

Spatiotemporal Variation in Carbon and Water Use Efficiency and Their Influencing Variables Based on Remote Sensing Data in the Nanling Mountains Region
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Acquisition
2.3. Methods
2.3.1. Estimation of CUE and WUE Values
2.3.2. Trends Analysis
2.3.3. Hurst Exponent Analysis
2.3.4. Stability Analysis
2.3.5. Partial Correlation Analysis
3. Results
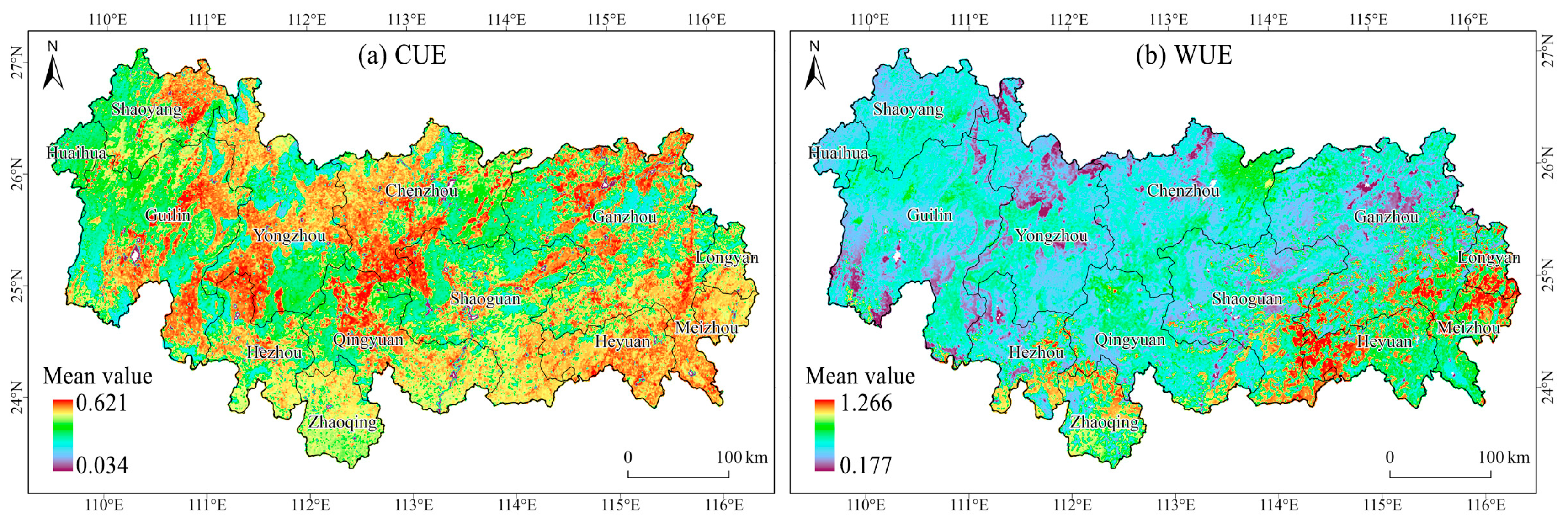
3.1. Spatial and Temporal Characteristics of Vegetation CUE and WUE
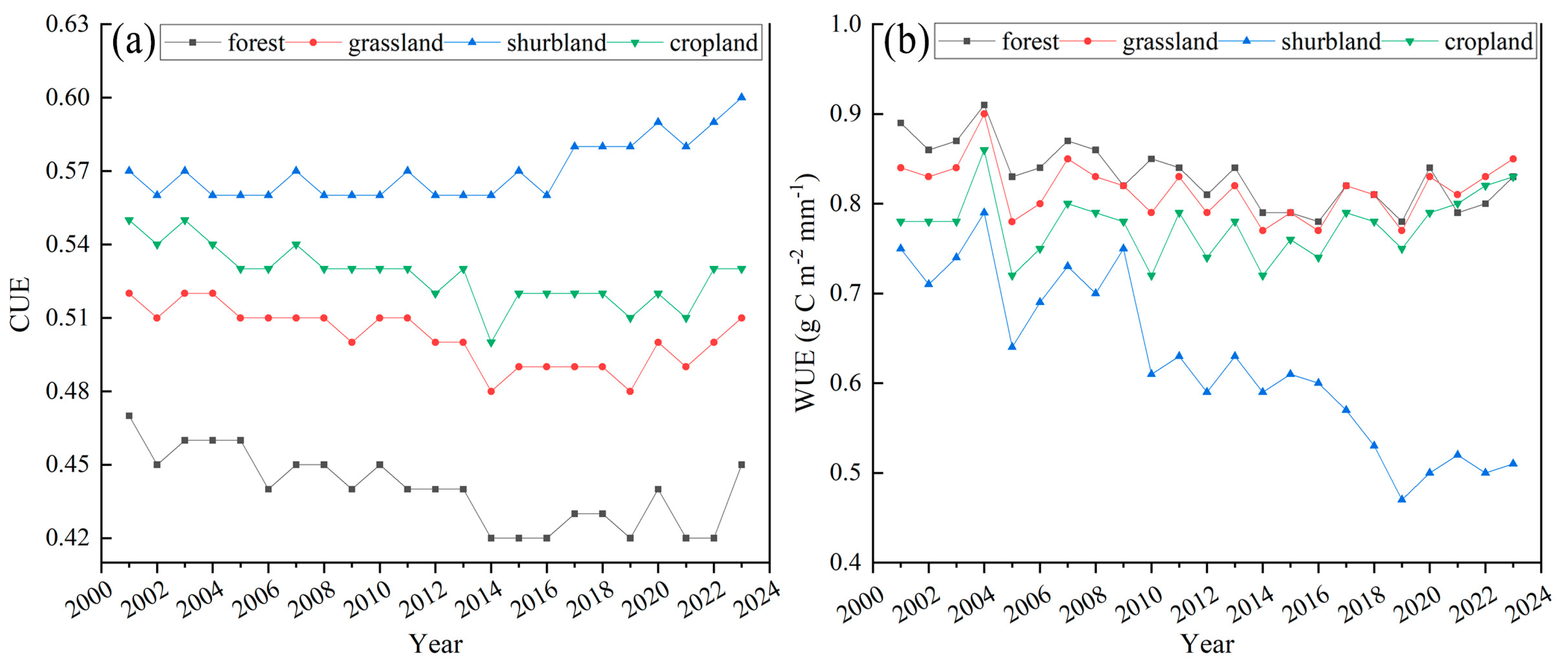
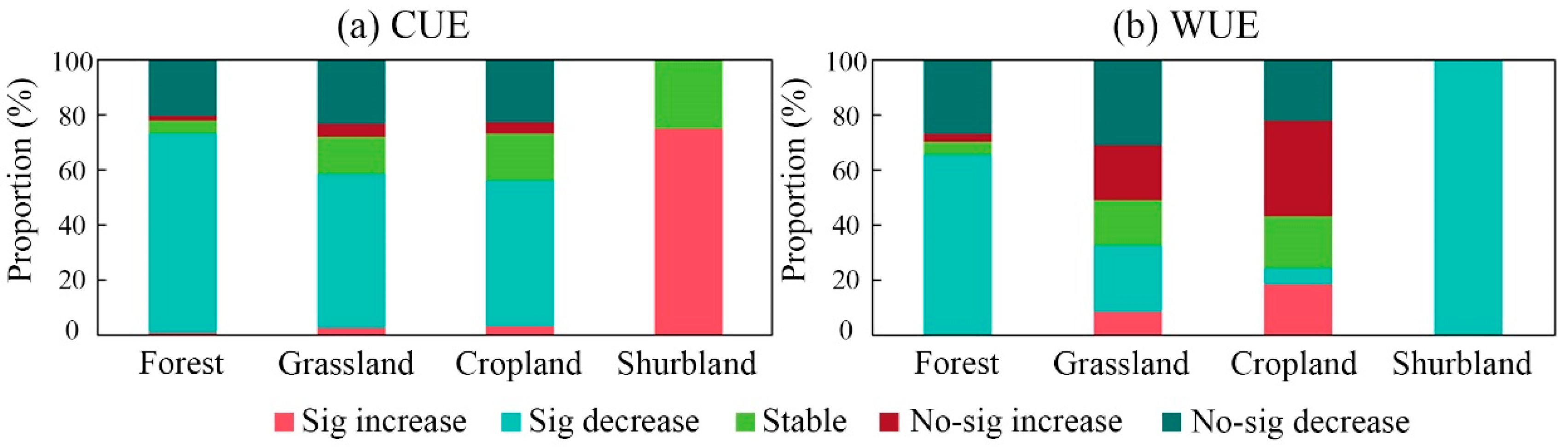
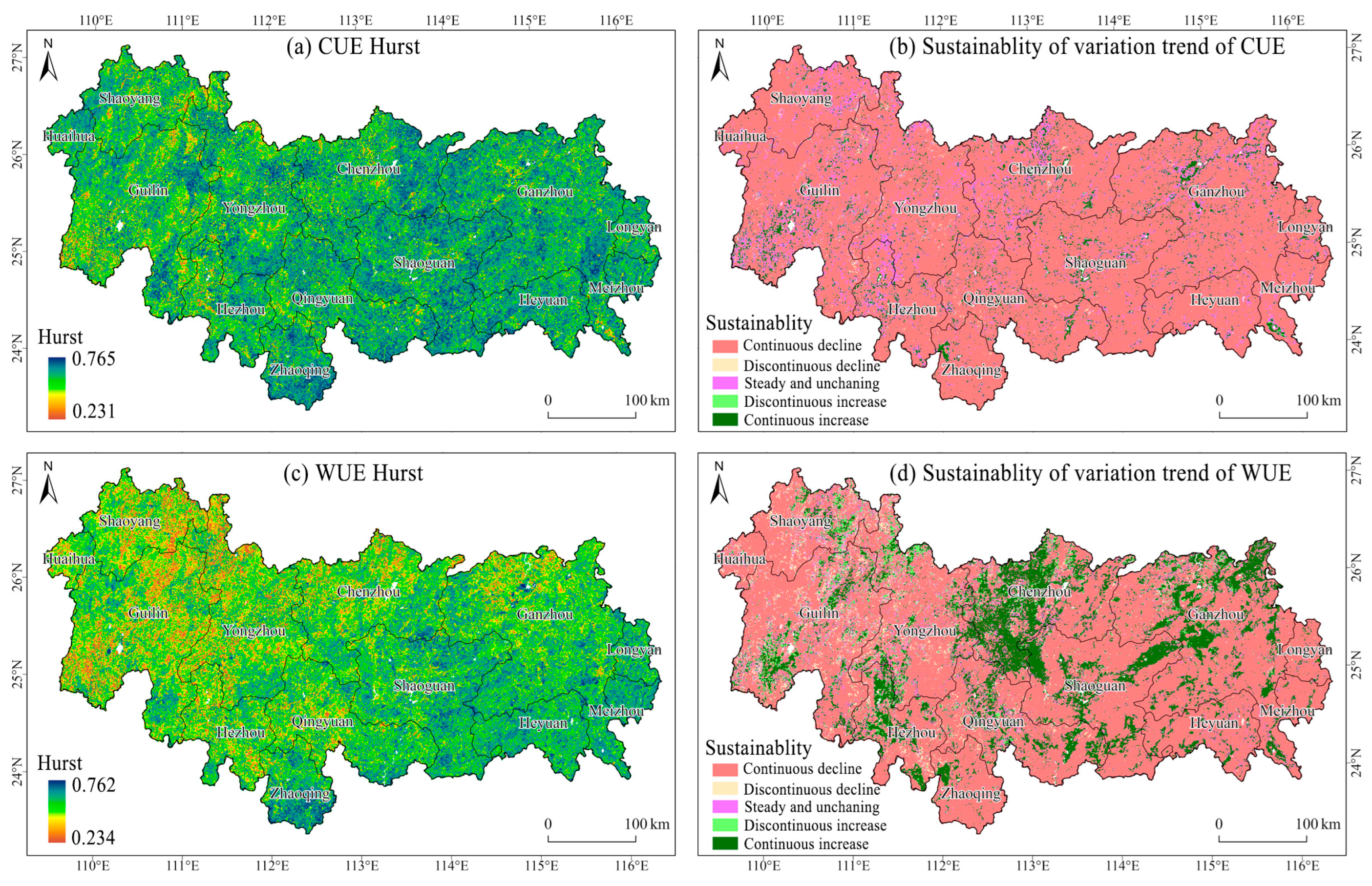
3.2. Analysis of Vegetation CUE and WUE Trends in the NMR
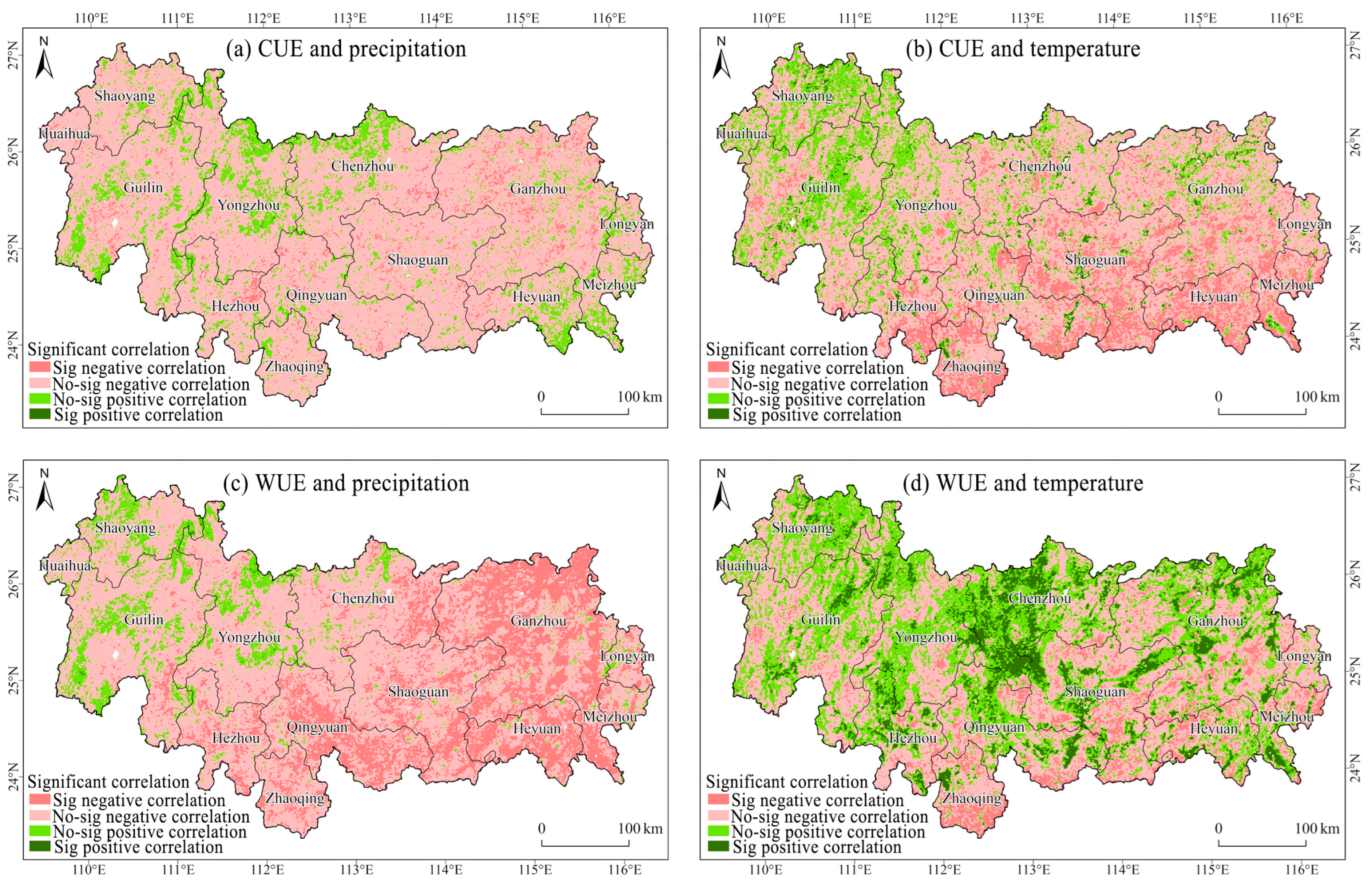
3.3. Response of Vegetation CUE and WUE to Changes in Precipitation and Temperature in the NMR
4. Discussion
4.1. Spatial and Temporal Dynamic and Distribution of CUE and WUE in NMR
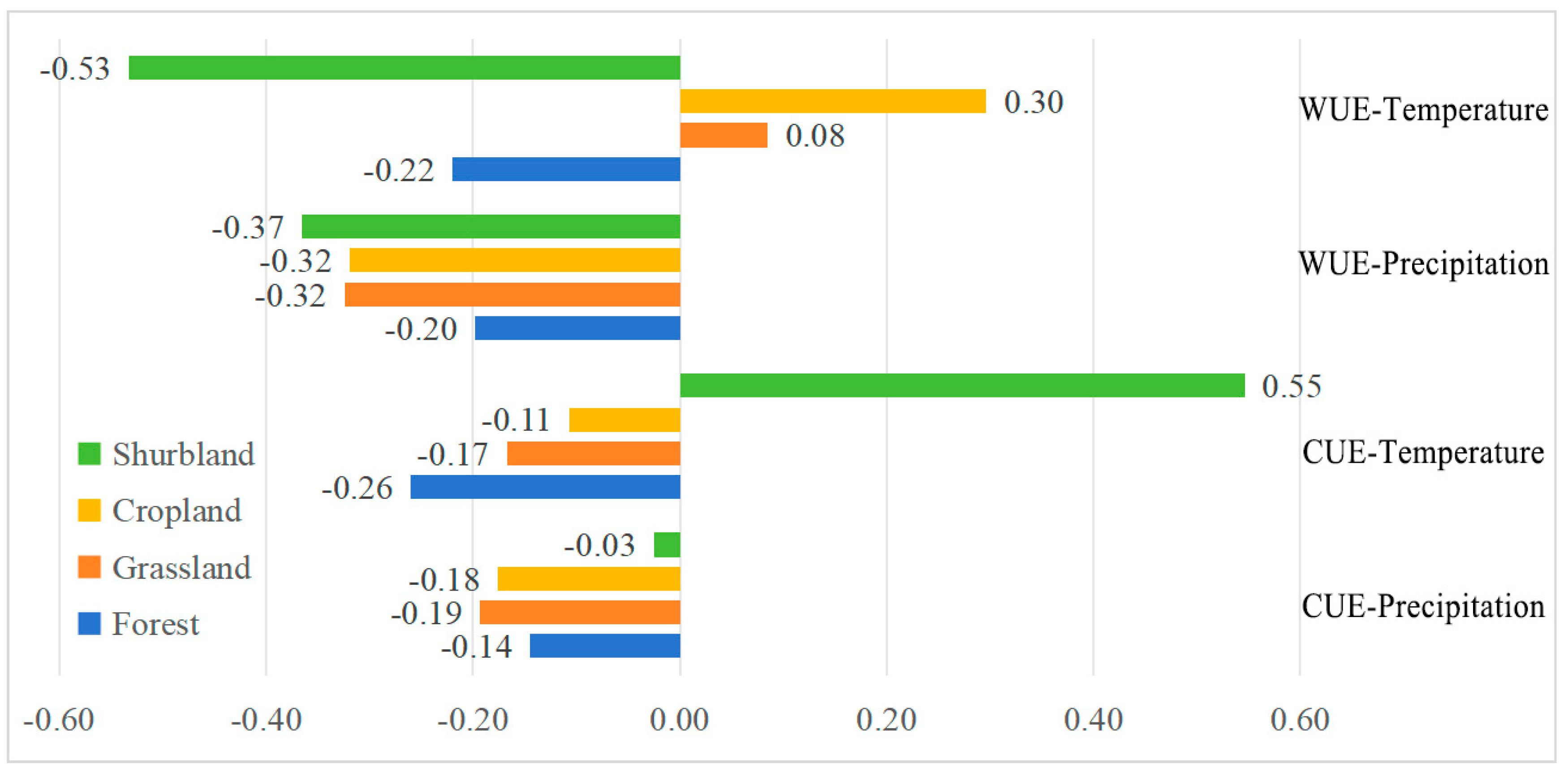
4.2. Influencing Factors of CUE and WUE Variation in the NMR
4.3. Uncertainty and Limitation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mystakidis, S.; Davin, E.L.; Gruber, N.; Seneviratne, S.I. Constraining future terrestrial carbon cycle projections using observation-based water and carbon flux estimates. Glob. Change Biol. 2016, 22, 2198–2215. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Fang, J.; Ciais, P.; Peylin, P.; Huang, Y.; Sitch, S.; Wang, T. The carbon balance of terrestrial ecosystems in China. Nature 2009, 458, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Ding, J.; Welp, M.; Huang, S.; Liu, B. Using MODIS data to analyse the ecosystem water use efficiency spatial-temporal variations across Central Asia from 2000 to 2014. Environ. Res. 2020, 182, 108985. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef]
- Umair, M.; Kim, D.; Choi, M. Impact of climate, rising atmospheric carbon dioxide, and other environmental factors on water-use efficiency at multiple land cover types. Sci. Rep. 2020, 10, 11644. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Bai, X.; Zhao, C.; Tan, Q.; Li, Y.; Luo, G.; Wu, L.; Chen, F.; Li, C.; Ran, C.; et al. Responses of carbon and water use efficiencies to climate and land use changes in China’s karst areas. J. Hydrol. 2023, 617, 128968. [Google Scholar] [CrossRef]
