
Remote Sensing-Based Assessment of the 2005–2011 Bamboo Reproductive Event in the Arakan Mountain Range and Its Relation with Wildfires

Abstract
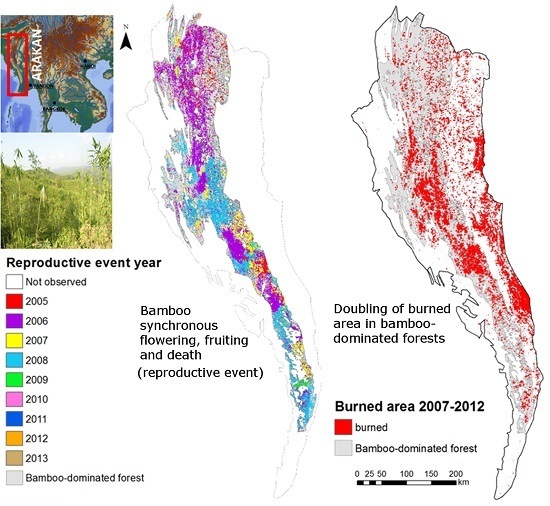
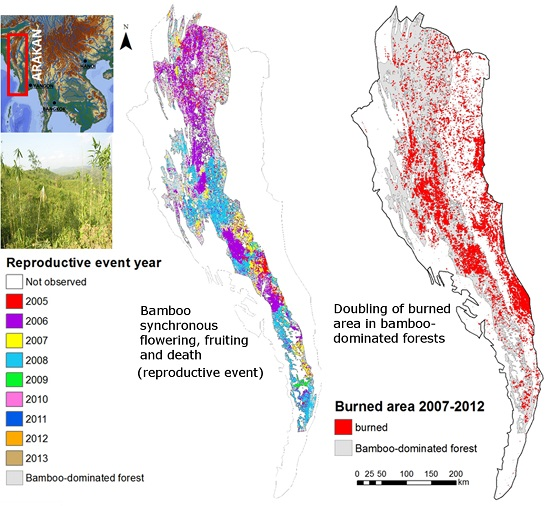
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- (a)
- What is the extent and geographic distribution of the bamboo-dominated forest in Arakan?
- (b)
- Which spatial and temporal dynamics characterized the bamboo reproductive event?
- (c)
- Was the wildfire regime impacted by the bamboo reproductive event?
2. Materials and Methods
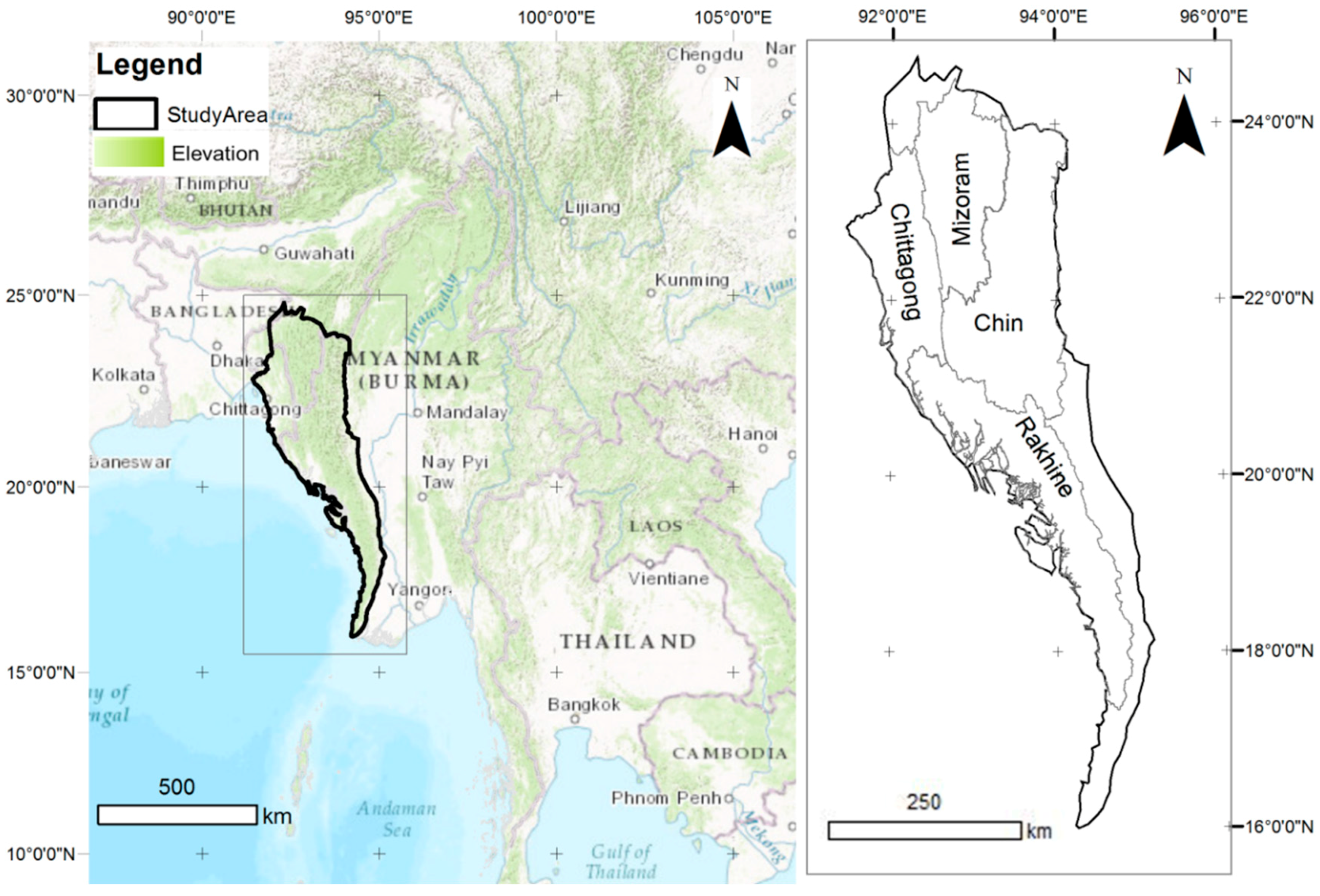
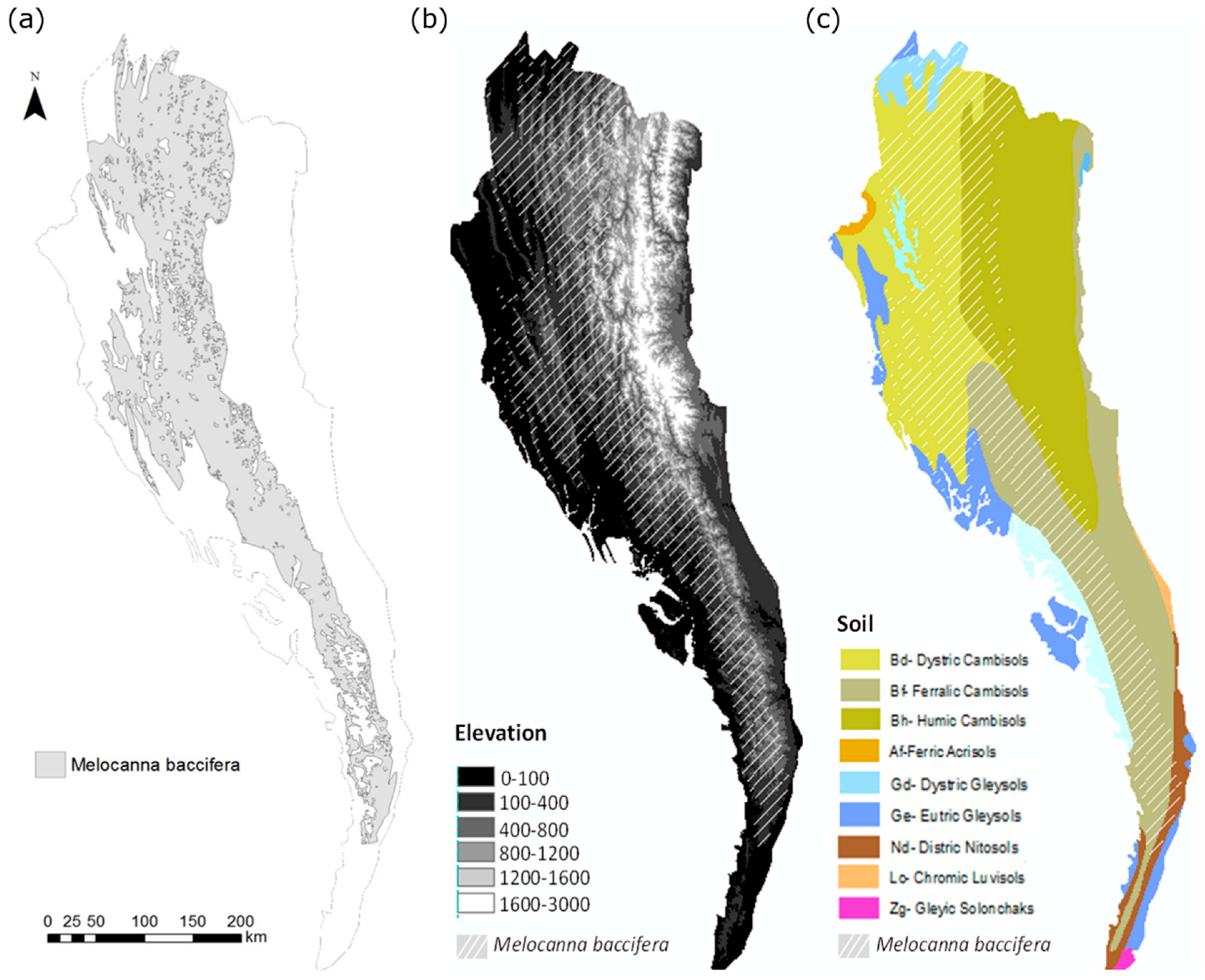
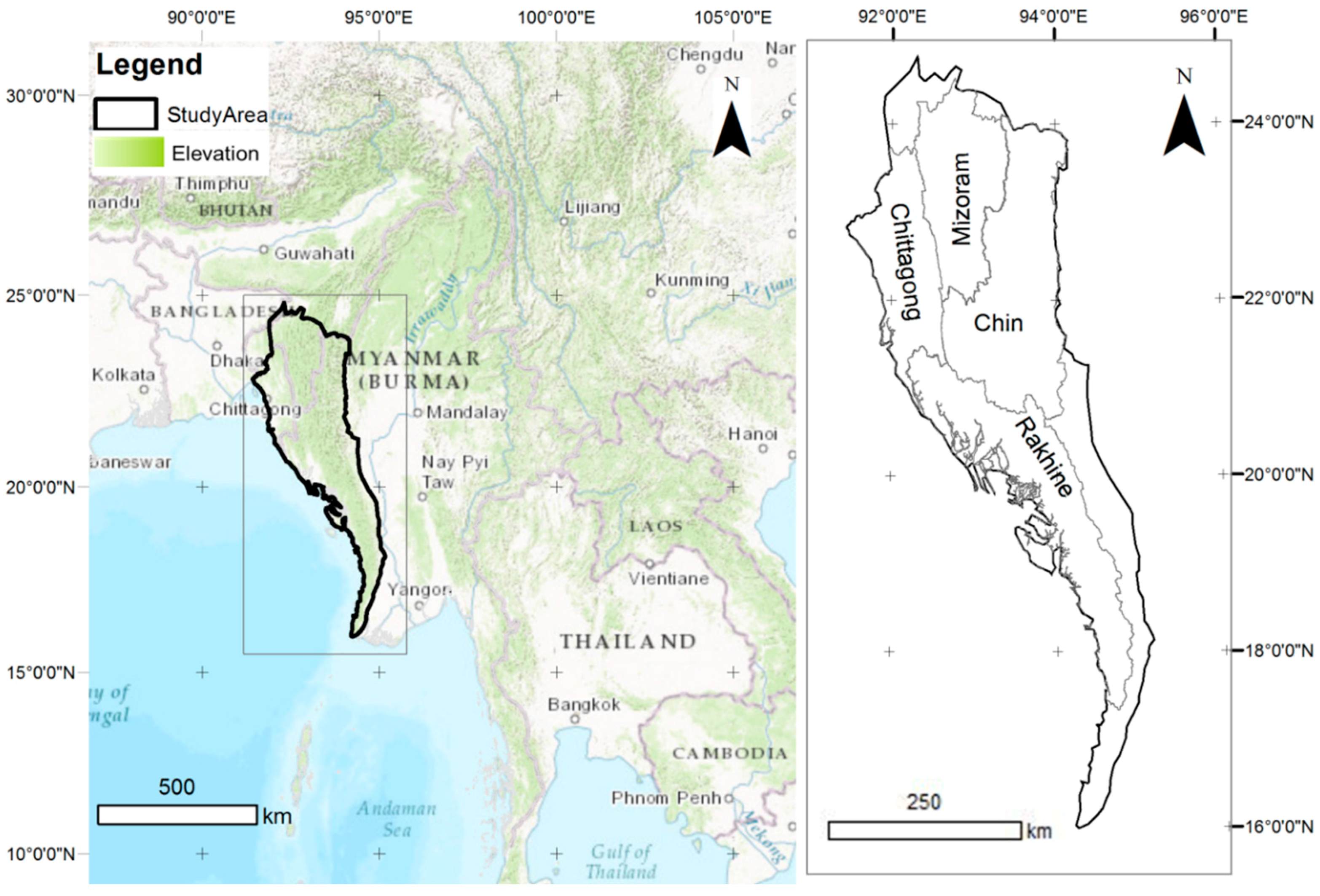
2.1. Study Area and Field Survey
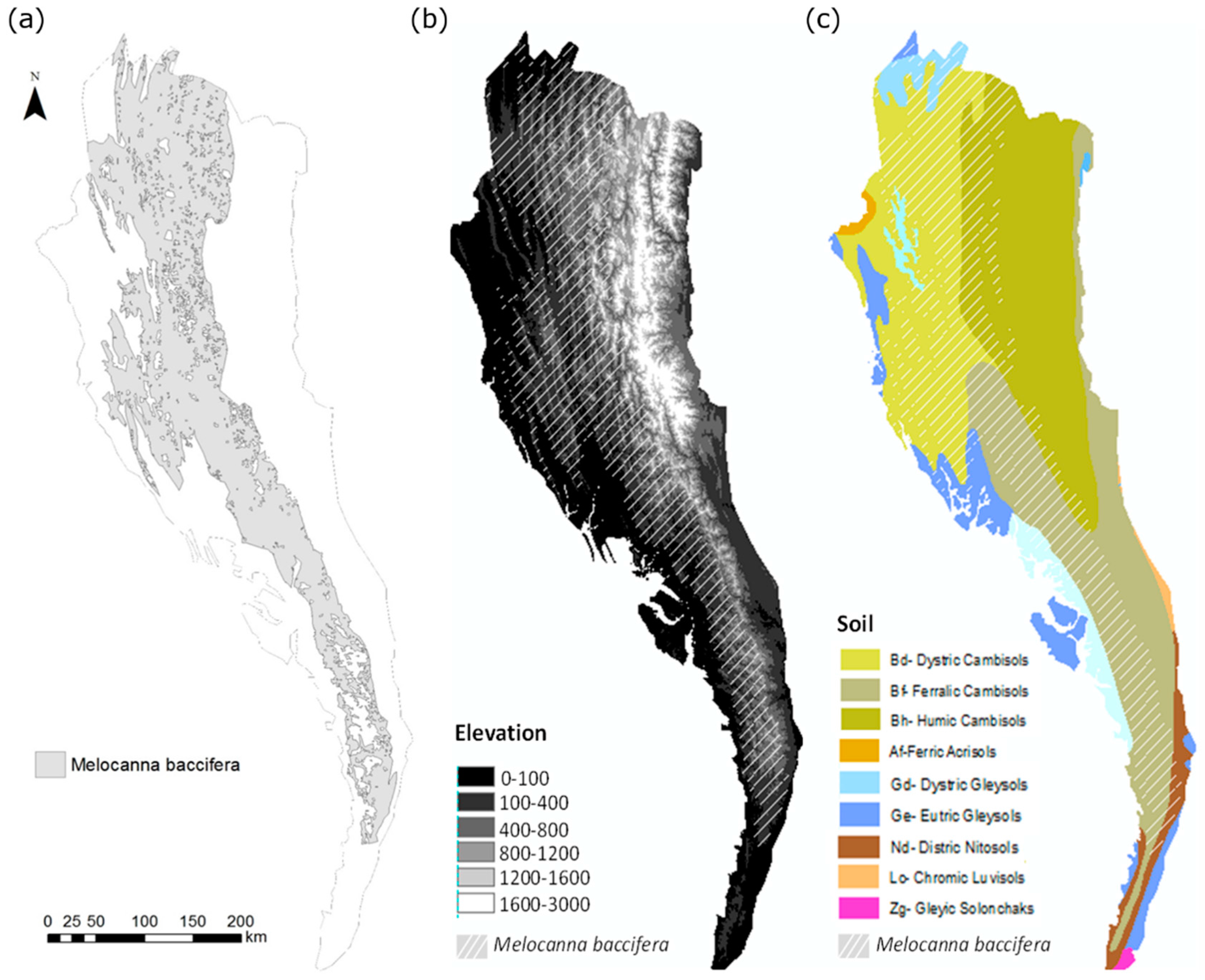
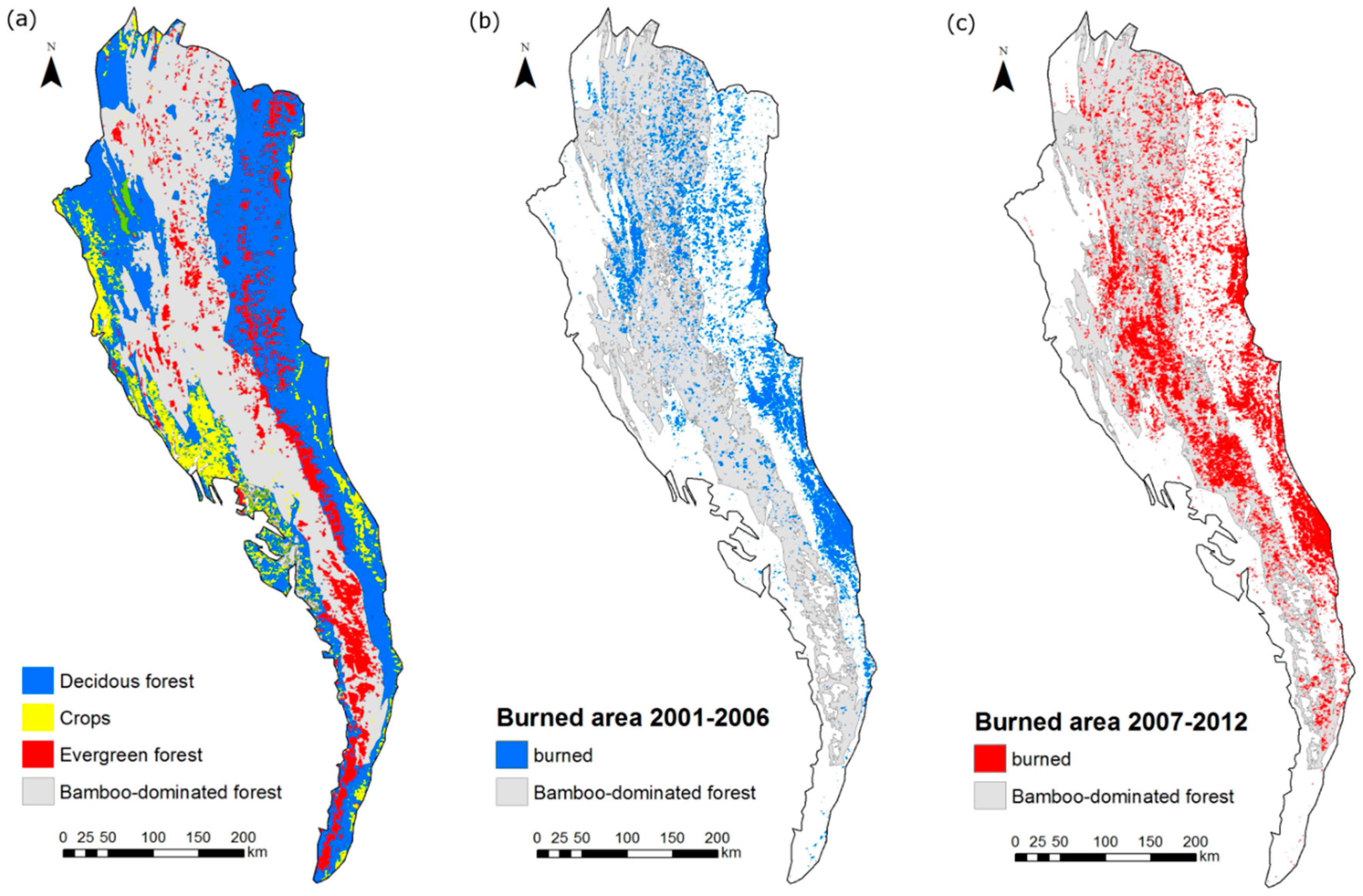
2.2. Bamboo Forest Mapping
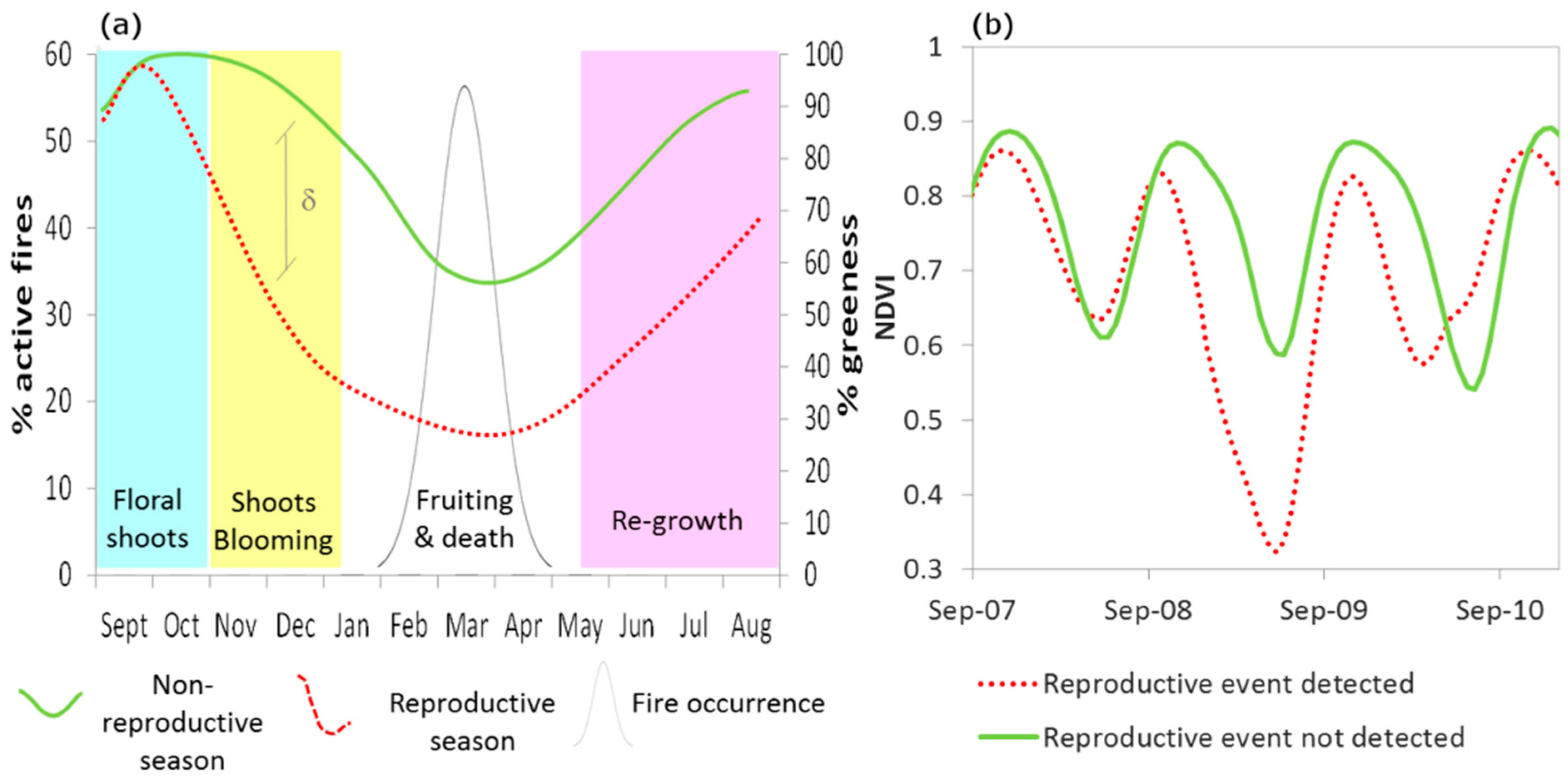
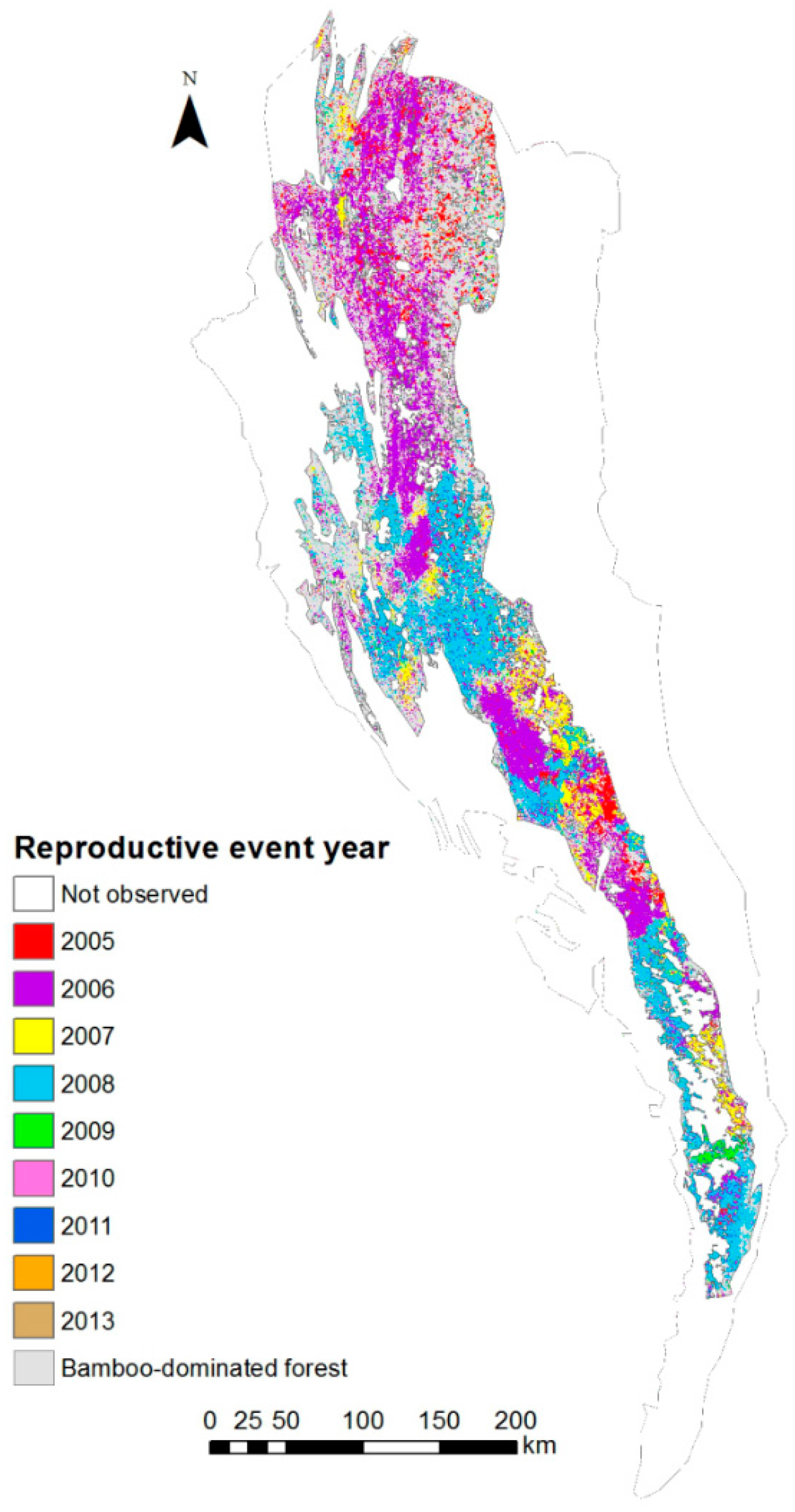
