Developed for Controlled Feeding Studies. J. Am. Diet. Assoc. 2009, 109, 2043–2051. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.M.; Ruggio, D.M.; Ashraf-Khorassani, M. Phytosterol composition of nuts and seeds commonly consumed in the United States. J. Agric. Food Chem. 2005, 53, 9436–9445. [Google Scholar] [CrossRef] [PubMed]
- Muller, R.; Walker, S.; Brauer, J.; Junquera, M. Does beer contain compounds that might interfere with cholesterol metabolism? J. Inst. Brew. 2007, 113, 102–109. [Google Scholar] [CrossRef]
- Decloedt, A.I.; van Landschoot, A.; Vanhaecke, L. Fractional factorial design-based optimisation and application of an extraction and UPLC-MS/MS detection method for the quantification of phytosterols in food, feed and beverages low in phytosterols. Anal. Bioanal. Chem. 2016, 408, 7731–7744. [Google Scholar] [CrossRef] [PubMed]
- Nes, W.R.; Nes, W.D. Lipids in Evolution, 1st ed.; Plenum Press: New York, NY, USA; London, UK, 1980; p. 243. [Google Scholar]
- Knapp, F.F.; Nicholas, H.J. Sterols and Triterpenes of Banana Pulp. J. Food Sci. 1969, 34, 584. [Google Scholar] [CrossRef]
- Knapp, F.F.; Nicholas, H.J. Distribution of Sterols and Steryl Esters in Banana Plant. Phytochemistry 1969, 8, 2091. [Google Scholar] [CrossRef]
- Ramu, R.; Shirahatti, P.S.; Zameer, F.; Prasad, N. Investigation of antihyperglycaemic activity of banana (Musa sp. var. Nanjangud rasa bale) pseudostem in normal and diabetic rats. J. Sci. Food Agric. 2015, 95, 165–173. [Google Scholar] [PubMed]
- Abidi, S.L. Chromatographic analysis of plant sterols in foods and vegetable oils. J. Chromatogr. A 2001, 935, 173–201. [Google Scholar] [CrossRef]
- Toivo, J.; Phillips, K.; Lampi, A.-M.; Piironen, V. Determination of sterols in foods: Recovery of free, esterified, and glycosidic sterols. J. Food Compos. Anal. 2001, 14, 631–643. [Google Scholar] [CrossRef]
- Piironen, V.; Toivo, J.; Puupponen-Pimiä, R.; Lampi, A.M. Plant sterols in vegetables, fruits and berries. J. Sci. Food Agric. 2003, 83, 330–337. [Google Scholar] [CrossRef]
- Normen, L.; Johnsson, M.; Andersson, H.; Van Gameren, Y.; Dutta, P. Plant sterols in vegetables and fruits commonly consumed in Sweden. Eur. J. Nutr. 1999, 38, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. Effect of processing techniques at industrial scale on orange juice antioxidant and beneficial health compounds. J. Agric. Food Chem. 2002, 50, 5107–5114. [Google Scholar] [CrossRef] [PubMed]
- Spanos, G.A.; Wrolstad, R.E.; Heatherbell, D.A. Influence of Processing and Storage on the Phenolic Composition of Apple Juice. J. Agric. Food Chem. 1990, 38, 1572–1579. [Google Scholar] [CrossRef]
