Spatio-Temporal LAI Modelling by Integrating Climate and MODIS LAI Data in a Mesoscale Catchment


Abstract
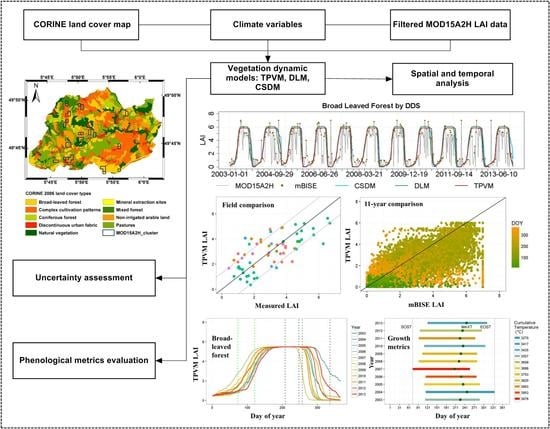
:
1. Introduction
2. Study Area and Data Sources
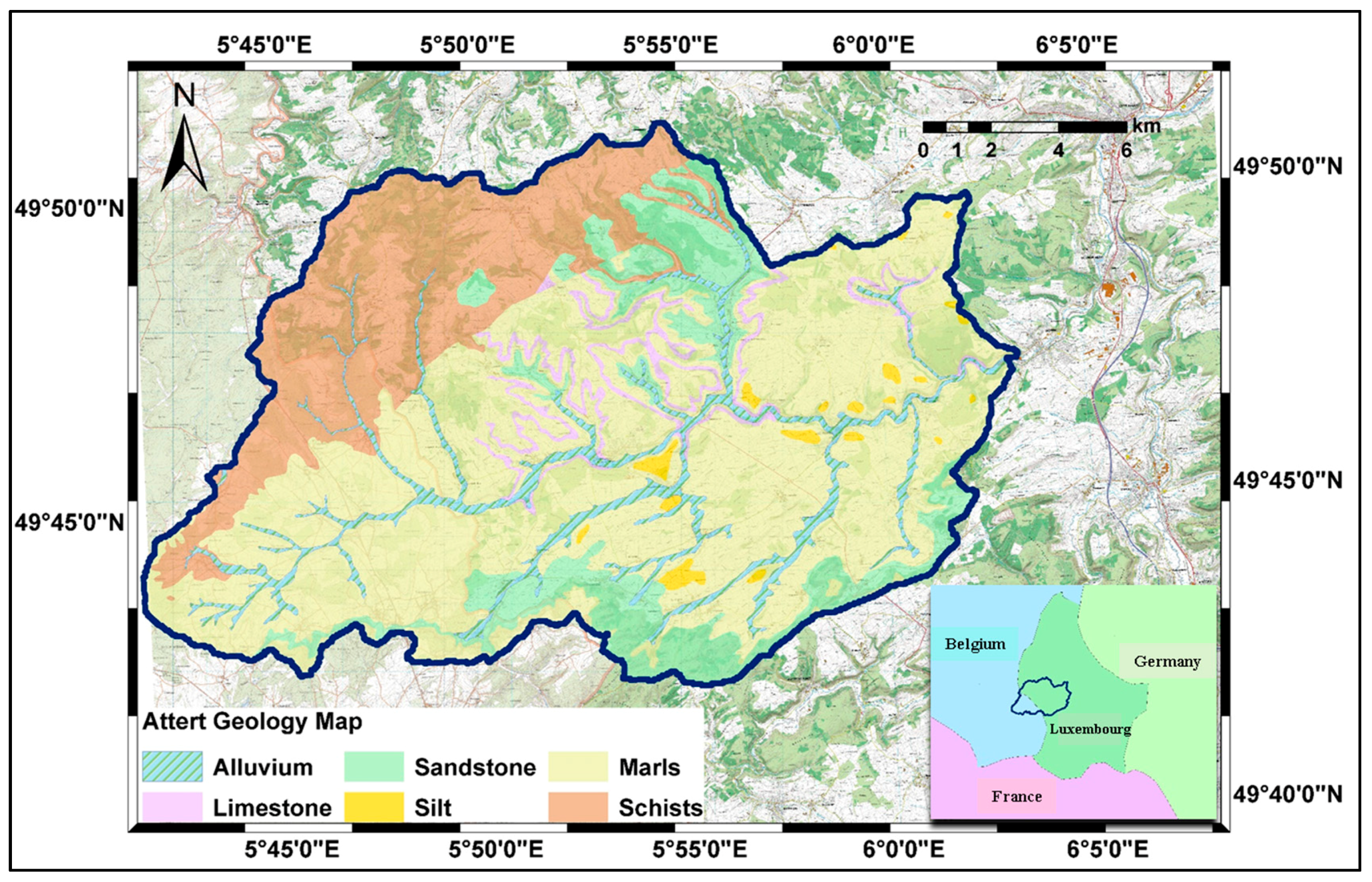
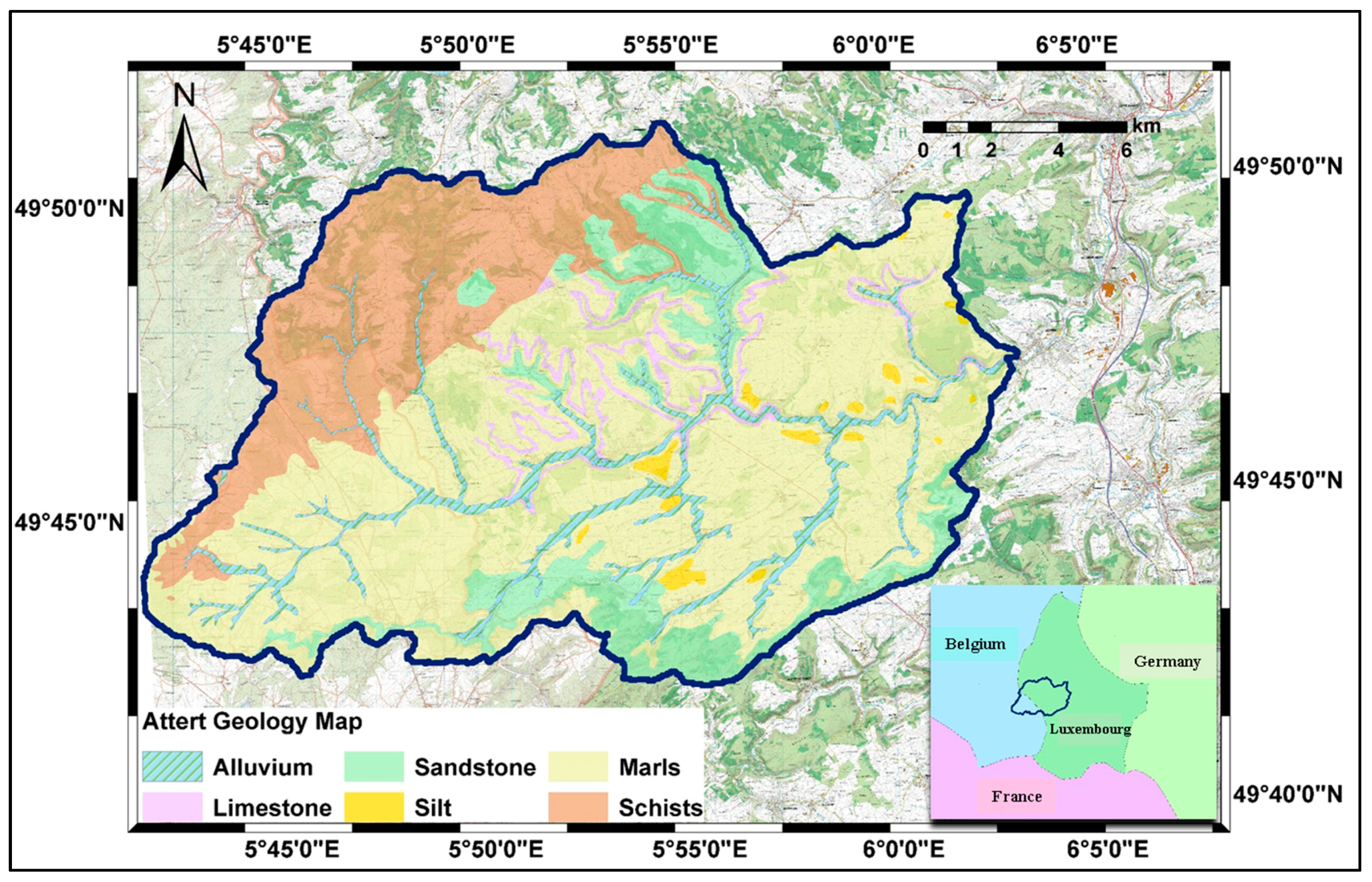
2.1. Study Area
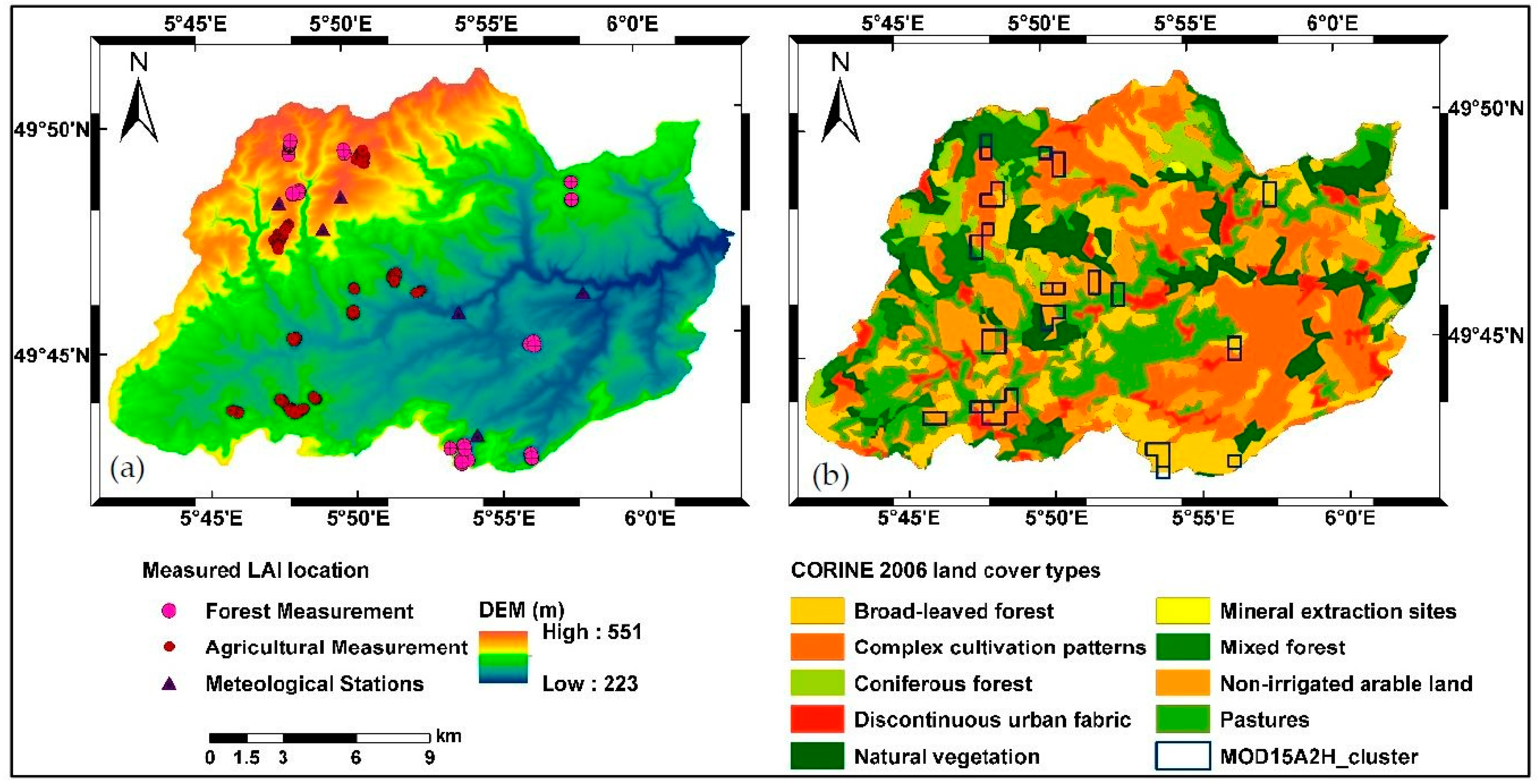
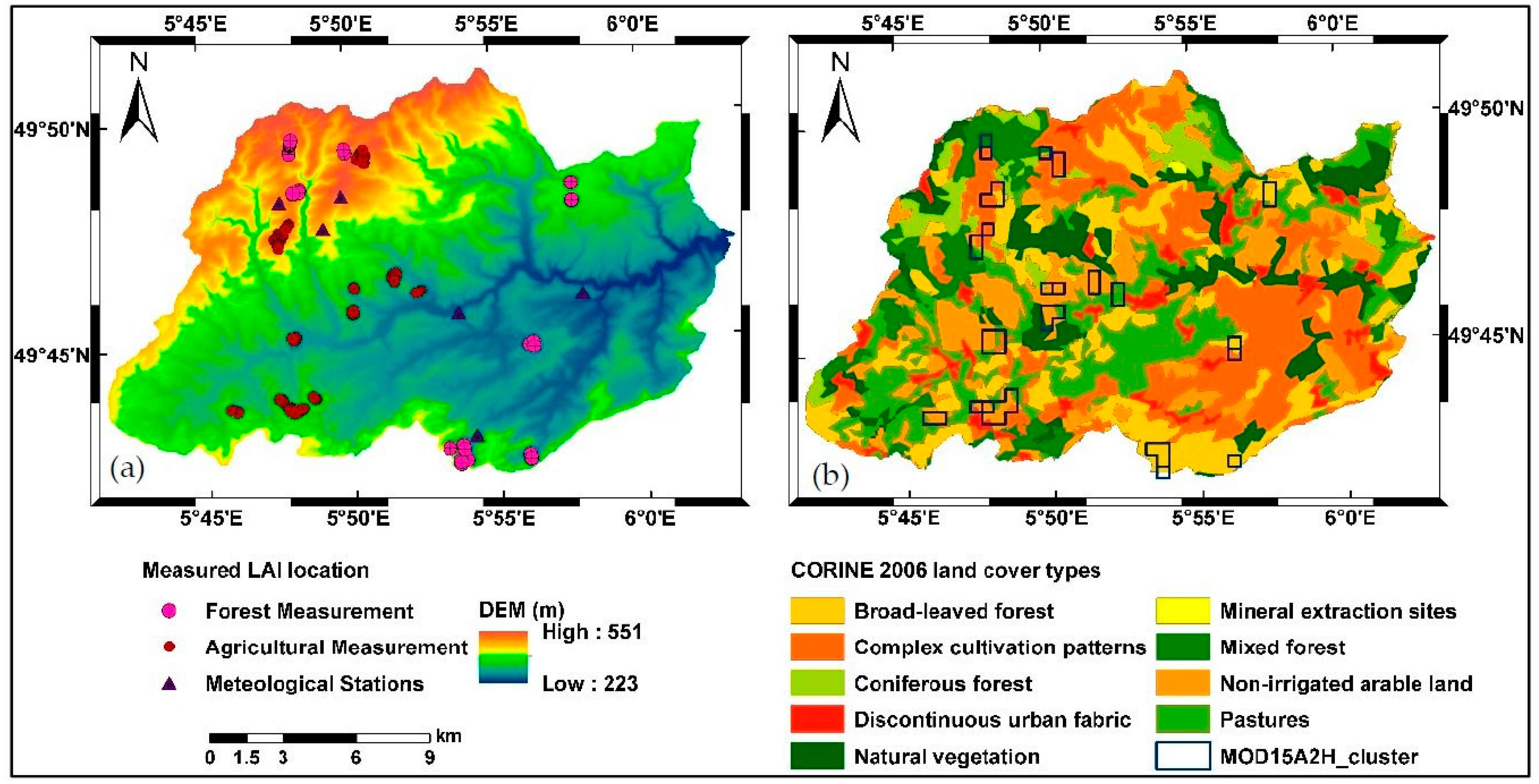
2.2. Data Sources
2.2.1. MODIS LAI Product
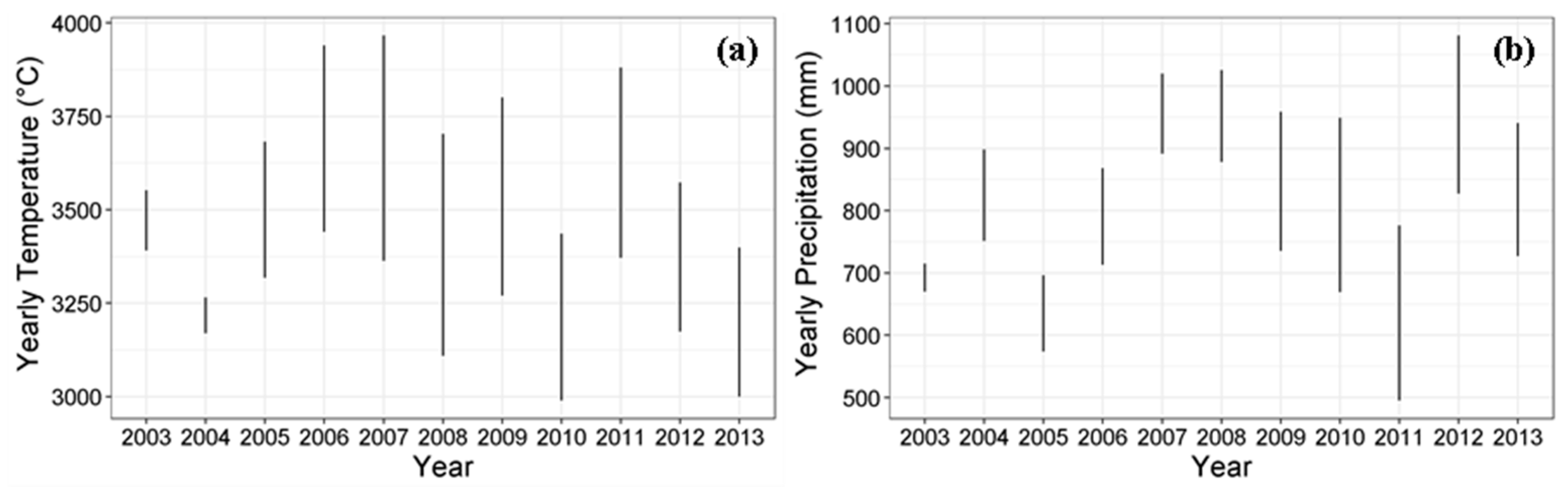
2.2.2. Climate Data
2.2.3. Land Cover Maps
2.2.4. Field LAI Measurement
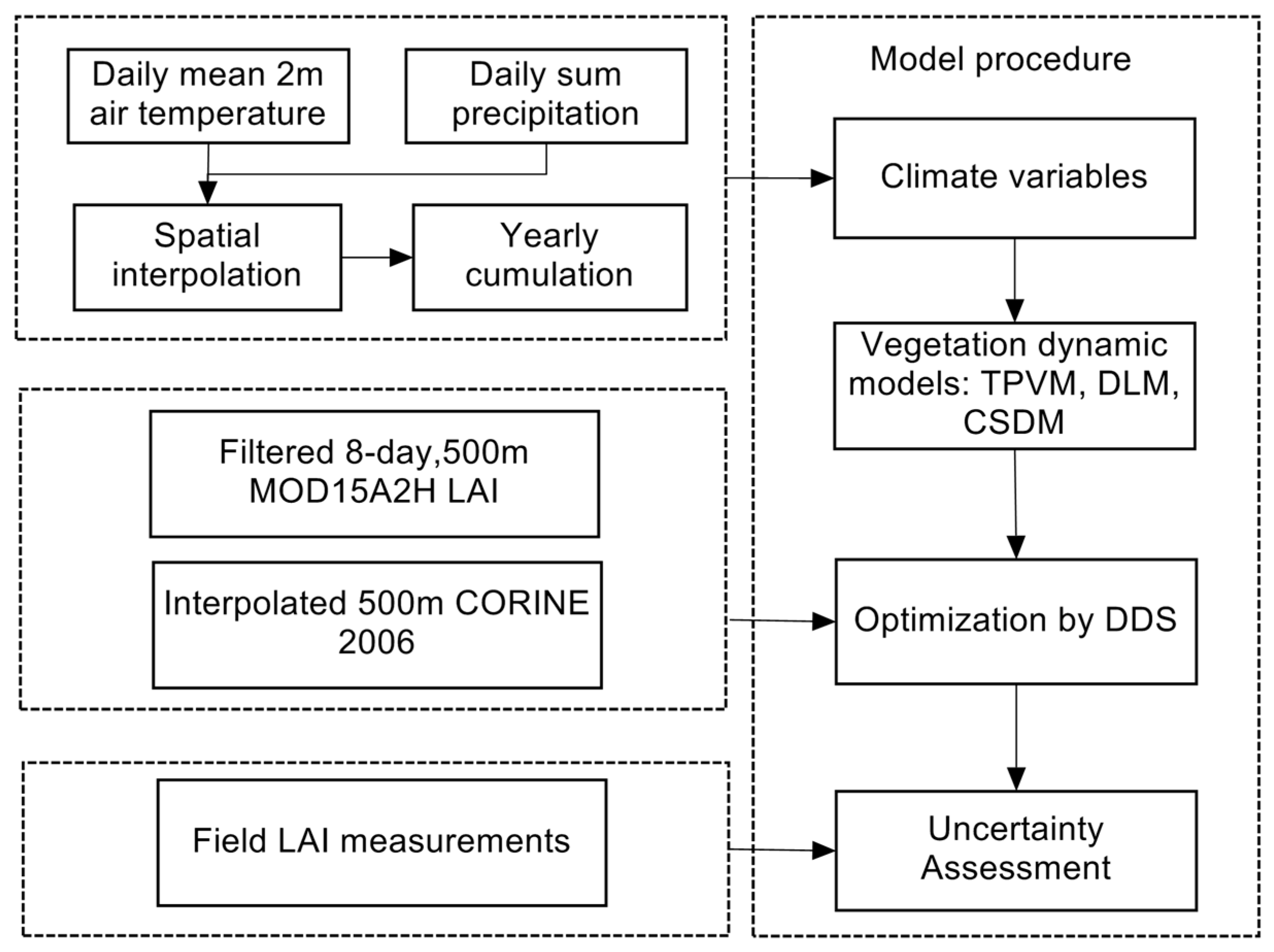
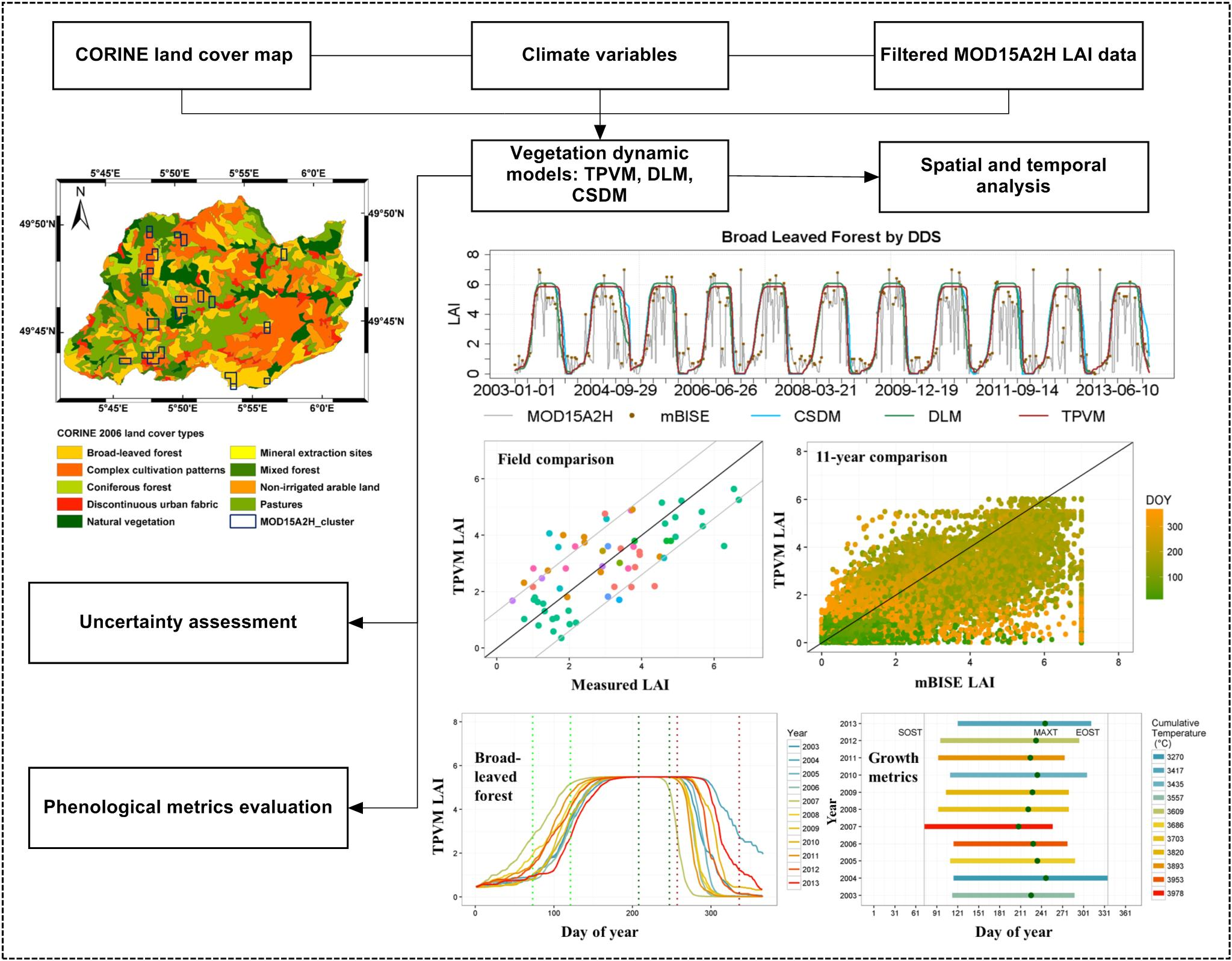
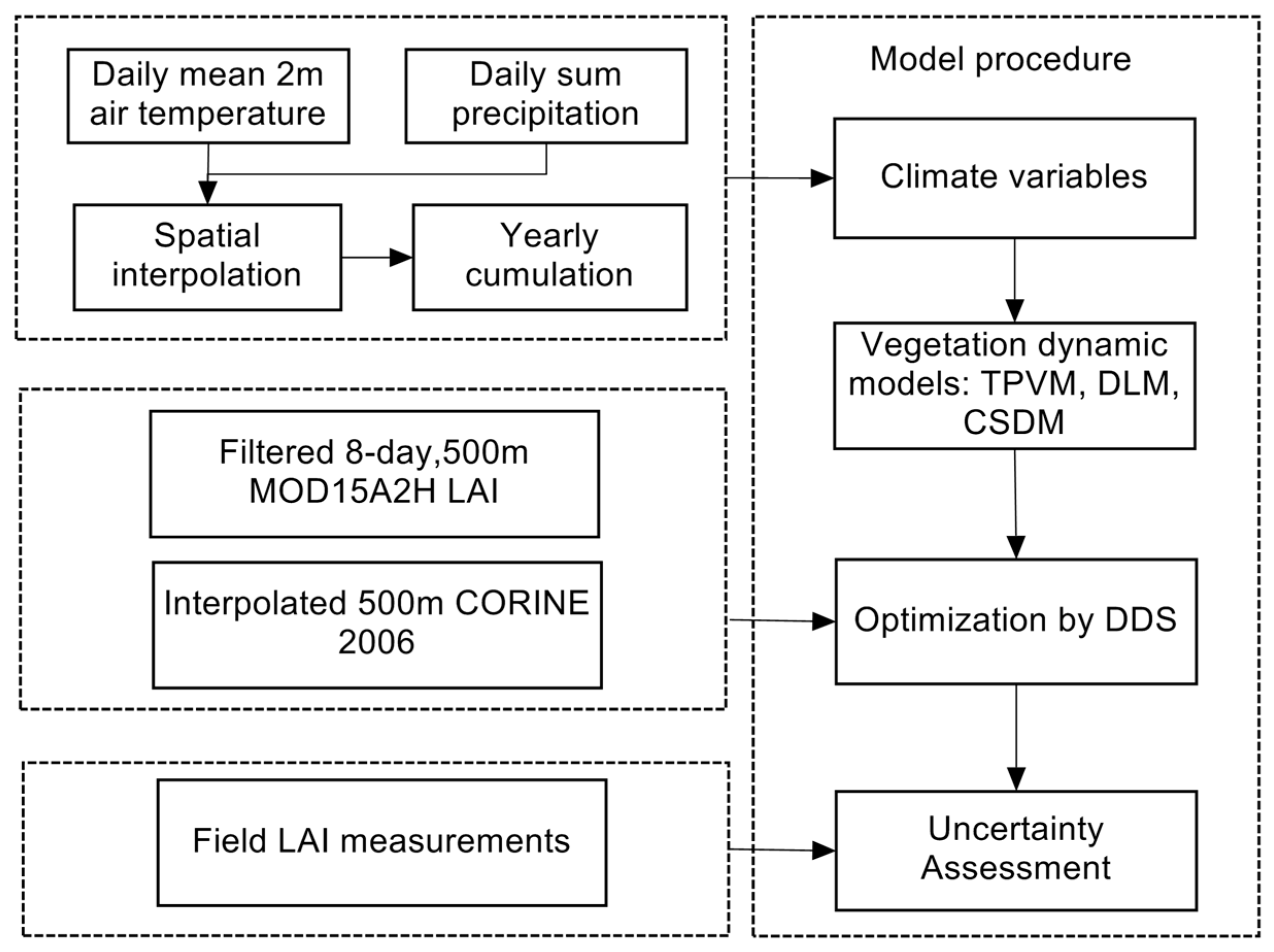
3. Methodology
3.1. Model Development
3.2. Optimization
3.3. Objective Function and Error Measures
4. Model Results and Evaluation
