1. Introduction
Grapevine (
Vitis vinifera L.) was grown for the production of table grapes and wine, in 2023, on about 6.6 million ha worldwide (
https://www.fao.org/faostat/en/#data, accessed on 3 January 2025). As many vineyards are in regions where water is a limiting resource and drought stress may reduce the quality and quantity of grapevine production, monitoring of the crop water status is very important in viticulture and has become a major research area [
1].
Stomatal conductance, water potential, canopy temperature, and vegetation indices are widely used to characterize the water status of grapevine and other crop species [
1,
2,
3,
4]. Measurements of stomatal conductance and water potential of leaves, stems, or plants result in a mean value per measuring object, deliver no spatial information, and require contact between plant tissue and sensor. In contrast, thermal and spectral sensing and imaging provide various vegetation indices, are contactless and may be applied on various platforms and scales; e.g., ground-based spectrometry and thermography [
5], aerial thermography [
6,
7], aerial multispectral and thermal imagery [
8,
9,
10], and ground-based hyperspectral imaging [
11].
The effects of water content on leaf reflectance are greatest in spectral bands centered at 1450, 1940, and 2500 nm. Secondary effects occur at 980 nm and 1240 nm [
12,
13,
14]. In satellite and airborne imagery, the SWIR wavebands are usually saturated with atmospheric water vapor, which has a higher concentration over the full atmospheric column between sensor and vegetation than the liquid water of the plant tissue [
15]. As the SWIR water absorption bands are not suitable for remote sensing of the water content of vegetation, wavebands from the NIR range are used instead. Spectral band ratios developed to estimate the water content of vegetation from remote sensing include the normalized difference water index (NDWI; [
16]) and the plant water index (PWI; [
17]). These spectral ratio indices use the (broad) wavebands provided by satellite-borne or airborne sensors.
Wavebands around 1200 nm and 1400 nm have been used to predict the water status of grapevines, also from aerial SWIR data [
18,
19]. The best models used SWIR data (750–1700 nm) from UAV and allowed predicting stem water potential with R² of 0.54. Random Forest and Support Vector Machine models used wavebands located around 1420 nm and 1460 nm, and 1050 nm and 1200 nm, respectively [
20].
In proximal sensing, the content of atmospheric water vapor of the column between leaf tissue and sensor is considerably lower than the content of liquid water in living plant tissue. Reflection from ground-based hyperspectral imaging at 1410 nm and 1520 nm was correlated with the water status of tomato leaves [
21]. For the period 2019 to 2022, Sanaeifar et al. [
22] listed 41 studies in which proximal hyperspectral sensors were used for drought detection of various plant species on the leaf and canopy scale; 20 non-imaging sensors, 18 in the VISNIR range, and 2 using SWIR data, in grapevine [
23] and in oak phenotyping [
24], respectively. Oerke & Steiner [
25] successfully used a normalized difference water index NDWI
1937 for characterizing the water status of apple leaves infected by
Venturia inaequalis.
Knowledge of the effect of leaf diseases on the water status of crop plants is limited (see review by Grimmer et al. [
26]). Leaf diseases such as downy mildew, powdery mildew, and various leaf spot diseases are described to affect yield formation of grapevine and to aggravate the effects of drought conditions on host physiology without knowing details on their effect on leaf water status [
27,
28].
Downy mildew of grapevine is caused by the obligate biotrophic oomycete
Plasmopara viticola (Berk. & M.A. Curtis) Berl & De Toni and is one of the most devastating diseases worldwide [
29]. Despite frequent fungicide use, it causes considerable production losses in grapevine production. As the completion of its life cycle depends on host plant resistance and environmental conditions, the use of (partially) resistant cultivars and the use of fungicides according to forecast models, depending on leaf wetness duration, are essential tools in downy mildew control [
30,
31,
32,
33].
Plant-pathogen interactions may be classified as compatible (successful infection leading to disease and pathogen reproduction) or incompatible (successful plant defense, no disease and no pathogen survival) [
34]. In more detail, however, there is a continuum in the level of compatibility of interactions. The interaction between the susceptible genotype Mueller-Thurgau, a typical representative of European grapevine
Vitis vinifera, and
P. viticola is fully compatible and results in the abundant formation of abaxial sporangia. Partial, i.e., incomplete resistance of grapevine cultivars to
P. viticola depends on genes (
Rpv) introgressed from
Vitis species from the Americas and Asia [
35,
36].
Rpv3 (derived from
V. rupestris) and
Rpv10 (from
V. amurensis) confer a high level of resistance of cultivars Regent (carrying
Rpv3-1) and Solaris (
Rpv3-3, Rpv10), respectively, to
P. viticola. Resistance reactions of Regent and Solaris include the formation of spots of brown tissue with some necrotic (dead) cells. Earlier studies, however, revealed that
P. viticola can survive and even sporulate on this tissue, which includes living cells containing haustoria of the pathogen [
37].
The influence of downy mildew on leaf transpiration has been investigated using thermography in cucumber [
38,
39] and grapevine [
40]. Hyperspectral imaging of the leaf water content of apple leaves infested by
Venturia inaequalis demonstrated that increased transpiration does not inevitably reduce the water content of colonized leaf tissue, as the plant may compensate for the increased water loss through the cuticle provided water supply is not limiting [
25]. Only in advanced stages of pathogenesis, the imaging sensor system revealed highly localized water gradients around infections. The study also revealed that the adaxial sides of non-infected apple leaves had higher leaf water content than the abaxial side. However, the relationship between leaf water content (LWC) and Normalized Difference Leaf Water Index was calibrated only for adaxial apple leaf tissue. It was not clear whether the difference in NDLWI
1937 values between leaf sides resulted from an effect on the SWIR water absorption bands, on the tissue structure-characterizing NIR waveband at 1047 nm, or on both leaf characteristics. In dicotyledonous plants, the arrangement of leaf tissues is strongly asymmetric, with the chloroplast-rich palisade parenchyma near the upper surface to intercept incoming light and the spongy mesophyll and large air spaces on the abaxial side of the leaf near the lower surface where the stomata are located [
15]. This bifacial arrangement is likely to cause differences in the spectral characteristics between adaxial and abaxial leaf surfaces. The water-transporting vascular bundle is oriented more to the abaxial side of leaves, with xylem and phloem often located underneath the palisade parenchyma.
Imaging sensors enable the detection, mapping, and modelling of the biochemical and physiological processes in compatible and incompatible host–pathogen interactions [
37,
41,
42,
43]. The objective of the study reported herein was to improve our understanding of the effect of
P. viticola on the leaf water status of grapevine leaves, depending on the compatibility of the host–pathogen interaction, by applying an innovative method of LWC mapping on the tissue scale. Four objectives were addressed; (I) to establish the relationship between LWC measured gravimetrically and a normalized difference vegetation index based on the SWIR water absorption band(s) for adaxial and abaxial sides of grapevine leaves; (II) to check the validity of this relationship under various environmental conditions, e.g., background and grapevine genotype; (III) to map the effect of downy mildew on the leaf water status in compatible and incompatible interactions; and (IV) to track the changes in LWC during pathogenesis in longitudinal studies.
2. Materials and Methods
2.1. Plant Growth
Grapevine (
Vitis vinifera L.) plants of cultivars Mueller-Thurgau, Regent, and Solaris were grown from green cuttings in commercial substrate ED 73 (Balster Einheitserdewerk, Froendenberg, Germany) in plastic pots (10 × 10 cm, 0.9 L volume) in a greenhouse at 23/20 °C (day/night), 60% relative humidity (RH), and a photoperiod of 16 h (>300 μmol m
−2 s
−1). Plants were watered as necessary and fertilized before the experiment once with 100 mL of a 0.2% solution of Poly Crescal (Aglukon, Düsseldorf, Germany). The cultivars differ in field susceptibility to downy mildew: cv. Mueller-Thurgau (white wine, rating 7), cv. Regent (red wine, 3), cv. Solaris (white wine, 3) (scale 1 = resistant, to 9 = susceptible; [
44]).
2.2. Inoculum Preparation and Inoculation
Sporangia of Plasmopara viticola produced on the abaxial side of grapevine leaves of the susceptible cv. Mueller-Thurgau were stored at −18 °C. Sporangia were washed off the leaves with water containing 0.1% Tween-20, adjusted to a concentration of 5 × 104 mL−1, and sprayed onto the abaxial side of the upper four leaves (1.2 mL per leaf) of grapevines by turning the plants upside down during spraying.
For each cultivar, 4–6 plants were inoculated with the pathogen when at least six leaves were fully developed. Inoculated plants were incubated for 24 h at 100% RH, 23/20 °C, 16 h photoperiod; subsequently, they were grown at 60% RH. Non-inoculated control plants were not subjected to spells of 100% RH.
