
Spatiotemporal Changes of Pine Caterpillar Infestation Risk and the Driving Effect of Habitat Factors in Northeast China

 , , and
, , and
Abstract

1. Introduction
2. Study Area and Data
2.1. Study Area
2.2. Data Source
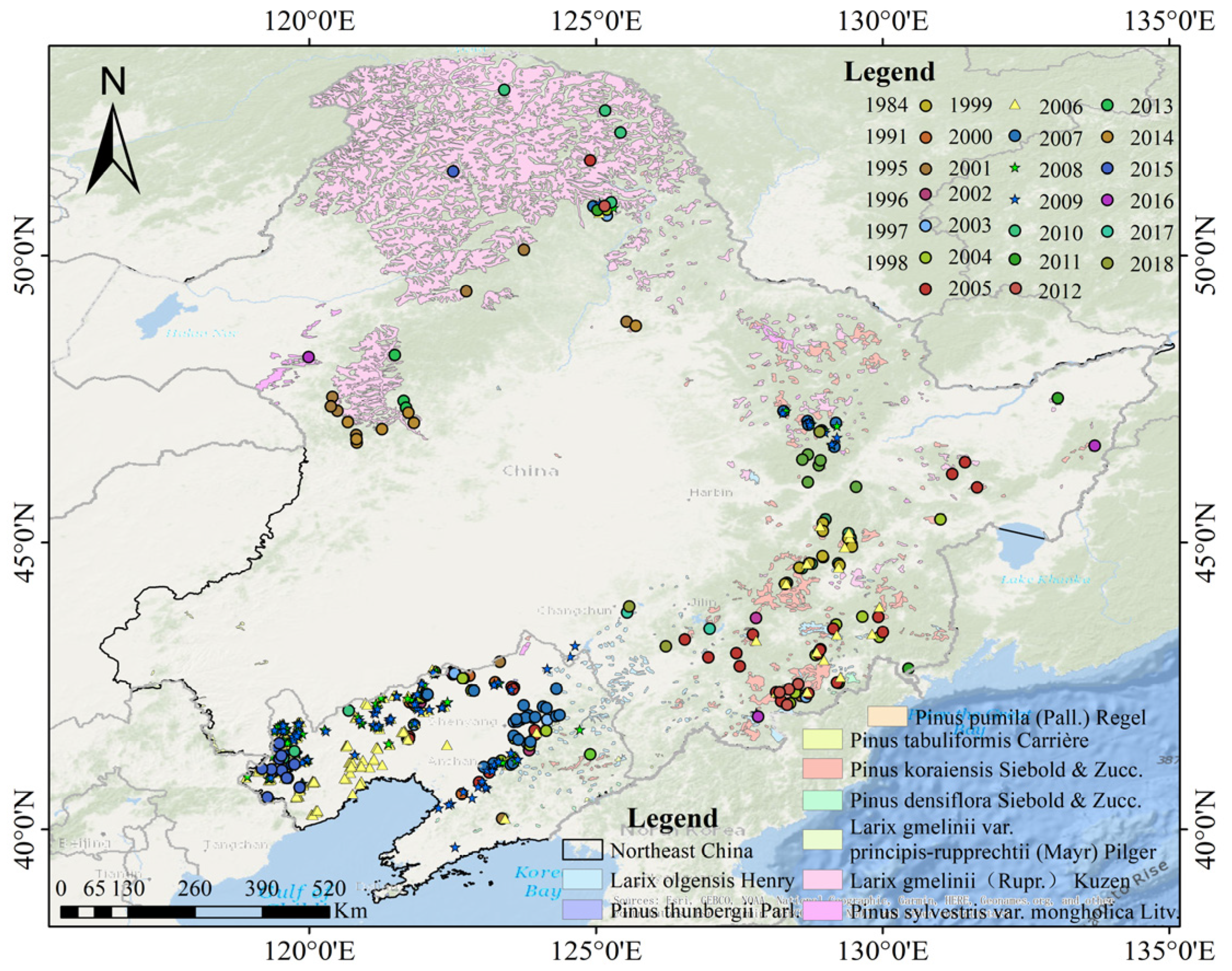
2.2.1. Distribution Points of Pine Caterpillar Infestation and Non-Infestation
2.2.2. Habitat Factor Data
2.3. Shapiro–Wilk Test
2.4. Risk Assessment and Habitat Factor Analysis Methods
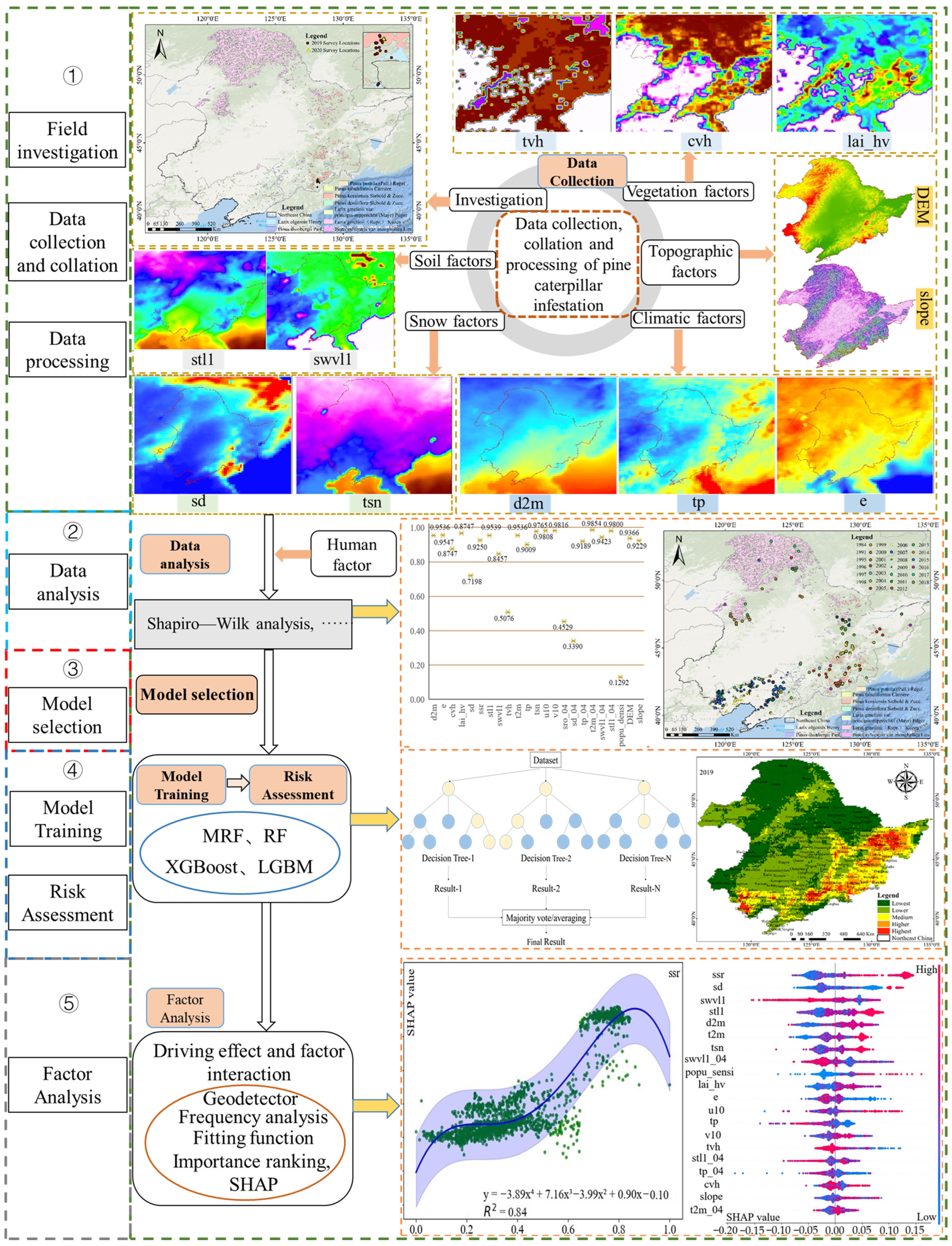
2.4.1. Conceptual Framework
2.4.2. The Risk Assessment Model
2.4.3. SHAP and Fitting Function
2.4.4. Frequency Analysis
2.4.5. GeoDetector
3. Results and Analysis
3.1. Comparison of Accuracy of Different Models
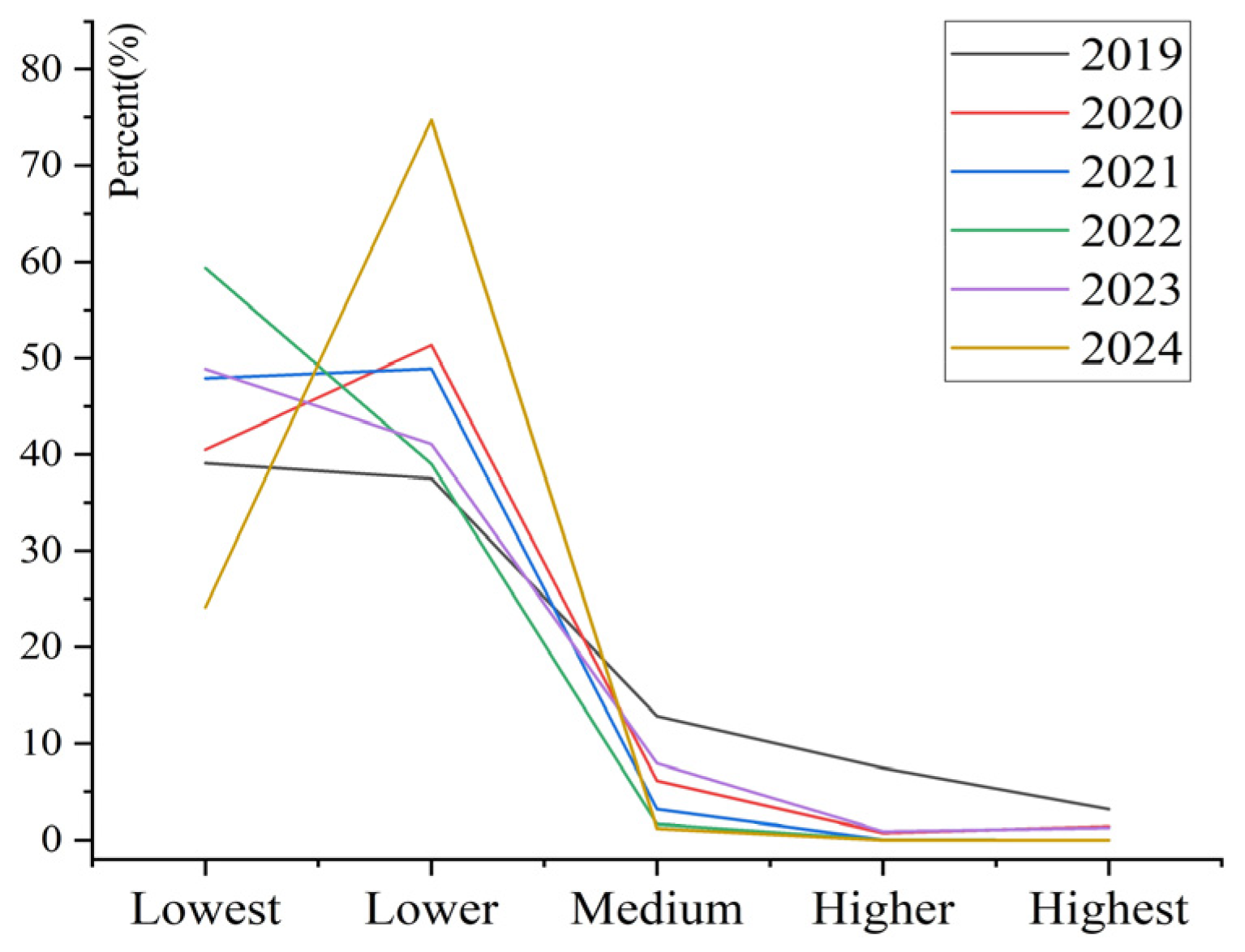
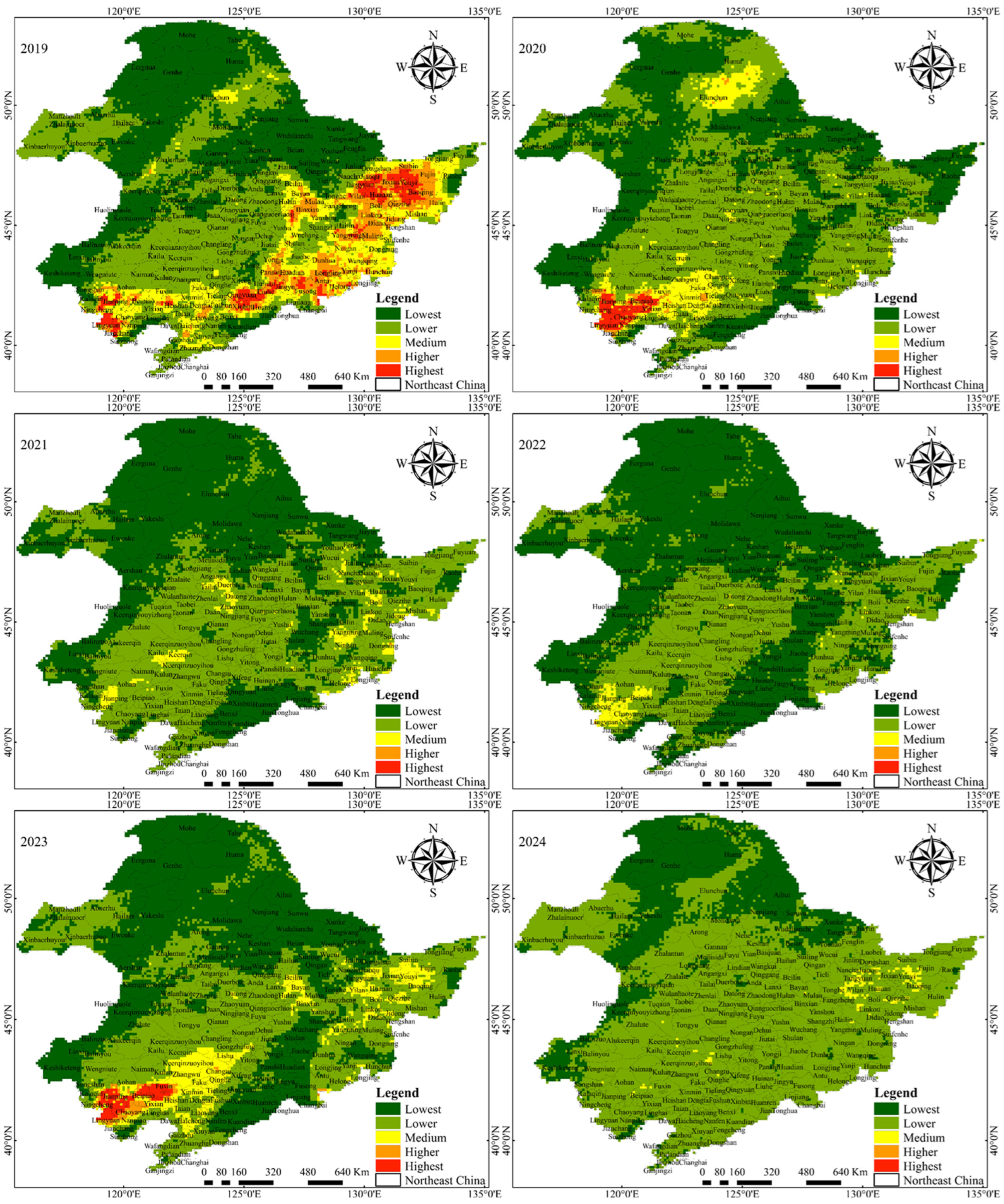
3.2. Spatiotemporal Changes of Infestation Risk Levels of Pine Caterpillar
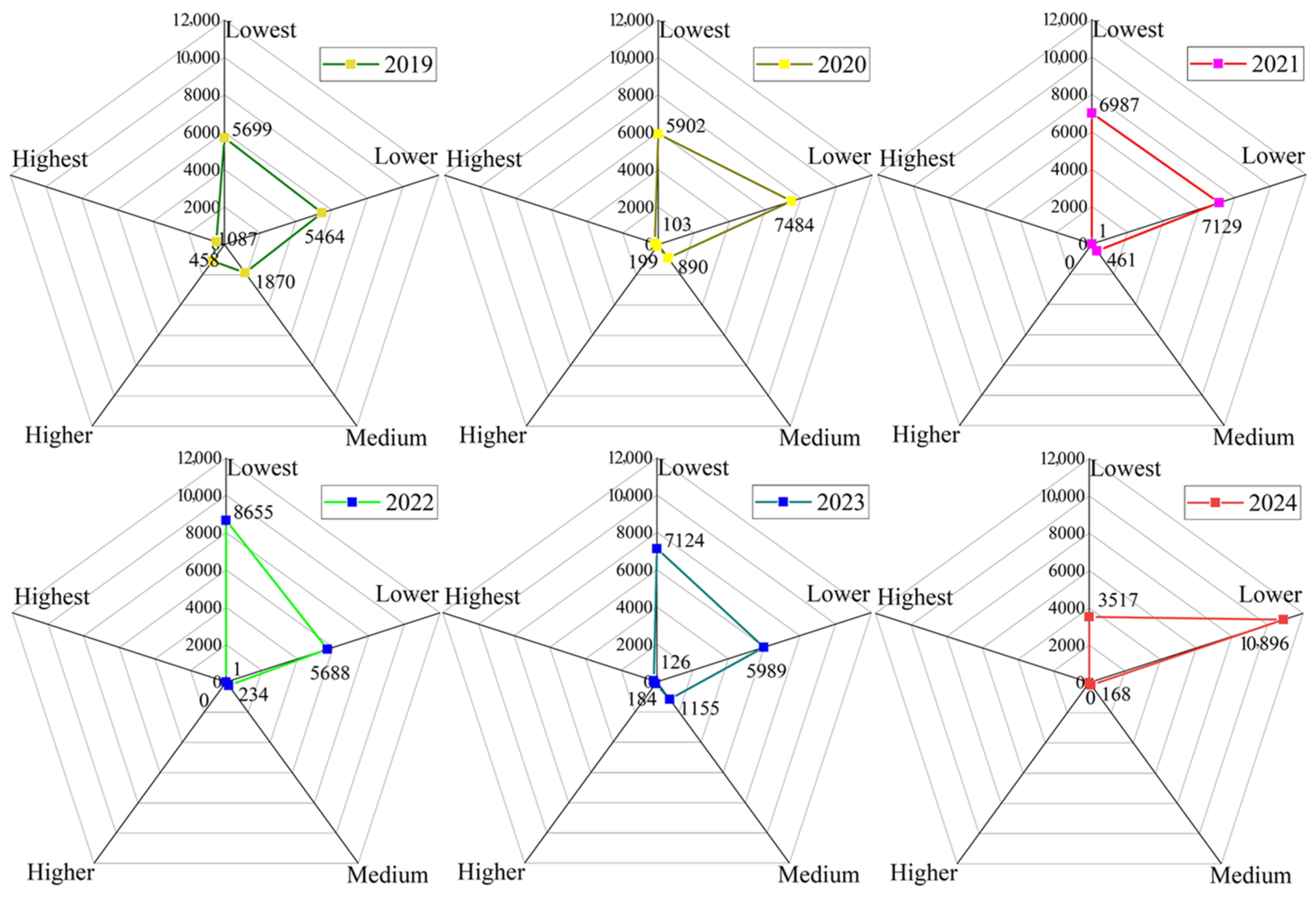
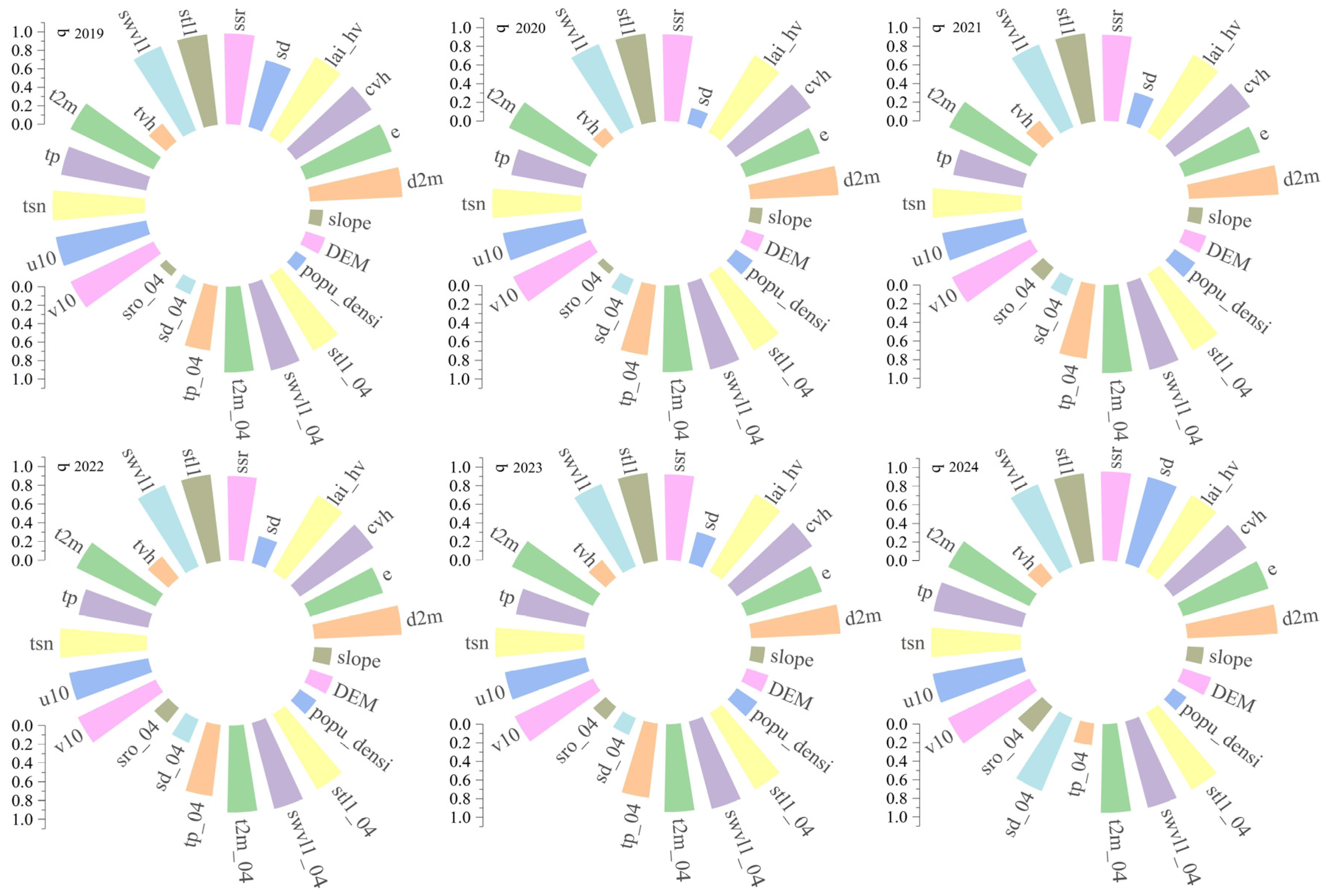
3.3. Frequency Analysis of Infestation Risk Levels of Pine Caterpillar
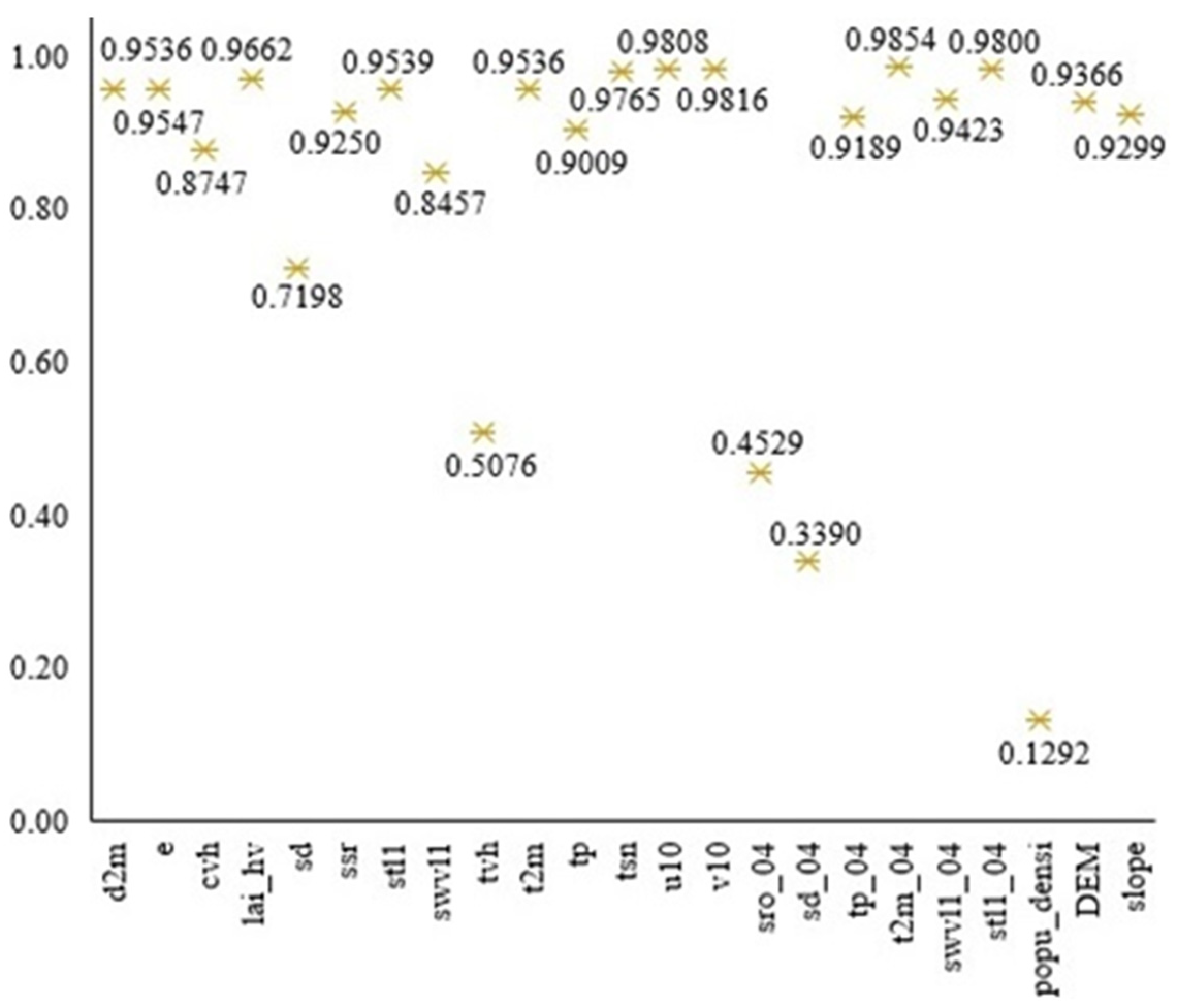
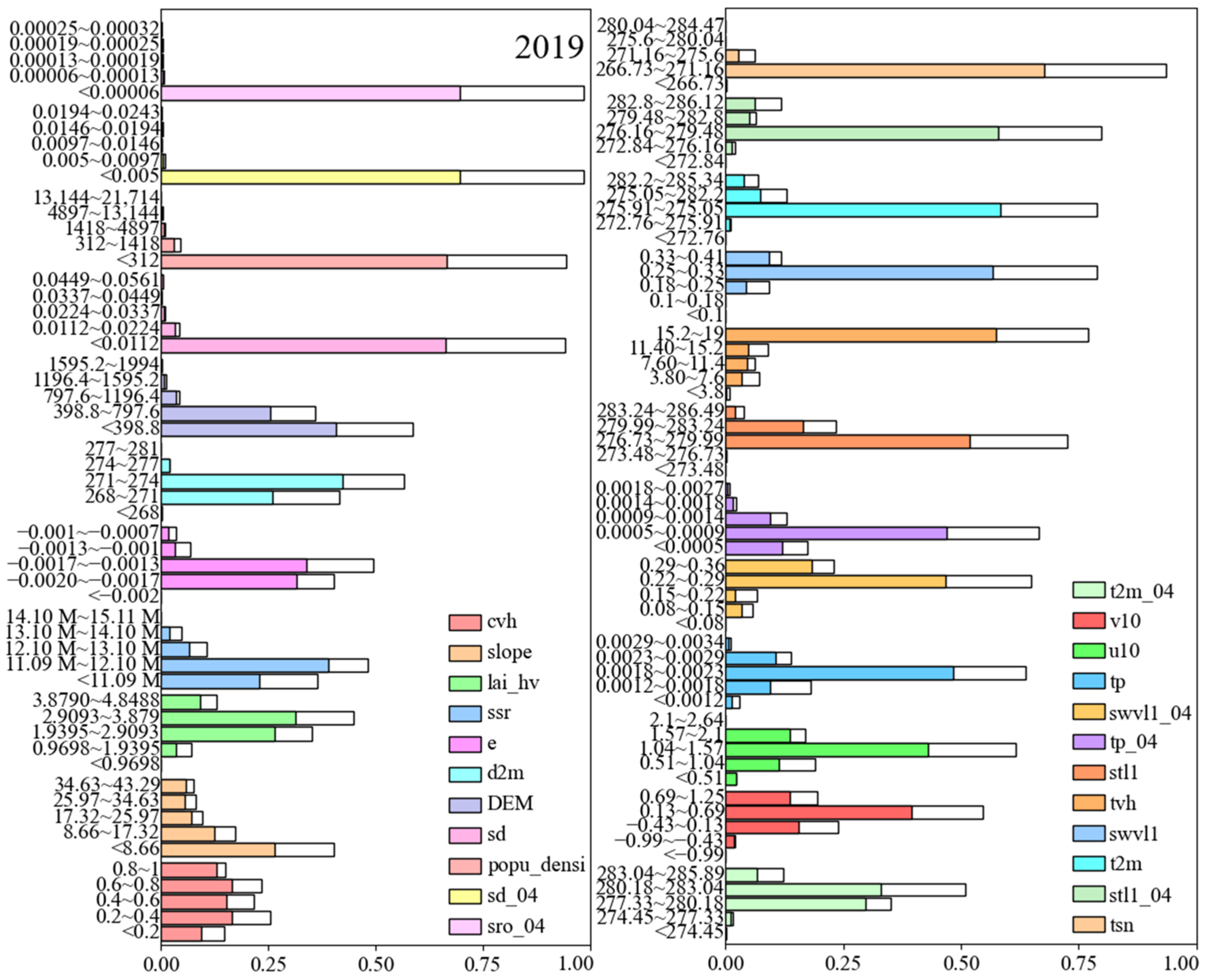
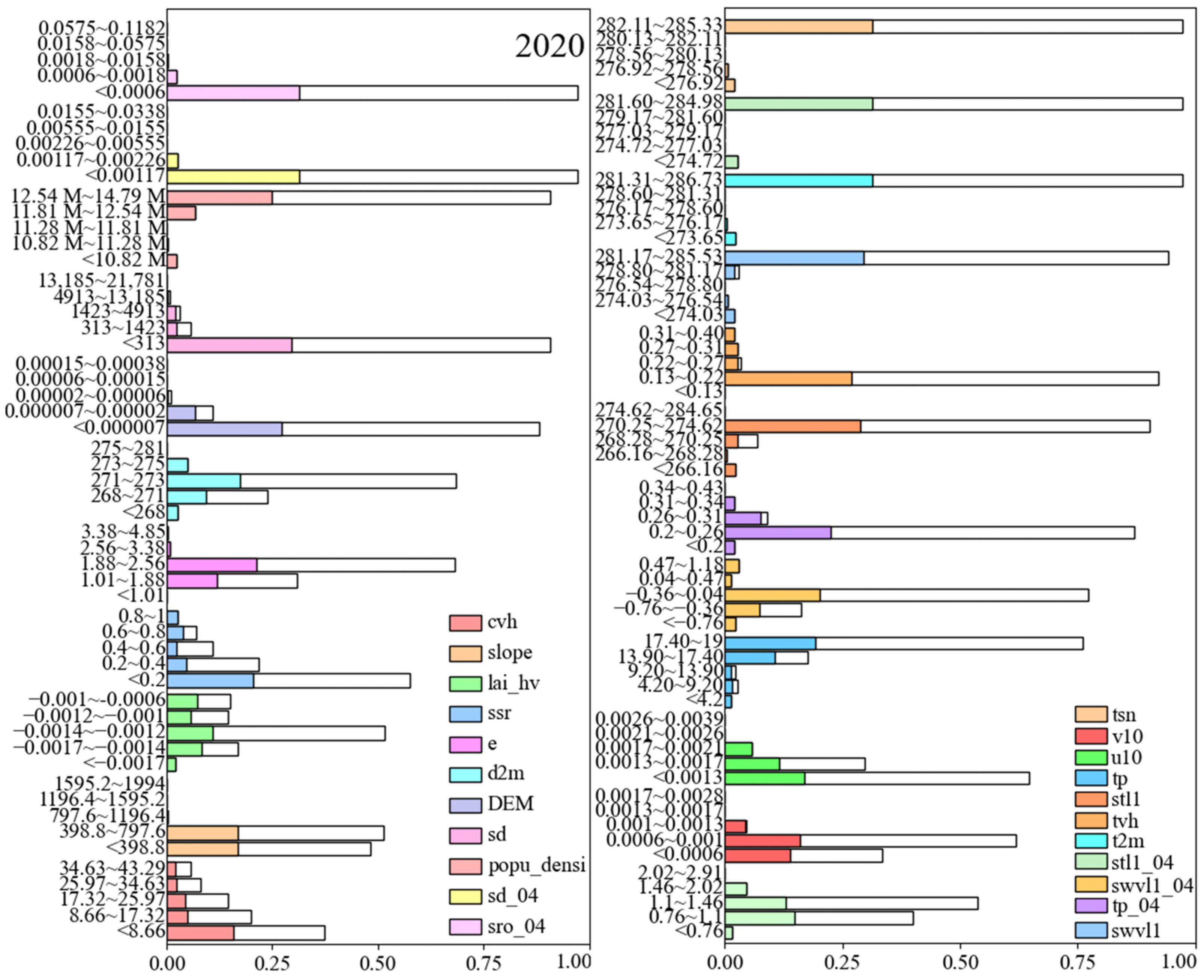
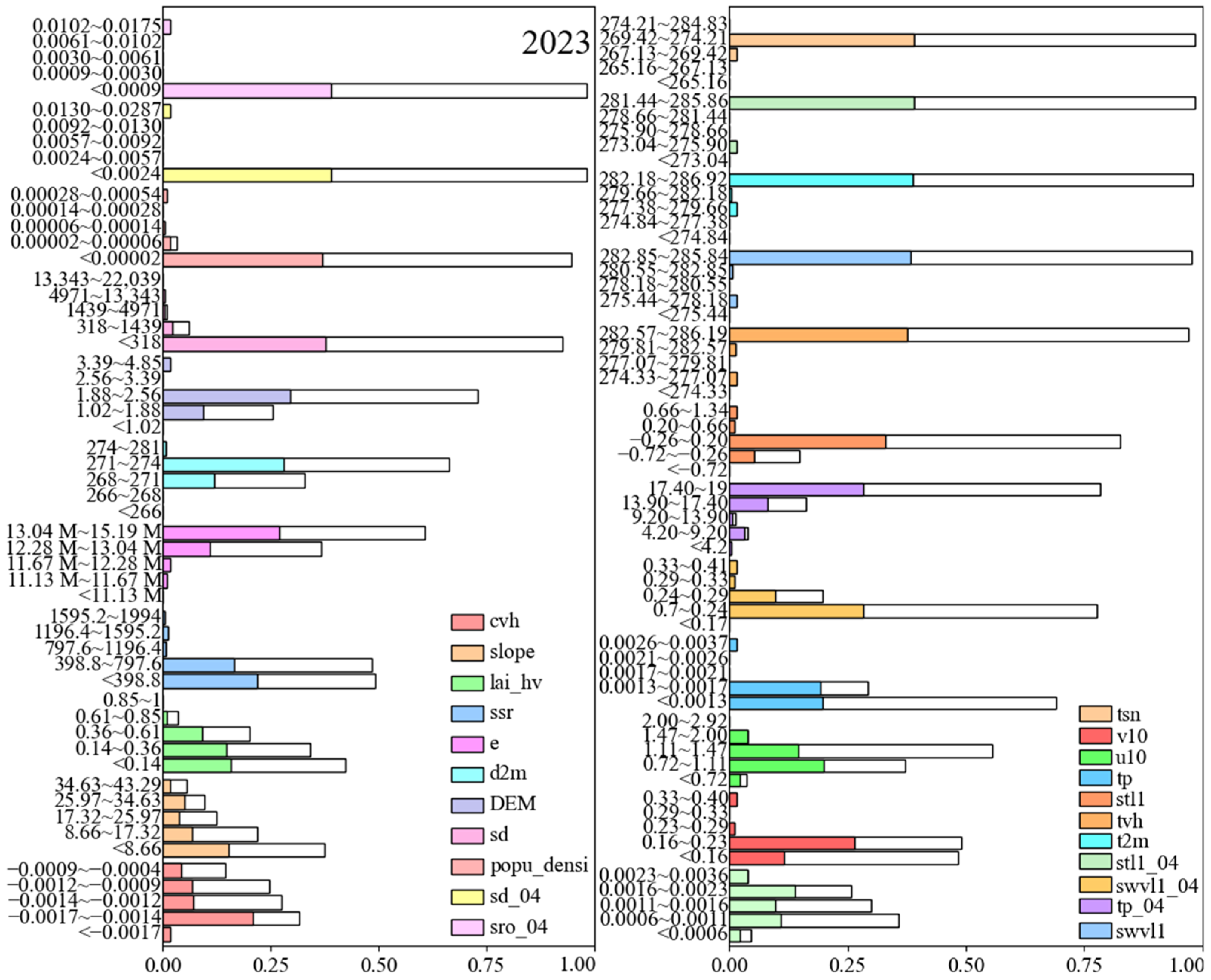
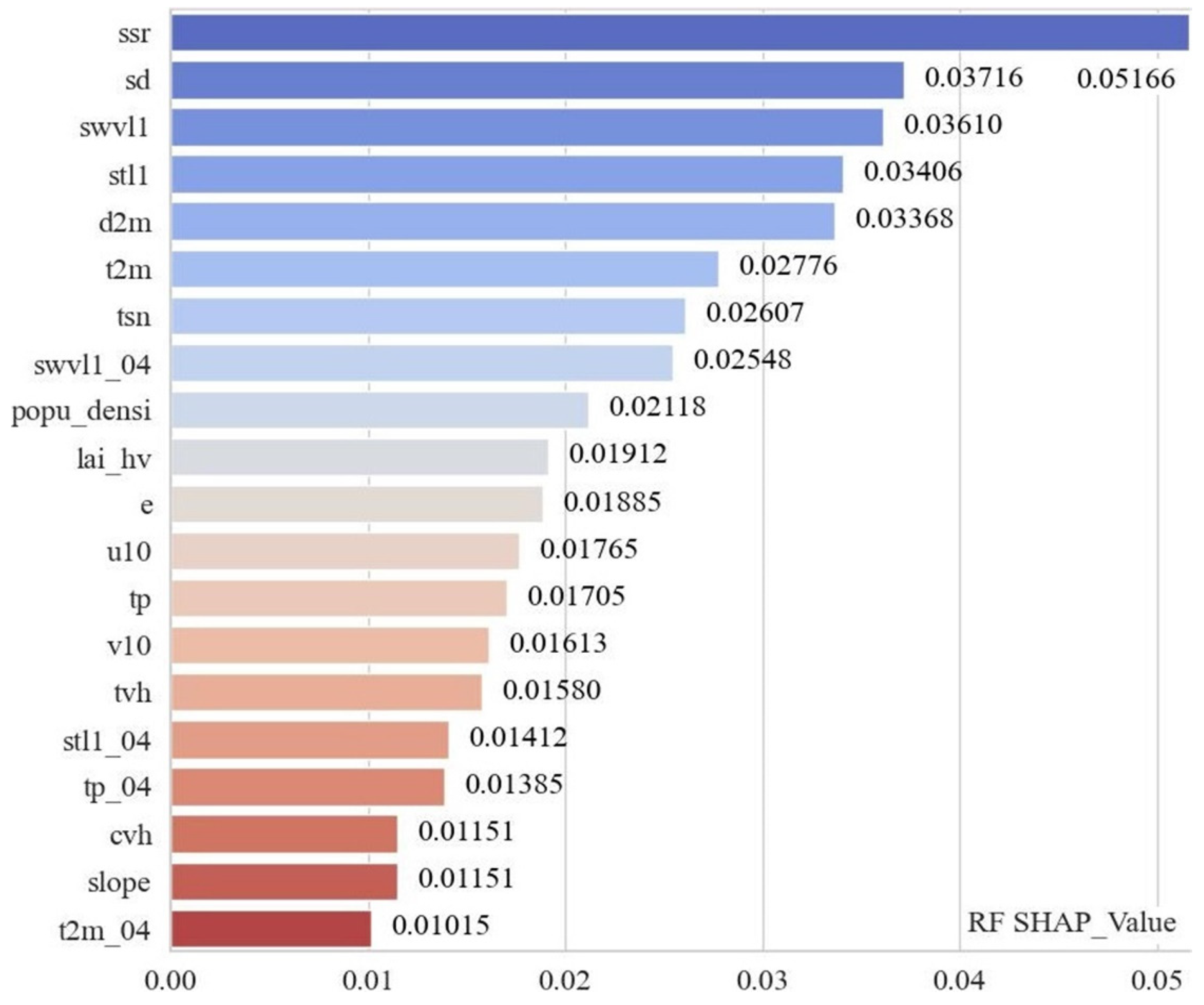
3.4. Identification of Key Habitat Factors
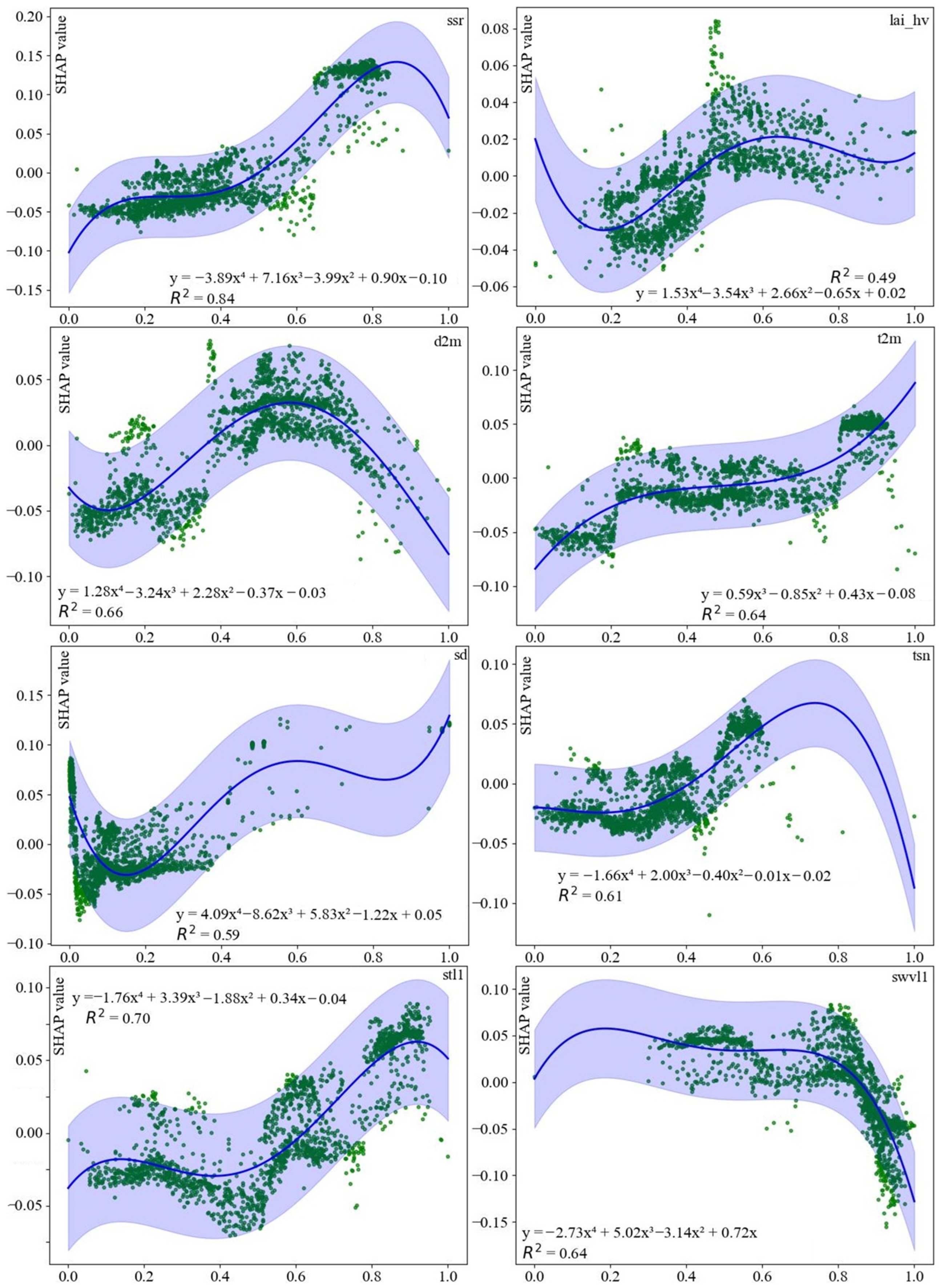
3.5. SHAP and Fitting Function Analysis
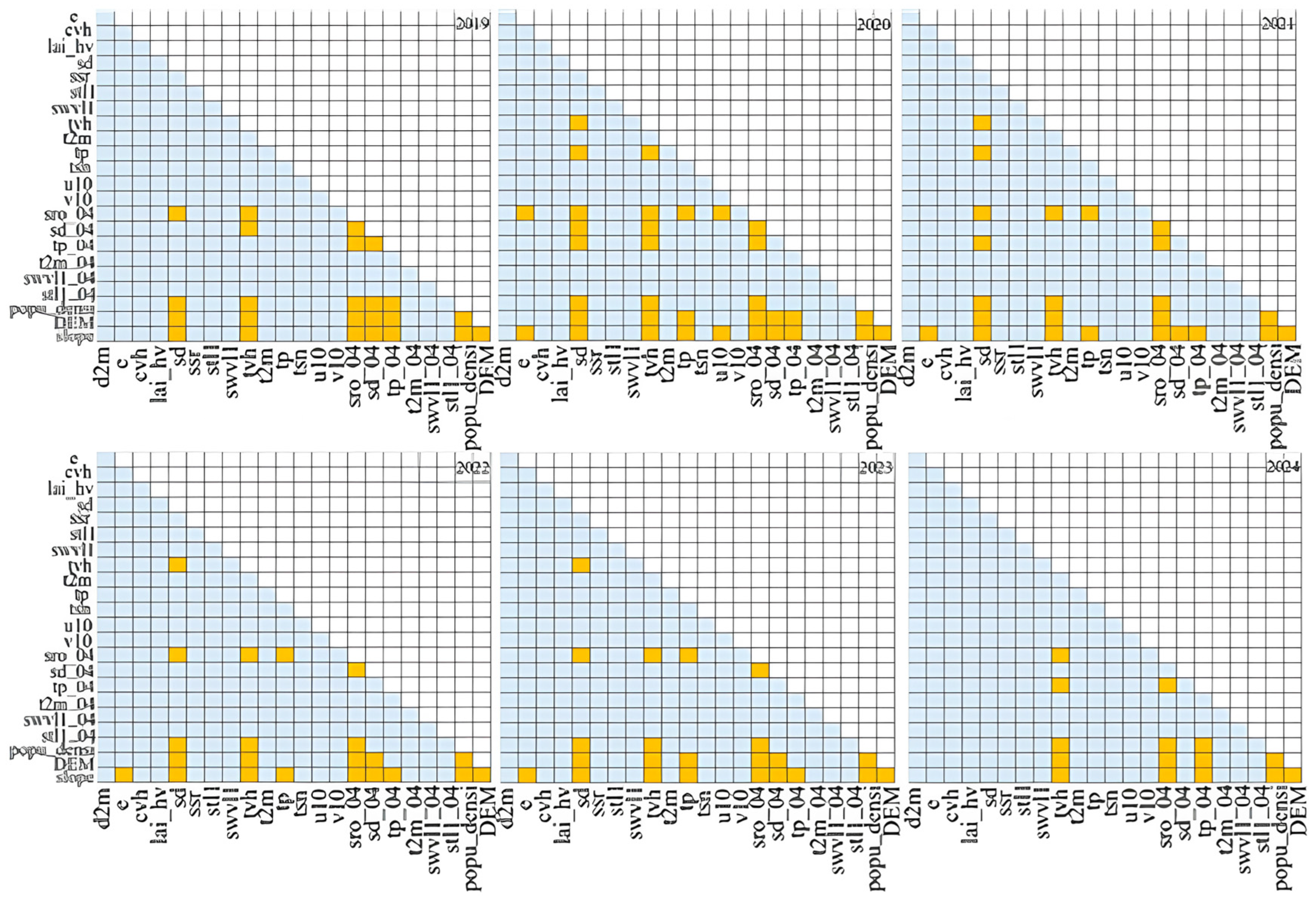
3.6. Interaction Detector Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Grêt-Regamey, A.; Weibel, B. Global Assessment of Mountain Ecosystem Services Using Earth Observation Data. Ecosyst. Serv. 2020, 46, 101213. [Google Scholar] [CrossRef]
- Xie, Y.; Cheng, C.; Zhang, T.; Wu, X.; Wang, P. Donor-Side Valuation of Forest Ecosystem Services in China during 1990–2020. Energy Ecol. Environ. 2023, 8, 503–521. [Google Scholar] [CrossRef]
- Cortini, F.; Comeau, P.G. Pests, Climate and Competition Effects on Survival and Growth of Trembling Aspen in Western Canada. New For. 2020, 51, 175–190. [Google Scholar] [CrossRef]
- Han, D.; Wang, S.; Zhang, J.; Cui, R.; Wang, Q. Evaluating Dendrolimus superans (Lepidoptera: Lasiocampidae) Occurrence and Density Modeling with Habitat Conditions. Forests 2024, 15, 388. [Google Scholar] [CrossRef]
- Schroeder, M.; Cocoş, D. Performance of the Tree-Killing Bark Beetles Ips typographus and Pityogenes chalcographus in Non-Indigenous Lodgepole Pine and Their Historical Host Norway Spruce. Agric. For. Entomol. 2018, 20, 347–357. [Google Scholar] [CrossRef]
- Chen, H.; Hu, Y.; Chang, Y.; Bu, R.; Li, Y.; Liu, M. Simulating Impact of Larch Caterpillar (Dendrolimus superans) on Fire Regime and Forest Landscape in Da Hinggan Mountains, Northeast China. Chin. Geogr. Sci. 2011, 21, 575–586. [Google Scholar] [CrossRef]
- Cheng, X.; Qian, G.; Song, X.; Zhang, S.; Zhou, X.; Zou, Y.; Zhang, G.; Fang, G.; Song, Y.; Bi, S. The Catastrophe Prediction Models of Dendrolimus punctatus Based on Disaster Index. Int. J. Pest Manag. 2021, 70, 616–625. [Google Scholar] [CrossRef]