- Demir, N.; Bahceci, K.S.; Acar, J. The effect of processing method on the characteristics of carrot juice. J. Food Qual. 2007, 30, 813–822. [Google Scholar] [CrossRef]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The potential benefits of red beetroot supplementation in health and disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef] [PubMed]
- Parsi, Z.; Gorecki, T. Determination of ergosterol as an indicator of fungal biomass in various samples using non-discriminating flash pyrolysis. J. Chromatogr. A 2006, 1130, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Stuper-Szablewska, K.; Perkowski, J. Contamination of wheat grain with microscopic fungi and their metabolites in Poland in 2006-2009. Ann. Agric. Environ. Med. 2014, 21, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Arnezeder, C.; Koliander, W.; Hampel, W.A. Rapid-Determination of Ergosterol in Yeast-Cells. Anal. Chim. Acta 1989, 225, 129–136. [Google Scholar] [CrossRef]
- Pasanen, A.L.; Yli-Pietilä, K.; Pasanen, P.; Kalliokoski, P.; Tarhanen, J. Ergosterol content in various fungal species and biocontaminated building materials. Appl. Environ. Microbiol. 1999, 65, 138–142. [Google Scholar] [PubMed]
- Yamaya, A.; Endo, Y.; Fujimoto, K.; Kitamura, K. Effects of genetic variability and planting location on the phytosterols content and composition in soybean seeds. Food Chem. 2007, 102, 1071–1075. [Google Scholar] [CrossRef]
- Mo, S.; Dong, L.; Hurst, W.J.; Van Breemen, R.B. Quantitative analysis of phytosterols in edible oils using APCI liquid chromatography-tandem mass spectrometry. Lipids 2013, 48, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Diaz, T.G.; Merás, I.D.; Casas, J.S.; Franco, M.A. Characterization of virgin olive oils according to its triglycerides and sterols composition by chemometric methods. Food Control 2005, 16, 339–347. [Google Scholar] [CrossRef]
- Vrbkova, B.; Roblová, V.; Yeung, E.S.; Preisler, J. Determination of sterols using liquid chromatography with off-line surface-assisted laser desorption/ionization mass spectrometry. J. Chromatogr. A 2014, 1358, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Flakelar, C.L.; Prenzler, P.D.; Luckett, D.J.; Howitt, J.A.; Doran, G. A rapid method for the simultaneous quantification of the major tocopherols, carotenoids, free and esterified sterols in canola (Brassica napus) oil using normal phase liquid chromatography. Food Chem. 2017, 214, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Rapota, M.O.; Tyrsin, Y.A. Extraction of lipids from the raw materials for beer production and development of methods from phytosterols’ determination by high performance liquid chromatography (HPLC). Biol. Med. 2015, 7, 5. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