2.3. Disease Assessment
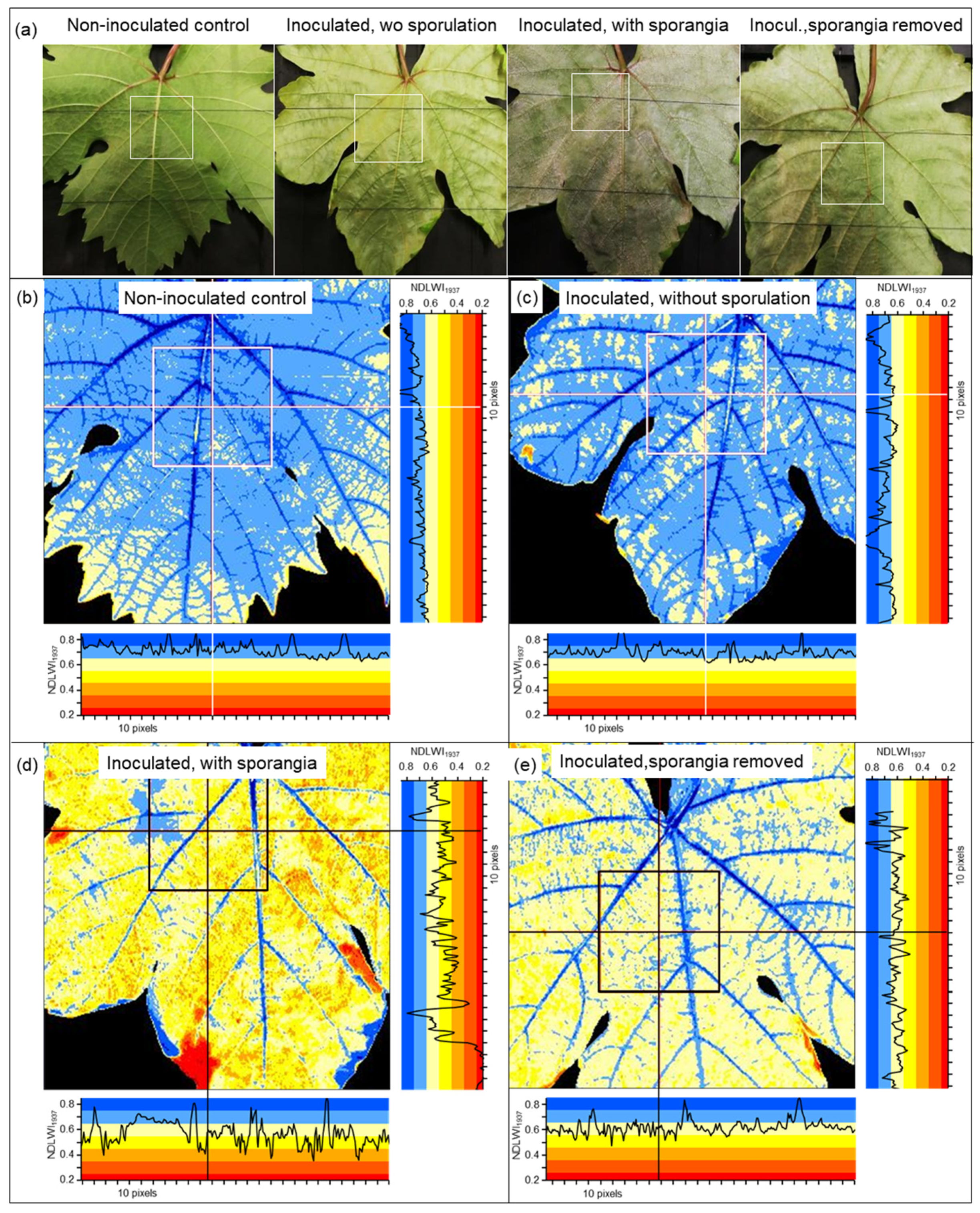
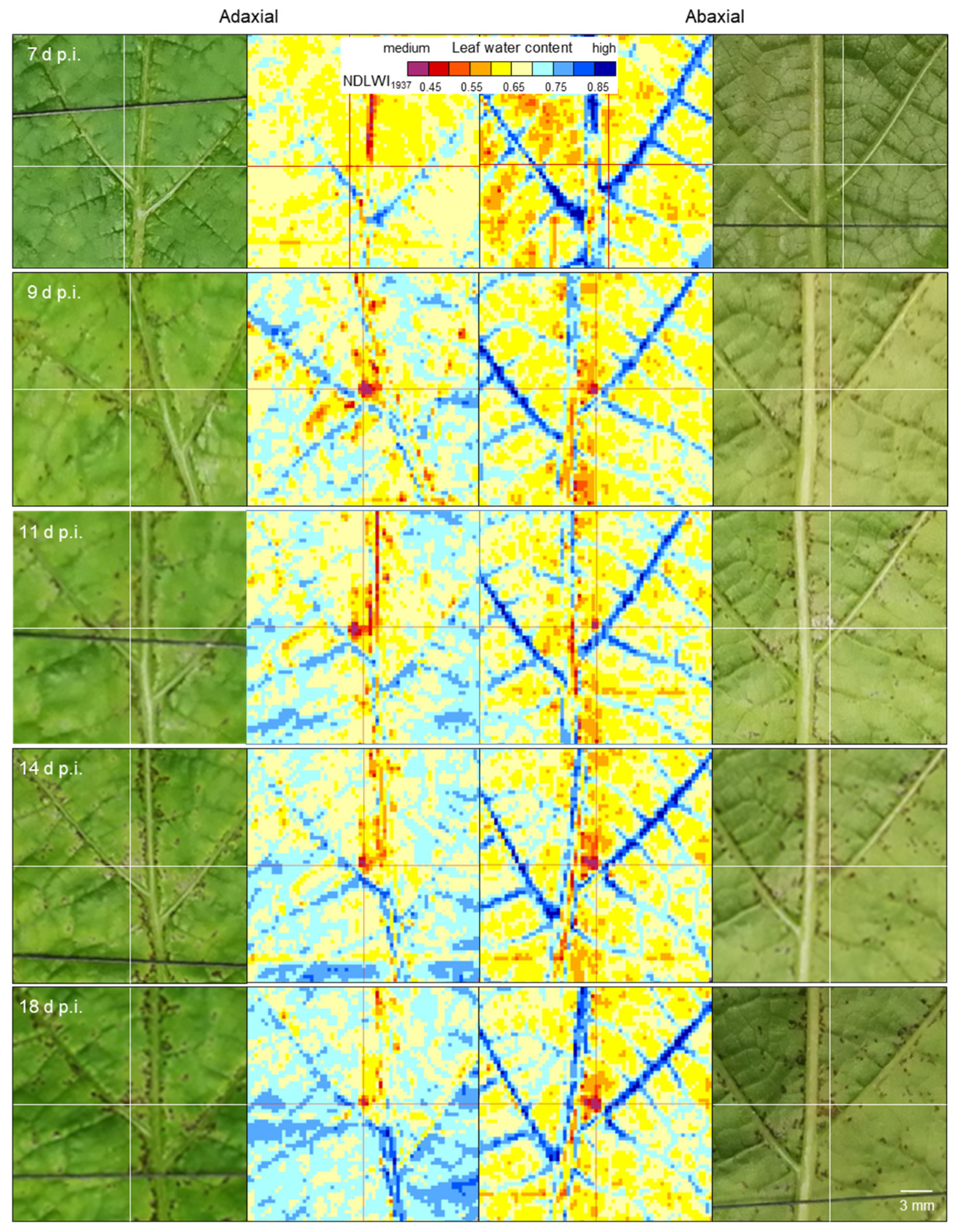
The severity of P. viticola colonization of grapevine leaves was assessed visually starting 3 days post-inoculation (d p.i.). To induce sporulation of P. viticola, infected plants were incubated for 14 h (19.00 to 09.00) at 100% RH. For cv. Mueller-Thurgau, the percent (abaxial) leaf area covered by whitish sporangia was estimated using the classes 0, 1, 2, 5, 10, 20, 30, …, 90, 100%. For cvs. Regent and Solaris, the percentage leaf area covered by tiny brownish leaf spots was estimated using the same classes.
On images of leaf tissue, the size of individual leaf spots was assessed by counting the number of pixels with NDLWI1937 values < 0.55.