- Wu, S.J.; Zhu, T.H.; Qiao, T.M.; Li, S.J.; Shan, H. Prediction of the Potential Distribution of Dendrolimus Houi Lajonquiere in Sichuan of China Based on the Species Distribution Model. Appl. Ecol. Environ. Res. 2021, 19, 2227–2240. [Google Scholar] [CrossRef]
- Bao, Y.; Han, A.; Zhang, J.; Liu, X.; Tong, Z.; Bao, Y. Contribution of the Synergistic Interaction between Topography and Climate Variables to Pine Caterpillar (Dendrolimus spp.) Outbreaks in Shandong Province, China. Agric. For. Meteorol. 2022, 322, 109023. [Google Scholar] [CrossRef]
- Bao, Y.; Na, L.; Han, A.; Guna, A.; Wang, F.; Liu, X.; Zhang, J.; Wang, C.; Tong, S.; Bao, Y. Drought Drives the Pine Caterpillars (Dendrolimus spp.) Outbreaks and Their Prediction under Different RCPs Scenarios: A Case Study of Shandong Province, China. For. Ecol. Manag. 2020, 475, 118446. [Google Scholar] [CrossRef]
- You, W.; You, H.; Wu, L.; Ji, Z.; He, D. Landscape-Level Spatiotemporal Patterns of Dendrolimus punctatus Walker and Its Driving Forces: Evidence from a Pinus massoniana Forest. Trees-Struct. Funct. 2020, 34, 553–562. [Google Scholar] [CrossRef]
- Liu, N.; Zhao, X.; Zhang, X.; Zhao, J.; Wang, H.; Wu, D. Remotely Sensed Evidence of the Divergent Climate Impacts of Wind Farms on Croplands and Grasslands. Sci. Total Environ. 2023, 905, 167203. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Tang, Y.; Wang, Z.; Peng, C.; Huang, C.; Dian, Y.; Teng, M.; Zhou, Z. Seasonal Variations of the Relationship between Spectral Indexes and Land Surface Temperature Based on Local Climate Zones: A Study in Three Yangtze River Megacities. Remote Sens. 2023, 15, 870. [Google Scholar] [CrossRef]
- Xiang, Y.; Yuan, C.; Cen, Q.; Huang, C.; Wu, C.; Teng, M.; Zhou, Z. Heat Risk Assessment and Response to Green Infrastructure Based on Local Climate Zones. Build. Environ. 2024, 248, 111040. [Google Scholar] [CrossRef]
- Han, R.D.; Parajulee, M.; Zhong, H.; Feng, G. Effects of Environmental Humidity on the Survival and Development of Pine Caterpillars, Dendrolimus tabulaeformis (Lepidoptera: Lasiocampidae). Insect Sci. 2008, 15, 147–152. [Google Scholar] [CrossRef]
- Fang, L.; Yu, Y.; Fang, G.; Zhang, X.; Yu, Z.; Zhang, X.; Crocker, E.; Yang, J. Effects of Meteorological Factors on the Defoliation Dynamics of the Larch Caterpillar (Dendrolimus superans Butler) in the Great Xing’an Boreal Forests. J. For. Res. 2021, 32, 2683–2697. [Google Scholar] [CrossRef]
- Hua, H.; Wu, C.; Jassal, R.S.; Huang, J.; Liu, R.; Wang, Y. Pine Caterpillar Occurrence Modeling Using Satellite Spring Phenology and Meteorological Variables. Environ. Res. Lett. 2022, 17, 104046. [Google Scholar] [CrossRef]
- Gao, H.; Wang, C.; Wang, G.; Zhu, J.; Tang, Y.; Shen, P.; Zhu, Z. A Crop Classification Method Integrating GF-3 PolSAR and Sentinel-2A Optical Data in the Dongting Lake Basin. Sensors 2018, 18, 3139. [Google Scholar] [CrossRef]
- Mercier, A.; Betbeder, J.; Denize, J.; Roger, J.L.; Spicher, F.; Lacoux, J.; Roger, D.; Baudry, J.; Hubert-Moy, L. Estimating Crop Parameters Using Sentinel-1 and 2 Datasets and Geospatial Field Data. Data Brief 2021, 38, 107408. [Google Scholar] [CrossRef]