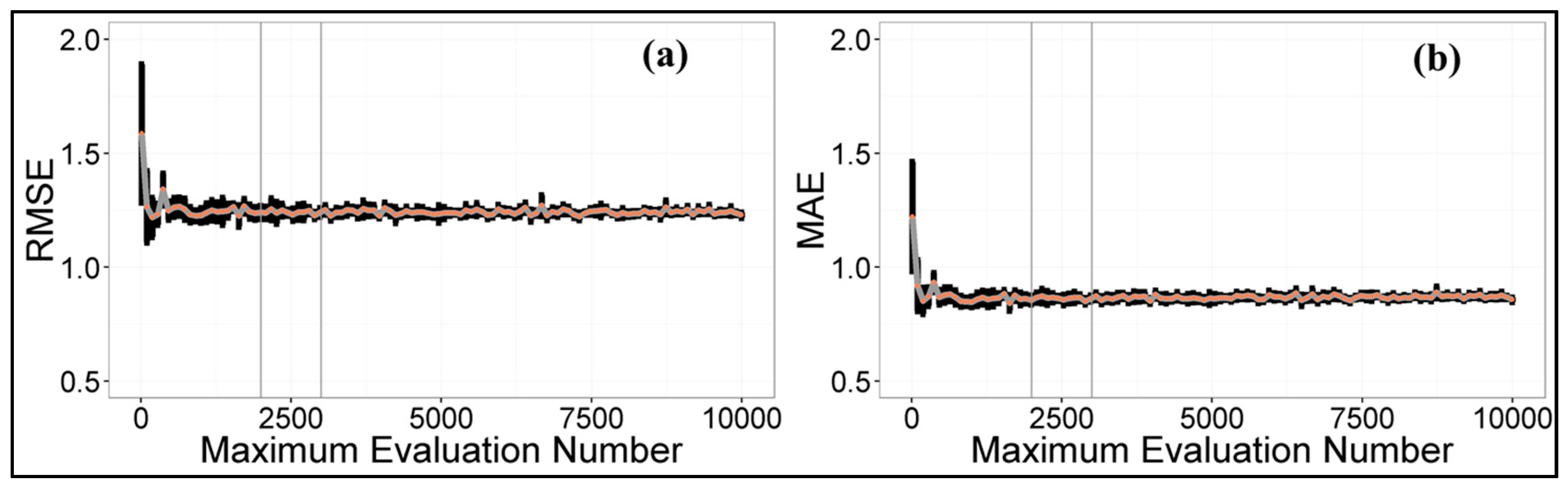
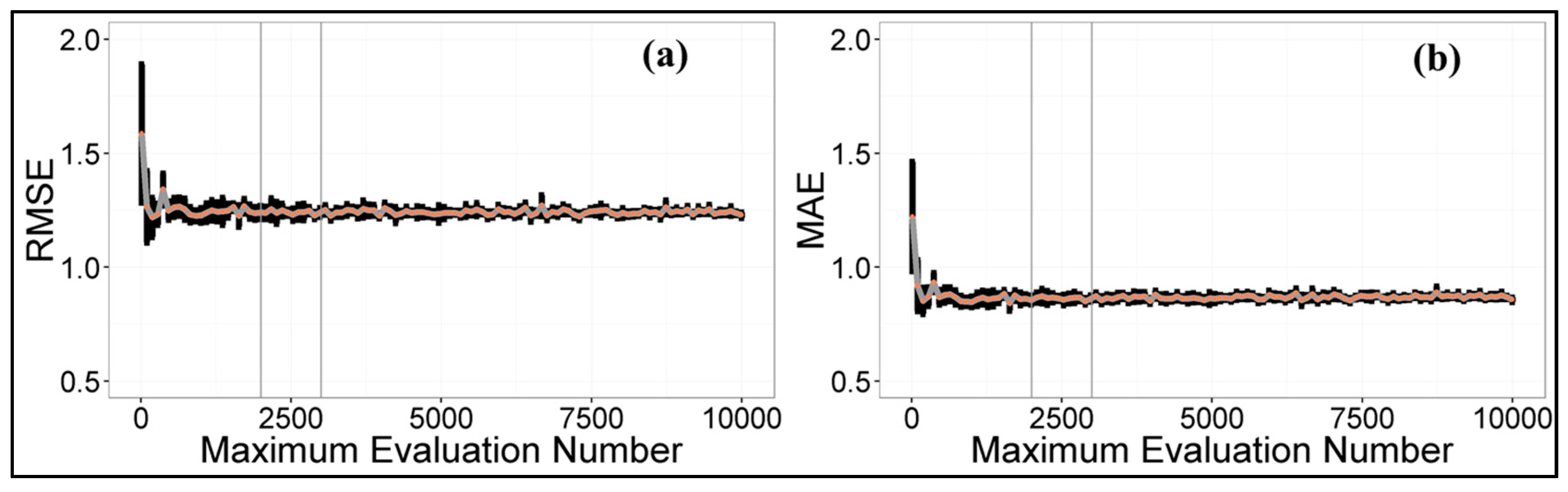
4.1. Maximum Evaluation Number Determination of DDS
4.2. Model Results
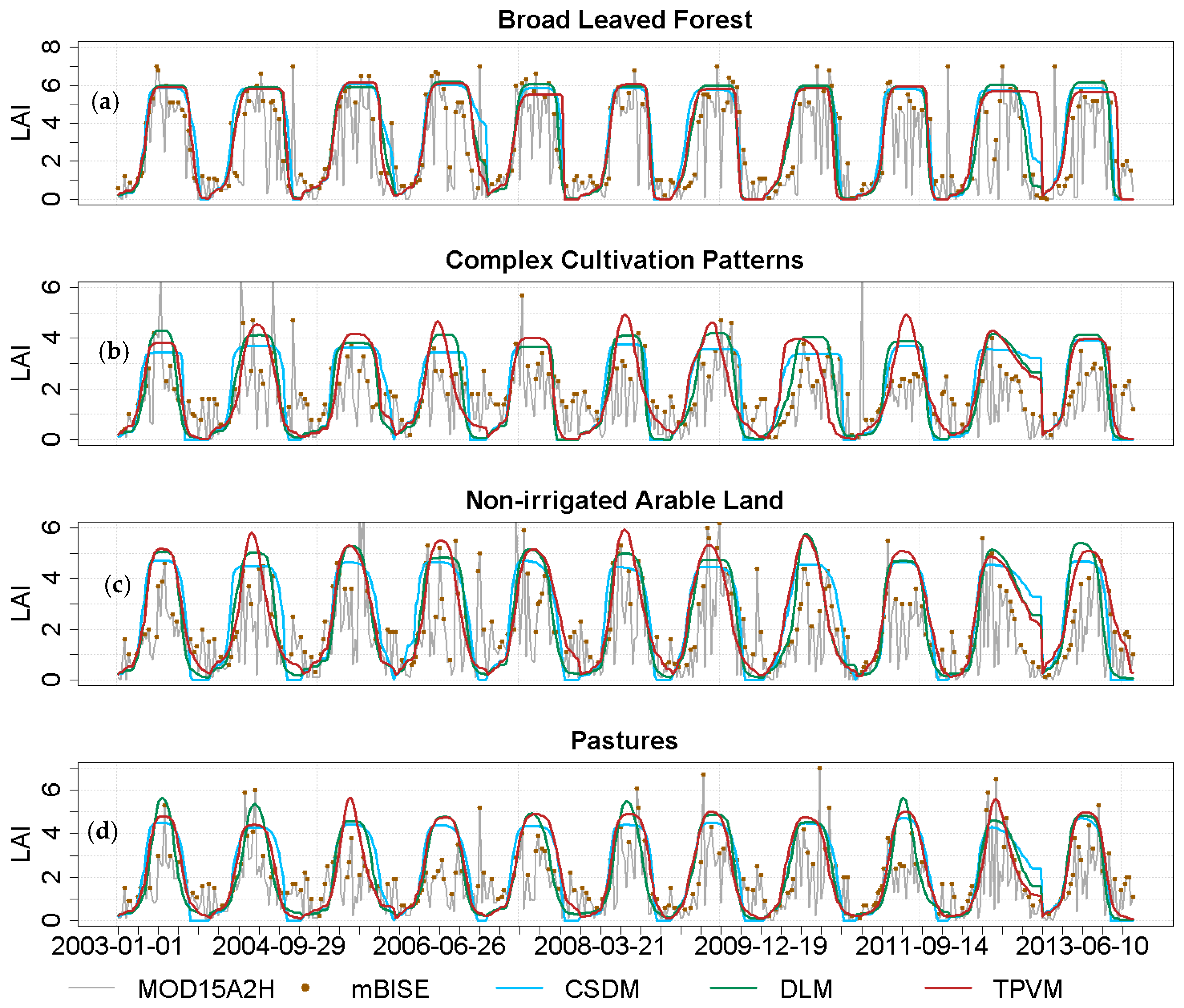
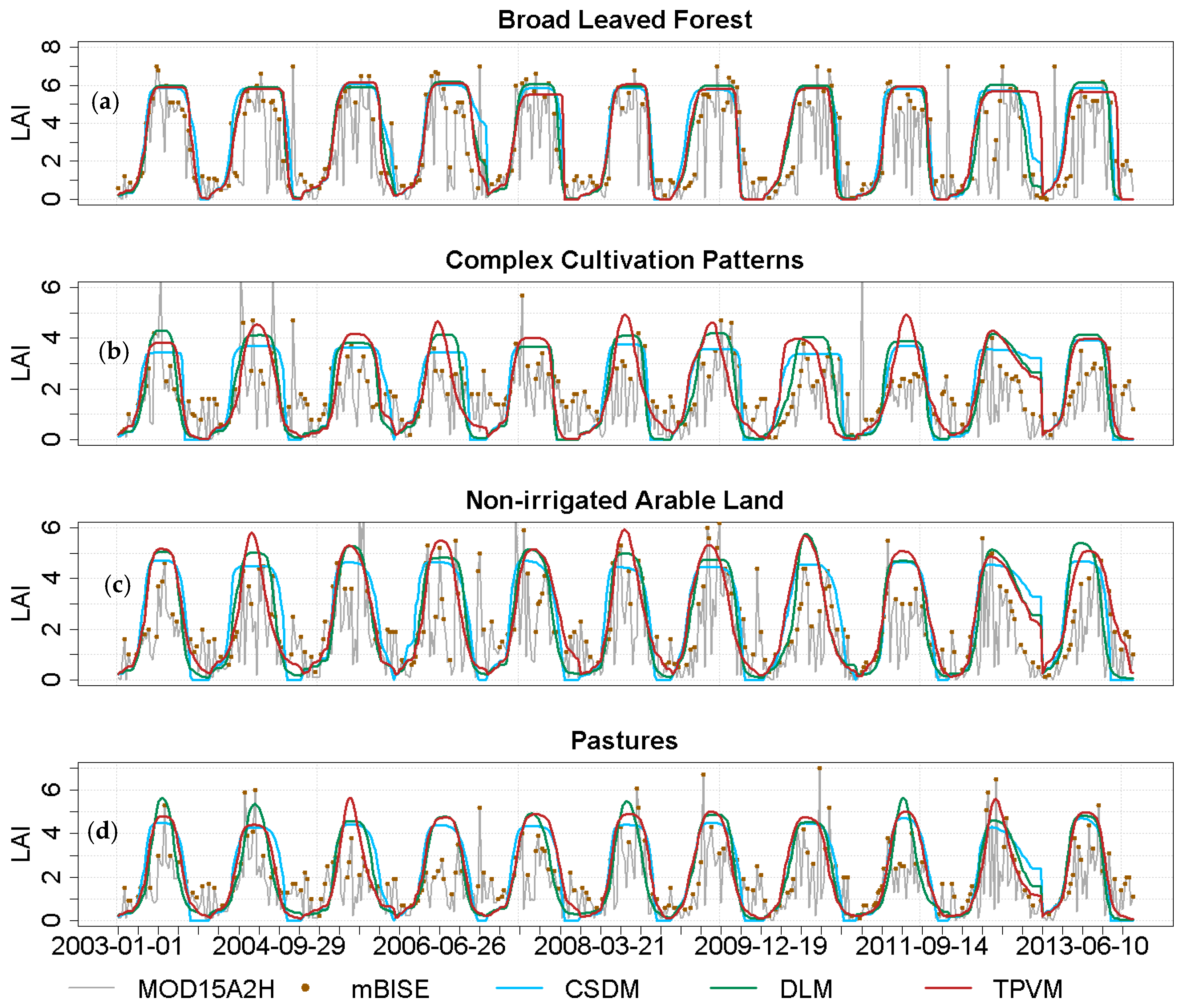
4.2.1. Analysis of Temporal LAI Pattern
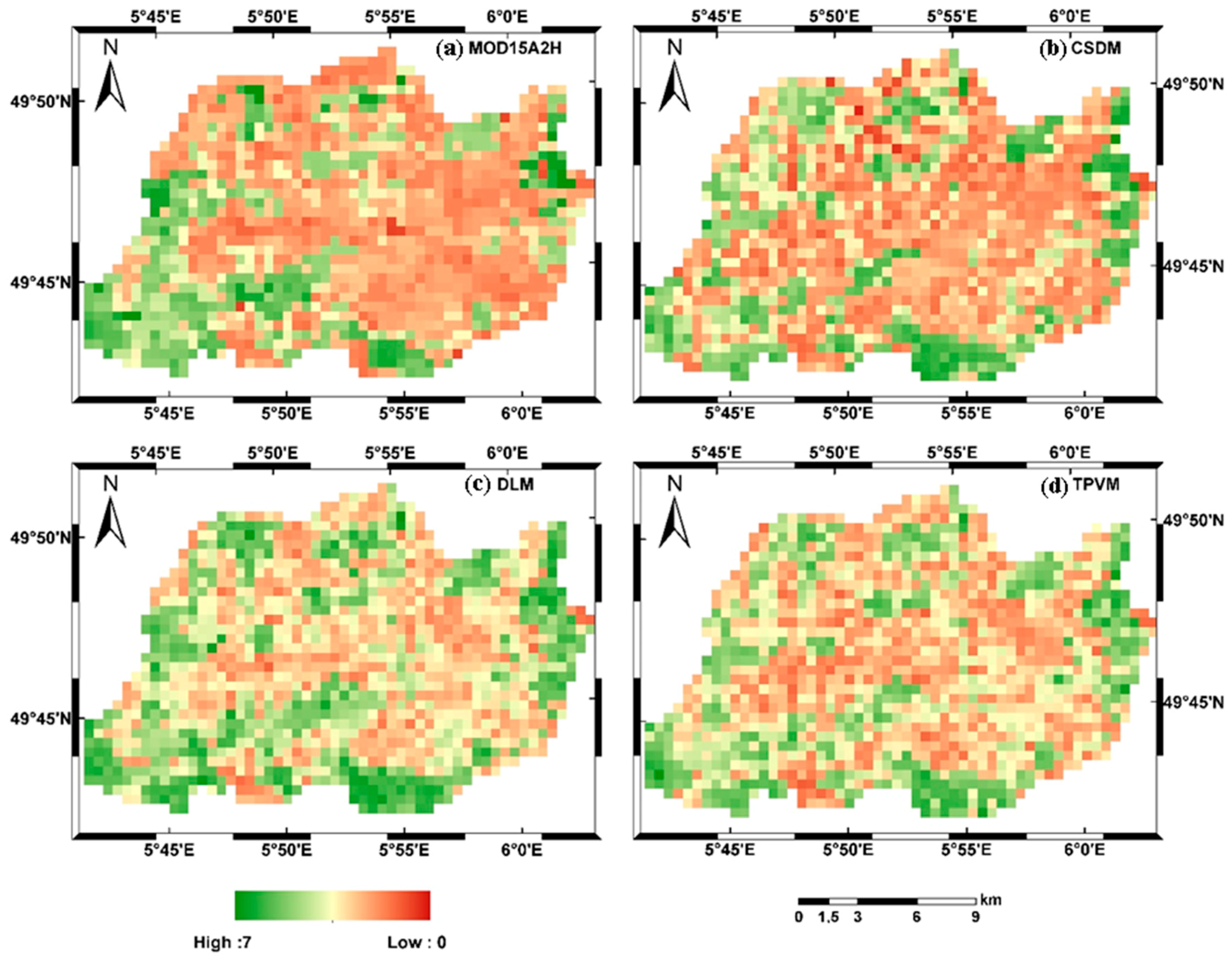
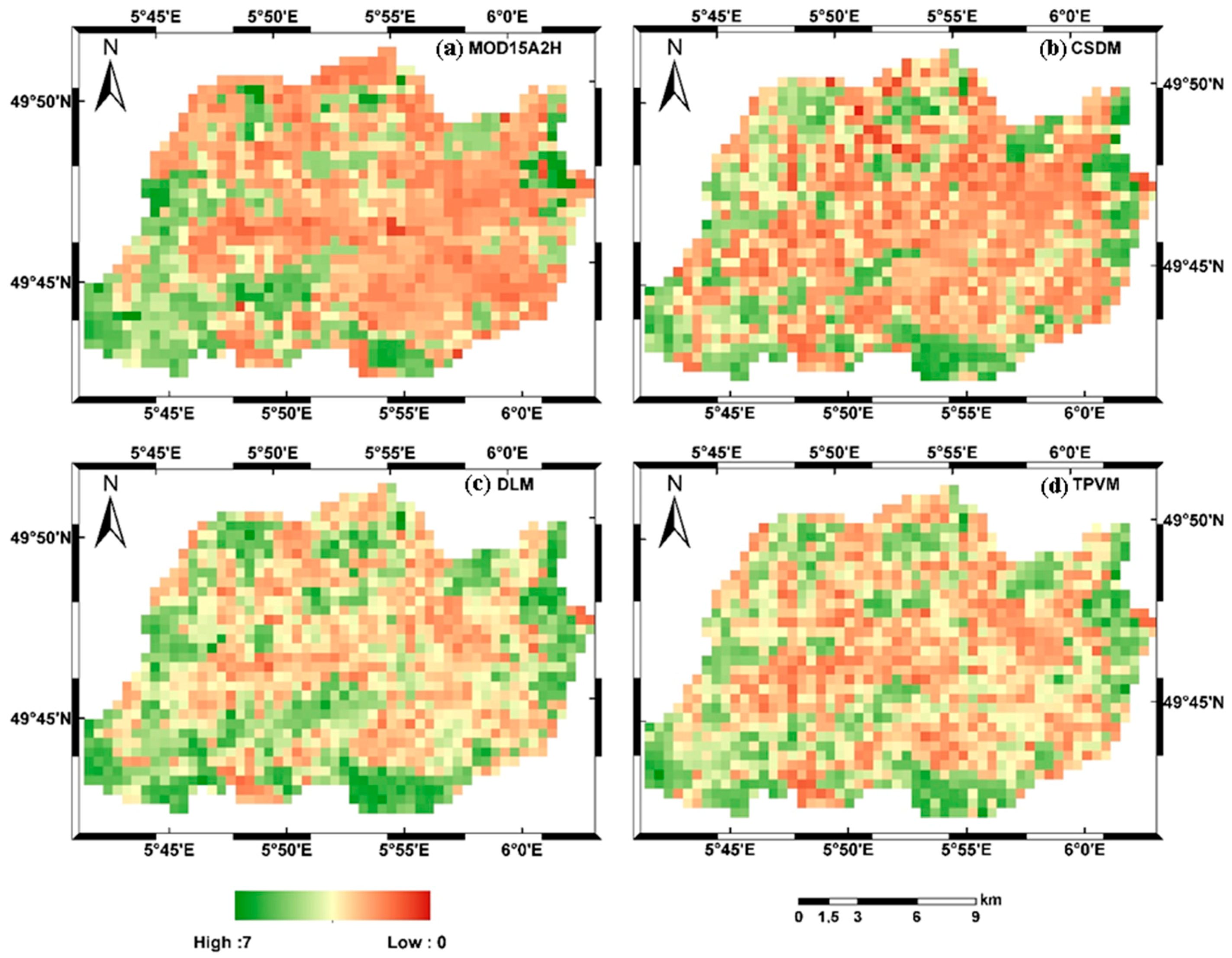
4.2.2. Analysis of Spatial LAI Pattern
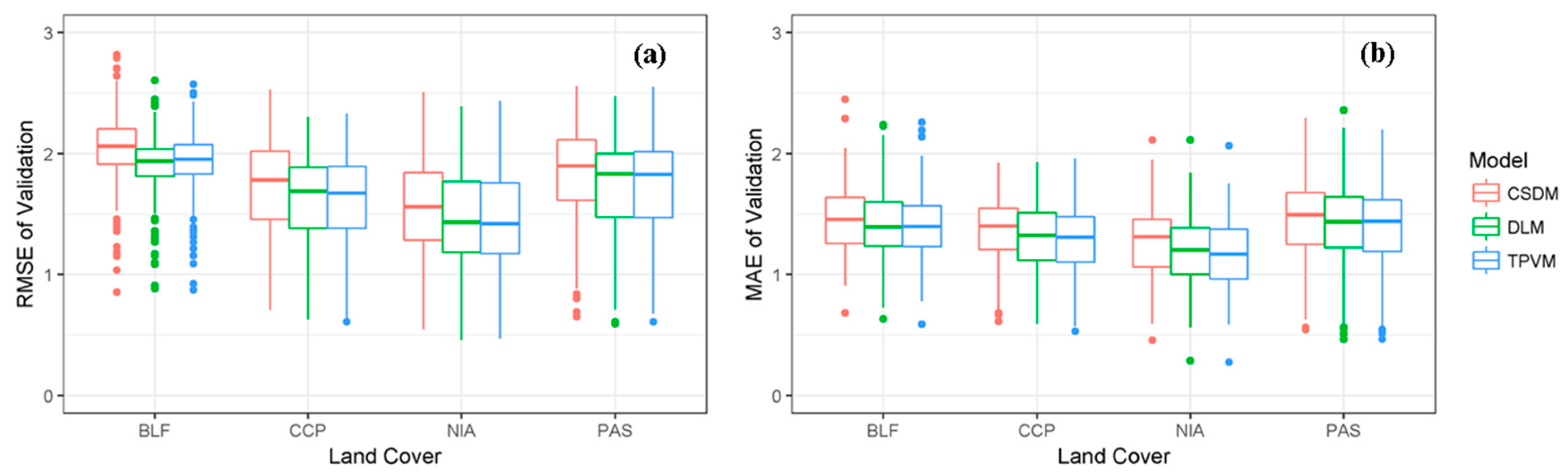
4.3. Uncertainty Assessment
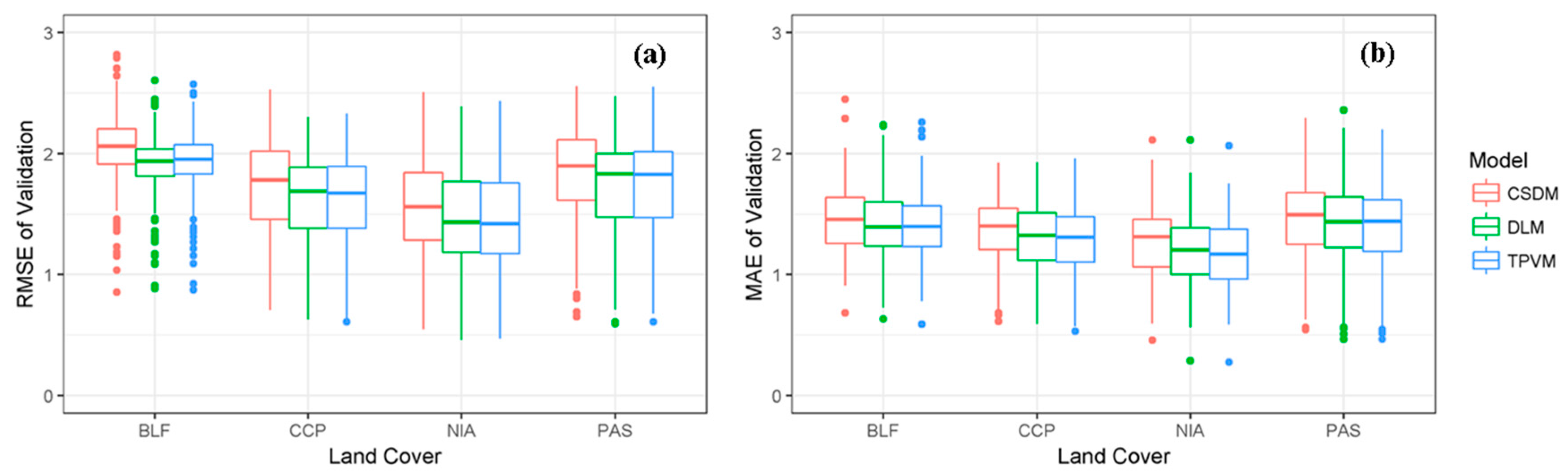
4.3.1. Accuracy of Model Fit Evaluation
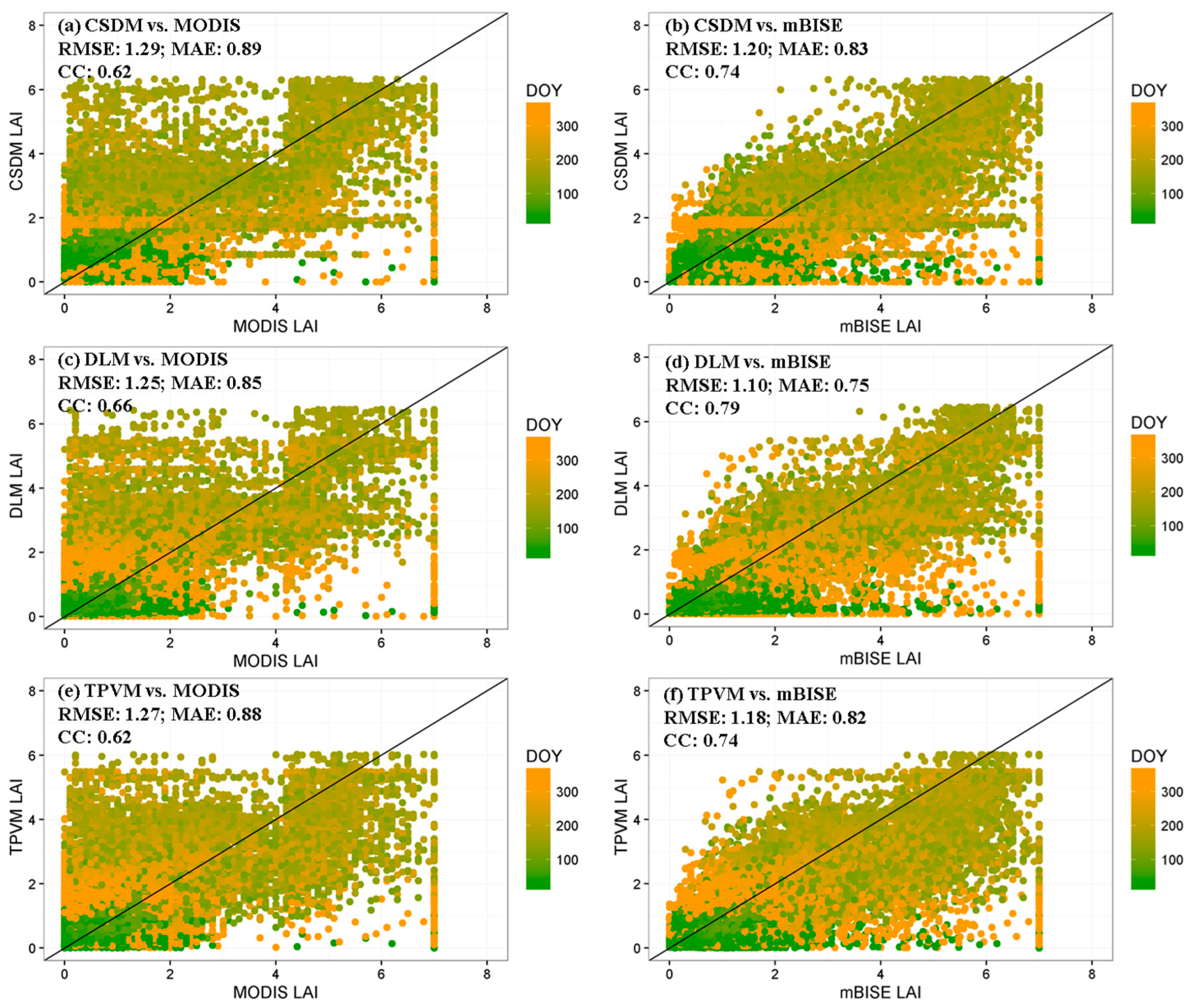
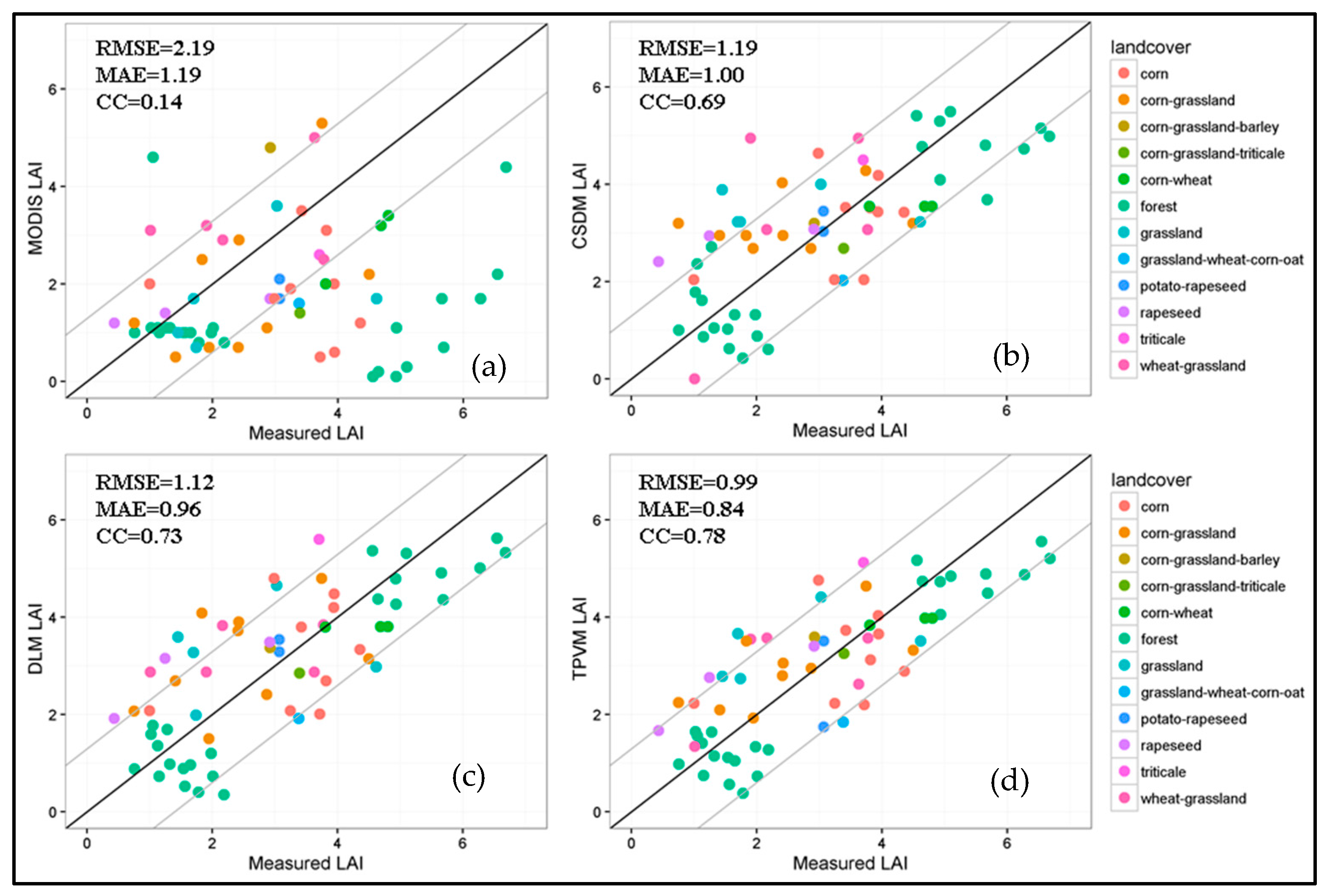
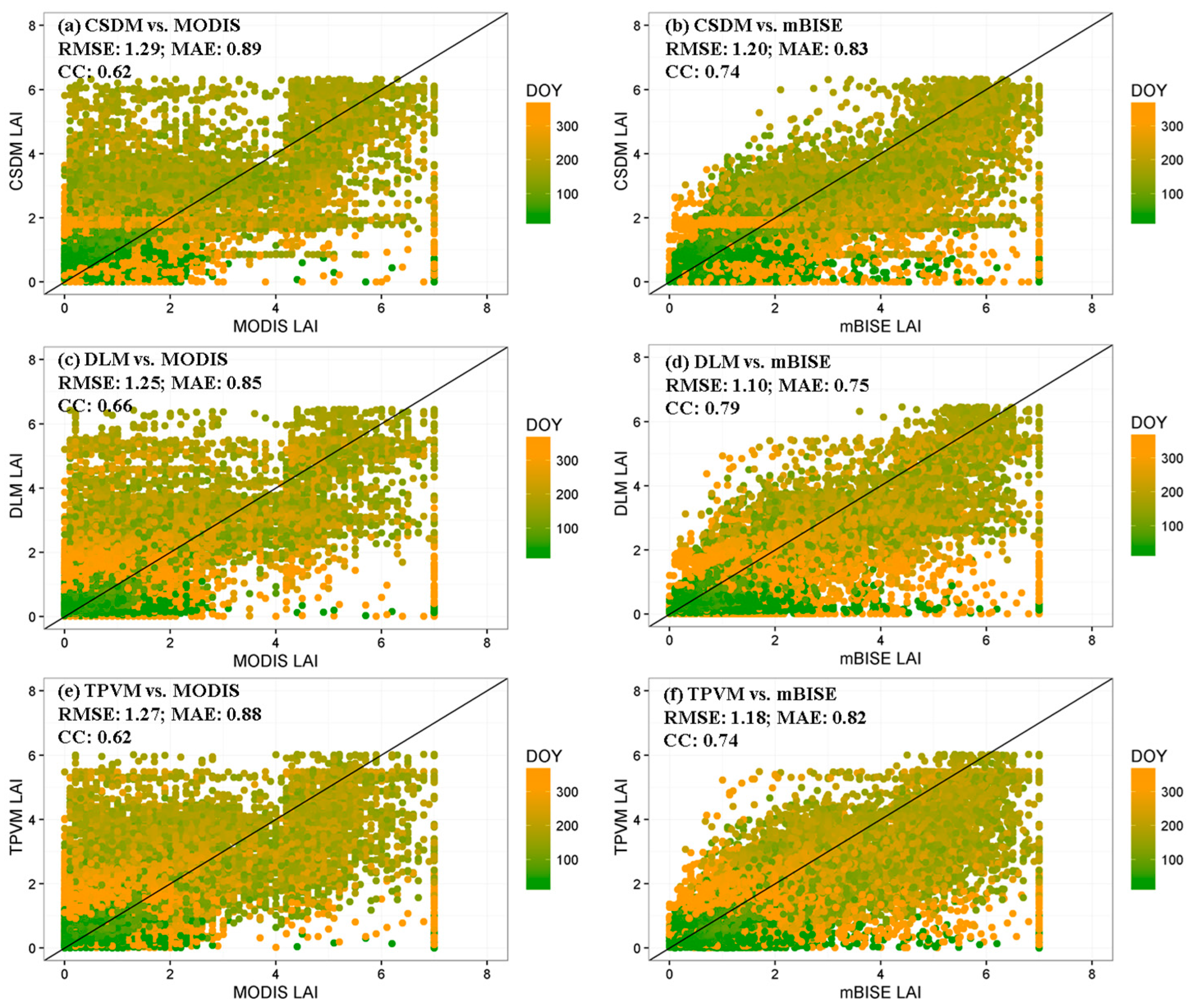
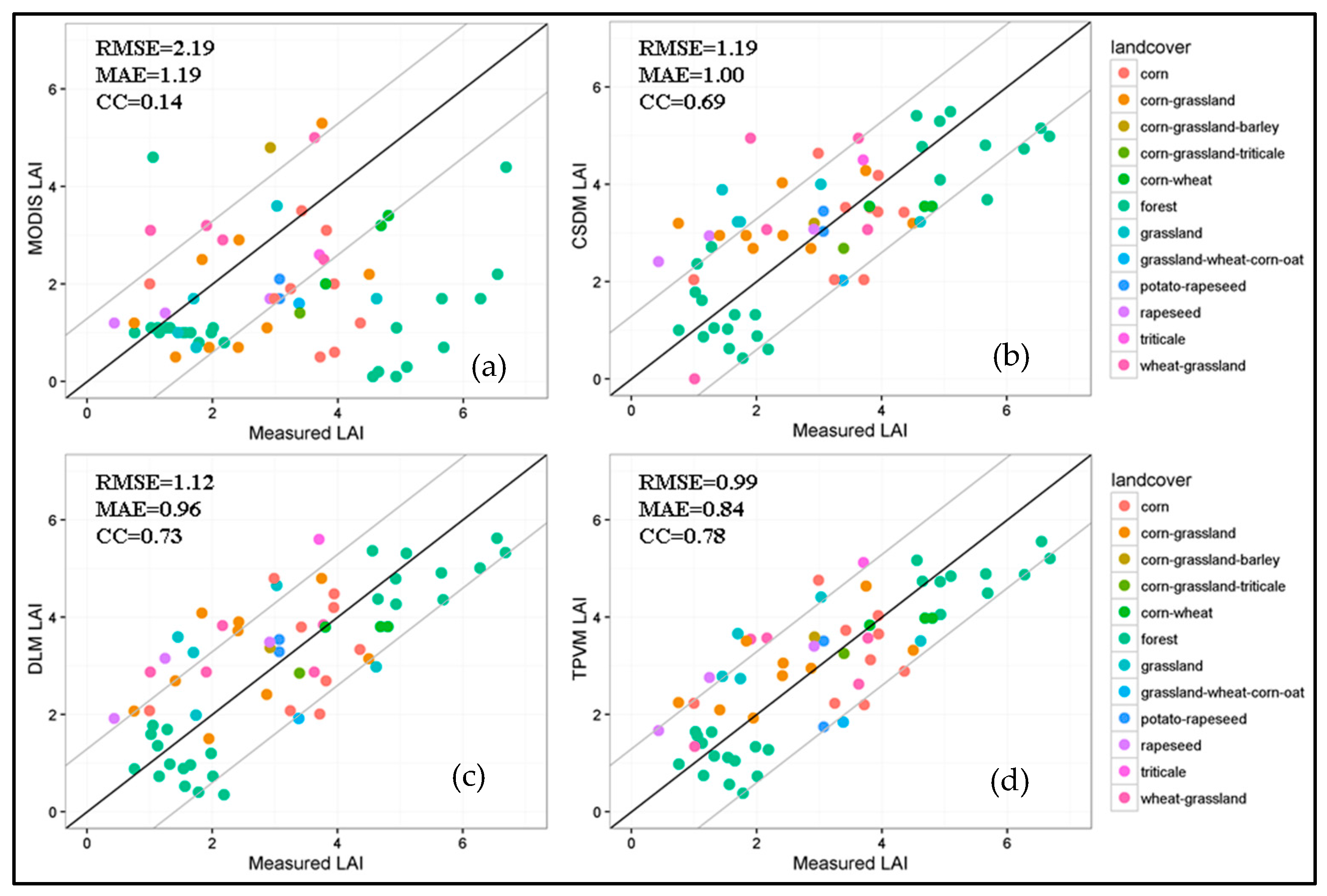
4.3.2. Comparison with MODIS Data and Field Measurement