2.4. Leaf Thickness
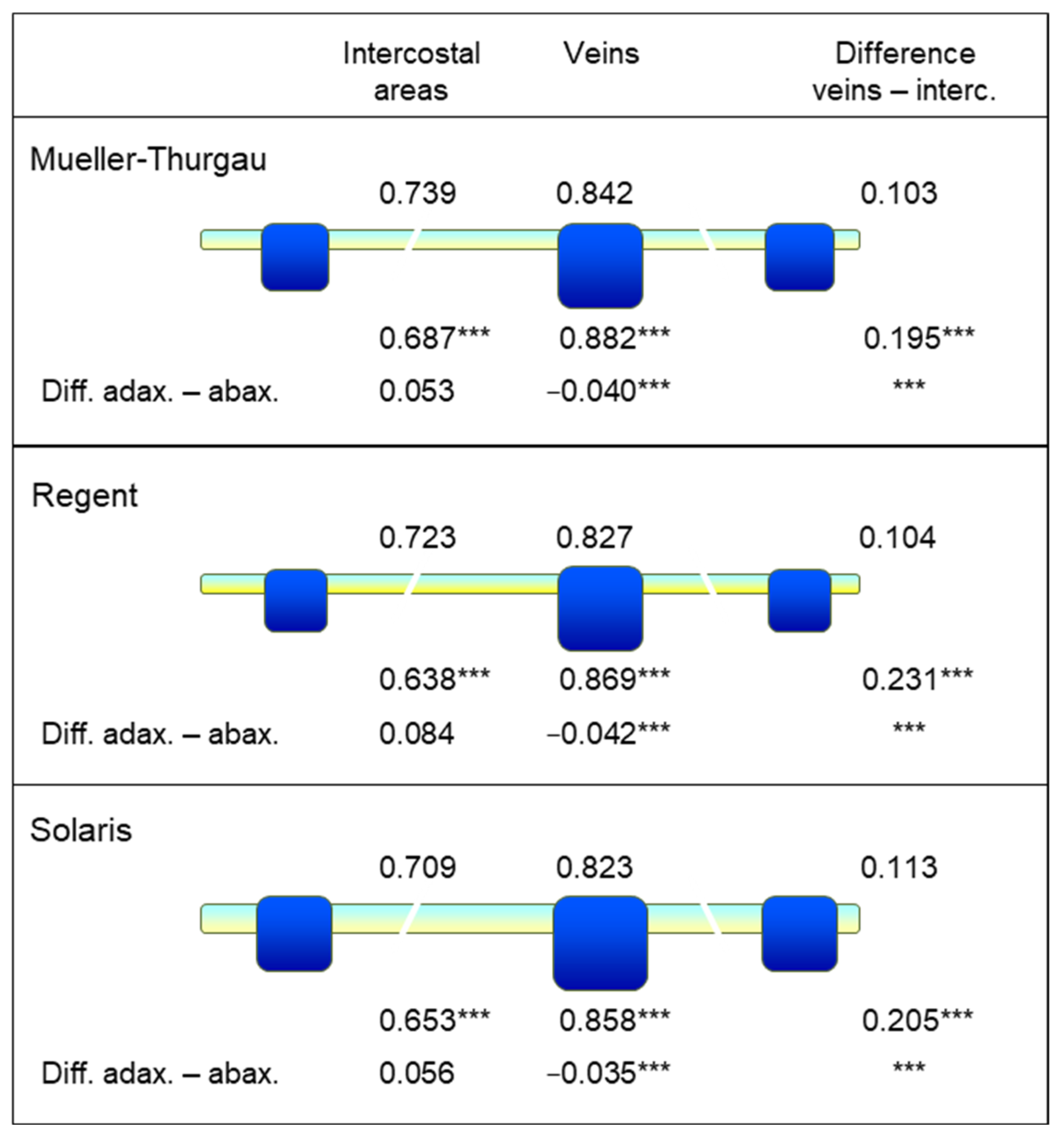
The thickness of grapevine leaves was assessed using a digital sliding caliper (Workzone DMV-SL05, Dario GmbH & Co KG, Hamburg, Germany). The thickness of intercostal areas and second-order leaf veins of ten mature, fully developed leaves and 10 young leaves (leaf level 2 and 3 from top) per grapevine genotype was measured.
2.5. Gravimetric Assessment of Leaf Water Content
The leaf water content (LWC) of grapevine leaves was measured gravimetrically as LWC = (fresh mass − dry mass)/fresh mass × 100. Per treatment, the fresh mass of 8–10 leaves was weighed directly after leaf detachment, and the dry mass after drying at 72 °C for at least 12 h. The LWC value is in contrast to the relative water content, as it does not consider the maximum water content of plant tissue. The experiment was carried out twice.
2.6. Hyperspectral Imaging (HSI)
2.6.1. Measuring System
Hyperspectral images in the shortwave infrared (SWIR) range (940 to 2544 nm) were recorded using a camera with the ImSpector N25E spectrograph (Spectral Imaging Ltd., Oulu, Finland) and an objective with a focal length of 30 mm and a field-of-view of 18°. An effective sensor slit length of 9.6 mm and 320 pixels per line result in a sensor pixel size of 30 µm. Spectral resolution was 6.3 nm (256 bands). Three ASD-Pro-Lamps (Analytical Spectral Devices Inc., Boulder, CO, USA) on each side of the camera provided a near-solar light spectrum and gave even illumination of the scene. Camera and lamps were arranged on a motorized line stage (SP-X-Stage-Dual2, Spectral Imaging Ltd.), which moved the measuring system above the plants. Camera and line stage were controlled using the SpectralCube software (Spectral Imaging Ltd.). Imaging data were recorded in a darkened room for optimal and reproducible measuring conditions.
During HSI measurements, the leaves attached to grapevine stems were fixed between two grids (mesh size 30 × 40 mm) of black fibers in a frame 0.35 m above a black polypropylene background to support the leaves from below and to smooth them from above. The potted grapevine plants and the supporting system were placed on a motorized table in order to vertically adjust the leaves to a working distance of 0.3 m (for maximum spatial resolution) to 0.6 m (for maximal spatial coverage). A set of four reflection images per object were recorded in order to allow the calculation of reflectance values: (I) white reference bar (Spectral Imaging Ltd.) with horizontal size and vertical level similar to the object area; (II) dark current image; (III) image of the object area of interest with optimized exposure time; and (IV) dark current image of the object area.
Hyperspectral images were recorded from leaves without visible water droplets on the surfaces. Leaf surfaces of plants incubated under 100% RH for 14 h overnight in order to induce P. viticola sporulation had to be dried under ambient conditions for >6 h before measurements.
2.6.2. Processing of Hyperspectral Data
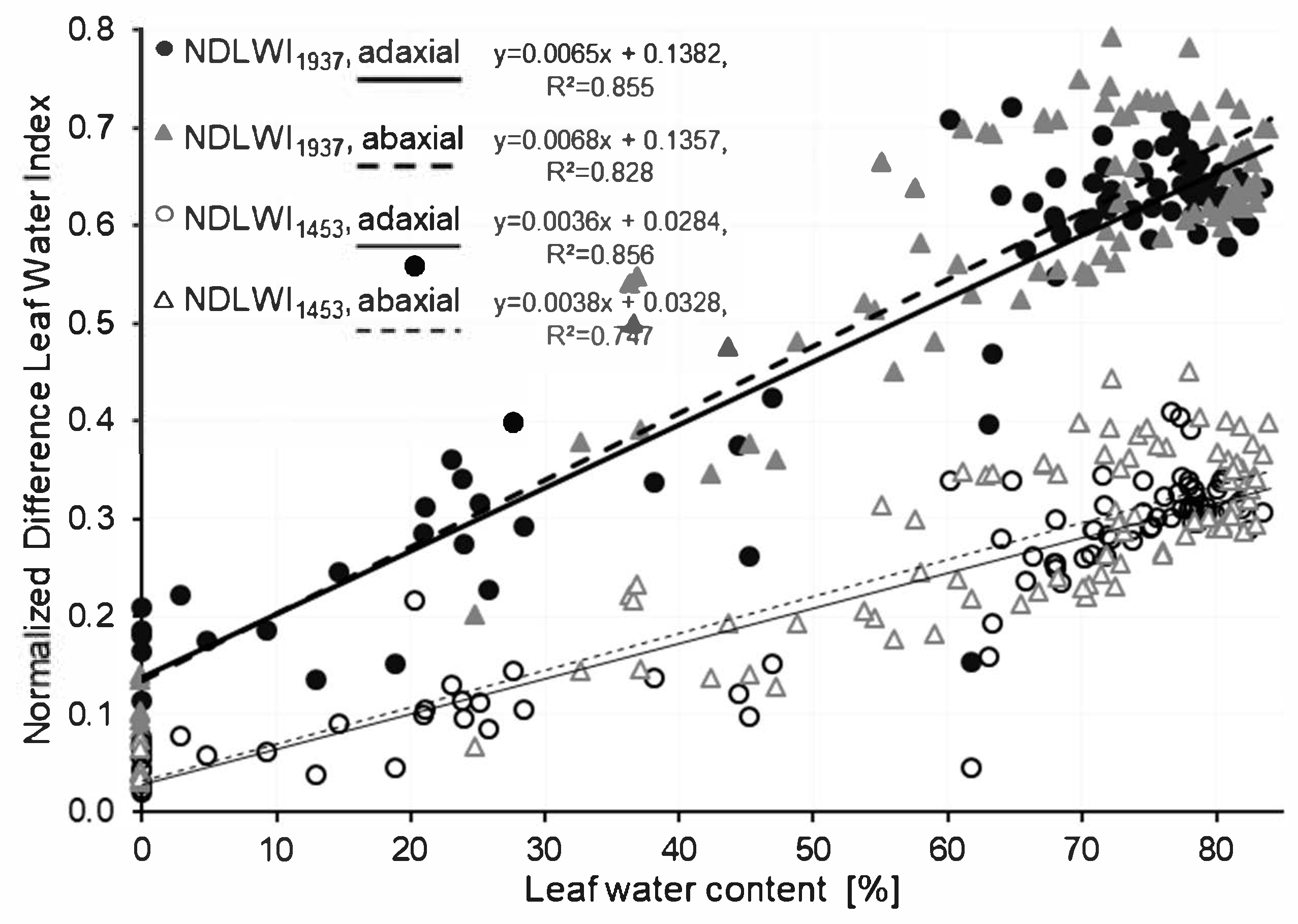
The raw image data (reflection values, 240 wavebands in the range from 997 to 2500 nm) were converted into hyperspectral reflectance images using the software ENVI + IDL 5.0 (EXELIS Visual Information Solutions, Boulder, CO, USA). Data from the 4 images of an object were imported and normalized by a customized IDL tool. Spectral vegetation indices—Normalized Difference Leaf Water Index (NDLWI) values—were calculated using the values of the water absorption bands at 1453 nm (band #82) and 1937 nm (band #159), respectively, and 1124 nm (near infrared, band #30) for normalization: NDLWI1453 = (R1124 − R1453)/(R1124 + R1453) and NDLWI1937 = (R1124 − R1937)/(R1124 + R1937), respectively.
