

Drought Amplifies the Suppressive Effect of Afforestation on Net Primary Productivity in Semi-Arid Ecosystems: A Case Study of the Yellow River Basin



Abstract

1. Introduction
2. Data and Methodology
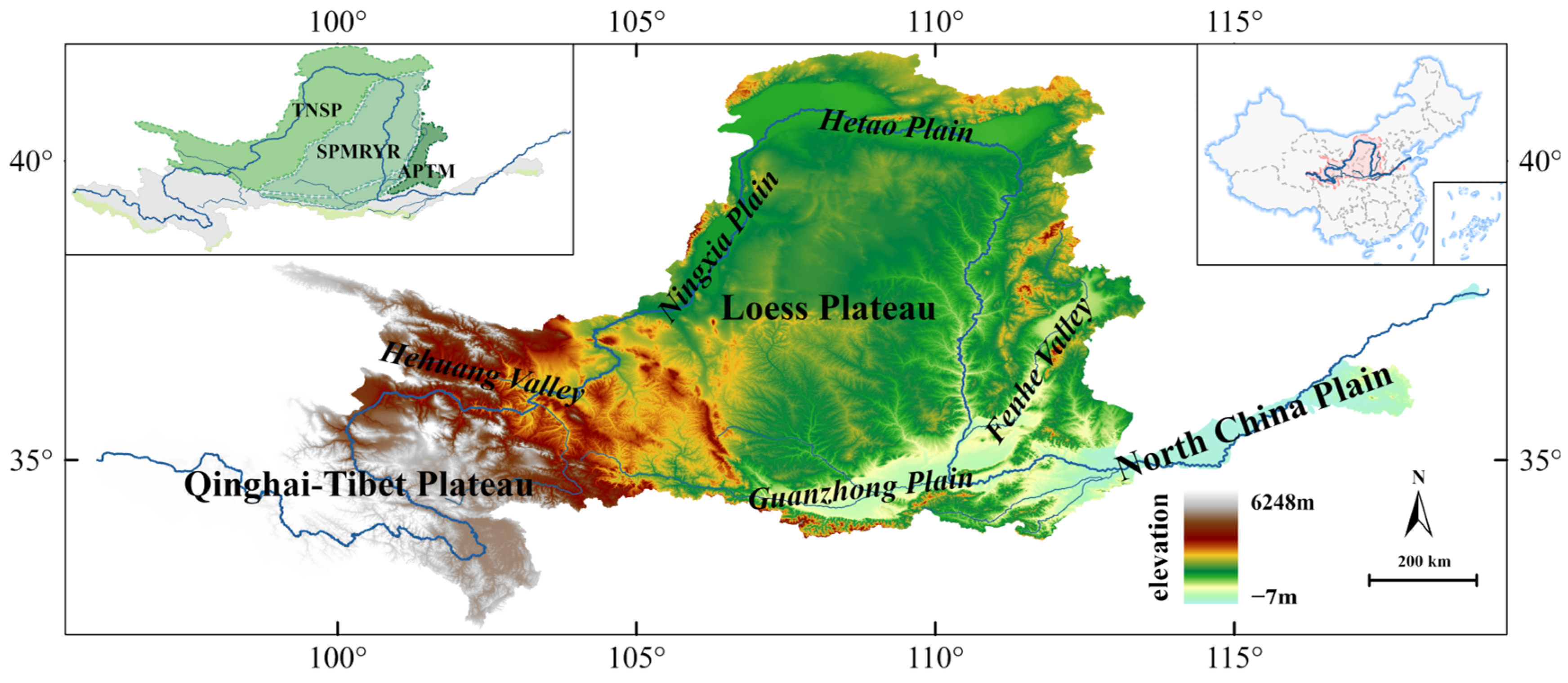
2.1. Study Area
2.2. Dataset
2.3. Research Methodology
2.3.1. Drought Event Recognition and Multidimensional Feature Extraction
2.3.2. NDVI Dynamic Attribution and Driving Mechanisms Analysis
2.3.3. Vegetation NPP Simulation and Scenario Experiments
3. Results
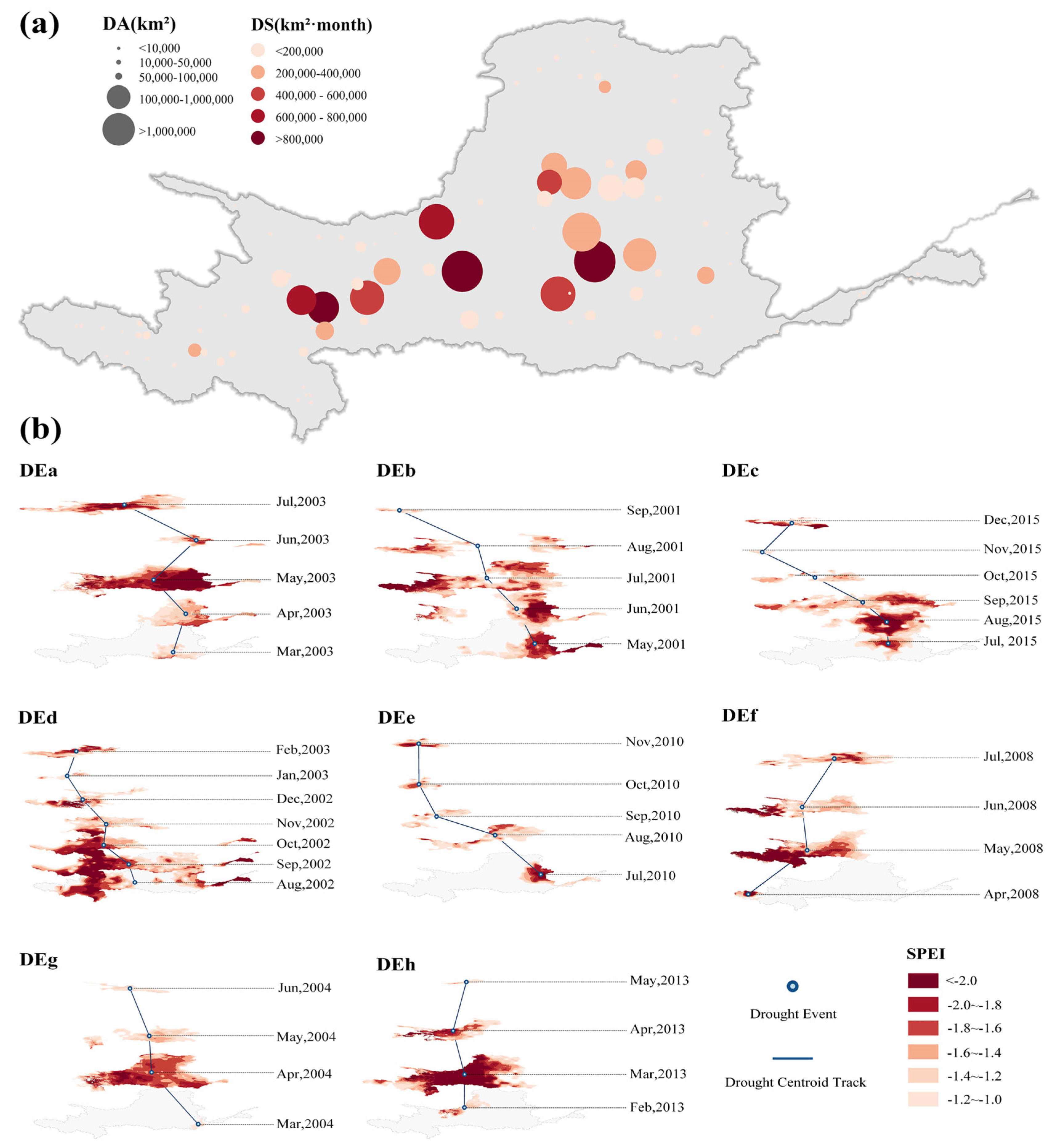
3.1. Spatial and Temporal Characteristics of Extreme Drought Events
3.2. The Suppression of Drought on NPP
3.3. The Suppressive Effect of Afforestation Projects on NPP
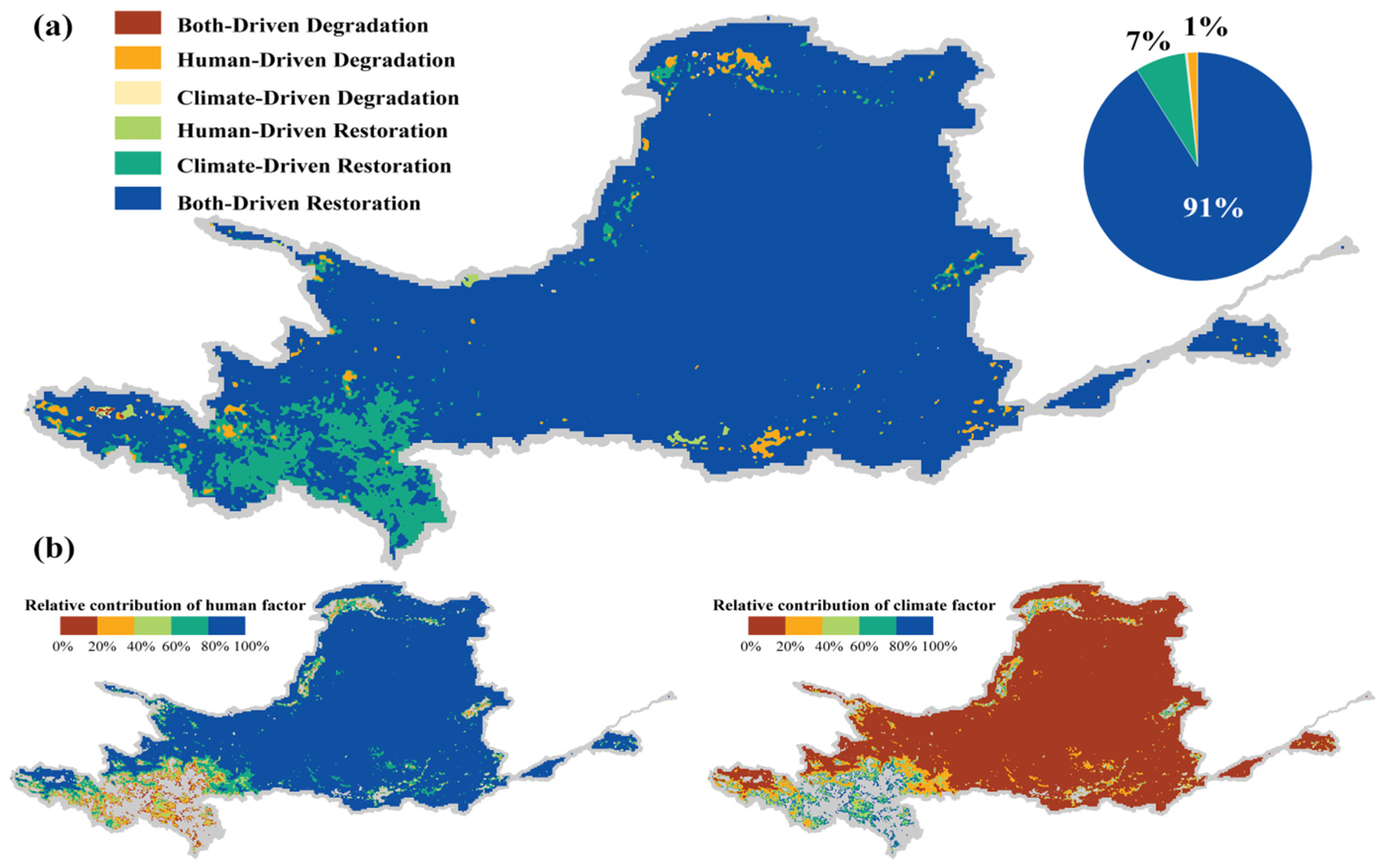
3.3.1. The Leading Role of Afforestation Projects in NDVI Growth
3.3.2. Decoupling of NDVI-NPP
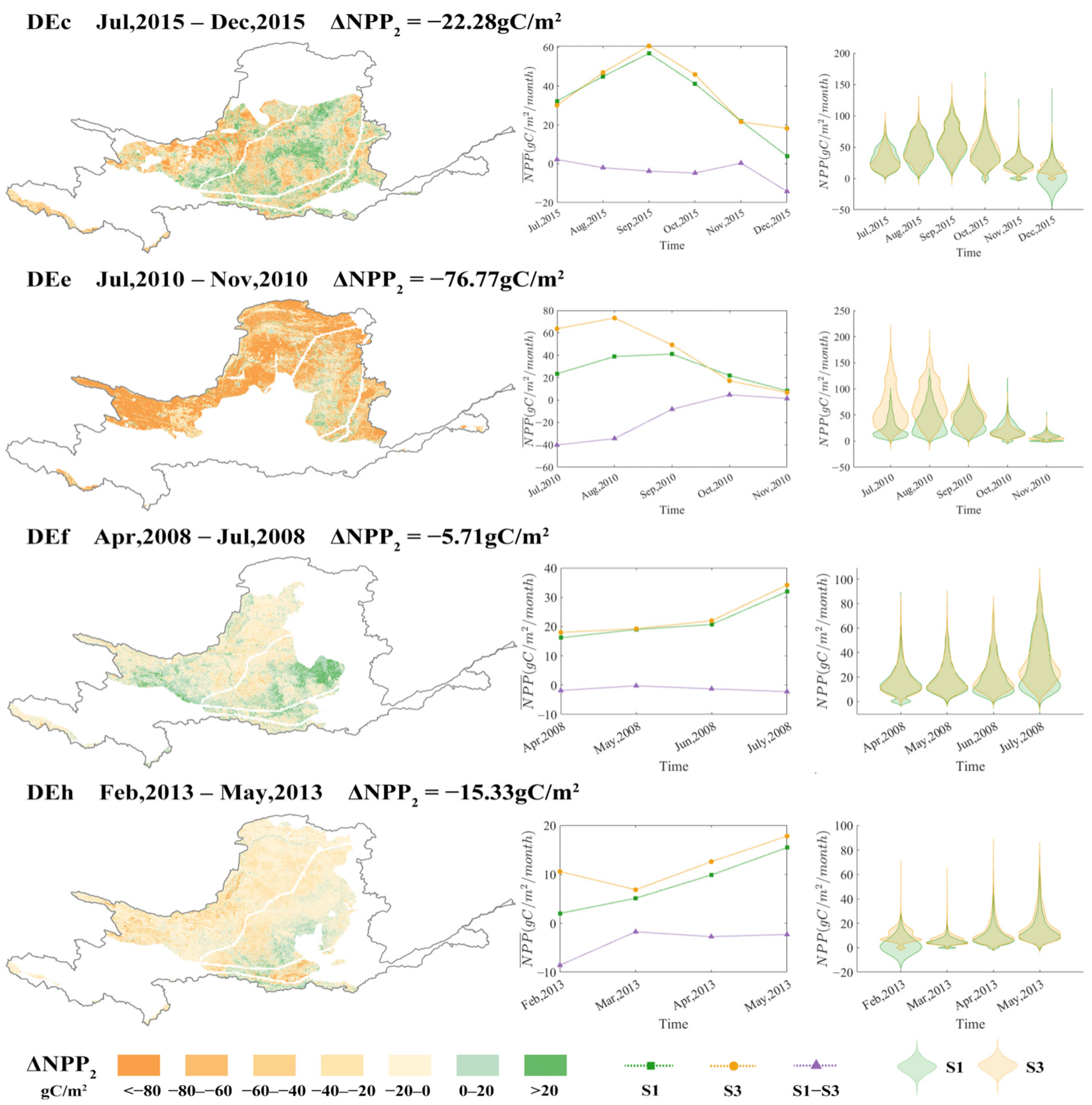
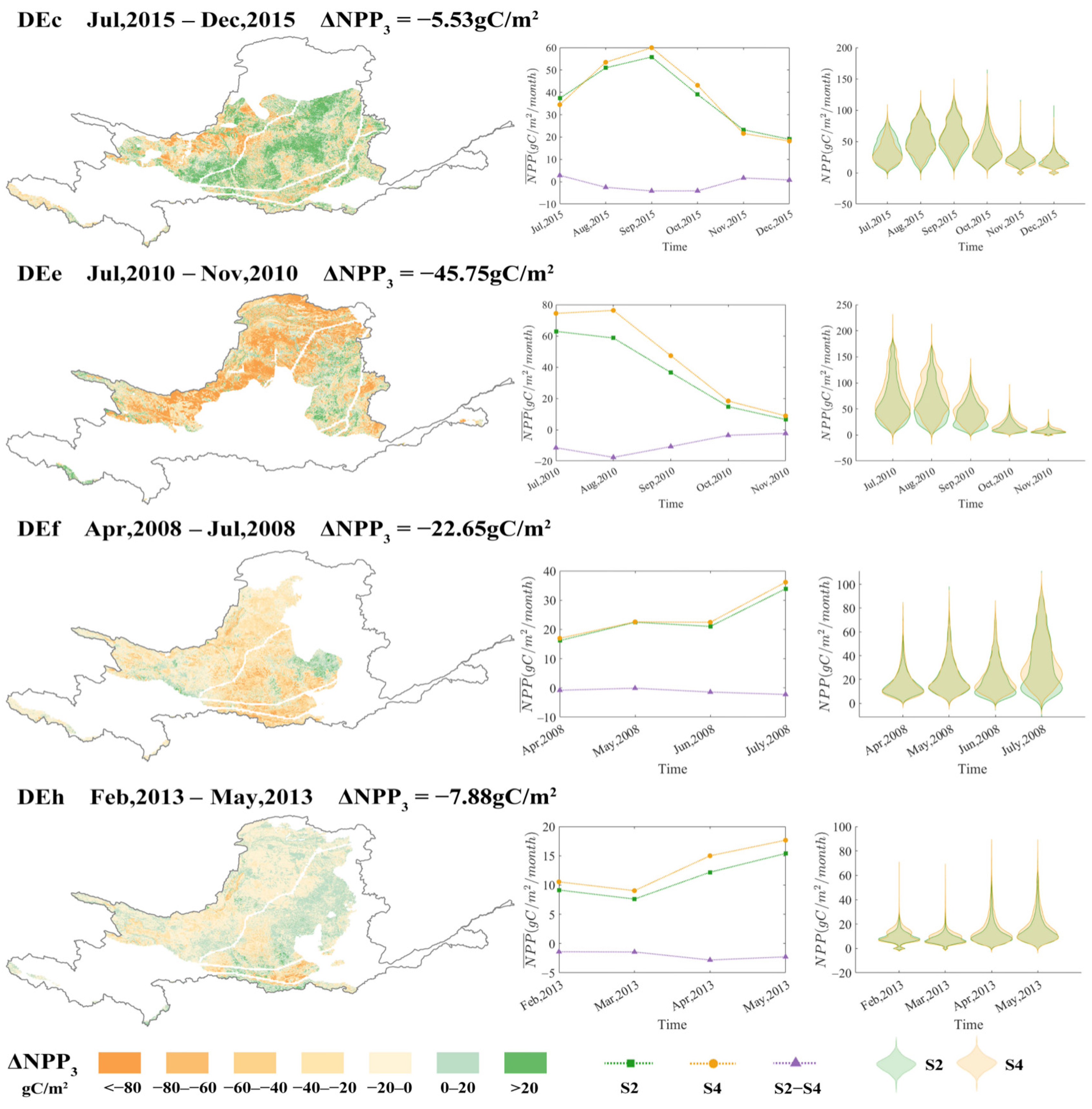
3.4. Amplification of the Suppressive Effect by Drought
4. Discussion
4.1. The Ecological Mechanism of the Suppressive Effect of Afforestation on NPP
4.2. Mechanisms of Drought Amplification of Suppressive Effects
4.3. The Applicability of 3D Clustering Algorithm
4.4. Ecological Management Strategy Optimization Path
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NPP | Net Primary Productivity |
| CASA | Carnegie–Ames–Stanford Approach |
| STL | Seasonal-Trend Decomposition using Loess |
| SPEI | Standardized Precipitation Evapotranspiration Index |
| TNSP | Three-North Shelterbelt Forest Program |
| PAR | Photosynthetically Active Radiation |
References
- Berthelot, M.; Friedlingstein, P.; Ciais, P.; Monfray, P.; Dufresne, J.L.; Le Treut, H.; Fairhead, L. Global response of the terrestrial biosphere to CO2 and climate change using a coupled climate-carbon cycle model. Glob. Biogeochem. Cycles 2002, 16, 31-1–31-15. [Google Scholar] [CrossRef]
- Cao, M.K.; Prince, S.D.; Li, K.R.; Tao, B.; Small, J.; Shao, X.M. Response of terrestrial carbon uptake to climate interannual variability in China. Glob. Change Biol. 2003, 9, 536–546. [Google Scholar] [CrossRef]
- Wahyudi, A.J.; Afdal. Climate Change Mitigation: From Carbon Cycle to Policy. In Proceedings of the 9th International Conference on Global Resource Conservation (ICGRC) and AJI from Ritsumeikan University, Malang, Indonesia, 7–8 March 2018; Volume 2019, p. 040019. [Google Scholar]