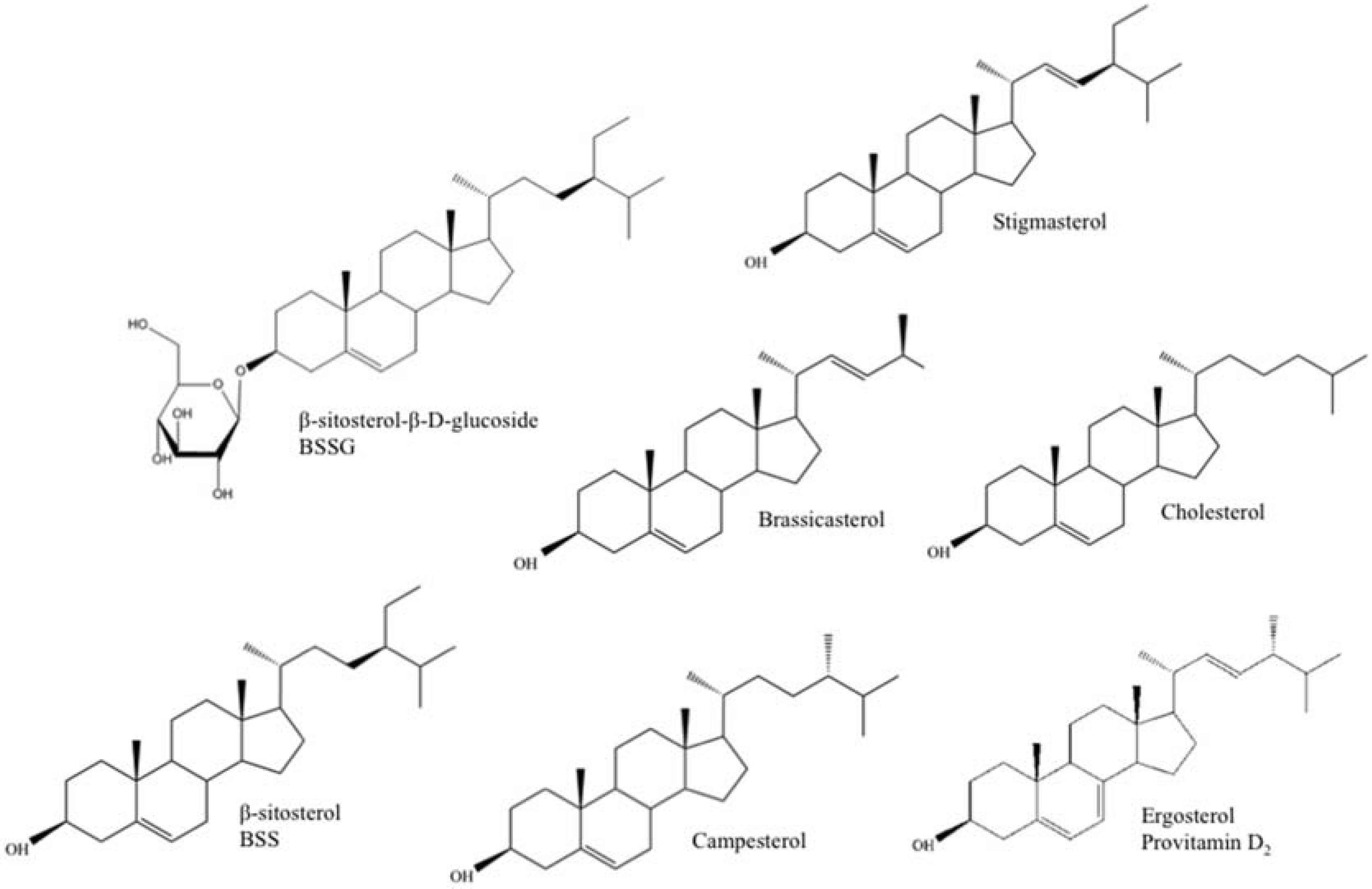
| Sterol Name | Mean Mass (Da) | Molecular Formula | Structural Characteristics | ||
|---|---|---|---|---|---|
| Double Bounds | Position Double Bounds | Alkyl Group at C24 | |||
| Cholesterol | 386.654 | C27H46O | 1 | C5–C6 | / |
| Ergosterol (Provitamin D2) | 396.648 | C28H44O | 3 | C5–C6 C7–C8 C22–C23 | Methyl |
| β-sitosterol-β-d-glucoside (BSSG) | 576.847 | C35H60O6 | 1 | C5–C6 | Ethyl |
| β-sitosterol (BSS) | 414.707 | C29H50O | 1 | C5–C6 | Ethyl |
| Brassicasterol | 398.675 | C28H46O | 2 | C5–C6 C22–C23 | Methyl |
| Stigmasterol | 412.691 | C29H48O | 2 | C5–C6 C22–C23 | Ethyl |
| Campesterol | 400.691 | C28H48O | 1 | C5–C6 | Methyl |
| 1. | 5 mL (diluted) ∆ beverage in a 50 mL tube |
| 2. | Addition of cholesterol (100 µL, 10 ng or 50 ng per µL) |
| 3. | (Calibration samples: fortified with different plant sterols) |
| 4. | Liquid-liquid extraction with 8 mL chloroform:methanol (2:1) |
| 5. | Vortex (30 s) + ultrasonication * (10 min) |
| 6. | Centrifugation (4400× g, 10 min) |
| 7. | Cottonwool filter |
| 8. | Second liquid-liquid extraction (repeat step 2 to 5) |
| 9. | 1 mL fresh chloroform:methanol added to the filter (filter wash out) |
| 10. | Transfer 2 mL extract to new tube |
| 11. | 15–20 min drying (under liquid N2, 46 °C) |
| 12. | 180 μL methanol (vortex 30 s, ultrasonication * 10 min) |
| 13. | 20 μL ultrapure H2O (vortex 30 s, ultrasonication * 3 min, vortex 30 s) |
| 14. | Centrifugation (12,300× g, 10 min) |
| 15. | Transfer 150 µL to plastic LC-MS vial with insert for analysis |
| Analyte | Precursor Ion | Product Ions | Mean Relative Ion Abundancy * | Retention Time (Relative) | S-Lens Voltage | Collision Energy |
|---|---|---|---|---|---|---|
| (m/z) | (m/z) | (%) | (min) | (V) | (eV) | |
| β-Sitosterol-β-d-glucoside BSSG | 397.3 | 91 | 70 | 1.80 (0.77) | 88 | 47 |
| 95 | 73 | 35 | ||||
| 105 | 100 | 40 | ||||
| 147 | 93 | 28 | ||||
| Ergosterol Provitamine D2 | 379.3 | 69 | 78 | 2.04 (0.87) | 120 | 23 |
| 91 | 100 | 53 | ||||
| 105 | 90 | 34 | ||||
| 15 | 82 | 24 | ||||
| Brassicasterol | 381.3 | 105 | 100 | 2.27 (0.97) | 82 | 43 |
| 159 | 67 | 23 | ||||
| 297 | 93 | 14 | ||||
| 311 | 40 | 13 | ||||
| Cholesterol internal standard | 369.3 | 91 | 83 | 2.35 (1.00) | 84 | 52 |
| 95 | 69 | 34 | ||||