- Zheng, Q.; Huang, W.; Cui, X.; Shi, Y.; Liu, L. New Spectral Index for Detecting Wheat Yellow Rust Using Sentinel-2 Multispectral Imagery. Sensors 2018, 18, 868. [Google Scholar] [CrossRef]
- Booth, T.H. Checking Bioclimatic Variables That Combine Temperature and Precipitation Data before Their Use in Species Distribution Models. Austral Ecol. 2022, 47, 1506–1514. [Google Scholar] [CrossRef]
- Soria-Auza, R.W.; Kessler, M.; Bach, K.; Barajas-Barbosa, P.M.; Lehnert, M.; Herzog, S.K.; Böhner, J. Impact of the Quality of Climate Models for Modelling Species Occurrences in Countries with Poor Climatic Documentation: A Case Study from Bolivia. Ecol. Model. 2010, 221, 1221–1229. [Google Scholar] [CrossRef]
- Archaux, F.; Bergès, L. Optimising Vegetation Monitoring. A Case Study in A French Lowland Forest. Environ. Monit. Assess. 2008, 141, 19–25. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Z.; Wang, W.; Fang, W.T.; Chiang, Y.T.; Liu, X.; Ju, H. Vegetation Successions of Coastal Wetlands in Southern Laizhou Bay, Bohai Sea, Northern China, Influenced by the Changes in Relative Surface Elevation and Soil Salinity. J. Environ. Manag. 2021, 293, 112964. [Google Scholar] [CrossRef]
- Christiansen, B. The Shortcomings of Nonlinear Principal Component Analysis in Identifying Circulation Regimes. J. Clim. 2005, 18, 4814–4823. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, M.; Liu, X.; Wang, F.; Li, X.; Wang, J.; Hou, G.; Zhao, S. Ecological Security Assessment and Warning of Cultivated Land Quality in the Black Soil Region of Northeast China. Land 2023, 12, 1005. [Google Scholar] [CrossRef]
- Yang, X.; Hao, Z.; Liu, K.; Tao, Z.; Shi, G. An Improved Unascertained Measure-Set Pair Analysis Model Based on Fuzzy AHP and Entropy for Landslide Susceptibility Zonation Mapping. Sustainability 2023, 15, 6205. [Google Scholar] [CrossRef]
- Campos, J.C.; Garcia, N.; Alírio, J.; Arenas-Castro, S.; Teodoro, A.C.; Sillero, N. Ecological Niche Models Using MaxEnt in Google Earth Engine: Evaluation, Guidelines and Recommendations. Ecol. Inform. 2023, 76, 102147. [Google Scholar] [CrossRef]
- Gagula, A.; Campana, M.B.D.; Narit, M.G.; Guerrero, P.D.; Parac, E.P. Using Maxent in Quantifying the Impacts of Climate Change in Land Suitability of Abaca (Musa Testilis) in Caraga Region, Philippines. In Proceedings of the 8th Geoinformation Science Symposium 2023: Geoinformation Science for Sustainable Planet, Yogyakarta, Indonesia, 28–30 August 2023. [Google Scholar] [CrossRef]
- Yalcin, M.; Sari, F.; Yildiz, A. Exploration of Potential Geothermal Fields Using MAXENT and AHP: A Case Study of the Büyük Menderes Graben. Geothermics 2023, 114, 102792. [Google Scholar] [CrossRef]
- Bera, D.; Das Chatterjee, N.; Bera, S. Comparative Performance of Linear Regression, Polynomial Regression and Generalized Additive Model for Canopy Cover Estimation in the Dry Deciduous Forest of West Bengal. Remote Sens. Appl. Soc. Environ. 2021, 22, 100502. [Google Scholar] [CrossRef]