- Wang, B.; Sun, R.; Deng, Y. The Variability of Net Primary Productivity and Its Response to Climatic Changes Based on the Methods of Spatiotemporal Decomposition in the Yellow River Basin, China. Pol. J. Environ. Stud. 2022, 31, 4229–4312. [Google Scholar] [CrossRef]
- White, A.; Cannell, M.G.R.; Friend, A.D. CO2 stabilization, climate change and the terrestrial carbon sink. Glob. Change Biol. 2000, 6, 817–833. [Google Scholar] [CrossRef]
- Pan, S.; Tian, H.; Dangal, S.R.; Zhang, C.; Yang, J.; Tao, B.; Ouyang, Z.; Wang, X.; Lu, C.; Ren, W.; et al. Complex Spatiotemporal Responses of Global Terrestrial Primary Production to Climate Change and Increasing Atmospheric CO2 in the 21st Century. PLoS ONE 2014, 9, e112810. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J.Y. High ecosystem stability of evergreen broadleaf forests under severe droughts. Glob. Change Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.S.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- Zhao, D.D.; Jia, W.Y.; Liu, J.P. Dynamic Changes and Driving Mechanisms of Net Primary Production (NPP) in a Semi-Arid Region of China. Sustainability 2023, 15, 11829. [Google Scholar] [CrossRef]
- Bao, G.; Bao, Y.H.; Qin, Z.H.; Xin, X.P.; Bao, Y.L.; Bayarsaikan, S.; Zhou, Y.; Chuntai, B. Modeling net primary productivity of terrestrial ecosystems in the semi-arid climate of the Mongolian Plateau using LSWI-based CASA ecosystem model. Int. J. Appl. Earth Obs. Geoinf. 2016, 46, 84–93. [Google Scholar] [CrossRef]
- Doughty, C.E.; Metcalfe, D.B.; Girardin, C.A.J.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, M.C.; Rocha, W.; et al. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef]
- Lawlor, D.W. Limitation to photosynthesis in water-stressed leaves: Stomata vs. metabolism and the role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Dai, X.A.; Li, W.Y.; Zhou, J.Y.; Zhang, J.J.; Li, C.; Dai, T.R.; Li, W.L.; Lu, H.; Ye, Y.K.; et al. Response of net primary productivity of vegetation to drought: A case study of Qinba Mountainous area, China (2001–2018). Ecol. Indic. 2023, 149, 110148. [Google Scholar] [CrossRef]