| 105 | 100 | 40 | ||||
| Campesterol | 383.3 | 81 | 67 | 2.61 (1.11) | 86 | 35 |
| 91 | 85 | 49 | ||||
| 95 | 74 | 34 | ||||
| 105 | 100 | 43 | ||||
| Stigmasterol | 395.3 | 81 | 64 | 2.63 (1.12) | 59 | 37 |
| 91 | 91 | 52 | ||||
| 105 | 100 | 44 | ||||
| 297 | 90 | 18 | ||||
| β-sitosterol BSS | 397.3 | 91 | 70 | 2.90 (1.23) | 88 | 47 |
| 95 | 73 | 35 | ||||
| 105 | 100 | 40 | ||||
| 147 | 93 | 28 |
| Category | Product Name | mg per 100 mL | µg per 100 mL | |||||
|---|---|---|---|---|---|---|---|---|
| BSS | BSSG | Brassicasterol | Campesterol | Stigmasterol | Ergosterol | |||
| Fruit juices | Apple juice | 0.21 ± 0.01 | 4.1 ± 1.2 * | NF (<0.75) | 27 ± 3 | 2.6 ± 0.4 | ND | |
| Orange juice | 0.42 ± 0.09 | 8.3 ± 2.3 * | NF (<1.5) | 71 ± 12 | 23 ± 3 | ND | ||
| Pomegranate juice | 2.1 ± 0.3 | 32 ± 7 | NF (<3) | 139 ± 18 | <30 | 17 ± 6 | ||
| Multifruit-carrot juice | 2.5 ± 0.2 | 16 ± 3 | NF (<3) | 607 ± 12 | 224 ± 10 | ND | ||
| Fresh orange-banana juice | 5.3 ± 2.2 | >90 | NF (<3) | 846 ± 93 | 610 ± 35 | NF (<3) | ||
| Vegetable juices | Tomato juice | 0.36 ± 0.02 | 4.4 ± 0.5 | NF (<2) | 155 ± 10 | 331 ± 27 | ND | |
| Mixed vegetable juice (a) | 0.74 ± 0.05 | 6.2 ± 0.5 | NF (<3) | 242 ± 24 | 596 ± 64 | ND | ||
| Mixed vegetable juice (b) | 0.72 ± 0.10 | 12 ± 3 | NF (<3.75) | 177 ± 25 | 359 ± 72 | 29 ± 4 | ||
| Beetroot juice | 0.42 ± 0.07 | 7.3 ± 1.2 | NF (<3) | 47 ± 8 | 40 ± 3 | NF (<3) | ||
| Carrot juice | 2.7 ± 0.4 | 18 ± 4 | NF (<3) | 677 ± 68 | 1270 ± 65 | NF (<3) | ||
| Plant-based milk alternatives | Coconut-rice | 0.51 ± 0.07 | 2.8 ± 0.9 | NF (<3) | 72 ± 10 | 76 ± 13 | ND | |
| Rice | 1.4 ± 0.1 | 2.4 ± 0.6 | 10 ± 3 | 260 ± 28 | 234 ± 23 | ND | ||
| Soy | 2.5 ± 0.5 | 4.9 ± 2.1 | 4.6 ± 0.4 | 1290 ± 291 | 998 ± 111 | ND | ||
| Cashew | 2.7 ± 0.4 | >60 | NF (<3) | 279 ± 44 | 15 ± 1 | NF (<3) | ||
| Almond (a) unroasted | 2.6 ± 0.6 | 78 ± 14 | NF (<3) | 101 ± 30 | <30 | ND | ||
| Almond (b) roasted | 2.5 ± 0.1 | 13 ± 2 | NF (<2) | 62 ± 4 | 1915 ± 109 | ND | ||
| Oat (a) | 2.1 ± 0.2 | 26 ± 4 | NF (<3) | 475 ± 30 | 182 ± 16 | ND | ||
| Oat (b) | 3.9 ± 0.7 | 33 ± 4 | 217 ± 12 | 1098 ± 61 | <15 | NF (<3) | ||
| Gel | Aloe vera gel beverage | 0.22 ± 0.03 | 17 ± 5 | NF (<0.75) | 23 ± 5 | 2.2 ± 0.9 | ND | |
| Sodas | Lemonade (a) (orange) | 0.48 ± 0.04 | 1.5 ± 0.3* | NF (<0.75) | 73 ± 3 | 19 ± 2 | NF (<0.75) | |
| Biolemonade (a) (elderberry) | 0.19 ± 0.02 | 2.3 ± 0.7 | NF (<1.5) | 24 ± 2 | 52 ± 15 | NF (<0.75) | ||
| Biolemonade (b) (ginger-orange) | 0.17 ± 0.05 | 2.5 ± 0.7 | NF (<0.75) | 24 ± 3 | 4.3 ± 1.0 | NF (<0.75) | ||