- Ye, X.; Liu, F.; Zhang, Z.; Xu, C.; Liu, J. Spatio-temporal variations of vegetation carbon use efficiency and potential driving meteorological factors in the Yangtze River Basin. J. Mt. Sci. 2020, 17, 1959–1973. [Google Scholar] [CrossRef]
- Li, G.; Chen, W.; Li, R.; Zhang, X.; Liu, J. Assessing the spatiotemporal dynamics of ecosystem water use efficiency across China and the response to natural and human activities. Ecol. Indic. 2021, 126, 107680. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, X.; Tian, H.; Wu, X.; Gao, Z.; Feng, Y.; Piao, S.; Lv, N.; Pan, N.; Fu, B. Accelerated increase in vegetation carbon sequestration in China after 2010: A turning point resulting from climate and human interaction. Glob. Change Biol. 2021, 27, 5848–5864. [Google Scholar] [CrossRef]
- Yu, L.; Josey, S.A.; Bingham, F.M.; Lee, T. Intensification of the global water cycle and evidence from ocean salinity: A synthesis review. Ann. N. Y. Acad. Sci. 2020, 1472, 76–94. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Ullah, S.; Yan, L.; Lin, Y. Remote sensing of ecosystem water use efficiency: A review of direct and indirect estimation methods. Remote Sens. 2021, 13, 2393. [Google Scholar] [CrossRef]
- Jiang, S.; Liang, C.; Cui, N.; Zhao, L.; Liu, C.; Feng, Y.; Hu, X.; Gong, D.; Zou, Q. Water use efficiency and its drivers in four typical agroecosystems based on flux tower measurements. Agric. For. Meteorol. 2020, 295, 108200. [Google Scholar] [CrossRef]
- Bai, Y.; Li, X.; Zhou, S.; Yang, X.; Yu, K.; Wang, M.; Liu, S.; Wang, P.; Wu, X.; Wang, X.; et al. Quantifying plant transpiration and canopy conductance using eddy flux data: An underlying water use efficiency method. Agric. For. Meteorol. 2019, 271, 375–384. [Google Scholar] [CrossRef]
- Gago, J.; Douthe, C.; Florez-Sarasa, I.; Escalona, J.M.; Galmes, J.; Fernie, A.R.; Flexas, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lu, J.; Xu, X.; Ren, X.; Wang, J.; Chai, X.; Wang, W. Spatial and Temporal Patterns of Carbon and Water Use Efficiency on the Loess Plateau and Their Influencing Factors. Land 2020, 12, 77. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Yang, Y.; Tong, L.; Li, J. Spatiotemporal Dynamic of Vegetation Carbon Use Efficiency and Its Relationship with Climate Factors in China During the Period 2000–2013. Res. Soil Water Conserv. 2019, 26, 278–286. [Google Scholar]
