
Forest Community Spatial Modeling Using Machine Learning and Remote Sensing Data




Abstract

1. Introduction
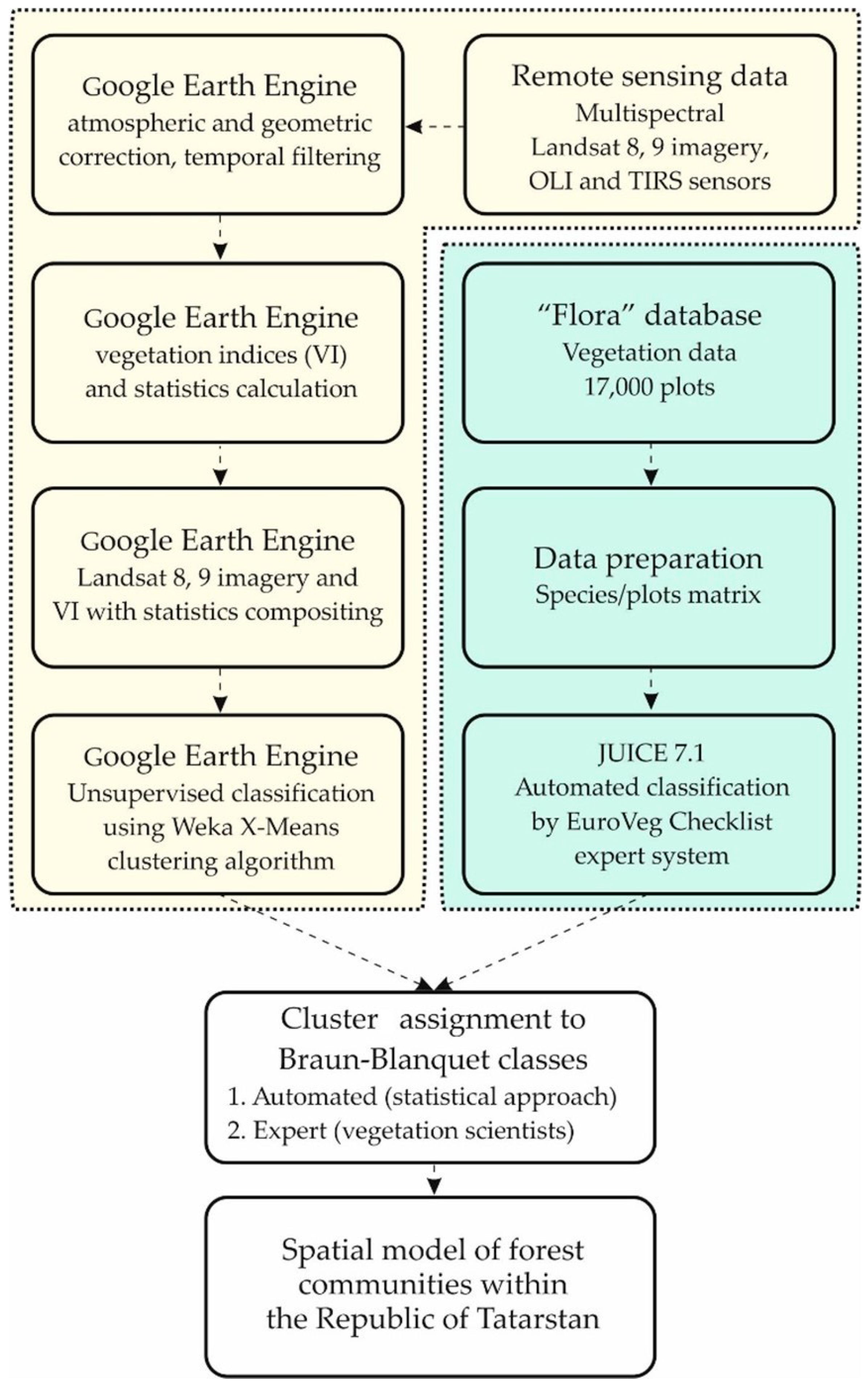
2. Materials and Methods
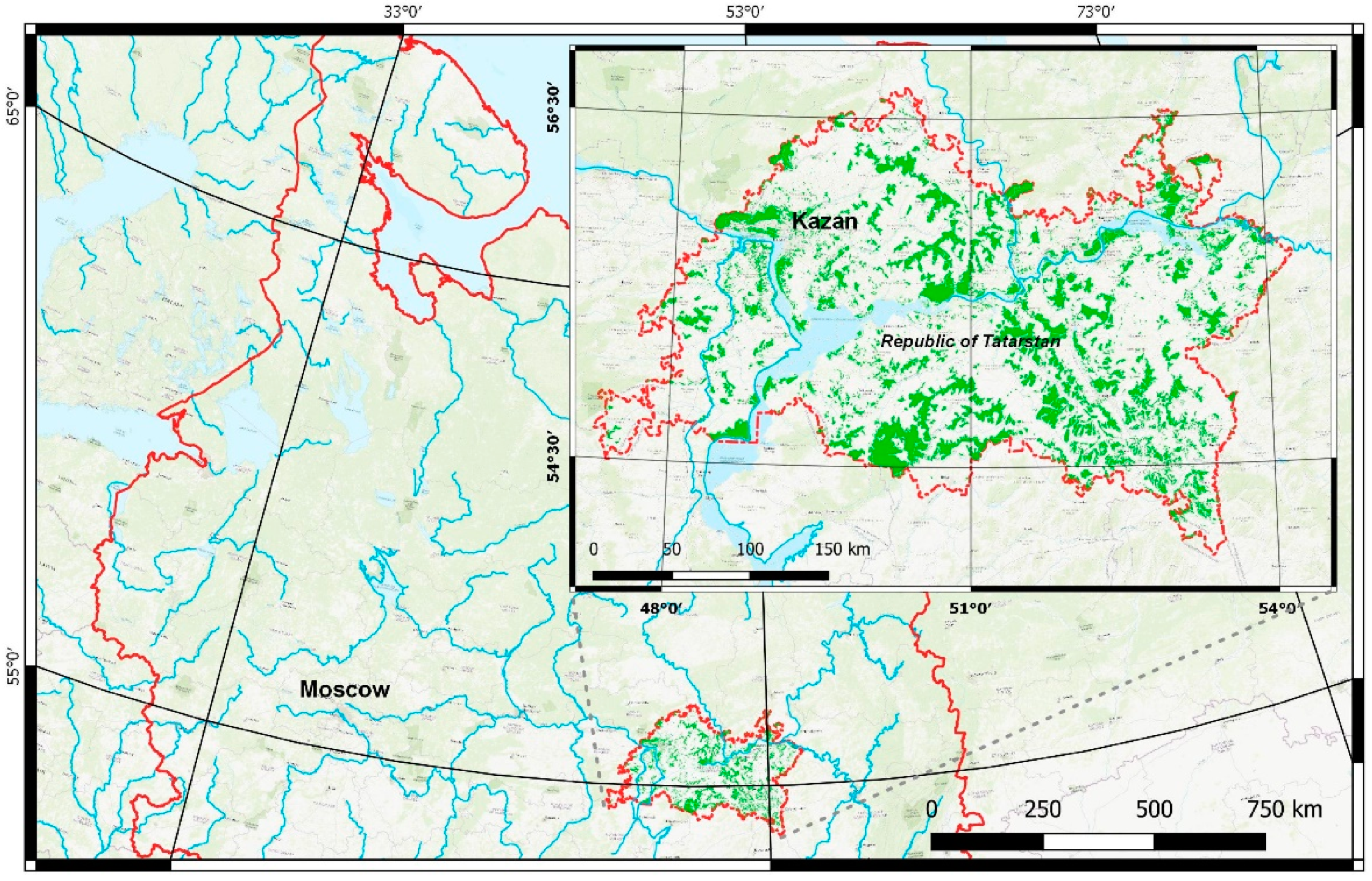
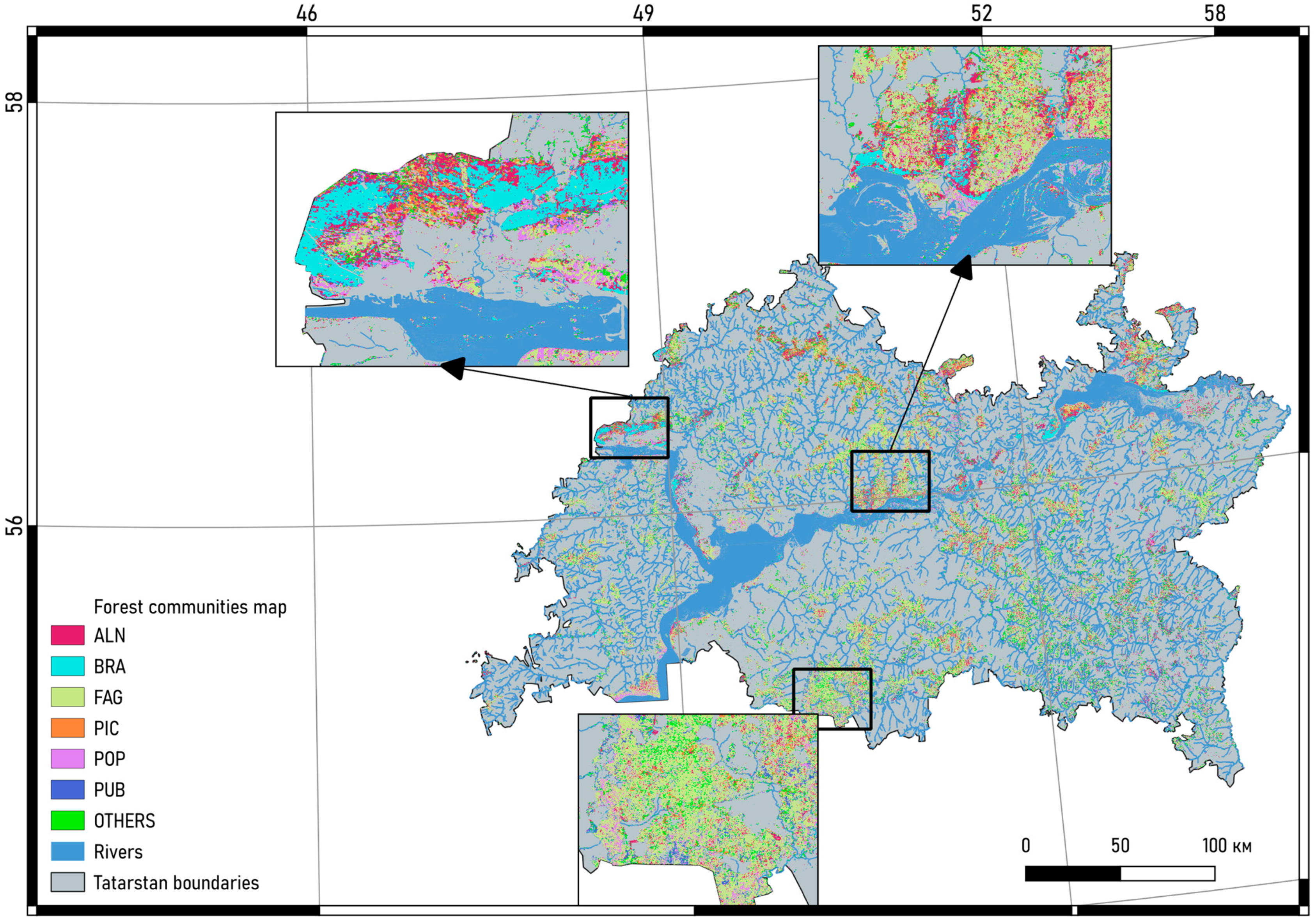
2.1. Study Area
2.2. Field Data Collection and Processing
2.3. Source Data Preparation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index Name | Description |
|---|---|
| Reflectance-based indices | |
| DVI (Difference Vegetation Index) [34] | Difference in reflectance between the near-infrared (NIR) and red bands. |
| DVIplus [35] | An extension of DVI, considering additional spectral bands or factors. |
| GNDVI (Green Normalized Difference Vegetation Index) [36] | Utilizes the green and NIR bands to emphasize chlorophyll content. |
| GRNDVI (Green-Red Normalized Difference Vegetation Index) [37] | Incorporates green and red bands for enhanced vegetation monitoring. |
| MNDVI (Modified NDVI) [38] | An adjusted form of NDVI to account for atmospheric and canopy background effects. |
| NDVI (Normalized Difference Vegetation Index) [39] | One of the most widely used indices, highlighting areas of active vegetation. |
| RDVI (Renormalized Difference Vegetation Index) [34] | Aims to minimize soil background influence while emphasizing vegetation. |
| TDVI (Transformed Difference Vegetation Index) [40] | A modified version of NDVI to enhance sensitivity to vegetation changes. |