Graphic illustrations of x-profiles and y-profiles along transects through leaf areas were prepared after data transfer from ENVI + IDL to Excel.
2.7. Calibration of Normalized Difference Leaf Water Indices
Non-inoculated young and mature leaves of grapevine (cv. Mueller-Thurgau) attached to the plants—with optimal leaf water content—and after detachment and drying for 2, 5, and 8 h, respectively, were recorded with the hyperspectral SWIR camera using a working distance of 60 cm. Young leaves (n = 10) dried under ambient air conditions (20–22 °C, 50% RH), mature leaves (n = 10) were dried at 40 °C in a drying chamber at maximum ventilation. After spectral recording, the weight of leaves was measured gravimetrically. After the fourth hyperspectral measurement, all leaves were desiccated to dryness at 70 °C for 14 h, and the dry matter of leaves was measured after equilibration with ambient air.
Marking the perimeter of the leaves recorded, averaging the spectral information of these regions of interest (ROIs), and extracting the reflectance values at 1124 nm, 1453 nm, and 1937 nm yielded the spectral information for the calibration curves NDLWI1453 vs. leaf water content and NDLWI1937 vs. leaf water content, respectively. Due to the venation of the grapevine leaf, ROIs included varying percentages of leaf veins and intercostal areas, respectively.
2.8. Statistical Analysis
For statistical analysis, individual grapevine leaves were used as the experimental unit. Statistical analyses were conducted using the software SPSS Statistics vers. 26.0 (IBM Germany GmbH, Ehningen, Germany). Data were tested for normal distribution and equality of variances using the Shapiro–Wilk test and Levene’s test, respectively. For normally distributed data, a standard ANOVA was performed. For significant F-values, mean comparisons were performed using the Tukey–Kramer test at a significance level of p ≤ 0.05. T-test statistics (significance level p ≤ 0.05) were applied to detect significant differences between healthy and infected leaves as well as adaxial and abaxial leaf sides. Data series were related to each other by the Pearson correlation coefficient (r). All experiments were conducted in biologically independent repetitions at least twice.
4. Discussion
4.1. Relationship Between Leaf Water Content and NDLWI1937
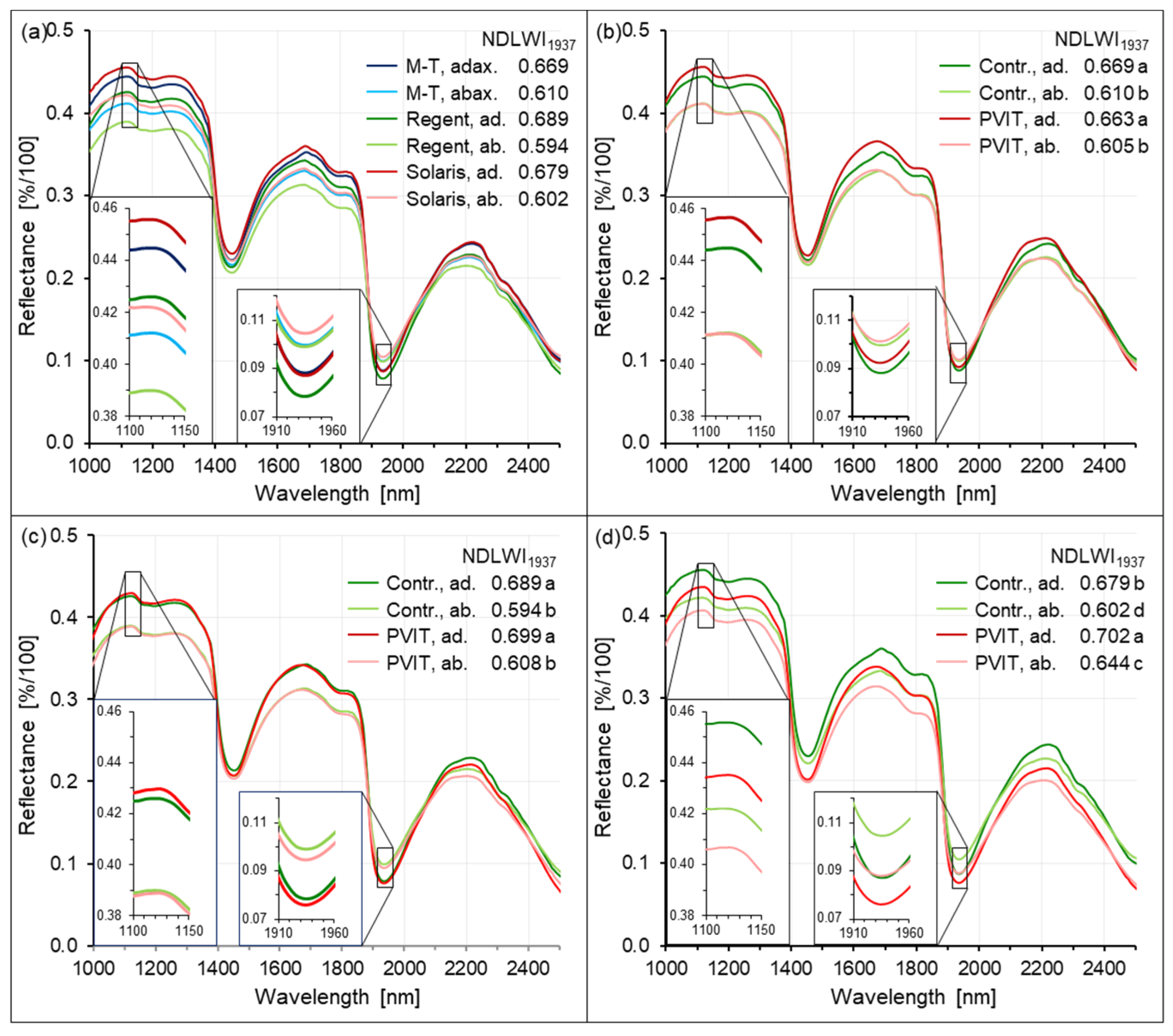
The Normalized Difference Leaf Water Index for grapevine uses the water absorption band at 1940 nm and the maximum NIR reflectance at 1124 nm, and is very similar to the NDLWI established for apple leaves [
25]. Proximal sensing enables the use of SWIR water absorption bands without the risk of corruption of the water signal on its path from leaf to sensor. Normalization by a NIR waveband proved to be very useful when studying the variation of this spectral information depending on grapevine genotype and leaf tissue variability. The greater y-axis intercept of the NDLWI
1937 as compared to apple leaves is likely to result from the inclusion of young grapevine leaves, which had higher reflectance of dried leaves than older ones. The higher tissue variability of grapevine leaves covering intercostal areas and veins of several orders also contributed to a lower coefficient of determination of the regression line for grapevine leaves (0.855 compared to 0.918 for apple leaves;
Figure S2). As downy mildew pathogens primarily produce abaxial disease symptoms, a calibration curve was also established for the lower leaf side of grapevine leaves in a separate experiment, which was almost identical to the relationship for the adaxial side.