- Qin, S.; Ding, J.; Ge, X.; Wang, J.; Wang, R.; Zou, J.; Tan, J.; Han, L. Spatio-temporal changes in water use efficiency and its driving factors in Central Asia (2001–2021). Remote Sens. 2023, 15, 767. [Google Scholar] [CrossRef]
- Yuan, M.; Li, M.; Cheng, H.; Ding, J.; Li, H. Future trends in carbon use efficiency for Chinese terrestrial ecosystem based on CMIP5 model results. J. Univ. Chin. Acad. Sci. 2017, 34, 452–461. [Google Scholar]
- Liu, F.; Ye, X.; Guo, Q.; Li, X.; Liu, J. Spatio-temporal variation of carbon and water use efficiency of different land cover in the Poyang Lake Basin and their correlations with climate factors. Acta Ecol. Sin. 2021, 41, 694–706. [Google Scholar]
- Liu, X.; Hu, B.; Ren, Z. Spatiotemporal variation of water use efficiency and its driving forces on the Loess Plateau during 2000–2014. Chin. Agric. Sci. 2018, 51, 302–314. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, M.; Chen, H.; Adams, J. Global pattern of NPP to GPP ratio derived from MODIS data: Effects of ecosystem type, geographical location and climate. Glob. Ecol. Biogeogr. 2009, 18, 280–290. [Google Scholar] [CrossRef]
- Yin, C.; Luo, M.; Meng, F.; Sa, L.; Bao, Y. The spatiotemporal variation and influencing factors of vegetation carbon and water use efficiency in the Mongolian Plateau. Chin. J. Ecol. 2022, 41, 1079–1089. [Google Scholar]
- Jiang, L.; Wu, Y.; He, X.; Fu, Q.; Wang, Z.; Jiang, Q. Dynamic simulation and coupling coordination evaluation of water footprint sustainability system in Heilongjiang province, China: A combined system dynamics and coupled coordination degree model. J. Clean. Prod. 2022, 380, 135044. [Google Scholar] [CrossRef]
- Lan, Y.; Cao, S.; Cao, G.; Li, X.; Yang, Y.; Lei, Y. Temporal and spatial dynamics of vegetation carbon use efficiency in Qinghai Lake Basin. Ecol. Sci. 2020, 39, 156–166. [Google Scholar]
- Zhao, J.; Feng, H.; Xu, T.; Xiao, J.; Guerrieri, R.; Liu, S.; Wu, X.; He, X.; He, X. Physiological and environmental control on ecosystem water use efficiency in response to drought across the northern hemisphere. Sci. Total Environ. 2021, 758, 143599. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, B.; Davis, M.; Sardans, J.; Peñuelas, J.; Billings, S. Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New Phytol. 2016, 210, 431–442. [Google Scholar] [CrossRef]