- Park, S.Y.; Yoon, D.K.; Park, S.H.; Jeon, J.I.; Lee, J.M.; Yang, W.H.; Cho, Y.S.; Kwon, J.; Lee, C.M. Proposal of a Methodology for Prediction of Indoor PM2.5 Concentration Using Sensor-Based Residential Environments Monitoring Data and Time-Divided Multiple Linear Regression Model. Toxics 2023, 11, 526. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, M. A Comparative Assessment of the Statistical Methods Based on Urban Population Density Estimation. Geocarto Int. 2023, 38, 2152494. [Google Scholar] [CrossRef]
- Early, R.; Rwomushana, I.; Chipabika, G.; Day, R. Comparing, Evaluating and Combining Statistical Species Distribution Models and CLIMEX to Forecast the Distributions of Emerging Crop Pests. Pest Manag. Sci. 2022, 78, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Fitzgibbon, A.; Pisut, D.; Fleisher, D. Evaluation of Maximum Entropy (Maxent) Machine Learning Model to Assess Relationships between Climate and Corn Suitability. Land 2022, 11, 1382. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between Optimized MaxEnt and Random Forest Modeling in Predicting Potential Distribution: A Case Study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]
- Dai, X.; Wu, W.; Ji, L.; Tian, S.; Yang, B.; Guan, B.; Wu, D. MaxEnt Model-Based Prediction of Potential Distributions of Parnassia Wightiana (Celastraceae) in China. Biodivers. Data J. 2022, 10, e81073. [Google Scholar] [CrossRef]
- Huercha; Song, R.; Ma, Y.; Hu, Z.; Li, Y.; Li, M.; Wu, L.; Li, C.; Dao, E.; Fan, X.; et al. MaxEnt Modeling of Dermacentor marginatus (Acari: Ixodidae) Distribution in Xinjiang, China. J. Med. Entomol. 2020, 57, 1659–1667. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, L.; Song, C.; Xue, Z.; Zheng, R.; Yan, X.; Hao, C. Modelling Potential Distribution of Tuta Absoluta in China under Climate Change Using CLIMEX and MaxEnt. J. Appl. Entomol. 2023, 147, 895–907. [Google Scholar] [CrossRef]
- Song, J.W.; Jung, J.M.; Nam, Y.; Jung, J.K.; Jung, S.; Lee, W.H. Spatial Ensemble Modeling for Predicting the Potential Distribution of Lymantria dispar asiatica (Lepidoptera: Erebidae: Lymantriinae) in South Korea. Environ. Monit. Assess. 2022, 194, 889. [Google Scholar] [CrossRef]
- Bellin, N.; Tesi, G.; Marchesani, N.; Rossi, V. Species Distribution Modeling and Machine Learning in Assessing the Potential Distribution of Freshwater Zooplankton in Northern Italy. Ecol. Inform. 2022, 69, 101682. [Google Scholar] [CrossRef]
- Chen, S.; Ding, Y. Machine Learning and Its Applications in Studying the Geographical Distribution of Ants. Diversity 2022, 14, 706. [Google Scholar] [CrossRef]
- El Alaoui, O.; Idri, A. Predicting the Potential Distribution of Wheatear Birds Using Stacked Generalization-Based Ensembles. Ecol. Inform. 2023, 75, 102084. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, H.; Qu, W.; Luan, P.; Sun, J. Research on Geological Safety Evaluation Index Systems and Methods for Assessing Underground Space in Coastal Bedrock Cities Based on a Back-Propagation Neural Network Comprehensive Evaluation–Analytic Hierarchy Process (BPCE-AHP). Sustainability 2023, 15, 8055. [Google Scholar] [CrossRef]