Grapevine genotypes significantly differed in the thickness of leaf tissue and veins, and intercostal areas caused considerable variability of tissue thickness within leaves. Although the normalization of the leaf water index using the sum of both wavebands proved to increase its robustness to tissue variability and illumination conditions, NDLWI1937 was used to visualize patterns of the leaf water status of infected leaves instead of LWC values calculated from the calibration curve. The variability in the relationship between spectral information and leaf water content resulted from using mean values for both the water content of whole leaves and averages of spectral information. Although spectral information may be sub-classified easily to veins (of various orders) and intercostal areas, it is hard to obtain the ground truth of the water content of specific leaf parts. The use of LWC values calculated from the calibration curve for maps of the leaf water status would neglect the effect of within-leaf variability. Moreover, the relative differences displayed in NDLWI1937 maps were used for leaves of grapevine cultivars differing in leaf thickness.
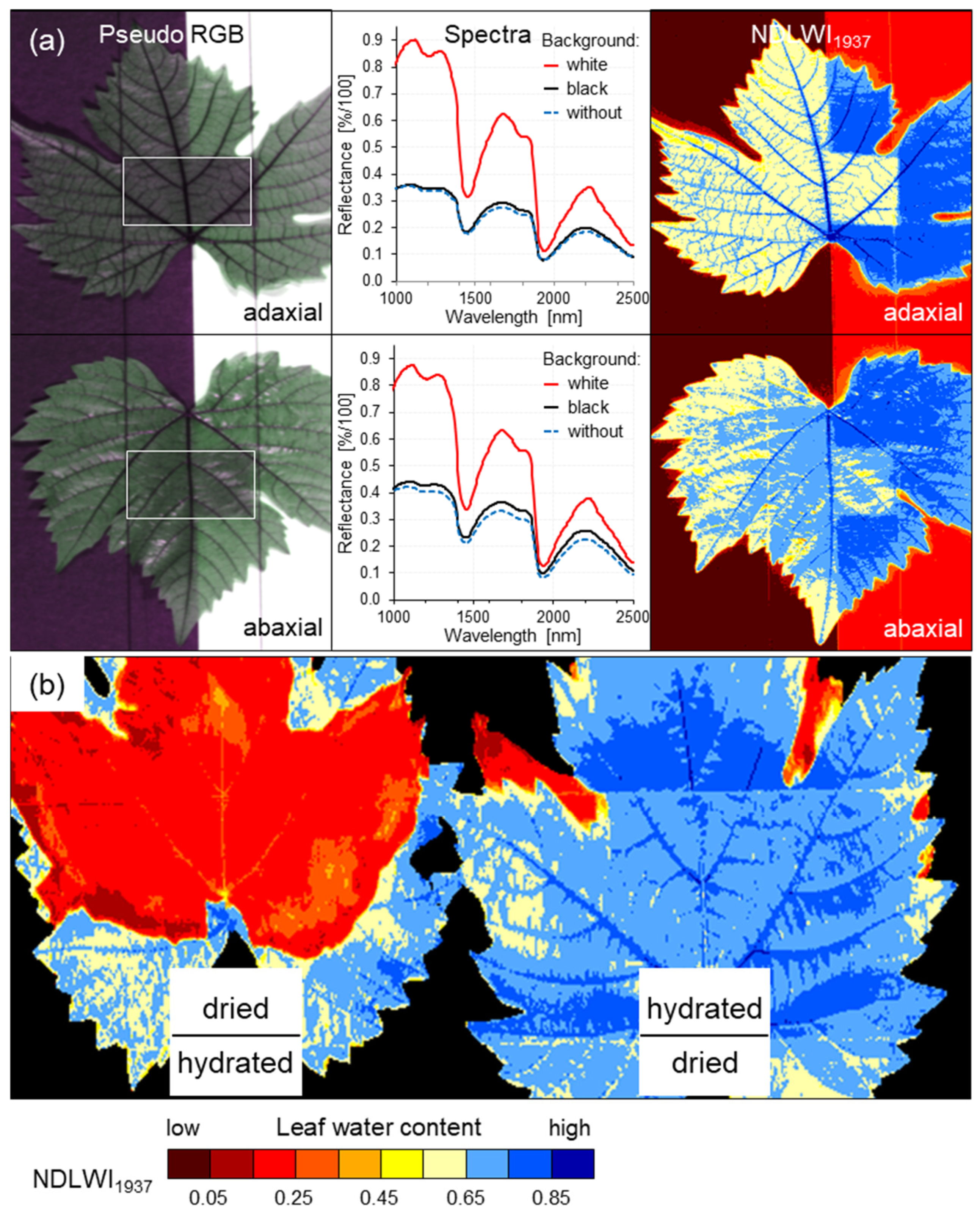
Interestingly, experiments with different backgrounds and with the stacking of leaves varying in leaf water content demonstrated that the reflection of the normally transmitted part of the irradiation by a white and dry background significantly increased SWIR reflectance and NDLWI1937 values by 0.15 to 0.20 units. Using a non-reflecting background about 0.35 m below the measured leaf, grapevine tissue with a leaf water content strongly differing from that of the leaf of interest above, in contrast, had hardly any effect on the spectral information of the incoming signal. This lack of influence suggested that back-scattering SWIR originates from the upper part of irradiated tissue. The idea was supported by slightly increased NDLWI1937 values for permanently overlapping parts of grapevine leaves where the lower tissue impairs the water loss through transpiration of the upper tissue.
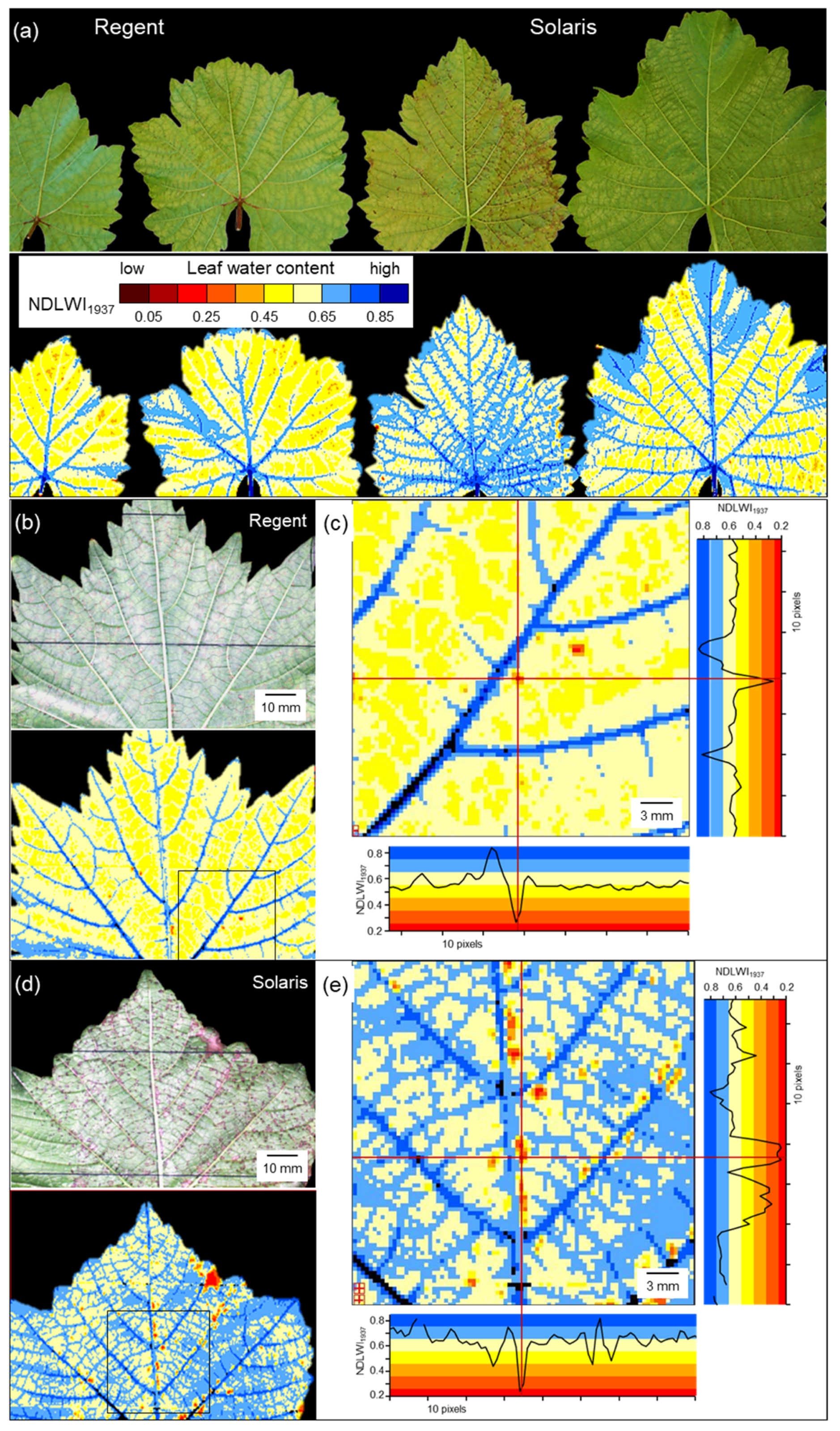
4.2. Effects of Leaf Side
Adaxial leaf sides of grapevine leaves had significantly higher mean NDLWI
1937 values than corresponding abaxial sides, irrespective of grapevine genotype and health status of living plant tissue. This result confirmed earlier observations on apple leaves [
25]. As valid for all cultivars, this observation was independent of leaf thickness. Interestingly, the difference between adaxial and abaxial tissue water content was observed only for living intercostal tissue. Necrotic tissue did not exhibit this difference.