| WDRVI (Wide Dynamic Range Vegetation Index) [41] | Offers a wider dynamic range than that of NDVI, improving sensitivity. |
| WDVI (Weighted Difference Vegetation Index) [42] | Focuses on minimizing soil background effects by applying a specific weight. |
| Water content indices | |
| DSWI1 (Drought Stress Water Index 1) [43] | Highlights areas undergoing drought stress, indicating lower water content. |
| GSAVI (Green Soil Adjusted Vegetation Index) [44] | A soil-adjusted index using the green band to minimize soil background influences. |
| NIRv (Near-Infrared Reflectance of Vegetation) [45] | Captures the NIR reflectance associated with vegetation’s structural characteristics. |
| NIRvH2 [46] | A variation of NIRv, considering additional factors for enhanced accuracy. |
| NDII (Normalized Difference Infrared Index) [47] | Helps in assessing vegetation water content. |
| NDMI (Normalized Difference Moisture Index) [48] | Another index for evaluating water content in vegetation. |
| NMDI (Normalized Multi-band Drought Index) [49] | Focuses on monitoring drought conditions across various bands. |
| MSI (Moisture Stress Index) [50] | Highlights moisture stress in vegetation, crucial for drought monitoring. |
| Soil-adjusted indices | |
| GDVI (Green Difference Vegetation Index) [51] | Minimizes soil background influences using the green band. |