- Azcárate, F.M.; Seoane, J.; Silvestre, M. Factors Affecting Pine Processionary Moth (Thaumetopoea pityocampa) Incidence in Mediterranean Pine Stands: A Multiscale Approach. For. Ecol. Manag. 2023, 529, 120728. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.G.; Dawson, A.; Zhai, L.; Stadt, K.J.; Comeau, P.G.; Whitehouse, C. Contributions of Insects and Droughts to Growth Decline of Trembling Aspen Mixed Boreal Forest of Western Canada. Glob. Change Biol. 2018, 24, 655–667. [Google Scholar] [CrossRef]
- Arabameri, A.; Nalivan, O.A.; Saha, S.; Roy, J.; Pradhan, B.; Tiefenbacher, J.P.; Ngo, P.T.T. Novel Ensemble Approaches of Machine Learning Techniques in Modeling the Gully Erosion Susceptibility. Remote Sens. 2020, 12, 1890. [Google Scholar] [CrossRef]
- Lu, S.; Ye, S.-J. Using an Image Segmentation and Support Vector Machine Method for Identifying Two Locust Species and Instars. J. Integr. Agric. 2020, 19, 1301–1313. [Google Scholar] [CrossRef]
- Zhang, C.; Park, D.S.; Yoon, S.; Zhang, S. Editorial: Machine Learning and Artificial Intelligence for Smart Agriculture. Front. Plant Sci. 2023, 13, 1121468. [Google Scholar] [CrossRef]
- Rafiq, D.; Bazaz, M.A. A Collection of Large-Scale Benchmark Models for Nonlinear Model Order Reduction. Arch. Comput. Methods Eng. 2023, 30, 69–83. [Google Scholar] [CrossRef]
- Wang, L.; Xu, M. Regression-Based Identification and Order Reduction Method for Nonlinear Dynamic Structural Models. Proc. Inst. Mech. Eng. Part G J. Aerosp. Eng. 2023, 237, 3508–3523. [Google Scholar] [CrossRef]
- Zhu, R.; Fei, Q.; Jiang, D.; Marchesiello, S.; Anastasio, D. Bayesian Model Selection in Nonlinear Subspace Identification. AIAA J. 2022, 60, 92–101. [Google Scholar] [CrossRef]
- Monter-Pozos, A.; González-Estrada, E. On Testing the Skew Normal Distribution by Using Shapiro–Wilk Test. J. Comput. Appl. Math. 2024, 440, 115649. [Google Scholar] [CrossRef]
- El Habib Daho, M.; Amine Chikh, M. Combining Bootstrapping Samples, Random Subspaces and Random Forests to Build Classifiers. J. Med. Imaging Health Inform. 2015, 5, 539–544. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, H.; Zhou, Q.; Cui, A. A XGBoost-Based Downscaling-Calibration Scheme for Extreme Precipitation Events. IEEE Trans. Geosci. Remote Sens. 2023, 61, 4103512. [Google Scholar] [CrossRef]
- Lyu, J.; Zheng, P.; Qi, Y.; Huang, G. LightGBM-LncLoc: A LightGBM-Based Computational Predictor for Recognizing Long Non-Coding RNA Subcellular Localization. Mathematics 2023, 11, 602. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, T.; Du, Q.; Quyang, N.; Nie, W.; Liu, Y.; Gou, P.; Li, G. Spatiotemporal changes of ecosystem health and the impact of its driving factors on the Loess Plateau in China. J. Ecol. Indic. 2025, 170, 1677–1680. [Google Scholar] [CrossRef]