A more detailed data analysis confirmed this effect for the intercostal areas, whereas NDLWI
1937 values of leaf veins were higher for the abaxial leaf side. Reflectance spectra revealed that grapevine genotype, leaf side, and host tissue reaction to
P. viticola infection affected reflectance in the NIR range (represented by R
1124) as well as at the water absorption bands. By calculating NDLWI
1937 as a proxy for the leaf water status on the tissue scale, the normalized difference index reliably eliminated spectral differences from tissue as influenced by the volume and structure of plant cells and the intercellular space typical for the bifacial leaves of dicotyledonous plant species [
15]. The higher water content of adaxial leaf sides is likely to result from the combination of the compact cell layer of the palisade parenchyma, rich in cytoplasm and metabolically highly active, and the effectiveness of an intact cuticle as a water barrier. The spongy parenchyma of the adaxial leaf side, in contrast, comprises a high portion of intercellular spaces responsible for the allocation of CO
2 and O
2 within tissue and is the site where constantly liquid water is transformed into water vapor, which escapes the leaf through the stomata of the abaxial epidermal layer.
Vascular strands are located below the palisade parenchyma and protrude the abaxial surface of intercostal areas, significantly in the case of lower-order veins (
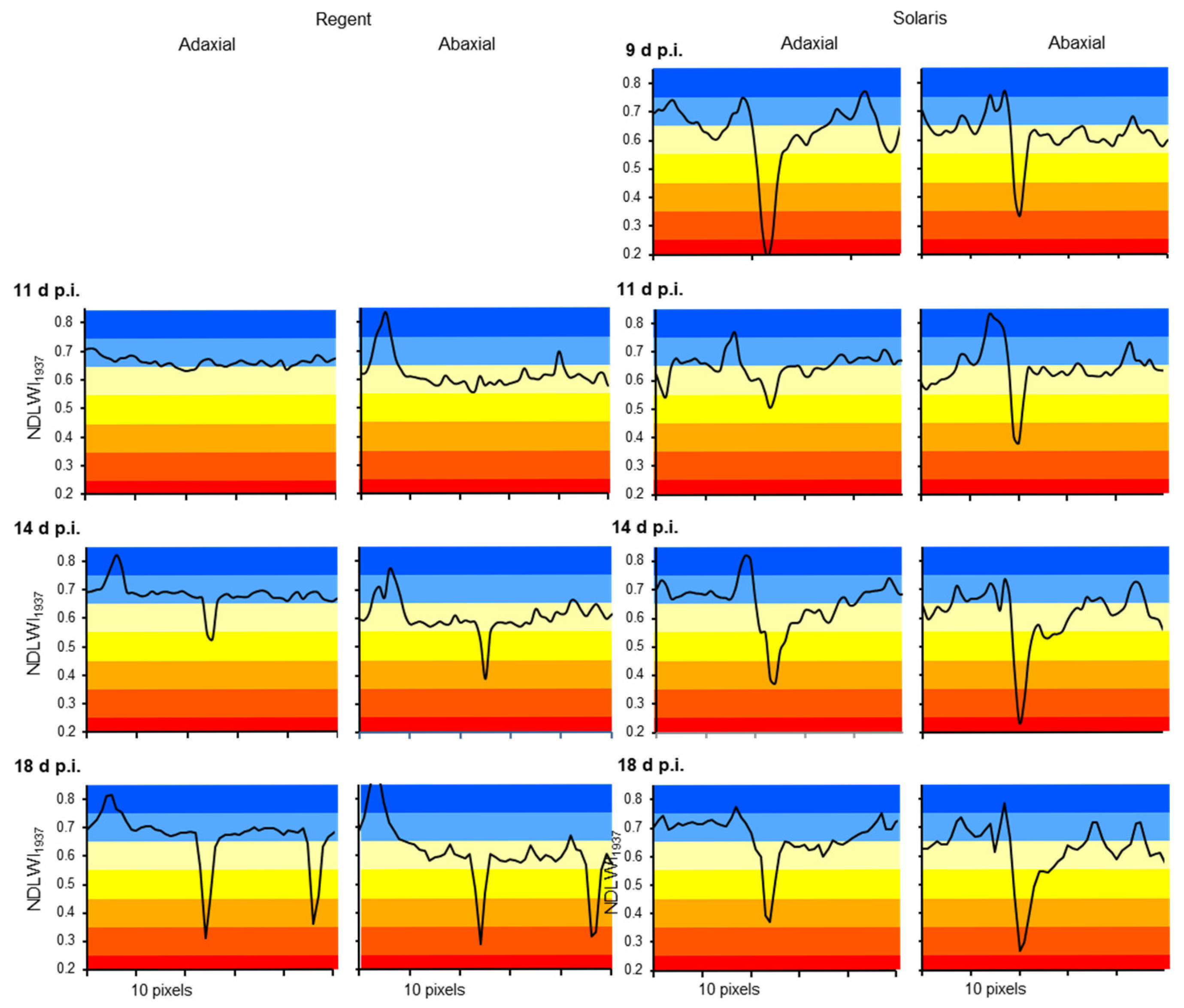
Figure S2). Localization, specific morphology (pear-shaped, prominent abaxial side considerably wider than adaxial side), and histological organization (less pronounced bifacial character, lack of stomata, large parenchymatic cells below the vascular bundle) of veins within the leaf plain result in a higher abaxial water content of veins. The higher contrast between veins and intercostal areas for the abaxial side confirms the fine differences in water content on a small scale. The ability to measure LWC differences between both sides of the same leaf can be explained only by the fact that reflectance from tissue near the surface directed to the sensor has a significantly higher contribution to the overall signal than lower tissue.
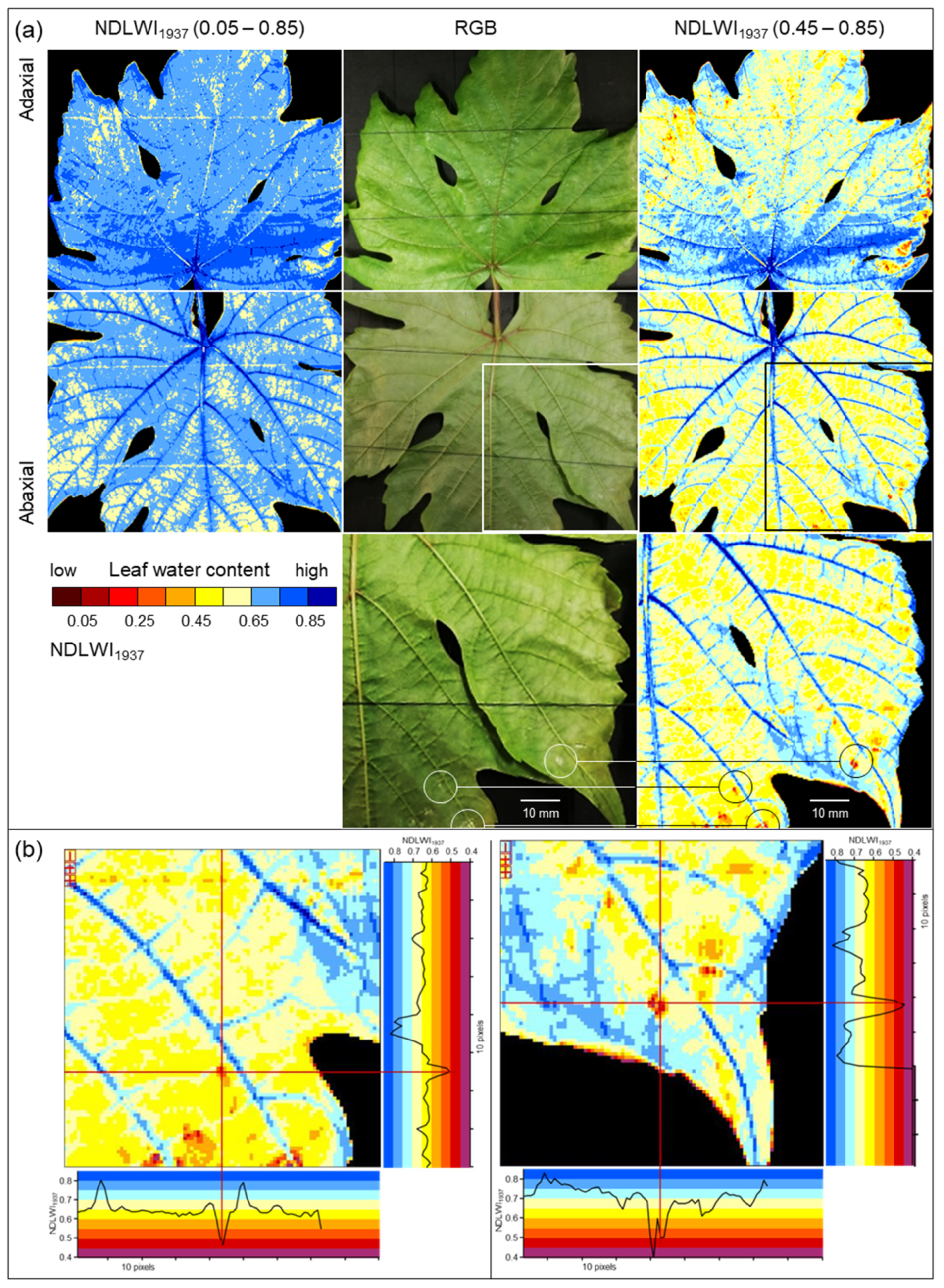
4.3. Spatial Resolution of Spectral Information
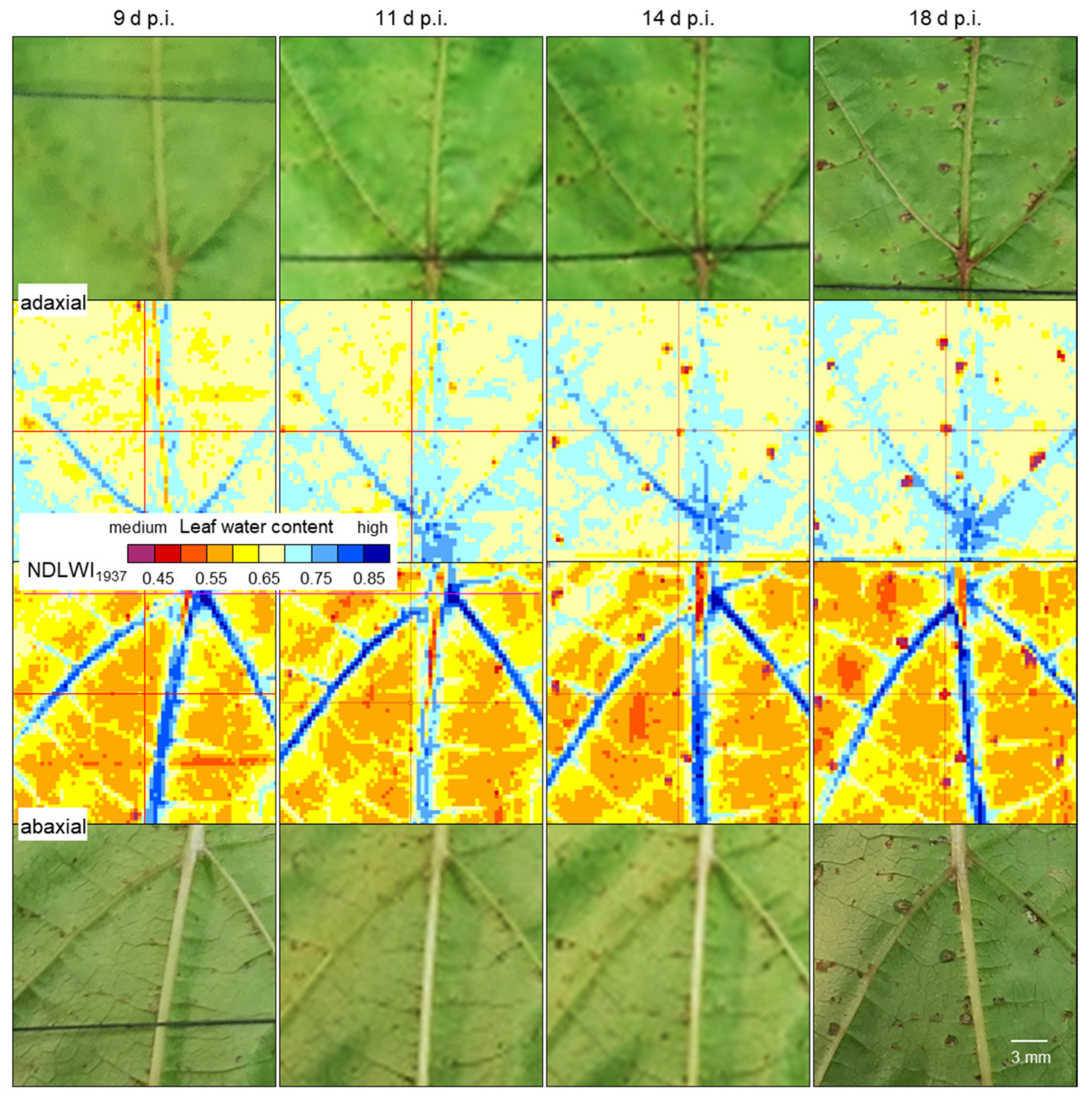