- Naeem, S.; Zhang, Y.; Zhang, X.; Rehman, A.U.; Tang, Z.; Xu, Z.; Li, C.; Azeem, T. Recent change in ecosystem water use efficiency in China mainly dominated by vegetation greening and increased CO2. Remote Sens. Environ. 2023, 298, 113811. [Google Scholar] [CrossRef]
- Du, L.; Zeng, Y.; Ma, L.; Qiao, C.; Wu, H.; Su, Z.; Bao, G. Effects of anthropogenic revegetation on the water and carbon cycles of a desert steppe ecosystem. Agric. For. Meteorol. 2021, 300, 108339. [Google Scholar] [CrossRef]
- Wu, D.; Zou, C.; Gao, J.; Lin, N. Ecological status changes of water conservation key ecological function areas in China. Environ. Sci. Technol. 2017, 40, 174–179. [Google Scholar]
- Zhou, G.; Zhang, H.; Zhou, P. Multi-disciplinary research values of the Nanling Mountains. Trop. Geogr. 2018, 38, 293–298. [Google Scholar]
- Duan, H.; Cao, F. Characteristics and trends of climate change of Chinese subtropical Nanling Mountain. J. Cent. South Univ. For. Technol. 2012, 32, 110–113. [Google Scholar]
- Lin, D.; Zhou, P.; Xu, W.; Li, J.; Lin, W. Carbon budget of different vegetation types in Nanling based on IBIS. Acta Ecol. Sin. 2024, 44, 1429–1440. [Google Scholar]
- Baeza, S.; Paruelo, J.M. Land use/land cover change (2000–2014) in the Rio de la Plata grasslands: An analysis based on MODIS NDVI time series. Remote Sens. 2020, 12, 381. [Google Scholar] [CrossRef]
- Zhao, H.; Li, X.; Zhang, D.; Xiao, R. Aboveground biomass in grasslands in Qinghai Province estimated from MODIS data and its influencing factors. Acta Prataculturae Sin. 2020, 29, 5. [Google Scholar]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Xu, L.; Meng, P.; Tong, X.; Zhang, J.; Li, J.; Wang, X.; Xie, H.; Liu, P. Productivity and water use efficiency of Pinus tabulaeformis responses to climate change in the temperate monsoon region. Agric. For. Meteorol. 2022, 327, 109188. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, C.; Chen, H.; Li, L.; Xu, H.; Li, H.; Jain, S.K. Trend and concentration characteristics of precipitation and related climatic teleconnections from 1982 to 2010 in the Beas River basin, India. Glob. Planet. Change 2016, 145, 116–129. [Google Scholar] [CrossRef]