- Jiang, X.W.; Bai, J.J. Quantifying the impacts of drought and ecological restoration on net primary production changes in the Chinese Loess Plateau. PLoS ONE 2020, 15, e0238997. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Yang, Q.Y.; Du, Z. Delineation of eco-geographic regional system of China. J. Geogr. Sci. 2003, 13, 309–315. [Google Scholar]
- Li, J.G.; Wang, Y.; Liu, L.L. Responses of the Terrestrial Ecosystem Productivity to Droughts in China. Front. Earth Sci. 2020, 8, 59. [Google Scholar] [CrossRef]
- Sun, B.F.; Zhao, H.; Wang, X.K. Effects of drought on net primary productivity: Roles of temperature, drought intensity, and duration. Chin. Geogr. Sci. 2016, 26, 270–282. [Google Scholar] [CrossRef]
- Li, X.Y.; Piao, S.L.; Huntingford, C.; Peñuelas, J.; Yang, H.; Xu, H.; Chen, A.P.; Friedlingstein, P.; Keenan, T.F.; Sitch, S.; et al. Global variations in critical drought thresholds that impact vegetation. Natl. Sci. Rev. 2023, 10, nwad049. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, Y.; Liu, Y.Y.; Tong, L.J.; Zhang, Q.P.; Li, J.L. Assessing the Impacts of Drought on Grassland Net Primary Production at the Global Scale. Sci. Rep. 2019, 9, 14041. [Google Scholar] [CrossRef]
- Li, C.; Filho, W.L.; Yin, J.; Hu, R.C.; Wang, J.; Yang, C.S.; Yin, S.; Bao, Y.H.; Ayal, D.Y. Assessing vegetation response to multi-time-scale drought across inner Mongolia plateau. J. Clean. Prod. 2018, 179, 210–216. [Google Scholar] [CrossRef]
- Zhan, C.; Liang, C.; Zhao, L.; Jiang, S.Z.; Niu, K.J.; Zhang, Y.L. Drought-related cumulative and time-lag effects on vegetation dynamics across the Yellow River Basin, China. Ecol. Indic. 2022, 143, 109409. [Google Scholar] [CrossRef]
- Jiao, Q.; Li, R.; Wang, F.; Mu, X.M.; Li, P.F.; An, C.C. Impacts of Re-Vegetation on Surface Soil Moisture over the Chinese Loess Plateau Based on Remote Sensing Datasets. Remote Sens. 2016, 8, 156. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhang, Y.L.; Wang, X.; He, T.H.; Xia, H.J.; Zhang, K.R.; Zhang, Q.F. National Forest Restoration Projects in China: Cost-Efficiency and Trade-Off Between Carbon Sequestration and Water Consumption. Earths Future 2024, 12, e2024EF004976. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.H.; Piao, S.L.; Xu, B.D.; Chaturvedi, R.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Berdugo, M.; Gaitán, J.J.; Delgado-Baquerizo, M.; Crowther, T.W.; Dakos, V. Prevalence and drivers of abrupt vegetation shifts in global drylands. Proc. Natl. Acad. Sci. USA 2022, 119, e2123393119. [Google Scholar] [CrossRef]