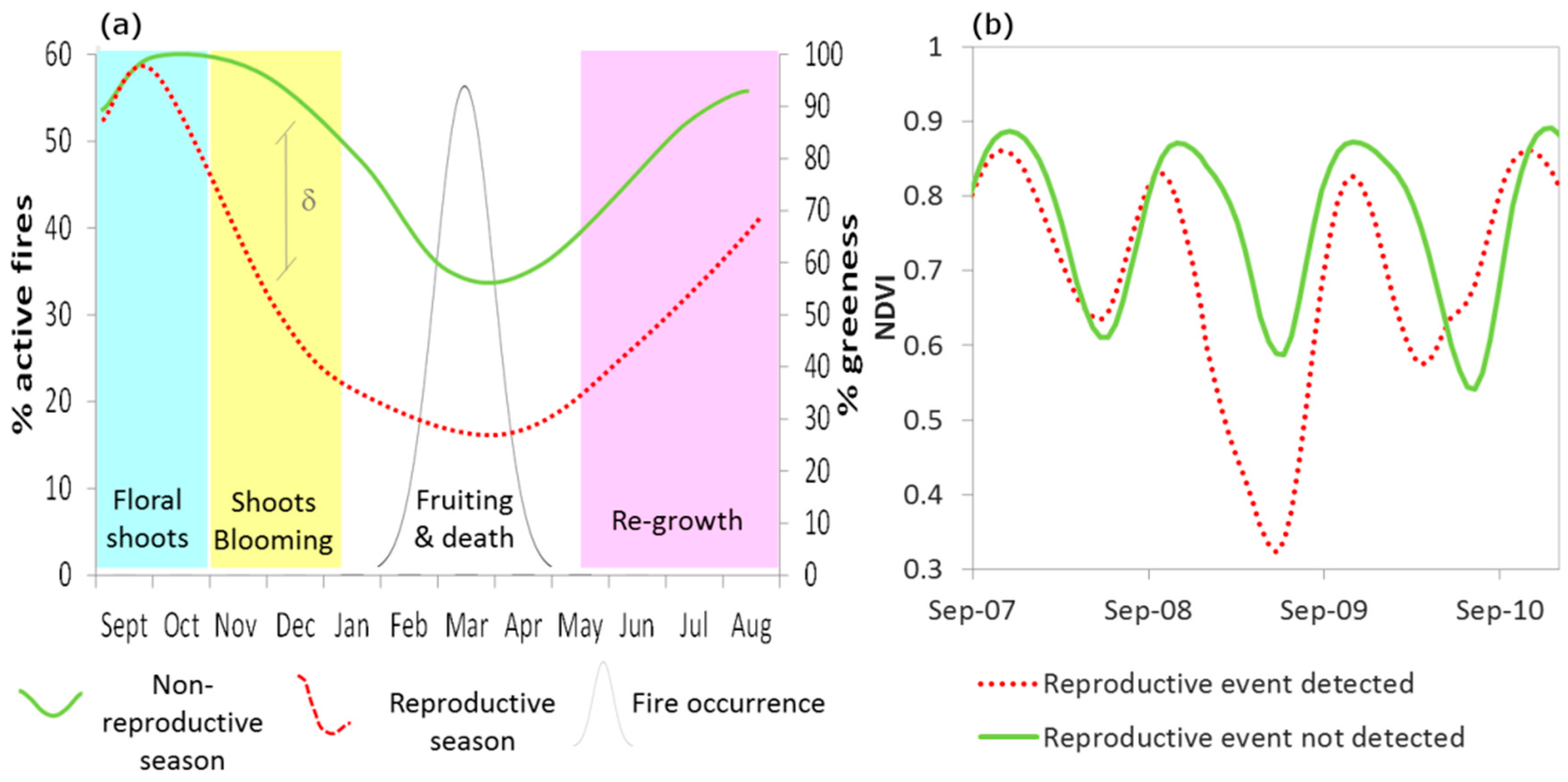
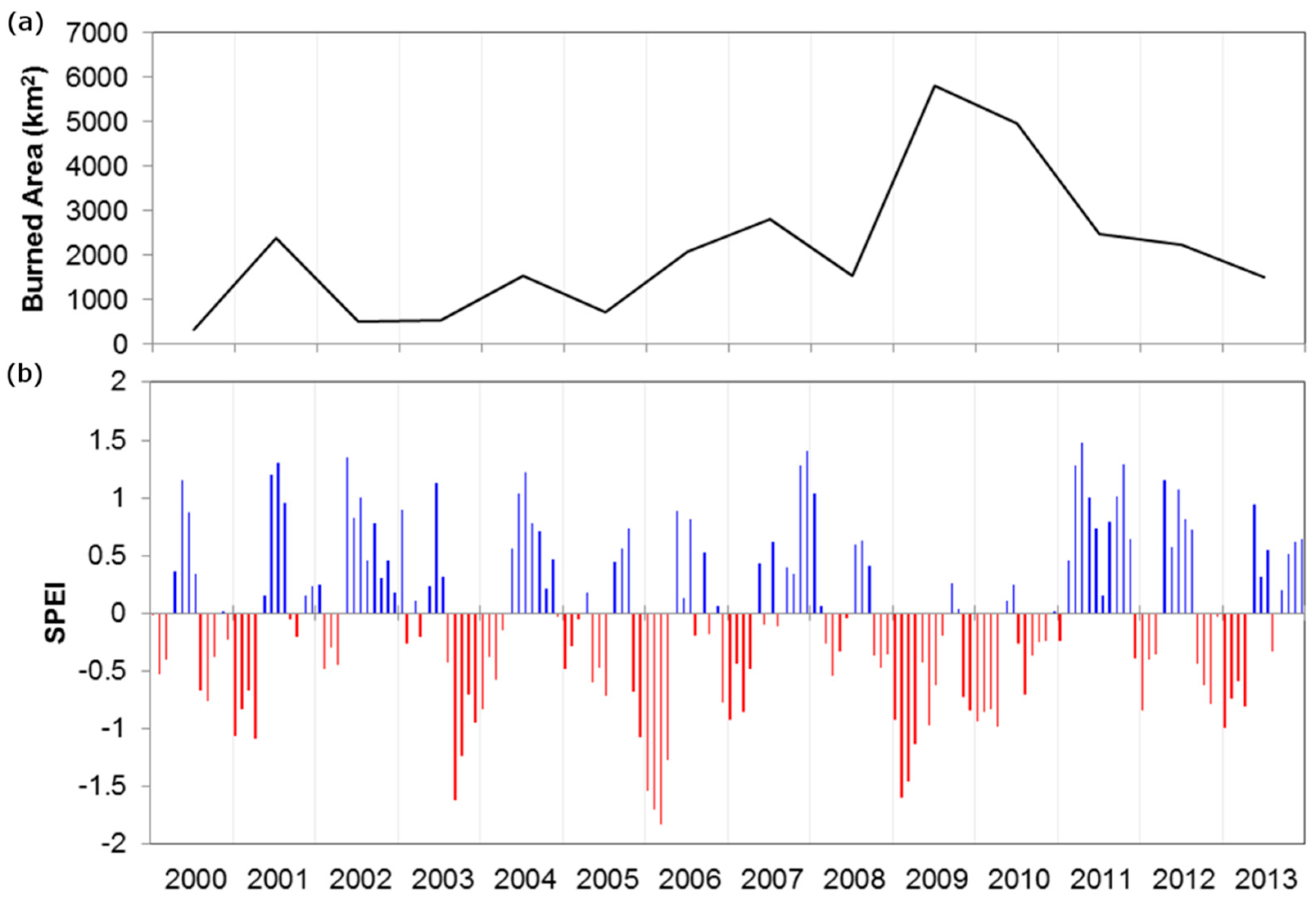
2.3. Bamboo Reproductive Event Detection
- (i)
- Computation of November-December mean NDVI (NDVIND).
- (ii)
- Starting from the year 2005 (as no bamboo flowering was reported before 2005 [7,8,9], a t-test was applied to identify statistically significant anomalies (p < 0.001) between NDVIND of the year i and mean and standard deviation of NDVIND for the period 2000—year i − 1. These seasons were marked as ‘reproductive seasons’.
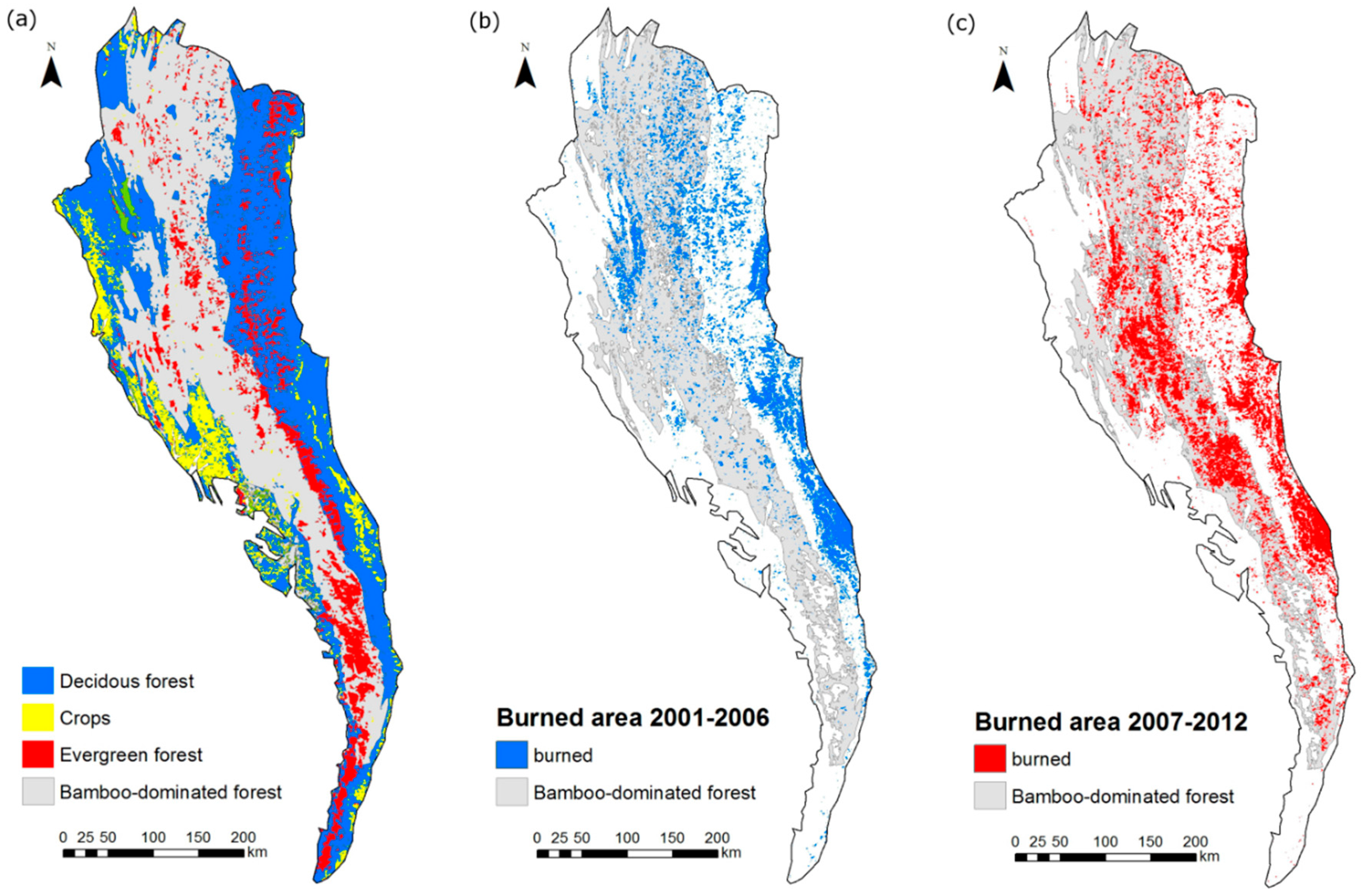
2.4. Evaluation of Wildfire Temporal and Spatial Patterns
3. Results and Discussion
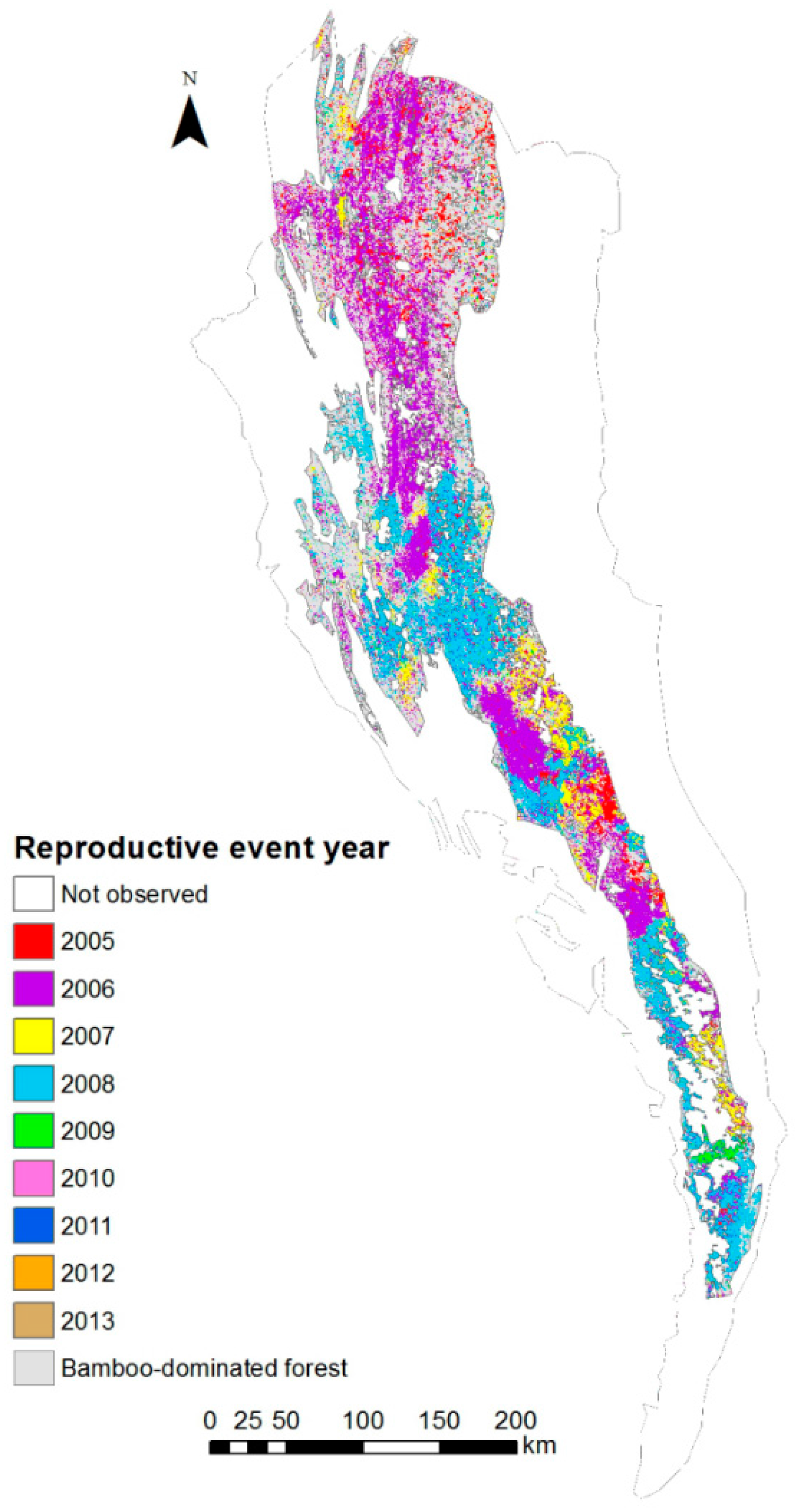
3.1. Bamboo Forest Extent and Reproductive Event Dynamics
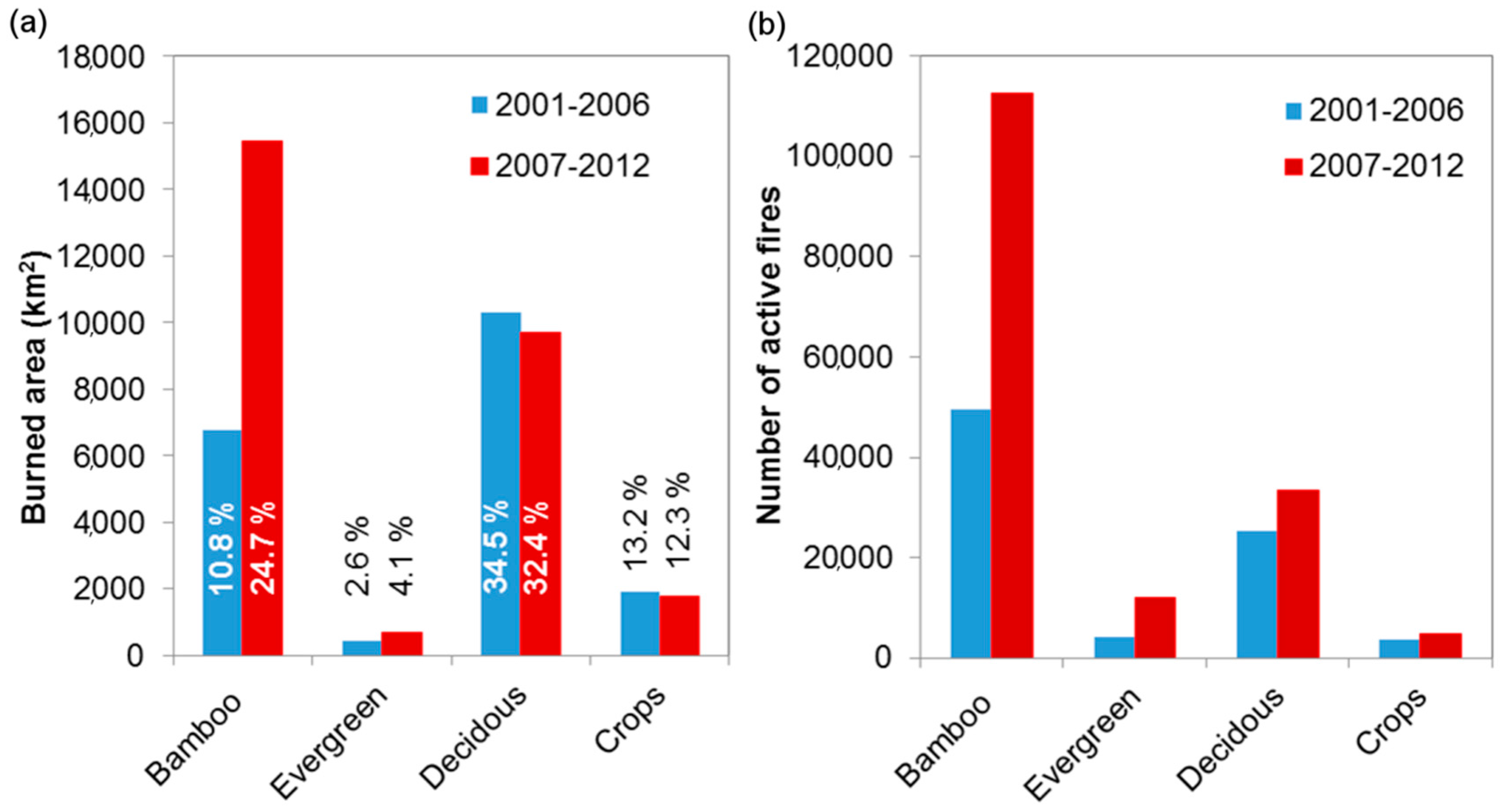
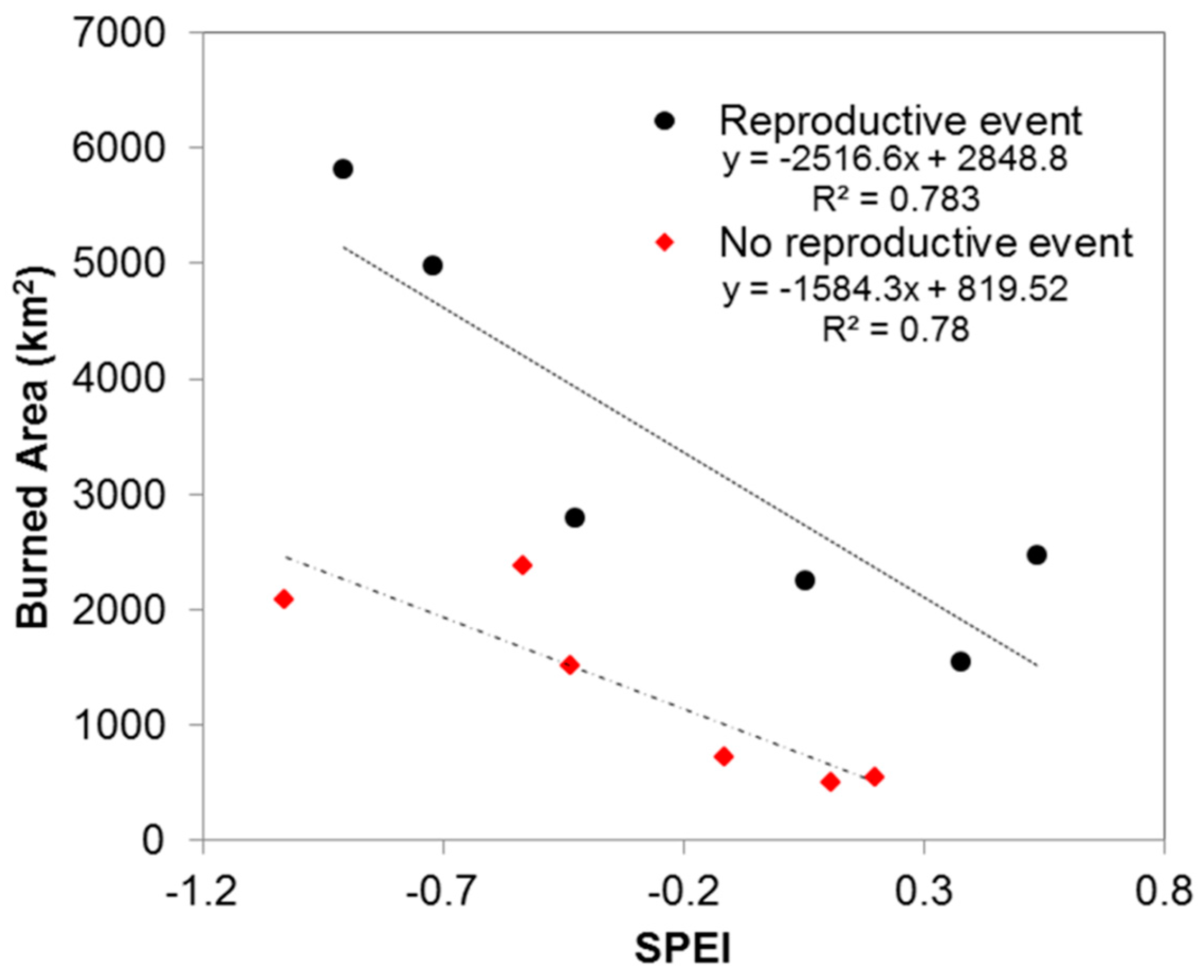
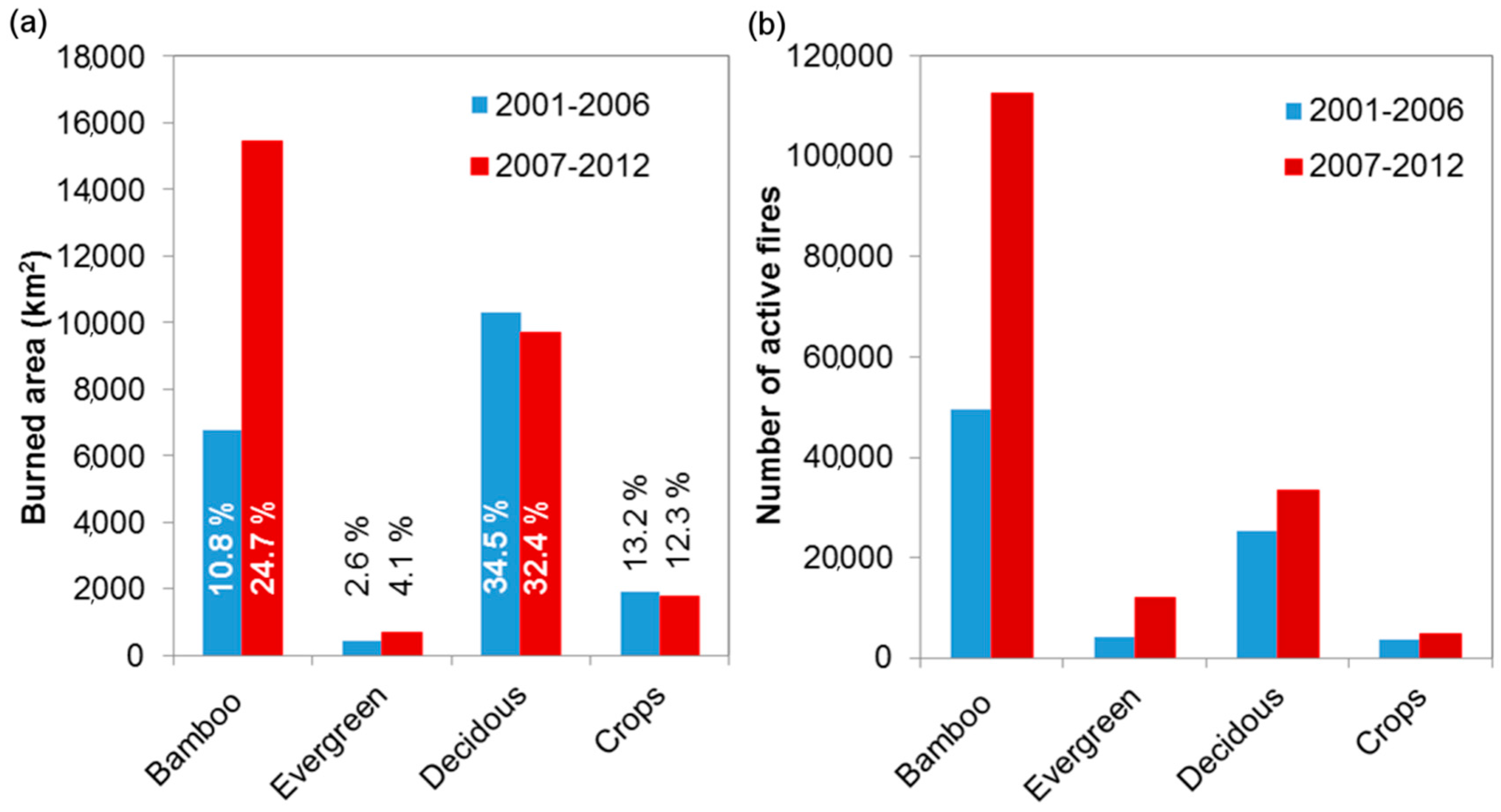
3.2. Analysis of Wildfire Dynamics
3.3. Possible Limitations of the Proposed Approach