- Pham, B.T.; Luu, C.; Van Phong, T.; Nguyen, H.D.; Van Le, H.; Tran, T.Q.; Ta, H.T.; Prakash, I. Flood Risk Assessment Using Hybrid Artificial Intelligence Models Integrated with Multi-Criteria Decision Analysis in Quang Nam Province, Vietnam. J. Hydrol. 2021, 592, 125815. [Google Scholar] [CrossRef]
- Liu, C.; Li, W.; Wang, W.; Zhou, H.; Liang, T.; Hou, F.; Xu, J.; Xue, P. Quantitative Spatial Analysis of Vegetation Dynamics and Potential Driving Factors in a Typical Alpine Region on the Northeastern Tibetan Plateau Using the Google Earth Engine. Catena 2021, 206, 105500. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial Insects along Elevation Gradients: Species and Community Responses to Altitude. Biol. Rev. Camb. Philos. Soc. 2005, 80, 489–513. [Google Scholar] [CrossRef]
- Marini, L.; Fontana, P.; Klimek, S.; Battisti, A.; Gaston, K.J. Impact of Farm Size and Topography on Plant and Insect Diversity of Managed Grasslands in the Alps. Biol. Conserv. 2009, 142, 394–403. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T. Potential Effects of Climate Change on Ecosystem and Tree Species Distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Hernández-Alonso, R.; Camarero, J.J. Patterns and Drivers of Pine Processionary Moth Defoliation in Mediterranean Mountain Forests. Front. Ecol. Evol. 2019, 7, 458. [Google Scholar] [CrossRef]
- Marini, L.; Ayres, M.P.; Battisti, A.; Faccoli, M. Climate Affects Severity and Altitudinal Distribution of Outbreaks in an Eruptive Bark Beetle. Clim. Chang. 2012, 115, 327–341. [Google Scholar] [CrossRef]
- Haynes, K.J.; Liebhold, A.M.; Lefcheck, J.S.; Morin, R.S.; Wang, G. Climate Affects the Outbreaks of a Forest Defoliator Indirectly through Its Tree Hosts. Oecologia 2022, 198, 407–418. [Google Scholar] [CrossRef]
- Lai, H.; Hales, S.; Woodward, A.; Walker, C.; Marks, E.; Pillai, A.; Chen, R.X.; Morton, S.M. Effects of Heavy Rainfall on Waterborne Disease Hospitalizations among Young Children in Wet and Dry Areas of New Zealand. Environ. Int. 2020, 145, 106136. [Google Scholar] [CrossRef]
- Aoki, C.F.; Cook, M.; Dunn, J.; Finley, D.; Fleming, L.; Yoo, R.; Ayres, M.P. Old Pests in New Places: Effects of Stand Structure and Forest Type on Susceptibility to a Bark Beetle on the Edge of Its Native Range. For. Ecol. Manag. 2018, 419–420, 206–219. [Google Scholar] [CrossRef]
- Bognounou, F.; De Grandprè, L.; Pureswaran, D.S.; Kneeshaw, D. Temporal Variation in Plant Neighborhood Effects on the Defoliation of Primary and Secondary Hosts by an Insect Pest. Ecosphere 2017, 8, e01759. [Google Scholar] [CrossRef]
- Dodds, K.J.; Aoki, C.F.; Arango-Velez, A.; Cancelliere, J.; D’Amato, A.W.; DiGirolomo, M.F.; Rabaglia, R.J. Expansion of Southern Pine Beetle into Northeastern Forests: Management and Impact of a Primary Bark Beetle in a New Region. J. For. 2018, 116, 178–191. [Google Scholar] [CrossRef]
- Sánchez-Cuesta, R.; Ruiz-Gómez, F.J.; Duque-Lazo, J.; González-Moreno, P.; Navarro-Cerrillo, R.M. The Environmental Drivers Influencing Spatio-Temporal Dynamics of Oak Defoliation and Mortality in Dehesas of Southern Spain. For. Ecol. Manag. 2021, 485, 118946. [Google Scholar] [CrossRef]