| GOSAVI (Green Optimized Soil Adjusted Vegetation Index) [44] | A soil-adjusted index designed for optimal vegetation monitoring. |
| GRVI (Green Red Vegetation Index) [44] | Utilizes green and red bands for vegetation analysis, adjusting for soil effects. |
| MSAVI (Modified Soil Adjusted Vegetation Index) [52] | A soil-adjusted index with modifications for better accuracy. |
| OSAVI (Optimized Soil Adjusted Vegetation Index) [53] | Optimizes soil adjustment to improve vegetation monitoring in low cover areas. |
| SARVI (Soil Adjusted and Atmospherically Resistant Vegetation Index) [54] | A soil-adjusted and atmospherically resistant index. |
| SAVI (Soil Adjusted Vegetation Index) [55] | Adjusts for the influence of soil bcenterness when vegetation cover is low. |
| TSAVI (Transformed Soil Adjusted Vegetation Index) [56] | A transformed index for better vegetation representation in diverse conditions. |
| Chlorophyll indices | |
| GARI (Green Atmospherically Resistant Vegetation Index) [36] | Designed to monitor chlorophyll content while minimizing atmospheric effects. |
| GCC (Green Chlorophyll Content) [57] | Directly related to chlorophyll content, crucial for assessing vegetation health. |
| MGRVI (Modified Green Red Vegetation Index) [58] | A modified index to enhance sensitivity to chlorophyll content. |
| MCARI2 (Modified Chlorophyll Absorption in Reflectance Index 2) [59] | Targets chlorophyll absorption features for accurate monitoring. |