- Lloyd-Hughes, B. A spatio-temporal structure-based approach to drought characterisation. Int. J. Climatol. 2012, 32, 406–418. [Google Scholar] [CrossRef]
- Xu, K.; Yang, D.W.; Xu, X.Y.; Lei, H.M. Copula based drought frequency analysis considering the spatio-temporal variability in Southwest China. J. Hydrol. 2015, 527, 630–640. [Google Scholar] [CrossRef]
- Guo, H.; Bao, A.M.; Ndayisaba, F.; Liu, T.; Jiapaer, G.; El-Tantawi, A.M.; Maeyer, P.D. Space-time characterization of drought events and their impacts on vegetation in Central Asia. J. Hydrol. 2018, 564, 1165–1178. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Kattel, G.R.; Zhang, J.; Shang, Y.; Gnyawali, K.R.; Zhang, F.; Miao, L.J. Slightly enhanced drought in the Yellow River Basin under future warming scenarios. Atmos. Res. 2022, 280, 106423. [Google Scholar] [CrossRef]
- Wang, L.; Shu, Z.K.; Wang, G.Q.; Sun, Z.L.; Yan, H.F.; Bao, Z.X. Analysis of Future Meteorological Drought Changes in the Yellow River Basin under Climate Change. Water 2022, 14, 1896. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, J.Z.; Liu, H.; Chen, X.L.; Yesou, H. Evidence of vegetation greening benefitting from the afforestation initiatives in China. Geo-Spat. Inf. Sci. 2024, 27, 683–702. [Google Scholar] [CrossRef]
- Tian, Q.; Lu, J.Z.; Chen, X.L. A novel comprehensive agricultural drought index reflecting time lag of soil moisture to meteorology: A case study in the Yangtze River basin, China. Catena 2022, 209, 105804. [Google Scholar] [CrossRef]
- Wu, J.R.; Chen, X.L.; Lu, J.Z. Assessment of long and short-term flood risk using the multi-criteria analysis model with the AHP-Entropy method in Poyang Lake basin. Int. J. Disaster Risk Reduct. 2022, 75, 102968. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017[dataset]. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Yuan, W.P.; Liu, S.G.; Yu, G.R.; Bonnefond, J.M.; Chen, J.Q.; Davis, K.; Desai, A.R.; Goldstein, A.H.; Gianelle, D.; Rossi, F.; et al. Global estimates of evapotranspiration and gross primary production based on MODIS and global meteorology data. Remote Sens. Environ. 2010, 114, 1416–1431. [Google Scholar] [CrossRef]