| Soda with plant extract (a) | 0.05 ± 0.01 | 1.1 ± 0.2 | NF (<0.75) | 8.0 ± 1.7 | 1.3 ± 0.3 | NF (<0.75) | ||
| Soda with plant extract (b) (stevia) | 0.05 ± 0.01 | 0.60 ± 0.10 | NF (<0.75) | 7.3 ± 1.1 | 1.9 ± 0.3 | NF (<0.75) | ||
| Soda with plant extract (c) (peach) | 0.06 ± 0.01 | 1.5 ± 0.5 | NF (<0.75) | 5.8 ± 1.2 | 0.80 ± 0.31 | NF (<0.75) | ||
| Teas | Tea infusion (a) | 0.08 ± 0.01 | 0.67 ± 0.12 | NF (<0.75) | 9.2 ± 1.7 | 1.1 ± 0.3 | NF (<0.75) | |
| Tea infusion (b) | 0.06 ± 0.01 | 0.65 ± 0.17 | NF (<0.75) | 9.6 ± 1.3 | 1.5 ± 0.2 | NF (<0.75) | ||
| Tea infusion (c) | 0.06 ± 0.01 | 0.68 ± 0.12 | NF (<0.75) | 6.9 ± 0.8 | 2.3 ± 0.2 | NF (<0.75) | ||
| Iced tea (b) | 0.06 ± 0.01 | 0.93 ± 0.32 | NF (<0.75) | 8.3 ± 1.6 | 5.8 ± 0.8 | NF (<0.75) | ||
| Iced tea (c) | 0.05 ± 0.01 | 0.84 ± 0.08 | NF (<0.75) | 6.7 ± 0.8 | 9.5 ± 1.4 | NF (<0.75) | ||
 Indicates a plant-based beverage that can be considered a good source of free and conjugated plant sterols;
Indicates a plant-based beverage that can be considered a good source of free and conjugated plant sterols;  Indicates a plant-based beverage that contains only moderate (yellow) or low concentrations (orange) of plant sterols. NF, not found; ND, not determined.
Indicates a plant-based beverage that contains only moderate (yellow) or low concentrations (orange) of plant sterols. NF, not found; ND, not determined.| Category | Product Name | mg per 100 mL | µg per 100 mL | |||||
|---|---|---|---|---|---|---|---|---|
| BSS | BSSG | Brassicasterol | Campesterol | Stigmasterol | Ergosterol | |||
| Non- alcoholic malt drinks | Chinese malt drink | 0.07 ± 0.02 | 0.95 ± 0.33 | NF (<0.75) | 6.6 ± 2.1 | <LOQ (<2) | ND | |
| Icelandic malt drink | 0.14 ± 0.04 | 2.74 ± 1.31 | NF (<1.5) | 19 ± 4 | 2.4 ± 0.9 | ND | ||
| Non-alcoholic lager (a) | 0.04 ± 0.01 | 0.50 ± 0.07 | NF (<0.75) | 6.8 ± 2.4 | 1.0 ± 0.4 | NF (<0.75) | ||
| Non-alcoholic lager (b) | 0.07 ± 0.03 | 1.6 ± 0.5 | NF (<0.75) | 11 ± 3 | 1.9 ± 0.8 | ND | ||
| Non-alcoholic wheat beer (a) | 0.07 ± 0.02 | 1.0 ± 0.1 | NF (<0.75) | 7.7 ± 1.9 | 0.88 ± 0.35 | ND | ||
| Non-alcoholic wheat beer (b) | 0.12 ± 0.03 | 1.4 ± 0.2 * | NF (<0.75) | 14 ± 3 | 2.1 ± 0.4 | ND | ||
| Beers | Lager (a) | 0.20 ± 0.04 | 2.2 ± 0.3 | NF (<0.75) | 24 ± 5 | 3 ± 1 | NF (<0.75) | |
| Lager (b) | 0.26 ± 0.02 | 1.7 ± 0.3 * | NF (<0.75) | 31 ± 4 | 5 ± 1 | ND | ||
| Lager (c) | 0.25 ± 0.04 | 1.2 ± 0.3 * | NF (<0.75) | 39 ± 5 | 7 ± 1 | ND | ||
| Lager (d) | 0.23 ± 0.03 | 1.7 ± 0.3 * | NF (<0.75) | 23 ± 4 | 5 ± 1 | ND | ||