| MSR (Modified Simple Ratio) [60] | A modified vegetation index to improve sensitivity to chlorophyll content. |
| MTVI2 (Modified Triangular Vegetation Index 2) [59] | Focuses on enhancing the representation of chlorophyll content. |
| OCVI (Optimized Chlorophyll Vegetation Index) [61] | Optimizes chlorophyll representation in vegetation monitoring. |
| VIG (Vegetation Index Green) [62] | Utilizes the green band for chlorophyll monitoring, essential for assessing plant health. |
| Structural indices | |
| CVI (Chlorophyll Vegetation Index) [63] | Represents vegetation structure and chlorophyll content. |
| DSI (Difference Structure Index) [64] | Highlights structural variations in vegetation. |
| SR (Simple Ratio) [65] | A basic ratio of NIR-to-red reflectance, indicating vegetation structure. |
| Non-linear indices | |
| EVI2 (Enhanced Vegetation Index 2) [66] | An improved version of NDVI, incorporating non-linear enhancements. |
| GBNDVI (Green Blue Normalized Difference Vegetation Index) [37] | A non-linear index utilizing green and blue bands. |
| GLI (Green Leaf Index) [67] | Represents vegetation greenness in a non-linear manner. |
| GEMI (Global Environment Monitoring Index) [68] | A global index for vegetation monitoring with non-linear properties. |
| RI (RapidEye Vegetation Index) [69] | A specific index for RapidEye satellite data. |