- Yuan, W.P.; Zheng, Y.; Piao, S.L.; Ciais, P.; Lombardozzi, D.; Wang, Y.P.; Ryu, Y.; Chen, G.X.; Dong, W.J.; Hu, Z.M.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Zheng, Y.; Shen, R.Q.; Wang, Y.W.; Li, X.Q.; Liu, S.G.; Liang, S.L.; Chen, J.M.; Ju, W.M.; Zhang, L.; Yuan, W.P. Improved estimate of global gross primary production for reproducing its long-term variation, 1982–2017. Earth Syst. Sci. Data 2020, 12, 2725–2746. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.L.; Lai, C.G. Severe drought events inducing large decrease of net primary productivity in mainland China during 1982–2015. Sci. Total Environ. 2020, 703, 135541. [Google Scholar] [CrossRef]
- Sheffield, J.; Andreadis, K.M.; Wood, E.F.; Lettenmaier, D.P. Global and continental drought in the second half of the twentieth century: Severity-area-duration analysis and temporal variability of large-scale events. J. Clim. 2009, 22, 1962–1981. [Google Scholar] [CrossRef]
- Guo, H.; Bao, A.M.; Liu, T.; Jiapaer, G.; Ndayisaba, F.; Jiang, L.L.; Kurban, A.; Maeyer, P.D. Spatial and temporal characteristics of droughts in Central Asia during 1966–2015. Sci. Total Environ. 2018, 624, 1523–1538. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.P.; Martin, E.; Franchistéguy, L.; Habets, F.; Soubeyroux, J.M.; Blanchard, M.; Baillon, M. Multilevel and multiscale drought reanalysis over France with the Safran-Isba-Modcou hydrometeorological suite. Hydrol. Earth Syst. Sci. 2010, 14, 459–478. [Google Scholar] [CrossRef]
- Diaz, V.; Perez, G.A.C.; Van Lanen, H.A.J.; Solomatine, D.; Varouchakis, E.A. Characterisation of the dynamics of past droughts. Sci. Total Environ. 2020, 718, 134588. [Google Scholar] [CrossRef]
- Huang, S.Z.; Chang, J.X.; Leng, G.Y.; Huang, Q. Integrated index for drought assessment based on variable fuzzy set theory: A case study in the Yellow River basin, China. J. Hydrol. 2015, 527, 608–618. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Charles Griffin & Company: London, UK, 1949. [Google Scholar]