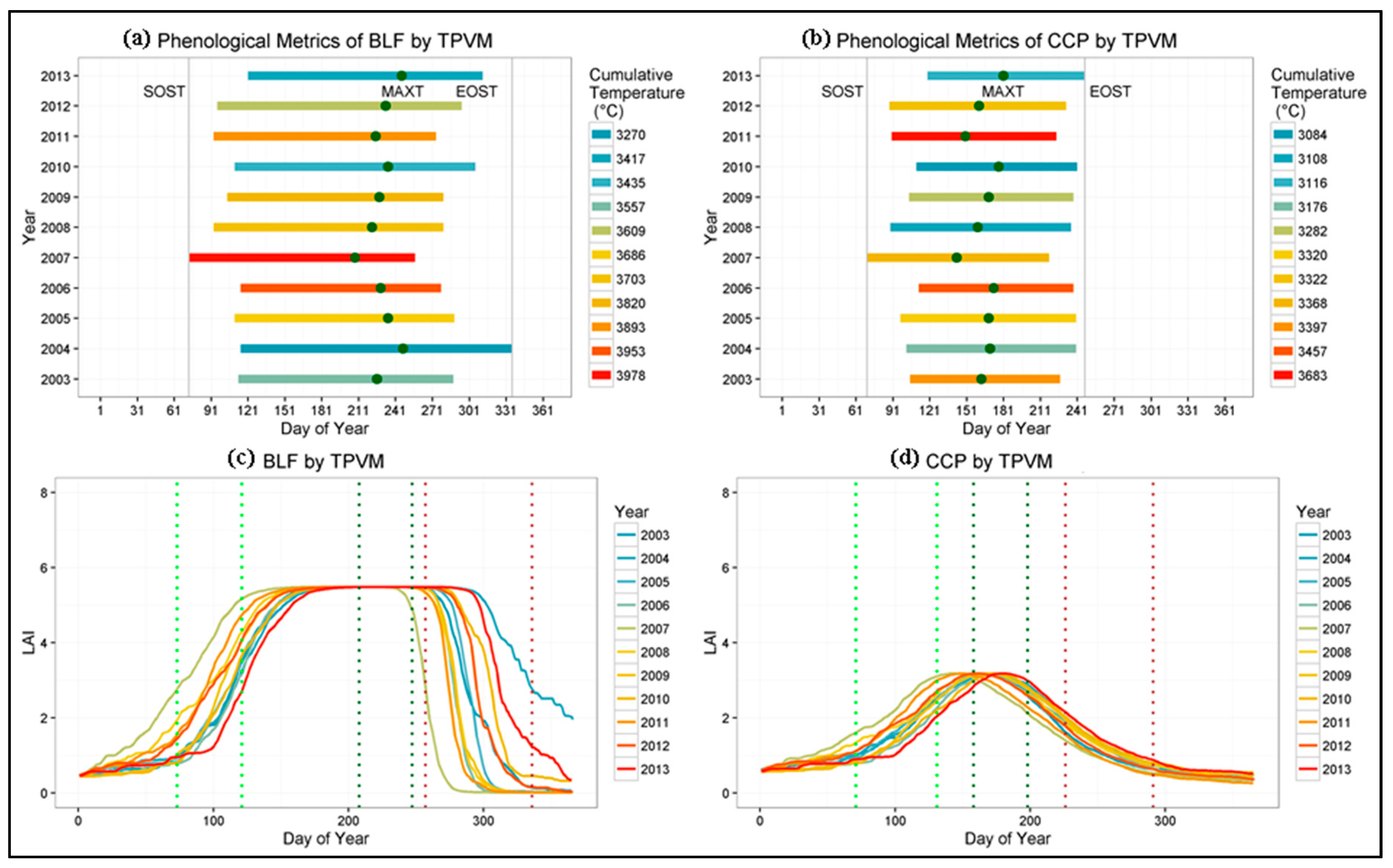
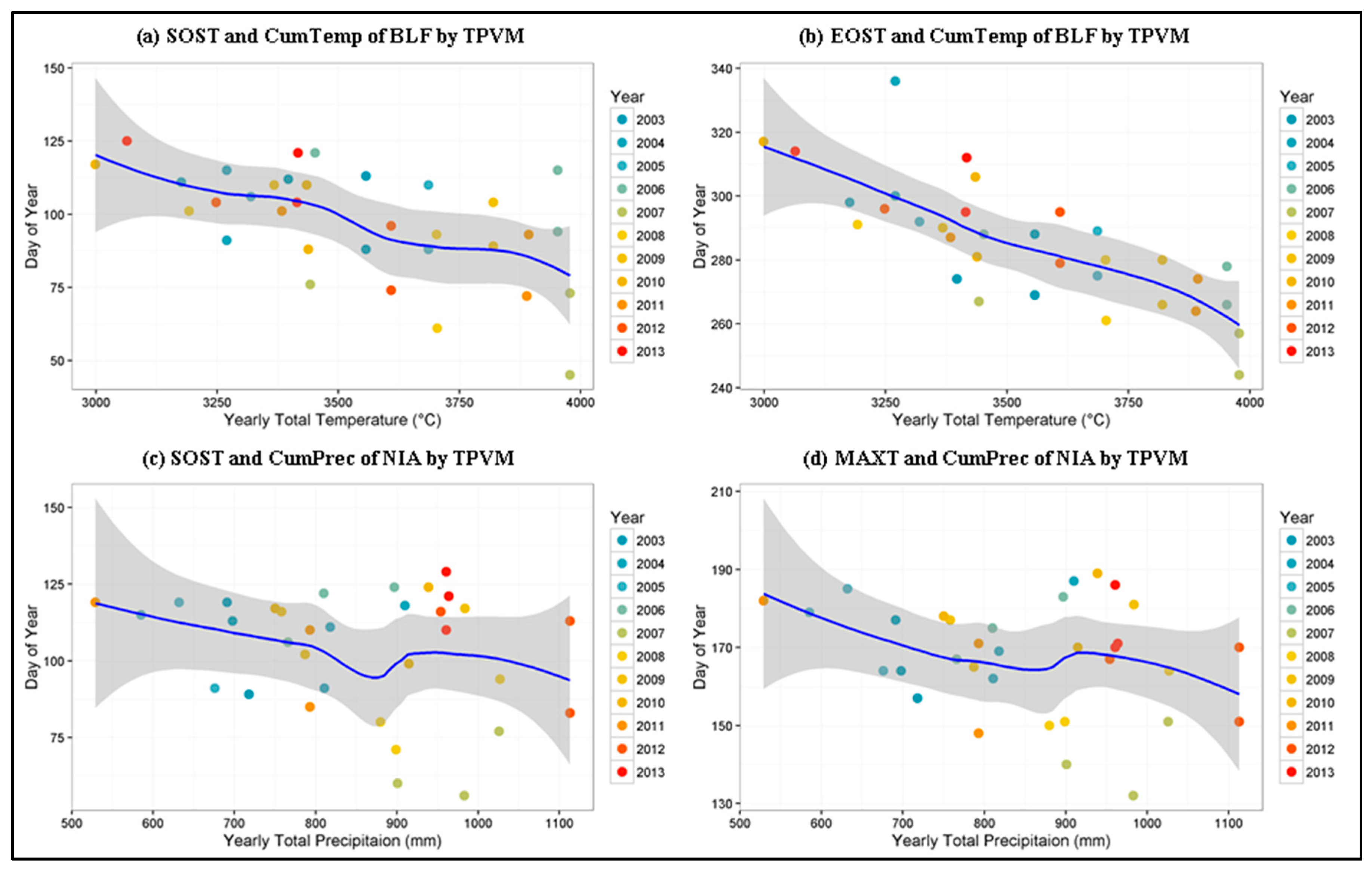
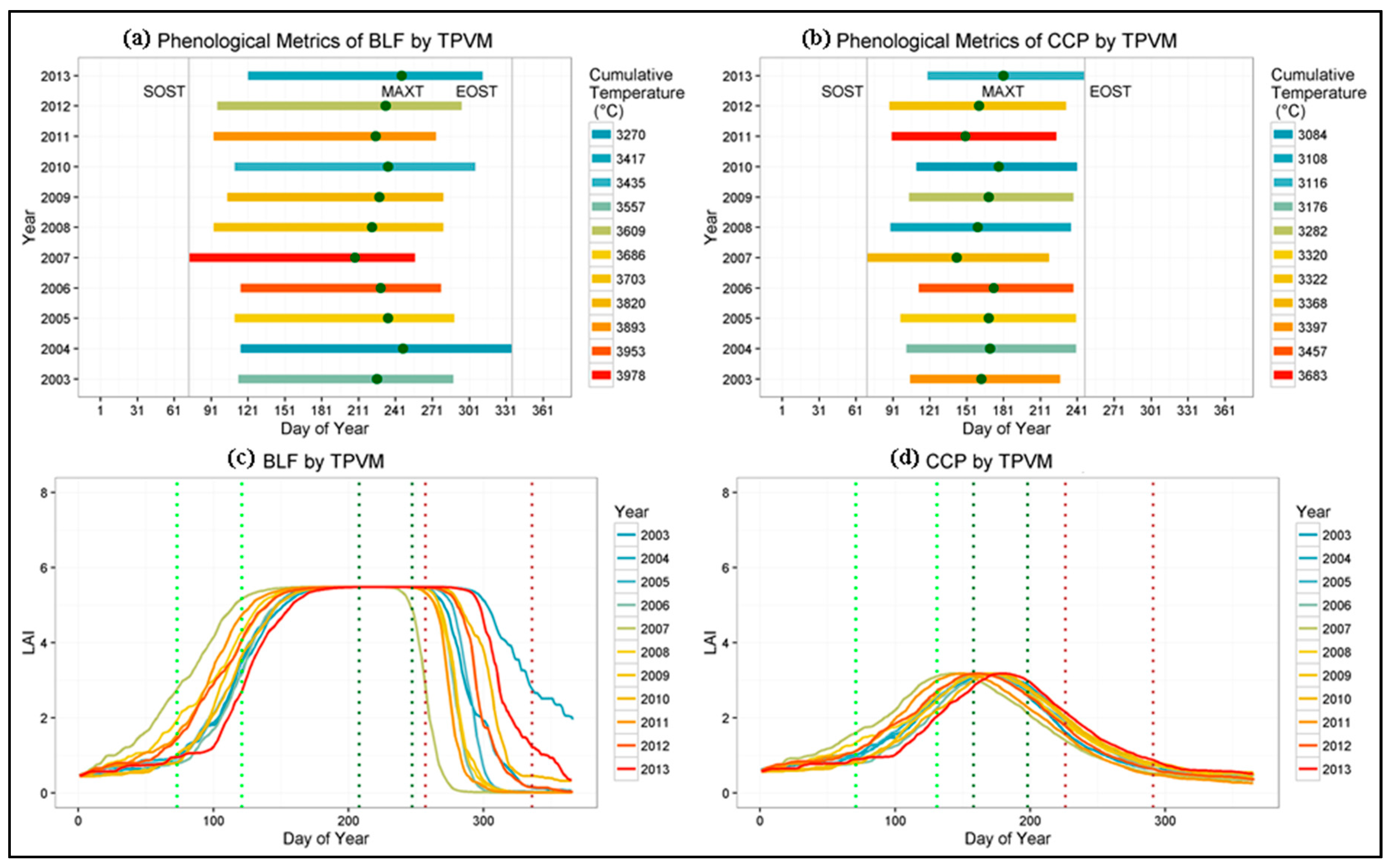
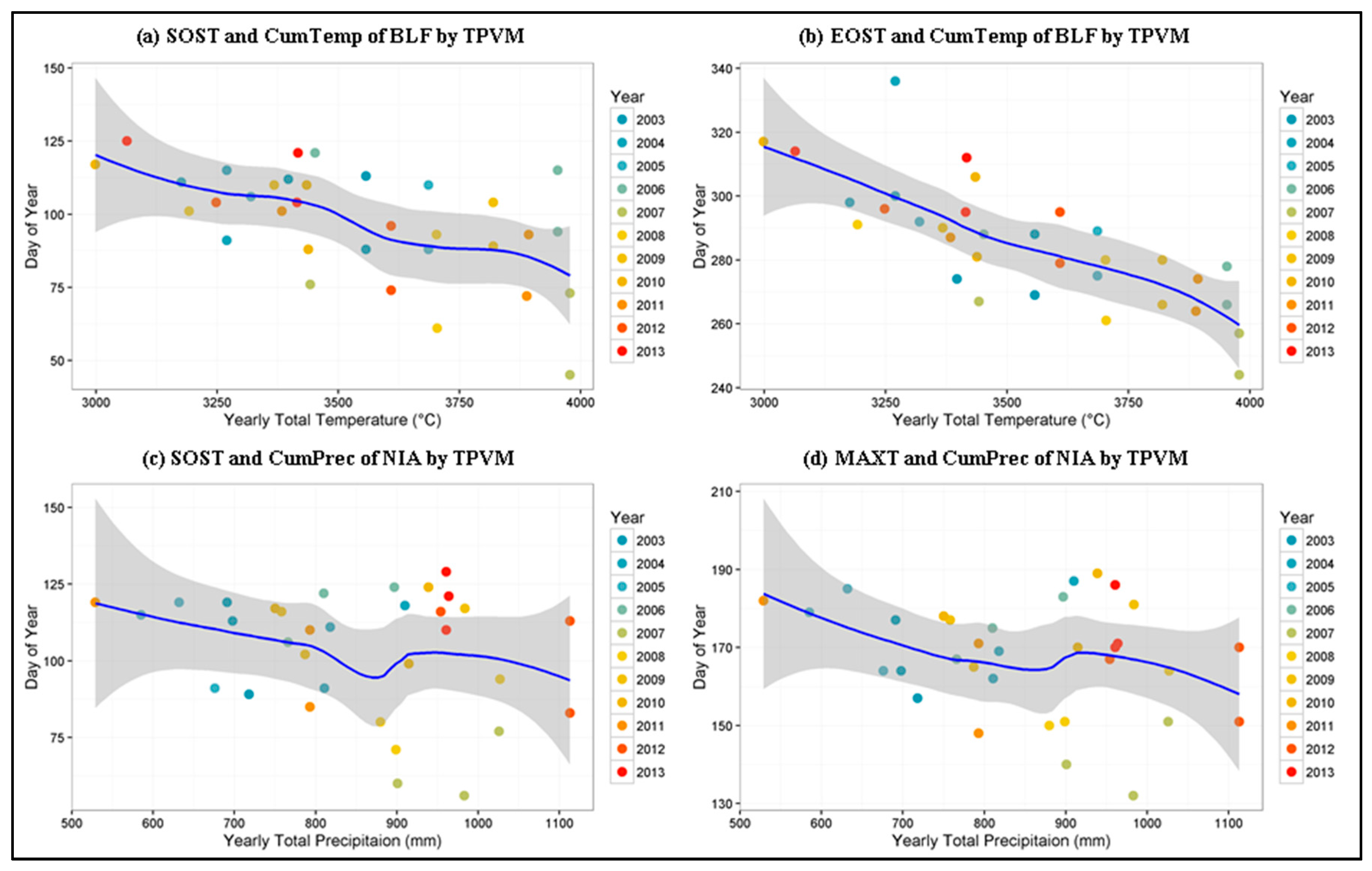
4.4. Phenological Metrics and Climate Controls Evaluation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Reed, B.C.; White, M.A. Assessing satellite-derived start-of-season measures in the conterminous USA. Int. J. Climatol. 2002, 22, 1793–1805. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Zhang, X.; Llorens, L.; Ogaya, R.; Lloret, F.; Comas, P.; Estiarte, M.; Terradas, J. Complex spatiotemporal phenological shifts as a response to rainfall changes. New Phytol. 2004, 161, 837–846. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in Africa by coupling MODIS and TRMM instruments. J. Geophys. Res. Atmos. 2005. [Google Scholar] [CrossRef]