| SI (Shadow Index) [70] | Represents vegetation shape characteristics. |
| SEVI (Soil and Atmospherically Resistant Vegetation Index) [71] | Minimizes soil and atmospheric effects using a non-linear approach. |
| VARI (Visible Atmospherically Resistant Index) [62] | Focuses on the visible spectrum for vegetation monitoring, applying non-linear corrections. |
| Other indices | |
| BWDRVI (Broadband Width Difference Vegetation Index) [72] | A unique index capturing broadband width differences. |
| CIG (Canopy Index Green) [73] | Represents canopy structure using the green band. |
| FCVI (Floating Canopy Vegetation Index) [74] | A special index for floating canopy vegetation. |
| IAVI (Inverted Attributed Vegetation Index) [75] | An inverted index for enhanced vegetation attribute representation. |
| IKAW (Kawashima Vegetation Index) [76] | A specific vegetation index developed by Kawashima. |
| IPVI (Infrared Percentage Vegetation Index) [77] | Utilizes infrared reflectance for vegetation percentage estimation. |
| MRBVI (Modified Ratio Vegetation Index) [78] | A modified ratio index for improved vegetation monitoring. |
| MNLI (Modified Non-Linear Vegetation Index) [79] | Incorporates non-linear adjustments for enhanced vegetation representation. |
| NDDI (Normalized Difference Drought Index) [80] | Focuses on drought monitoring and water stress assessment. |
| NDGI (Normalized Difference Greenness Index) [35] | Highlights vegetation greenness. |