- Yue, S.; Pilon, P.; Cavadias, G. Power of the Mann-Kendall and Spearmans rho tests for detecting monotonic trends in hydrological series. J. Hydrol. 2002, 259, 254–271. [Google Scholar] [CrossRef]
- Alexandersson, H. A homogeneity test applied to precipitation data. Int. J. Climatol. 1986, 6, 661–675. [Google Scholar] [CrossRef]
- Krake, T.; Kltzl, D.; Hgele, D. Uncertainty-Aware Seasonal-Trend Decomposition Based on Loess. IEEE Trans. Vis. Comput. Graph. 2025, 31, 1496–1512. [Google Scholar] [CrossRef]
- Evans, J.; Geerken, R. Discrimination between climate and human-induced dryland degradation. J. Arid. Environ. 2004, 57, 535–554. [Google Scholar] [CrossRef]
- Gao, W.D.; Zheng, C.; Liu, X.H.; Lu, Y.D.; Chen, Y.F.; Wei, Y.; Ma, Y.D. NDVI-based vegetation dynamics and their responses to climate change and human activities from 1982 to 2020: A case study in the Mu Us Sandy Land. China Ecol. Indic. 2022, 137, 108745. [Google Scholar] [CrossRef]
- Zhu, W.Q.; Pan, Y.Z.; He, H.; Wang, L.L.; Mou, M.J.; Liu, J.H. A Changing-Weight Filter Method for Reconstructing a High-Quality NDVI Time Series to Preserve the Integrity of Vegetation Phenology. IEEE Trans. Geosci. Remote Sens. 2012, 50, 1085–1094. [Google Scholar] [CrossRef]
- Zhu, W.Q.; Pan, Y.Z.; Zhang, J.S. Estimation of Net Primary Productivity of Chinese Terrestrial Vegetation Based on Remote Sensing. Chin. J. Plant Ecol. 2007, 3, 413–424. [Google Scholar]
- Xu, H.T.; He, B.; Guo, L.L.; Yan, X.; Dong, J.W.; Yuan, W.P.; Hao, X.M.; Lv, A.F.; He, X.Q.; Li, T.W. Changes in the Fine Composition of Global Forests from 2001 to 2020. J. Remote Sens. 2024, 4, 0119. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Gao, G.Y.; Ran, L.S.; Wang, Y.; Fu, B.J. Afforestation Reduces Deep Soil Carbon Sequestration in Semiarid Regions: Lessons From Variations of Soil Water and Carbon Along Afforestation Stages in Chinas Loess Plateau. J. Geophys. Res. Biogeosci. 2024, 129, e2024JG00828. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, J.Z.; Chen, X.L. Drought offsets the vegetation greenness-induced gross primary productivity from 1982 to 2018 in China. J. Hydrol. 2024, 632, 130881. [Google Scholar] [CrossRef]