- Peng, J.; Liu, Z.; Liu, Y.; Wu, J.; Han, Y. Trend analysis of vegetation dynamics in Qinghai–Tibet Plateau using Hurst Exponent. Ecol. Indic. 2012, 14, 28–39. [Google Scholar] [CrossRef]
- Granero, M.S.; Segovia, J.T.; Pérez, J.G. Some comments on Hurst exponent and the long memory processes on capital markets. Phys. A Stat. Mech. Its Appl. 2008, 387, 5543–5551. [Google Scholar] [CrossRef]
- Hurst, H.E. Long-term storage capacity of reservoirs. Trans. Am. Soc. Civ. Eng. 1951, 116, 770–799. [Google Scholar] [CrossRef]
- Mandelbrot, B.B.; Wallis, J.R. Robustness of the rescaled range R/S in the measurement of noncyclic long run statistical dependence. Water Resour. Res. 1969, 5, 967–988. [Google Scholar] [CrossRef]
- Lin, Z.; Wen, Z.; Liu, Y.; Yao, H.; Zhou, R.; Ren, H.; Yuan, L. Spatiotemporal variation and attribution of carbon and water use efficiency in the Yellow River Basin based on remote sensing data. Acta Ecol. Sin. 2024, 44, 377–391. [Google Scholar]
- Chen, Z.; Yu, G.; Wang, Q. Ecosystem carbon use efficiency in China: Variation and influence factors. Ecol. Indic. 2018, 90, 316–323. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Li, J.; Gang, C.; Zhang, Y.; Odeh, I.; Qi, J. Assessing the spatiotemporal dynamic of global grassland carbon use efficiency in response to climate change from 2000 to 2013. Acta Oecol. 2017, 81, 22–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, G.; Yang, J.; Wimberly, M.C.; Zhang, X.; Tao, J.; Jiang, Y.; Zhu, J. Climate-driven global changes in carbon use efficiency. Glob. Ecol. Biogeogr. 2014, 23, 144–155. [Google Scholar] [CrossRef]
- Xia, L.; Wang, F.; Mu, X.; Jin, K.; Sun, W.; Gao, P.; Zhao, G. Water use efficiency of net primary production in global terrestrial ecosystems. J. Earth Syst. Sci. 2015, 124, 921–931. [Google Scholar] [CrossRef]
- Zhao, F.; Wu, Y.; Ma, S.; Lei, X.; Liao, W. Increased water use efficiency in China and its drivers during 2000–2016. Ecosystems 2022, 25, 1476–1492. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, P.; Shen, W.; Rao, X.; Hu, Y. Physiological homeostasis and morphological plasticity of two tree species subjected to precipitation seasonal distribution changes. Perspect. Plant Ecol. Evol. Syst. 2017, 25, 1–19. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, J.; Huang, L.; Zhang, X.; Tan, X.; Zhang, L. Influence of canopy structure on understory vegetation of secondary evergreen broadleaf forest communities. J. Zhejiang A F Univ. 2019, 36, 1151–1157. [Google Scholar]