- Walter, J.A.; Platt, R.V. Multi-Temporal Analysis Reveals That Predictors of Mountain Pine Beetle Infestation Change during Outbreak Cycles. For. Ecol. Manag. 2013, 302, 308–318. [Google Scholar] [CrossRef]
- Figueredo, L.; Villa-Murillo, A.; Colmenarez, Y.; Vásquez, C. A Hybrid Artificial Intelligence Model for Aeneolamia varia (Hemiptera: Cercopidae) Populations in Sugarcane Crops. J. Insect Sci. 2021, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- DeRose, R.J.; Bentz, B.J.; Long, J.N.; Shaw, J.D. Effect of Increasing Temperatures on the Distribution of Spruce Beetle in Engelmann Spruce Forests of the Interior West, USA. For. Ecol. Manag. 2013, 308, 198–206. [Google Scholar] [CrossRef]
- Lalande, B.M.; Hughes, K.; Jacobi, W.R.; Tinkham, W.T.; Reich, R.; Stewart, J.E. Subalpine Fir Mortality in Colorado Is Associated with Stand Density, Warming Climates and Interactions among Fungal Diseases and the Western Balsam Bark Beetle. For. Ecol. Manag. 2020, 466, 118133. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Farooq, M.; Al-Sadi, A.M.; Nawaz, A.; Jabran, K.; Siddique, K.H.M. Impact of Climate Change on Biology and Management of Wheat Pests. Crop Prot. 2020, 137, 105304. [Google Scholar] [CrossRef]
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a Warming Climate: Insect Responses to Extreme High Temperatures. Annu. Rev. Entomol. 2021, 66, 163–184. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Factor Description | Abbreviation |
|---|---|---|
| Climate | 5-year average surface net solar radiation | ssr (J/m2) |
| 5-year average 2 m temperature | t2m (K) | |
| 5-year average 2 m dewpoint temperature | d2m (K) | |
| 5-year average 10 m u-component of wind | u10 (m/s) | |
| 5-year average 10 m v-component of wind | v10 (m/s) | |
| 5-year average evaporation | e (m of weq) | |
| 5-year average total precipitation | tp (m) | |
| average total precipitation in April | tp_04 (m) | |
| average 2 m temperature in April | t2m_04 (K) | |
| average surface runoff in April | sro_04 (m) | |
| Vegetation | 5-year average high vegetation coverage | cvh (0–1) |
| 5-year average leaf area index of high vegetations | lai_hv (m2/m2) | |
| 5-year average high vegetation types | tvh | |
| Soil | 5-year average soil moisture at 0–7 cm depth | swvl1 (m3/m3) |
| 5-year average temperature at 0–7 cm depth | stl1 (K) | |
| monthly mean soil moisture in April of the year of occurrence occurrence | swvl1_04 (m3/m3) | |
| monthly mean temperature in April of the year of occurrence | stl1_04 (K) | |
| Snow | 5-year average temperature of snow layer | tsn (K) |
| 5-year average snow depth | sd (m of weq) | |
| average snow depth in April | sd_04 (m of weq) | |
| Topography | digital elevation model | DEM (m) |
| slope | slope | |
| Human | 5-year average population density | popu_densi |
| Methods | Class | Precision | Recall | F1-Score | Val_Accuracy |
|---|---|---|---|---|---|
| MRF | No Occur | 0.97 | 0.99 | 0.98 | 0.9748 |
| Occur | 0.97 | 0.90 | 0.95 | ||
| RF | No Occur | 0.95 | 0.99 | 0.97 | 0.9505 |