- Thitiporn, M.; Thong-ob, T.; Wonprasaid, S. Effect of Soil Moisture and Determination of Critical Soil Moisture Contents of Cassava. Indian J. Agric. Res. 2020, 54, 483–488. [Google Scholar]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of Photosynthesis of C3 Plants in Response to Progressive Drought: Stomatal Conductance as a Reference Parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Tezara, W.; Fernández, M.D.; Donoso, C.; Herrera, A. Seasonal Changes in Photosynthesis and Stomatal Conductance of Five Plant Species from a Semiarid Ecosystem. Photosynthetica 1998, 35, 399–410. [Google Scholar] [CrossRef]
- Marchin, R.M.; Medlyn, B.E.; Tjoelker, M.G.; Ellsworth, D.S. Decoupling between stomatal conductance and photosynthesis occurs under extreme heat in broadleaf tree species regardless of water access. Glob. Change Biol. 2023, 29, 6319–6335. [Google Scholar] [CrossRef]
- Lei, Y.; Zhang, Z.H.; Li, Z.S. Effects of large-scale re-vegetation on soil desiccation in the Loess Plateau: Problems and perspectives. Acta Ecol. Sin. 2019, 39, 7382–7388. [Google Scholar]
- Wei, Y.Q.; Jin, J.L.; Li, H.C.; Zhou, Y.L.; Cui, Y.; Commey, N.A.; Zhang, Y.L.; Jiang, S.M. Assessment of Agricultural Drought Vulnerability Based on Crop Growth Stages: A Case Study of Huaibei Plain, China. Int. J. Disaster Risk Sci. 2023, 14, 209–222. [Google Scholar] [CrossRef]
- Wang, J.X.; Yang, Y.; Huang, J.K.; Adhikari, B. Adaptive irrigation measures in response to extreme weather events: Empirical evidence from the North China plain. Reg. Environ. Change 2019, 19, 1009–1022. [Google Scholar] [CrossRef]
- Chen, G.; Tan, Q.Y.; Xu, M.X.; Liu, G.B. Mixed-species plantations can alleviate water stress on the Loess Plateau. For. Ecol. Manag. 2020, 458, 117767. [Google Scholar]
- Fan, L.; Dong, G.; Frappart, F.; Wigneron, J.P.; Yue, Y.; Xiao, X.; Zhang, Y.; Tao, S.; Cao, L.; Li, Y.; et al. Satellite-Observed Increase in Aboveground Carbon over Southwest China during 2013–2021. J. Remote Sens. 2024, 4, 0113. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Driver Scenario | Relative Contribution of Climate Change | Relative Contribution of Human Activities | |||
|---|---|---|---|---|---|
| Climate-driven vegetation gain | >0 | >0 | <0 | 100% | 0% |
| Anthropogenically promoted vegetation gain | >0 | <0 | >0 | 0% | 100% |
| Climate-anthropogenic synergistic vegetation gain | >0 | >0 | >0 | ||