- Wu, Z.; Dai, E.; Lin, M. Simulating the effect of climate change and human activities on the forest ecosystem in the hilly red soil region of southern China: A case study in Taihe county, Jiangxi province. Geogr. Res. 2018, 37, 2141–2152. [Google Scholar]
- Ren, H.; Chen, H.; Li, L.; Li, P.; Hou, C.; Wang, H.; Zhang, Q.; Zhang, P. Spatial and temporal patterns of carbon storage from 1992 to 2002 in forest ecosystems in Guangdong, Southern China. Plant Soil 2013, 363, 123–138. [Google Scholar] [CrossRef]
- Wu, X.; Shi, W.; Guo, B.; Tao, F. Large spatial variations in the distributions of and factors affecting forest water retention capacity in China. Ecol. Indic. 2020, 113, 106152. [Google Scholar] [CrossRef]
- Liu, Y.; Cai, Z.; Yu, Q.; Wu, W.; Zhou, Q. Spatial variation evaluation of cultivated land quality from the perspective of crop rotation for a typical lateritic red soil farming area in south China. J. Agric. Resour. Environ. 2021, 38, 1051–1063. [Google Scholar]
- Feldman, A.F.; Feng, X.; Felton, A.J.; Konings, A.G.; Knapp, A.K.; Biederman, J.A.; Poulter, B. Plant responses to changing rainfall frequency and intensity. Nat. Rev. Earth Environ. 2024, 5, 276–294. [Google Scholar] [CrossRef]
- Lv, P.; Sun, S.; Zhao, X.; Li, Y.; Zhao, S.; Zhang, J.; Hu, Y.; Guo, A.; Yue, P.; Zuo, X. Effects of altered precipitation patterns on soil nitrogen transformation in different landscape types during the growing season in northern China. Catena 2023, 222, 106813. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, J.; Lambers, H.; Wu, J.; Qin, G.; Li, Y.; Li, Y.; Li, Z.; Wang, J.; Wang, F. Intensified rainfall in the wet season alters the microbial contribution to soil carbon storage. Plant Soil 2022, 476, 337–351. [Google Scholar] [CrossRef]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2023, 8, 343–351. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef]
- Hou, Q.; Pei, T.; Yu, X.; Chen, Y.; Ji, Z.; Xie, B. The seasonal response of vegetation water use efficiency to temperature and precipitation in the Loess Plateau, China. Glob. Ecol. Conserv. 2022, 33, e01984. [Google Scholar] [CrossRef]