- Watson, D.J. Comparative physiological studies on the growth of field crops: I. Variation in net assimilation rate and leaf area between species and varieties, and within and between years. Ann. Bot. 1947, 11, 41–76. [Google Scholar] [CrossRef]
- Tesemma, Z.K.; Wei, Y.; Western, A.W.; Peel, M.C. Leaf Area Index variation for crop, pasture, and tree in response to climatic variation in the goulburn–broken catchment, Australia. J. Hydrometeorol. 2014, 15, 1592–1606. [Google Scholar] [CrossRef]
- Glenn, E.; Huete, A.; Nagler, P.; Nelson, S. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: What vegetation indices can and cannot tell us about the landscape. Sensors 2008, 8, 2136. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; McPherson, E.G.; Simpson, J.R.; Ustin, S.L. Rainfall interception by sacramento’s urban forest. J. Arboric. 1998, 24, 235–244. [Google Scholar]
- Running, S.W.; Coughlan, J.C. A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecol. Model. 1988, 42, 125–154. [Google Scholar] [CrossRef]
- Liang, L.; Schwartz, M. Landscape phenology: An integrative approach to seasonal vegetation dynamics. Landsc. Ecol. 2009, 24, 465–472. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Karl, T.R. Spring phenology: Nature’s experiment to detect the effect of “green-up” on surface maximum temperatures. Mon. Weather Rev. 1990, 118, 883–890. [Google Scholar] [CrossRef]