| Wheat beer (a) | 0.28 ± 0.04 | 2.8 ± 0.4 * | NF (<0.75) | 52 ± 3 | 6 ± 1 | 4.1 ± 0.4 | ||
| Wheat beer (b) | 0.38 ± 0.09 | 3.2 ± 0.6 * | NF (<0.75) | 53 ± 11 | 7 ± 2 | ND | ||
| Wheat beer (c) | 0.26 ± 0.02 | 1.9 ± 0.3 * | NF (<0.75) | 30 ± 2 | 3 ± 1 | ND | ||
| Wheat beer (d) | 0.27 ± 0.04 | 3.4 ± 0.4 * | NF (<0.75) | 37 ± 6 | 5 ± 1 | ND | ||
| Ale (bottle fermented) (a) | 0.37 ± 0.05 | 2.0 ± 0.3 * | NF (<0.75) | 49 ± 7 | 9 ± 1 | 0.80 ± 0.18 | ||
| Ale (bottle fermented) (b) | 0.23 ± 0.03 | 4.5 ± 0.5 * | NF (<0.75) | 36 ± 4 | 6 ± 1 | 6.0 ± 0.3 | ||
| Ale (bottle fermented) (c) | 0.25 ± 0.03 | 2.5 ± 0.5 * | NF (<0.75) | 36 ± 4 | 4 ± 1 | ND | ||
| Ale (bottle fermented) (d) | 0.23 ± 0.03 | 1.3 ± 0.1 * | NF (<0.75) | 7 ± 2 | 2 ± 1 | 4.6 ± 0.8 | ||
| Ale (bottle fermented) (e) | 0.09 ± 0.02 | 0.9 ± 0.1 * | NF (<0.75) | 36 ± 4 | 6 ± 1 | ND | ||
 Indicates a plant-based beverage that contains only moderate (yellow) or low concentrations (orange) of plant sterols. NF, not found; ND, not determined.
Indicates a plant-based beverage that contains only moderate (yellow) or low concentrations (orange) of plant sterols. NF, not found; ND, not determined.| Species | Ergosterol (mg per 100 g DW) |
|---|---|
| Yeast | |
| Cryptococcus albidus | 4200 ± 1200 |
| Rhodotorulamucilaginosa | 3700 ± 760 |
| Rhodotorulaminuta | 3700 ± 630 |
| Saccharomycescerevisiae | 400–2000 |
| Fungi | |
| Acremoniumfurcatum | 1400 ± 780 |
| Stachybotryschartarum | 1200 ± 520 |
| Aspergillus versicolor | 1100 ± 1500 |
| Penicilliumbrevicompactum | 580 ± 1300 |
| Cladosporiumcladosporioides | 560 ± 1100 |
| Aureobasidiumpullulans | 260 ± 1600 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Decloedt, A.I.; Van Landschoot, A.; Watson, H.; Vanderputten, D.; Vanhaecke, L. Plant-Based Beverages as Good Sources of Free and Glycosidic Plant Sterols. Nutrients 2018, 10, 21. https://doi.org/10.3390/nu10010021
Decloedt AI, Van Landschoot A, Watson H, Vanderputten D, Vanhaecke L. Plant-Based Beverages as Good Sources of Free and Glycosidic Plant Sterols. Nutrients. 2018; 10(1):21. https://doi.org/10.3390/nu10010021
Chicago/Turabian StyleDecloedt, Anneleen I, Anita Van Landschoot, Hellen Watson, Dana Vanderputten, and Lynn Vanhaecke. 2018. "Plant-Based Beverages as Good Sources of Free and Glycosidic Plant Sterols" Nutrients 10, no. 1: 21. https://doi.org/10.3390/nu10010021
APA StyleDecloedt, A. I., Van Landschoot, A., Watson, H., Vanderputten, D., & Vanhaecke, L. (2018). Plant-Based Beverages as Good Sources of Free and Glycosidic Plant Sterols. Nutrients, 10(1), 21. https://doi.org/10.3390/nu10010021