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, L.H.; Edwards, K.F.; Byrnes, J.E.; Bastow, J.L.; Wright, A.N.; Spence, K.O. A meta-analysis of resource pulse–consumer interactions. Ecol. Monogr. 2010, 80, 125–151. [Google Scholar] [CrossRef]
- Yang, L.H.; Bastow, J.L.; Spence, K.O.; Wright, A.N. What can we learn from resource pulses? Ecology 2008, 89, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Silvertown, J.W. The evolutionary ecology of mast seeding trees. Biol. J. Linn. Soc. 1980, 14, 235–250. [Google Scholar] [CrossRef]
- Soderstrom, T.R.; Calderon, C.E. A commentary on the bamboos (Poaceae: Bambusoideae). Biotropica 1979, 11, 161–172. [Google Scholar] [CrossRef]
- Janzen, D.H. Why bamboos wait so long to flower? Annu. Rev. Ecol. Syst. 1976, 7, 347–391. [Google Scholar] [CrossRef]
- Blatter, E. The flowering of bamboos. J. Bombay Nat. Hist. Soc. 1930, 34, 135–141. [Google Scholar]
- Normile, D. Holding back a torrent of rats. Science 2010, 327, 806–807. [Google Scholar] [CrossRef] [PubMed]
- Singleton, G.R.; Belmain, S.; Brown, P.R.; Aplin, K.; Htwe, N.M. Impacts of rodent outbreaks on food security in Asia. Wildlife Res. 2010, 37, 355–359. [Google Scholar] [CrossRef]
- Singleton, G.R. Rodent Outbreaks: Ecology and Impacts; International Rice Research Institute: Los Banos, Philippines, 2010. [Google Scholar]
- Rao, M.; Htun, S.; Platt, S.G.; Tizard, R.; Poole, C.; Myint, T.; Watson, J.E.M. Biodiversity conservation in a changing climate: A review of threats and implications for conservation planning in Myanmar. AMBIO 2013, 42, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Bond, W.J. Mast flowering and semelparity in bamboos: The bamboo fire cycle hypothesis. Am. Nat. 1999, 154, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Nelson, B.W. Fire favours expansion of bamboo-dominated forests in the south-west Amazon. J. Trop. Ecol. 2011, 27, 59–64. [Google Scholar] [CrossRef]