- González-Sanpedro, M.C.; Le Toan, T.; Moreno, J.; Kergoat, L.; Rubio, E. Seasonal variations of Leaf Area Index of agricultural fields retrieved from Landsat data. Remote Sens. Environ. 2008, 112, 810–824. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the European continent from NDVI3g data and MODIS time series. Remote Sens. 2013, 6, 257. [Google Scholar] [CrossRef]
- Hwang, T.; Song, C.; Bolstad, P.V.; Band, L.E. Downscaling real-time vegetation dynamics by fusing multi-temporal MODIS and Landsat NDVI in topographically complex terrain. Remote Sens. Environ. 2011, 115, 2499–2512. [Google Scholar] [CrossRef]
- AghaKouchak, A.; Farahmand, A.; Melton, F.S.; Teixeira, J.; Anderson, M.C.; Wardlow, B.D.; Hain, C.R. Remote sensing of drought: Progress, challenges and opportunities. Rev. Geophys. 2015, 53, 452–480. [Google Scholar] [CrossRef]
- Thuiller, W. Biodiversity: Climate change and the ecologist. Nature 2007, 448, 550–552. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Leinonen, I.; Loustau, D. The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean Forests Ecosystems: An overview. Int. J. Biometeorol. 1999, 44, 67–75. [Google Scholar] [CrossRef]
- Chuine, I.; Kramer, K.; Hänninen, H. Plant development models. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 217–235. [Google Scholar]
- Forrest, J.; Miller-Rushing, A.J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos. Trans. R. Soc. London B Biol. Sci. 2010, 365, 3101–3112. [Google Scholar] [CrossRef] [PubMed]
- The Globe Program. A Worldwide Science and Education Program. Available online: https://www.globe.gov/globe-data (accessed on 1 May 2013).