| Climate-driven vegetation degradation | <0 | <0 | >0 | 100% | 0% |
| Anthropogenic disturbed vegetation degradation | <0 | >0 | <0 | 0% | 100% |
| Climate-anthropogenic synergistic vegetation degradation | <0 | <0 | <0 |
| Sight | Precipitation | Temperature | NDVI | PAR |
|---|---|---|---|---|
| S1 | ● | ● | ● | ● |
| S2 | ○ | ○ | ● | ● |
| S3 | ● | ● | ○ | ● |
| S4 | ○ | ○ | ○ | ● |
| Event ID | Time Period | DD (Months) | DA (×103 km2) | DS (×103 km2·h) | DL (km) | DI |
|---|---|---|---|---|---|---|
| DEa | March, 2000–July, 2000 | 5 | 710.65 | 946.30 | 1442.48 | 0.919 |
| DEb | May, 2001–September, 2001 | 5 | 702.86 | 930.11 | 1204.55 | 0.861 |
| DEc | July, 2015–December, 2015 | 6 | 579.54 | 567.59 | 1348.88 | 0.800 |
| DEd | August, 2002–February, 2003 | 7 | 537.02 | 928.34 | 714.93 | 0.794 |
| DEe | July, 2010–November, 2010 | 5 | 449.98 | 246.08 | 1269.23 | 0.618 |
| DEf | April, 2008–July, 2008 | 4 | 496.91 | 618.94 | 833.98 | 0.601 |
| DEg | March, 2004–June, 2004 | 4 | 591.45 | 464.96 | 811.71 | 0.579 |
| DEh | February, 2013–May, 2013 | 4 | 599.37 | 714.31 | 378.54 | 0.558 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Zhang, Z.; Du, M.; Lu, J.; Chen, X. Drought Amplifies the Suppressive Effect of Afforestation on Net Primary Productivity in Semi-Arid Ecosystems: A Case Study of the Yellow River Basin. Remote Sens. 2025, 17, 2100. https://doi.org/10.3390/rs17122100
Wang F, Zhang Z, Du M, Lu J, Chen X. Drought Amplifies the Suppressive Effect of Afforestation on Net Primary Productivity in Semi-Arid Ecosystems: A Case Study of the Yellow River Basin. Remote Sensing. 2025; 17(12):2100. https://doi.org/10.3390/rs17122100
Chicago/Turabian StyleWang, Futao, Ziqi Zhang, Mingxuan Du, Jianzhong Lu, and Xiaoling Chen. 2025. "Drought Amplifies the Suppressive Effect of Afforestation on Net Primary Productivity in Semi-Arid Ecosystems: A Case Study of the Yellow River Basin" Remote Sensing 17, no. 12: 2100. https://doi.org/10.3390/rs17122100
APA StyleWang, F., Zhang, Z., Du, M., Lu, J., & Chen, X. (2025). Drought Amplifies the Suppressive Effect of Afforestation on Net Primary Productivity in Semi-Arid Ecosystems: A Case Study of the Yellow River Basin. Remote Sensing, 17(12), 2100. https://doi.org/10.3390/rs17122100