- Saha, S.; Howe, H.F. The bamboo fire cycle hypothesis: A comment. Am. Nat. 2001, 158, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Bond, W.J. On incorporating fire into our thinking about natural ecosystems: A response to Saha and Howe. Am. Nat. 2001, 158, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.; Silveira, J.M.; Mestre, L.A.; Andrade, R.B.; D’Andrea, G.C.; Louzada, J.; Vaz-de-Mello, F.Z.; Numata, I.; Lacau, S.; Cochrane, M.A. Wildfires in bamboo-dominated Amazonian forest: Impacts on above-ground biomass and biodiversity. PLoS ONE 2012, 7, e33373. [Google Scholar] [CrossRef]
- Franklin, D.C.; Prior, L.D.; Hogarth, N.J.; McMahon, C.R. Bamboo, fire and flood: Consequences of disturbance for the vegetative growth of a clumping, clonal plant. Plant Ecol. 2010, 208, 319–332. [Google Scholar] [CrossRef]
- McMichael, C.H.; Bush, M.B.; Silman, M.R.; Piperno, D.R.; Raczka, M.; Lobato, L.C.; Zimmerman, M.; Hagen, S.; Palace, M. Historical fire and bamboo dynamics in western Amazonia. J. Biogeogr. 2013, 40, 299–309. [Google Scholar] [CrossRef]
- De Carvalho, A.L.; Nelson, B.W.; Bianchini, M.C.; Plagnol, D.; Kuplich, T.M.; Daly, D.C. Bamboo-dominated forests of the southwest Amazon: Detection, spatial extent, life cycle length and flowering waves. PLoS ONE 2013, 8, e54852. [Google Scholar] [CrossRef] [PubMed]
- Mouillot, F.; Schultz, M.G.; Yue, C.; Cadule, P.; Tansey, K.; Ciais, P.; Chuvieco, E. Ten years of global burned area products from spaceborne remote sensing—A review: Analysis of user needs and recommendations for future developments. Int. J. Appl. Earth Obs. 2014, 26, 64–79. [Google Scholar] [CrossRef]
- Giglio, L.; Loboda, T.; Roy, D.P.; Quayle, B.; Justice, C.O. An active-fire based burned area mapping algorithm for the MODIS sensor. Remote Sens. Environ. 2009, 113, 408–420. [Google Scholar] [CrossRef]
- Stibig, H.J.; Achard, F.; Fritz, S. A new forest cover map of continental Southeast Asia derived from SPOT-VEGETATION satellite imagery. Appl. Veg. Sci. 2004, 7, 153–162. [Google Scholar] [CrossRef]
- Stibig, H.J.; Belward, A.S.; Roy, P.S.; Rosalina-Wasrin, U.; Agrawal, S.; Joshi, P.K.; Beuchle, R.; Fritz, S.; Mubareka, S.; Giri, C. A land-cover map for South and Southeast Asia derived from SPOT-VEGETATION data. J. Biogeogr. 2007, 34, 625–637. [Google Scholar] [CrossRef]
- Giri, C.; Defourny, P.; Shrestha, S. Land cover characterization and mapping of continental Southeast Asia using multi-resolution satellite sensor data. Int. J. Remote Sens. 2003, 24, 4181–4196. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Global Forest Resources Assessment 2015. Available online: http://www.fao.org/forest-resources-assessment/en/ (accessed on 20 July 2016).