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Chang. Biol. 2006, 12, 343–351. [Google Scholar] [CrossRef]
- Migliavacca, M.; Cremonese, E.; Colombo, R.; Busetto, L.; Galvagno, M.; Ganis, L.; Meroni, M.; Pari, E.; Rossini, M.; Siniscalco, C.; et al. European larch phenology in the Alps: Can we grasp the role of ecological factors by combining field observations and inverse modelling? Int. J. Biometeorol. 2008, 52, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Orueta, A.; Huesca, M.; Whiting, M.L.; Litago, J.; Khanna, S.; Garcia, M.; Ustin, S.L. Derivation of phenological metrics by function fitting to time-series of spectral shape indexes AS1 and AS2: Mapping cotton phenological stages using MODIS time series. Remote Sens. Environ. 2012, 126, 148–159. [Google Scholar] [CrossRef]
- Kathuroju, N.; White, M.A.; Symanzik, J.; Schwartz, M.D.; Powell, J.A.; Nemani, R.R. On the use of the advanced very high resolution radiometer for development of prognostic land surface phenology models. Ecol. Model. 2007, 201, 144–156. [Google Scholar] [CrossRef]
- Chuine, I.; Belmonte, J.; Mignot, A. A modelling analysis of the genetic variation of phenology between tree populations. J. Ecol. 2000, 88, 561–570. [Google Scholar] [CrossRef]
- Chuine, I.; Yiou, P.; Viovy, N.; Seguin, B.; Daux, V.; Ladurie, E.L.R. Historical phenology: Grape ripening as a past climate indicator. Nature 2004, 432, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I.; Cour, P.; Rousseau, D.D. Selecting models to predict the timing of flowering of temperate trees: Implications for tree phenology modelling. Plant Cell Environ. 1999, 22, 1–13. [Google Scholar] [CrossRef]
- Chuine, I.; Cambon, G.; Comtois, P. Scaling phenology from the local to the regional level: Advances from species-specific phenological models. Glob. Chang. Biol. 2000, 6, 943–952. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Koetz, B.; Baret, F.; Poilvé, H.; Hill, J. Use of coupled canopy structure dynamic and radiative transfer models to estimate biophysical canopy characteristics. Remote Sens. Environ. 2005, 95, 115–124. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yokozawa, M.; Toritani, H.; Shibayama, M.; Ishitsuka, N.; Ohno, H. A crop phenology detection method using time-series MODIS data. Remote Sens. Environ. 2005, 96, 366–374. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green leaf phenology at landsat resolution: Scaling from the field to the satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B. Global vegetation phenology from Moderate Resolution Imaging Spectroradiometer (MODIS): Evaluation of global patterns and comparison with in situ measurements. J. Geophys. Res. Biogeosci. 2006. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Weiss, M.; Baret, F.; Garrigues, S.; Lacaze, R. LAI and FAPAR cyclopes global products derived from vegetation. Part 2: Validation and comparison with MODIS collection 4 products. Remote Sens. Environ. 2007, 110, 317–331. [Google Scholar] [CrossRef]
- Kandasamy, S.; Baret, F.; Verger, A.; Neveux, P.; Weiss, M. A comparison of methods for smoothing and gap filling time series of remote sensing observations – application to MODIS LAI products. Biogeosciences 2013, 10, 4055–4071. [Google Scholar]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Fang, H.; Liang, S.; Kuusk, A. Retrieving leaf area index using a genetic algorithm with a canopy radiative transfer model. Remote Sens. Environ. 2003, 85, 257–270. [Google Scholar] [CrossRef]
- Houborg, R.; Boegh, E. Mapping leaf chlorophyll and Leaf Area Index using inverse and forward canopy reflectance modeling and spot reflectance data. Remote Sens. Environ. 2008, 112, 186–202. [Google Scholar] [CrossRef]
- Riha, S.; Wilks, D.; Simoens, P. Impact of temperature and precipitation variability on crop model predictions. Clim. Chang. 1996, 32, 293–311. [Google Scholar] [CrossRef]
- Zehe, E.; Ehret, U.; Pfister, L.; Blume, T.; Schröder, B.; Westhoff, M.; Jackisch, C.; Schymanski, S.J.; Weiler, M.; Schulz, K.; et al. Hess opinions: From response units to functional units: A thermodynamic reinterpretation of the HRU concept to link spatial organization and functioning of intermediate scale catchments. Hydrol. Earth Syst. Sci. 2014, 18, 4635–4655. [Google Scholar] [CrossRef]
- Caos: From Catchments as Organised Systems to Models Based on Functional Units. Available online: http://www.caos-project.de/ (accessed on 6 May 2016).
- Sun, L.; Schulz, K. The improvement of land cover classification by thermal remote sensing. Remote Sens. 2015, 7. [Google Scholar] [CrossRef]
- Myneni, R.; Park, Y.K.T. MOD15A2H MODIS/TERRA Leaf Area Index/FPAR 8-Day l4 Global 500 m Sin Grid v006. Available online: https://lpdaac.usgs.gov/node/863 (accessed on 1 January 2016).
- Knyazikhin, Y.; Martonchik, J.V.; Myneni, R.B.; Diner, D.J.; Running, S.W. Synergistic algorithm for estimating vegetation canopy leaf area index and fraction of absorbed photosynthetically active radiation from MODIS and MISR data. J. Geophys. Res. Atmos. 1998, 103, 32257–32275. [Google Scholar] [CrossRef]
- Myneni, R.; Park, Y.K.T. MODIS Collection 6 (C6) LAI/FPAR Product User’s Guide. Available online: https://lpdaac.usgs.gov/sites/default/files/public/product_documentation/mod15_user_guide.pdf (accessed on 1 January 2016).
- Horn, J.; Schulz, K. Post-processing analysis of MODIS Leaf Area Index subsets. J. Appl. Remote Sens. 2010. [Google Scholar] [CrossRef]
- Lovell, J.L.; Graetz, R.D. Filtering pathfinder AVHRR land NDVI data for Australia. Int. J. Remote Sens. 2001, 22, 2649–2654. [Google Scholar] [CrossRef]
- LI-COR Inc. LAI-2200 Plant Canopy Analyzer: Instruction Manual; LI-COR Inc.: Lincoln, NE, USA, 2010. [Google Scholar]
- Jönsson, P.; Eklundh, L. Timesat—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Tolson, B.A.; Shoemaker, C.A. Dynamically dimensioned search algorithm for computationally efficient watershed model calibration. Water Resour. Res. 2007, 43, W01413. [Google Scholar] [CrossRef]
- Atkinson, P.M.; Jeganathan, C.; Dash, J.; Atzberger, C. Inter-comparison of four models for smoothing satellite sensor time-series data to estimate vegetation phenology. Remote Sens. Environ. 2012, 123, 400–417. [Google Scholar] [CrossRef]
- Borak, J.S.; Jasinski, M.F. Effective interpolation of incomplete satellite-derived leaf-area index time series for the continental united states. Agric. For. Meteorol. 2009, 149, 320–332. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- USGS Remote Sensing Phenology. Available online: http://phenology.cr.usgs.gov/methods_metrics.php (accessed on 7 June 2016).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Definition | Abbreviation | Definition |
|---|---|---|---|
| AVHRR | Advanced Very High-Resolution Radiometer | LOGS | Length of Growing Season |
| BISE | Best Index Slope Extraction | LP DAAC | Land Processes Distributed Active Archive Center |
| CAOS | Catchments as Organised Systems | LUT | Look-Up Table |
| CC | Person’s Correlation Coefficient | MAE | Mean Absolute Error |
| CORINE | CoORdination of INformation on the Environment | MAXT | Time of Maximum LAI value |
| CSDM | Canopy Structure Dynamic Model | mBISE | Modified Best Index Slope Extraction |
| DDS | Dynamically Dimensioned Search | MODIS | Moderate Resolution Imaging Spectroradiometer |
| DEM | Digital Elevation Map | NDVI | Normalized Difference Vegetation Index |
| DLM | Double Logistic Model | QC | Quality Criteria |
| DOY | Day of Year | RMSE | Root Mean Squared Error |
| EOST | End of Season Time | SEB | Surface Energy Balance |
| ESDIS | Earth Science Data and Information System | SOST | Start of Season Time |
| EVI | Enhanced Vegetation Index | SVAT | Soil-Vegetation-Atmosphere Transfer |
| FPAR | Fraction of absorbed Photosynthetically Active Radiation | TPVM | Temperature Precipitation Vegetation Model |
| GCM | Global Climate Models | VI | Vegetative Indicator |
| LAI | Leaf Area Index |
| Year | Yearly Cumulative Temperature (°C) | Yearly Cumulative Precipitation (mm) | ||
|---|---|---|---|---|
| Minimum | Maximum | Minimum | Maximum | |
| 2003 | 3391 | 3552 | 670 | 715 |
| 2004 | 3170 | 3265 | 751 | 899 |
| 2005 | 3318 | 3683 | 574 | 697 |
| 2006 | 3441 | 3940 | 713 | 868 |
| 2007 | 3363 | 3967 | 892 | 1020 |
| 2008 | 3108 | 3704 | 878 | 1025 |
| 2009 | 3270 | 3801 | 736 | 959 |
| 2010 | 2989 | 3436 | 669 | 949 |
| 2011 | 3371 | 3881 | 495 | 777 |
| 2012 | 3174 | 3573 | 827 | 1082 |
| 2013 | 3000 | 3399 | 727 | 941 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Schulz, K. Spatio-Temporal LAI Modelling by Integrating Climate and MODIS LAI Data in a Mesoscale Catchment. Remote Sens. 2017, 9, 144. https://doi.org/10.3390/rs9020144
Sun L, Schulz K. Spatio-Temporal LAI Modelling by Integrating Climate and MODIS LAI Data in a Mesoscale Catchment. Remote Sensing. 2017; 9(2):144. https://doi.org/10.3390/rs9020144
Chicago/Turabian StyleSun, Liya, and Karsten Schulz. 2017. "Spatio-Temporal LAI Modelling by Integrating Climate and MODIS LAI Data in a Mesoscale Catchment" Remote Sensing 9, no. 2: 144. https://doi.org/10.3390/rs9020144
APA StyleSun, L., & Schulz, K. (2017). Spatio-Temporal LAI Modelling by Integrating Climate and MODIS LAI Data in a Mesoscale Catchment. Remote Sensing, 9(2), 144. https://doi.org/10.3390/rs9020144