Compared to RGB cameras and hyperspectral sensor systems working in the VISNIR range, the spatial resolution of SWIR cameras is often considerably lower. This discrepancy may result in uncertainty whether no differences in SWIR data really indicate no effect on leaf water status or whether the effect is too small to be detected because of technical limitations. Spatial resolution limits the sensing system’s capability to detect small deviations from the normal status (healthy), i.e., in the case of plant pathology, to detect tiny primary disease symptoms with a diameter often <1 mm. Spectral anomaly may be small in spatial dimension as well as in spectral reflectance: The important role of spatial resolution in the detection and monitoring of brown leaf spots, i.e., the resistance reaction in interactions with reduced compatibility to
P. viticola infection, highlighted that it also limits sensor sensitivity. The spatial resolution of the sensor (with squared sensing units) has to be high; the pixel size of reflectance images has to be smaller than a third of the diameter of the usually circular symptoms of pathogen infection. Only when spatial resolution was adequate the full dimension of leaf water reduction due to the tiny spots of brown leaf tissue could be assessed (
Figure 13). For large-scale symptoms (of downy mildew sporangia or disease symptoms from other pathogens) or factors like nutrient supply, which affect all plant parts similarly, spatial resolution is not critical, and effects on plant water status may be detectable using non-imaging devices.
The highest spatial resolution of the SWIR camera was not only suitable to detect the tiny brown tissue spots in response to
P. viticola infection, but also to quantify and differentiate between response levels. Brown tissue spots had NDLWI
1937 values lower than grapevine tissue with abaxial downy mildew symptoms; however, levels were often higher than those of leaf necroses and water status of leaf spots varied with the time of pathogenesis, sometimes in both directions. This information is in agreement with observations of the little, but consistent development of biotrophic
P. viticola in these brown spots of cvs. Regent and Solaris, which also results in pathogen sporulation and spread [
37]. The transient resurge in the water content of cv. Solaris spots also indicated that brownish tissue differs from necroses and may represent a combination of necrotic and living cells. The area of brown spots visible in the VIS range differed from the area with modified tissue water content, stressing the difference between visible symptom and the effect on the water status as assessed from SWIR information.