- Crous, K.Y.; Uddling, J.; De Kauwe, M.G. Temperature responses of photosynthesis and respiration in evergreen trees from boreal to tropical latitudes. New Phytol. 2022, 234, 353–374. [Google Scholar] [CrossRef]
- Liu, Q.; Piao, S.; Janssens, I.A.; Fu, Y.; Peng, S.; Lian, X.; Ciais, P.; Myneni, R.B.; Peñuelas, J.; Wang, T. Extension of the growing season increases vegetation exposure to frost. Nat. Commun. 2018, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; She, W.; Zhang, Y.; Qiao, Y.; Fu, J.; Qin, S. N enrichment, increased precipitation, and the effect of shrubs collectively shape the plant community in a desert ecosystem in northern China. Sci. Total Environ. 2020, 716, 135379. [Google Scholar] [CrossRef]
- Chen, J.; Pu, J.; Li, J.; Zhang, T. Response of carbon-and water-use efficiency to climate change and human activities in China. Ecol. Indic. 2024, 160, 111829. [Google Scholar] [CrossRef]
- Hai, X.; Li, J.; Li, J.; Liu, Y.; Dong, L.; Wang, X.; Lv, W.; Hu, Z.; Shangguan, Z.; Deng, L. Variations in plant water use efficiency response to manipulated precipitation in a temperate grassland. Front. Recent Dev. Plant Sci. 2022, 13, 881282. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Li, H.; Desai, A.R.; Nagy, Z.; Luo, J.; Kolb, T.E.; Olioso, A.; Xu, X.; Yao, L.; Kutsch, W.; et al. How is water-use efficiency of terrestrial ecosystems distributed and changing on Earth? Sci. Rep. 2014, 4, 7483. [Google Scholar] [CrossRef] [PubMed]
- He, L.M.; Chen, J.M.; Pan, Y.; Pan, Y.D.; Birdsey, R.; Kattge, J. Relationships between net primary productivity and forest stand age in US forests. Glob. Biogeochem. Cycles 2012, 26, GB3009. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, G. Spatial variations and controls of carbon use efficiency in China’s terrestrial ecosystems. Sci. Rep. 2019, 9, 19516. [Google Scholar] [CrossRef]
- Fu, Y.; Jian, S.; Yu, X. Water use efficiency in China is impacted by climate change and land use and land cover. Environ. Sci. Pollut. Res. 2024, 31, 42840–42856. [Google Scholar] [CrossRef]
- Boretti, A. Evaluating water use efficiency and CO₂ absorption in plants under rising atmospheric carbon dioxide levels. J. Atmos. Sol.-Terr. Phys. 2024, 266, 106409. [Google Scholar] [CrossRef]