- Giri, C.; Pradhan, D.; Ofren, R.S.; Kratzschmar, E. Land Use/Land Cover Change in South and South-East Asia; UNEP Environment Assessment Programme for Asia and the Pacific: Bangkok, Thailand, 1998. [Google Scholar]
- Rao, A.N.; Ramanatha Rao, V. Bamboo-Conservation, diversity, ecogeography, germplasm, resource utilization and taxonomy. In Proceedings of the Training Course Cum Workshop, Kunming and Xishuanbanna, Yunnan, China, 10–17 May 1998.
- Landsat Orthorectified Dataset (January 2005–December 2011). The Global Land Cover Facility 2005. Available online: http://www.glovis.org (accessed on 31 December 2015).
- Bystriakova, N.; Kapos, V.; Stapleton, C.M.A.; Lysenko, I. Bamboo Biodiversity; UNEP-WCMC: Cambridge, UK, 2003. [Google Scholar]
- Bystriakova, N.; Kapos, V.; Lysenko, I.; Stapleton, C.M.A. Distribution and conservation status of forest bamboo biodiversity in the Asia-Pacific Region. Biodivers. Conserv. 2003, 12, 1833–1841. [Google Scholar] [CrossRef]
- Vuolo, F.; Mattiuzzi, M.; Klisch, A.; Atzberger, C. Data service platform for MODIS vegetation indices time series processing at BOKU Vienna: Current status and future perspectives. Proc. SPIE 2012. [Google Scholar] [CrossRef]
- Atzberger, C.; Eilers, P.H.C. Evaluating the effectiveness of smoothing algorithms in the absence of ground reference measurements. Int. J. Remote Sens. 2011, 32, 3689–3709. [Google Scholar] [CrossRef]
- Atkinson, P.M.; Jeganathan, C.; Dash, J.; Atzberger, C. Intercomparison of four models for smoothing satellite sensor time-series data to estimate vegetation phenology. Remote Sens. Environ. 2012, 123, 400–417. [Google Scholar] [CrossRef]
- Eilers, P.H.C. A perfect smoother. Anal. Chem. 2003, 75, 3631–3636. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.P.; Jin, Y.; Lewis, P.E.; Justice, C.O. Prototyping a global algorithm for systematic fire-affected area mapping using MODIS time series data. Remote Sens. Environ. 2005, 97, 137–162. [Google Scholar] [CrossRef]
- Giglio, L.; Descloitres, J.; Justice, C.O.; Kaufman, Y.J. An enhanced contextual fire detection algorithm for MODIS. Remote Sens. Environ. 2003, 87, 273–282. [Google Scholar] [CrossRef]
- FAO. Agricultural Atlas of the Union of Myanmar-Agricultural Year 2001–2002. Available online: http://dwms.fao.org/atlases/myanmar/index_en.htm (accessed on 20 July 2016).
- Attri, S.D.; Tyagi, A. Climate profile of India. Met. Monogr. Environ. Meteorol. 2010, 1, 1–122. [Google Scholar]
- FAO. Digital Soil Map of the World 2007. Available online: http://www.fao.org/geonetwork/srv/en/metadata.show?id=14116 (accessed on 10 July 2016).
- Dudal, R. Soils of Southeast Asia. In The Physical Geography of Southeast Asia, 1st ed.; Gupta, A., Ed.; Oxford University Press: Oxford, UK, 2005; pp. 94–104. [Google Scholar]
- Franklin, D.C. Synchrony and asynchrony: Observations and hypotheses for the flowering wave in a long-lived semelparous bamboo. J. Biogeogr. 2004, 31, 773–786. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multi-scalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Planning & Programme Implementation Department, Government of Mizoram. Government of Mizoram Comprehensive Action Plan for Bamboo Flowering and Famine Combat Schemes (BAFFACOS) (2004–2005 to 2008–2009). Available online: http://mizoram.nic.in/dept/planning/baffacos.htm (accessed on 20 July 2016).
- United Nations Development Program, Bangladesh. Scientific Report on Bamboo Flowering, Rodent Outbreaks and Food Security: Rodent Ecology, Pest Management, and Socio-Economic Impact in the Chittagong Hill Tracts. 2008. Available online: http://projects.nri.org/bandicoot/docs/bamboo.pdf (accessed on 20 July 2016).
- Platt, S.G.; Ko, W.K.; Myo, K.M.; Khaing, L.L.; Platt, K.; Maung, A.; Rainwater, T.R. Notes on Melocanna baccifera and bamboo brakes in the Rakhine hills of western Myanmar. Bamboo Sci. Cult. 2010, 23, 1–12. [Google Scholar]
- Rayle, C.M.; Franklin, S.B.; Lu, Z. Fire occurrence in relation to bamboo dominance in the Qinling Mountains of China: Evidence from phytolith and charcoal records in Holocene sediments. Sci. China Earth Sci. 2015, 58, 1182–1190. [Google Scholar] [CrossRef]
- Linderman, M.; Liu, J.; Qi, J.; An, L.; Ouyang, Z.; Yang, J.; Tan, Y. Using artificial neural networks to map the spatial distribution of understorey bamboo from remote sensing data. Int. J. Remote Sens. 2004, 25, 1685–1700. [Google Scholar] [CrossRef]
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fava, F.; Colombo, R. Remote Sensing-Based Assessment of the 2005–2011 Bamboo Reproductive Event in the Arakan Mountain Range and Its Relation with Wildfires. Remote Sens. 2017, 9, 85. https://doi.org/10.3390/rs9010085
Fava F, Colombo R. Remote Sensing-Based Assessment of the 2005–2011 Bamboo Reproductive Event in the Arakan Mountain Range and Its Relation with Wildfires. Remote Sensing. 2017; 9(1):85. https://doi.org/10.3390/rs9010085
Chicago/Turabian StyleFava, Francesco, and Roberto Colombo. 2017. "Remote Sensing-Based Assessment of the 2005–2011 Bamboo Reproductive Event in the Arakan Mountain Range and Its Relation with Wildfires" Remote Sensing 9, no. 1: 85. https://doi.org/10.3390/rs9010085
APA StyleFava, F., & Colombo, R. (2017). Remote Sensing-Based Assessment of the 2005–2011 Bamboo Reproductive Event in the Arakan Mountain Range and Its Relation with Wildfires. Remote Sensing, 9(1), 85. https://doi.org/10.3390/rs9010085