| Occur | 0.96 | 0.86 | 0.91 | ||
| XGBoost | No Occur | 0.93 | 0.98 | 0.96 | 0.9385 |
| Occur | 0.95 | 0.83 | 0.89 | ||
| LGBM | No Occur | 0.94 | 0.98 | 0.96 | 0.9417 |
| Occur | 0.95 | 0.85 | 0.90 |
| Lowest | Lower | Medium | Higher | Highest | |
|---|---|---|---|---|---|
| Probability value | 0~0.25 | 0.25~0.50 | 0.50~0.70 | 0.70~0.85 | 0.85~1.00 |
| 2019 | 2020 | 2021 | 2022 | 2023 | 2024 | |
|---|---|---|---|---|---|---|
| d2m | 0.9865 | 0.9211 | 0.9437 | 0.9193 | 0.9437 | 0.9553 |
| e | 0.9642 | 0.8111 | 0.8355 | 0.8089 | 0.8355 | 0.9513 |
| cvh | 0.9739 | 0.9463 | 0.9598 | 0.9322 | 0.9598 | 0.9383 |
| lai_hv | 0.9846 | 0.9529 | 0.9614 | 0.9393 | 0.9614 | 0.9562 |
| sd | 0.7452 | 0.1751 | 0.3443 | 0.2967 | 0.3443 | 0.9615 |
| ssr | 0.9815 | 0.9279 | 0.9223 | 0.8983 | 0.9223 | 0.9595 |
| stl1 | 0.9860 | 0.9473 | 0.9509 | 0.9277 | 0.9509 | 0.9524 |
| swvl1 | 0.9868 | 0.9616 | 0.9565 | 0.9392 | 0.9565 | 0.9602 |
| tvh | 0.2477 | 0.1472 | 0.2414 | 0.2893 | 0.2414 | 0.2049 |
| t2m | 0.9870 | 0.9454 | 0.9526 | 0.9263 | 0.9526 | 0.9560 |
| tp | 0.9249 | 0.7601 | 0.7484 | 0.7441 | 0.7484 | 0.9524 |
| tsn | 0.9855 | 0.9312 | 0.9395 | 0.9138 | 0.9395 | 0.9477 |
| u10 | 0.9818 | 0.8495 | 0.8671 | 0.8475 | 0.8671 | 0.9616 |
| v10 | 0.9863 | 0.8983 | 0.9196 | 0.9119 | 0.9196 | 0.9616 |
| sro_04 | 0.0901 | 0.0779 | 0.1760 | 0.2003 | 0.1760 | 0.3464 |
| sd_04 | 0.1627 | 0.1819 | 0.1979 | 0.2719 | 0.1979 | 0.8530 |
| tp_04 | 0.7016 | 0.7572 | 0.8022 | 0.7637 | 0.8022 | 0.2371 |
| t2m_04 | 0.9288 | 0.9275 | 0.9437 | 0.9272 | 0.9437 | 0.9567 |
| swvl1_04 | 0.9671 | 0.9577 | 0.9692 | 0.9474 | 0.9692 | 0.9611 |
| stl1_04 | 0.9407 | 0.9411 | 0.9491 | 0.9284 | 0.9491 | 0.9572 |
| popu_densi | 0.1389 | 0.2135 | 0.2626 | 0.2088 | 0.2626 | 0.1719 |
| DEM | 0.2193 | 0.2018 | 0.2398 | 0.2555 | 0.2398 | 0.3126 |
| slope | 0.1395 | 0.1291 | 0.1472 | 0.1853 | 0.1472 | 0.1746 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Wang, M.; Cai, D.; Wu, L.; Ji, X.; Ding, Q.; Wang, F.; Wang, M. Spatiotemporal Changes of Pine Caterpillar Infestation Risk and the Driving Effect of Habitat Factors in Northeast China. Remote Sens. 2025, 17, 1738. https://doi.org/10.3390/rs17101738
Zhao J, Wang M, Cai D, Wu L, Ji X, Ding Q, Wang F, Wang M. Spatiotemporal Changes of Pine Caterpillar Infestation Risk and the Driving Effect of Habitat Factors in Northeast China. Remote Sensing. 2025; 17(10):1738. https://doi.org/10.3390/rs17101738
Chicago/Turabian StyleZhao, Jingzheng, Mingchang Wang, Dong Cai, Linlin Wu, Xue Ji, Qing Ding, Fengyan Wang, and Minshui Wang. 2025. "Spatiotemporal Changes of Pine Caterpillar Infestation Risk and the Driving Effect of Habitat Factors in Northeast China" Remote Sensing 17, no. 10: 1738. https://doi.org/10.3390/rs17101738
APA StyleZhao, J., Wang, M., Cai, D., Wu, L., Ji, X., Ding, Q., Wang, F., & Wang, M. (2025). Spatiotemporal Changes of Pine Caterpillar Infestation Risk and the Driving Effect of Habitat Factors in Northeast China. Remote Sensing, 17(10), 1738. https://doi.org/10.3390/rs17101738