- Chen, T.; Peng, L.; Wang, Y. Prediction of global water use efficiency and its response to vapor pressure deficit and soil moisture coupling in the 21st century. J. Hydrol. 2024, 635, 131203. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Data Source | Spatial Resolution | Temporal Resolution |
|---|---|---|---|
| GPP | MOD17A3HGFv061 (https://lpdaac.usgs.gov/products/mod17a3hgfv061/, accessed on 1 September 2024) | 500 m | 1a |
| NPP | MOD17A3HGFv061 (https://lpdaac.usgs.gov/products/mod17a3hgfv061/, accessed on 1 September 2024) | 500 m | 1a |
| ET | MOD16A2GFv061 (https://lpdaac.usgs.gov/products/mod16a2gfv061/, accessed on 1 September 2024) | 500 m | 8d |
| Temperature | National Earth System Science Data Center (https://loess.geodata.cn, accessed on 1 September 2024) | 1 km | 1 mon |
| Precipitation | National Earth System Science Data Center (https://loess.geodata.cn, accessed on 1 September 2024) | 1 km | 1 mon |
| Land use | MCD12Q1v061 (https://lpdaac.usgs.gov/products/mcd12q1v061/, accessed on 1 September 2024) | 500 m | 1a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, S.; Zhou, P.; Lin, J.; Tan, Z.; Huang, J.; Yan, P.; Chen, H. Spatiotemporal Variation in Carbon and Water Use Efficiency and Their Influencing Variables Based on Remote Sensing Data in the Nanling Mountains Region. Remote Sens. 2025, 17, 648. https://doi.org/10.3390/rs17040648
Lei S, Zhou P, Lin J, Tan Z, Huang J, Yan P, Chen H. Spatiotemporal Variation in Carbon and Water Use Efficiency and Their Influencing Variables Based on Remote Sensing Data in the Nanling Mountains Region. Remote Sensing. 2025; 17(4):648. https://doi.org/10.3390/rs17040648
Chicago/Turabian StyleLei, Sha, Ping Zhou, Jiaying Lin, Zhaowei Tan, Junxiang Huang, Ping Yan, and Hui Chen. 2025. "Spatiotemporal Variation in Carbon and Water Use Efficiency and Their Influencing Variables Based on Remote Sensing Data in the Nanling Mountains Region" Remote Sensing 17, no. 4: 648. https://doi.org/10.3390/rs17040648
APA StyleLei, S., Zhou, P., Lin, J., Tan, Z., Huang, J., Yan, P., & Chen, H. (2025). Spatiotemporal Variation in Carbon and Water Use Efficiency and Their Influencing Variables Based on Remote Sensing Data in the Nanling Mountains Region. Remote Sensing, 17(4), 648. https://doi.org/10.3390/rs17040648