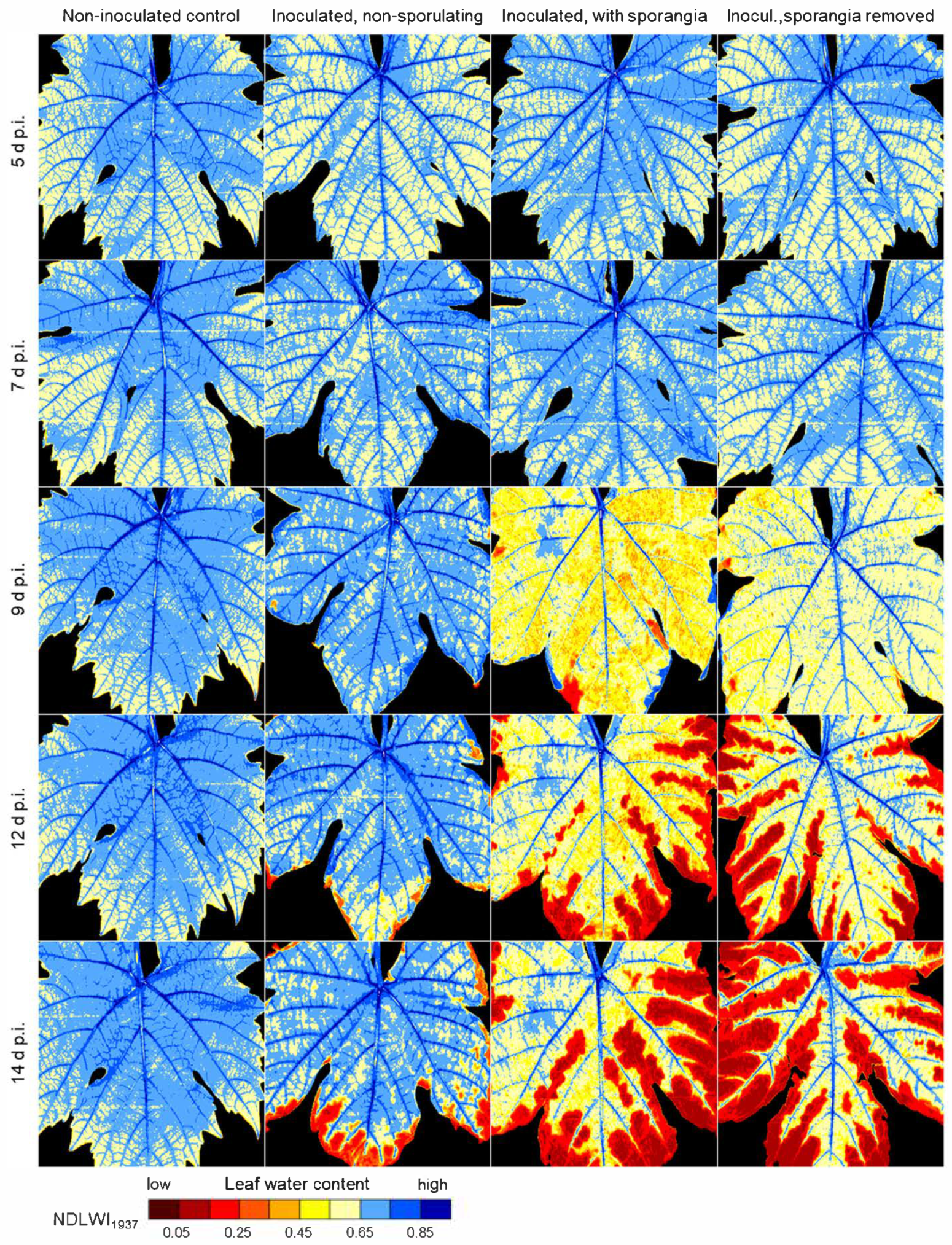
4.4. Leaf Water Response to P. viticola Infection
As expected, the leaf water status of grapevine leaves infected by the downy mildew pathogen P. viticola depended on the compatibility of the interaction. For partial resistant grapevine genotypes, infections resulted in the formation of tiny spots of brownish tissue associated with highly localized considerable LWC reduction; however, as the affected area generally remained small and LWC of non-infected tissue was hardly affected, the leaves maintained greenness and functionality. Disease resistance, though only partial, successfully protected leaves from severe damage and safeguarded their contribution to overall plant development.
The
Rpv3-1-mediated resistance of cv. “Regent” is associated with a defense mechanism that triggers the synthesis of fungi-toxic stilbenes and programmed cell death [
45]. Cultivar “Solaris” confers resistance to
P. viticola associated with necrosis formation, callose deposition, and stilbene accumulation [
46]. In the early stages of pathogenesis in cv. Solaris, the collapse of infected palisade cells largely restricts the expansion of
P. viticola to the spongy parenchyma, where tissue colonization ultimately may result in the formation of sporangiophores with sporangia on the abaxial leaf surface [
37,
47]. The formation of callose and amorphous material on stomata has been reported to impede
P. viticola sporulation on cv. Solaris [
47,
48]. Reduced or delayed sporulation may also (transiently) limit the effect on leaf water status of brownish leaf spots as observed 11 d p.i. in this study.
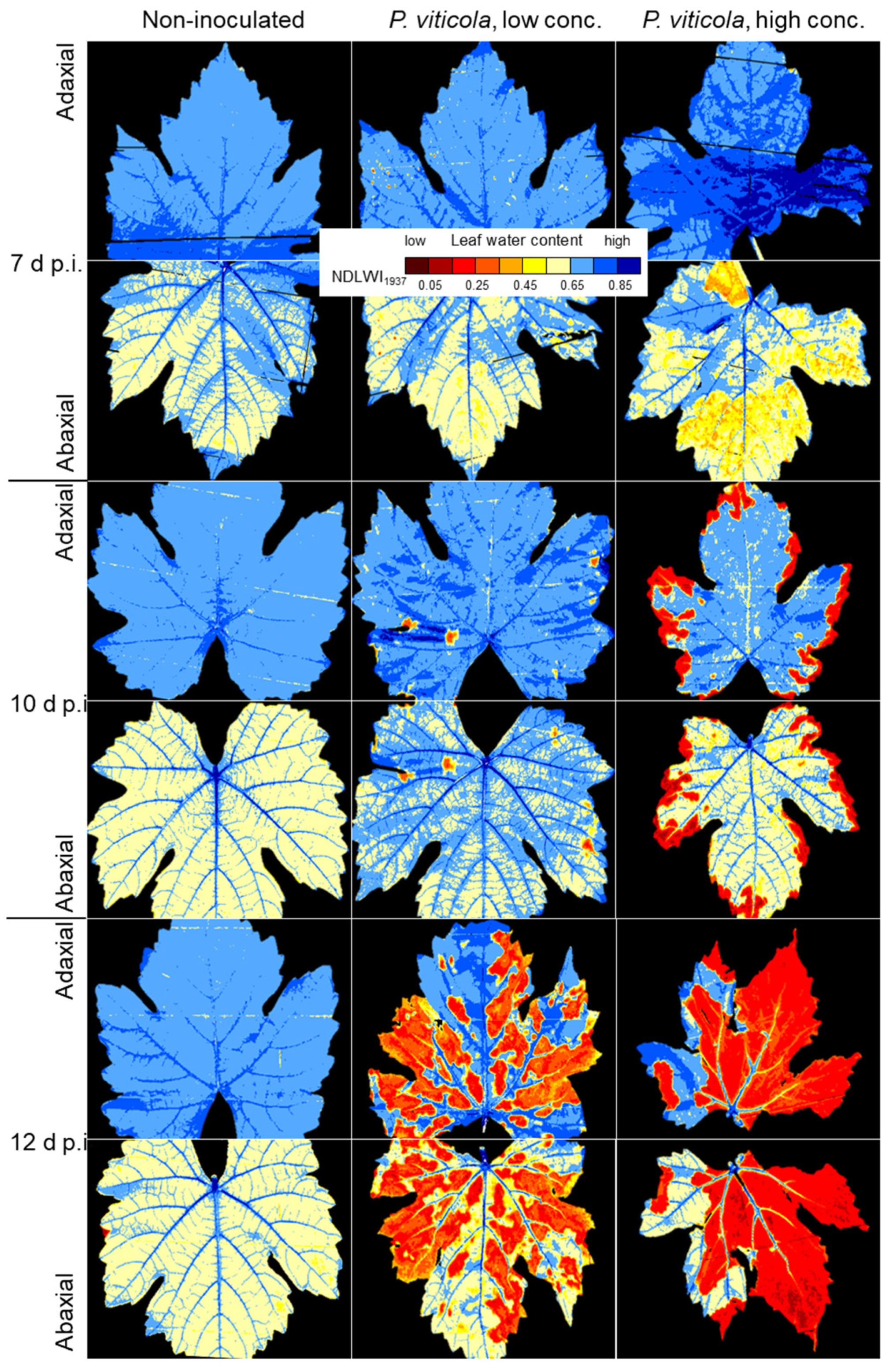
In the compatible interaction, the first effects of P. viticola on the LWC pattern of leaves coincided with first sporangia formation or, without the induction of sporulation by high RH, with strong colonization (and production of pathogen biomass) of leaf tissue. The effect on leaf water status, initially limited to sites of primary sporangia, increased with the density of sporulation and caused large-scale LWC reduction of colonized leaf tissue and ultimately resulting in tissue collapse. Although necrotic tissue had NDLWI values of 0.2 and lower, xylem and phloem retained their functionality as long as possible.
The reduced LWC of leaf tissue with abundant sporangia formation revealed a rather uniform and large-scale effect of P. viticola on the water status of the susceptible cv. Mueller-Thurgau. Although non-diseased intercostal areas of infected leaves were not affected, the high number of sporangiophores with sporangia protruding through stomata had a wicking activity on apoplastic leaf water and resulted in LWC values 10 to 20% lower than those of non-diseased leaves. As sporangiophores prevented diurnal closure of stomata, the slow but continuing loss of water led to a dehydration of intercostal areas, which started at leaf margins and expanded to central leaf parts in later stages of pathogenesis; leaf veins remained functional for longer periods of time. The pathogen biomass withdraws water (and nutrients) from the apoplast and (via haustoria) from the plant symplast and transfers it to ambient air, to the plant’s disadvantage. Without another spell of high RH of ambient air, P. viticola sporangia dried within 24 h, but preserved pathogenicity for several days. The large surface of sporangia and sporangiophores protruding into the relatively dry ambient air explains the severe effect of downy mildew on leaf vitality observed in vineyards, which may be even aggravated by insufficient water supply from the soil.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}