| NDPI (Normalized Difference Phenology Index) [81] | Utilized for monitoring vegetation phenology. |
| NDYI (Normalized Difference Yellowness Index) [82] | Highlights vegetation yellowness, crucial for certain phenological stages. |
| NGRDI (Normalized Green Red Difference Index) [83] | Utilizes green and red bands for vegetation monitoring. |
| NRFIg (Normalized Red/Far-Red Index Green) [84] | Focuses on the red-to-far-red ratio using the green band. |
| NRFIr (Normalized Red/Far-Red Index Red) [84] | Utilizes the red-to-far-red ratio in the red band. |
| NormG (Normalized Green) [44] | Represents normalized green reflectance. |
| NormNIR (Normalized NIR) [44] | Represents normalized near-infrared reflectance. |
| NormR (Normalized Red) [44] | Represents normalized red reflectance. |
| RCC (Red Chromatic Coordinate) [57] | Directly related to chlorophyll content using the red band. |
| RGBVI (Red Green Blue Vegetation Index) [58] | Utilizes the RGB bands for vegetation analysis. |
| RGRI (Red Green Ratio Index) [85] | A ratio index using red and green bands. |
| TGI (Triangular Greenness Index) [86] | Represents vegetation greenness using a triangular approach. |
| TriVI (Triangular Vegetation Index) [87] | A triangular index for enhanced vegetation representation. |
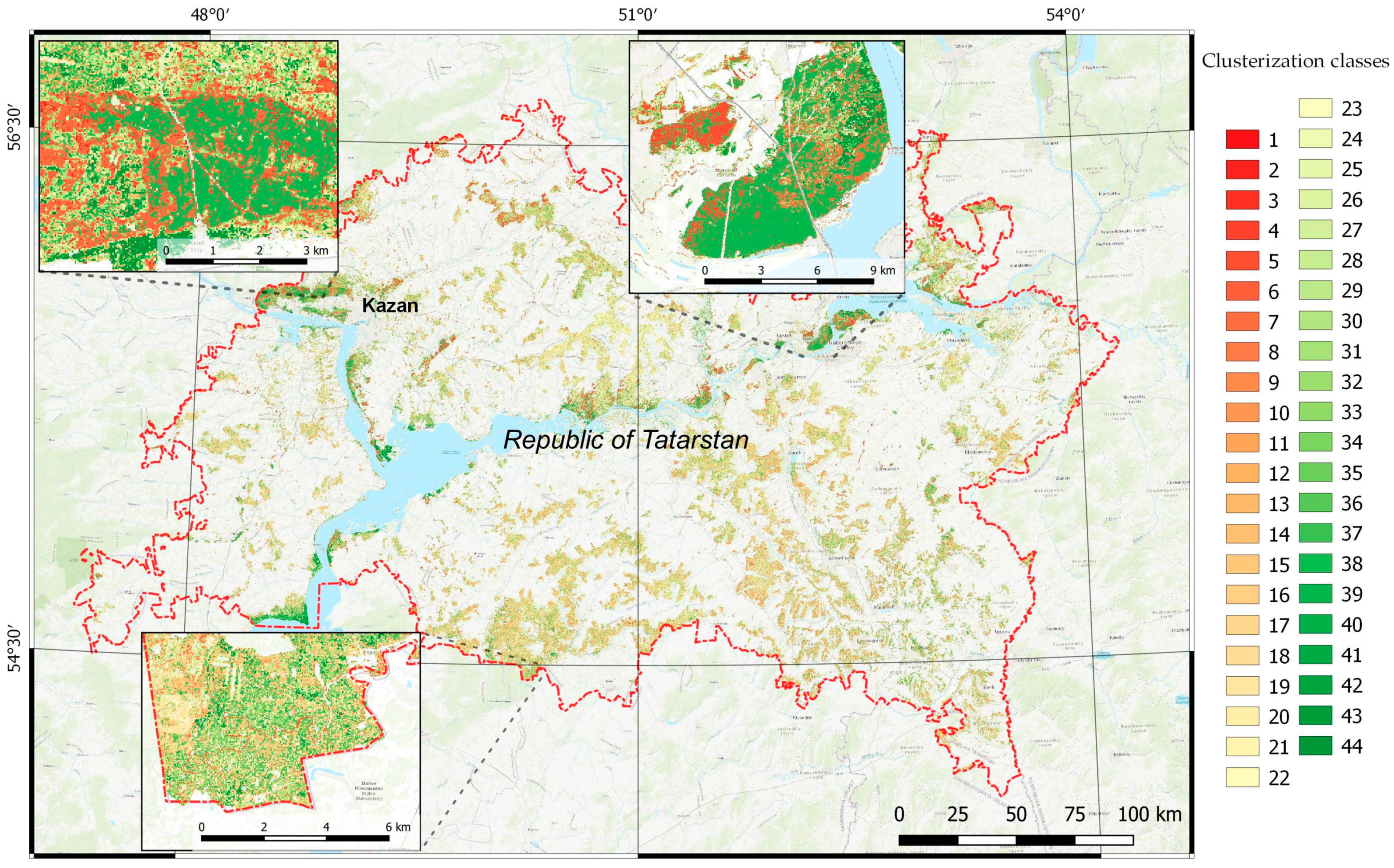
2.4. Remote-Sensing-Data-Based Clusterization
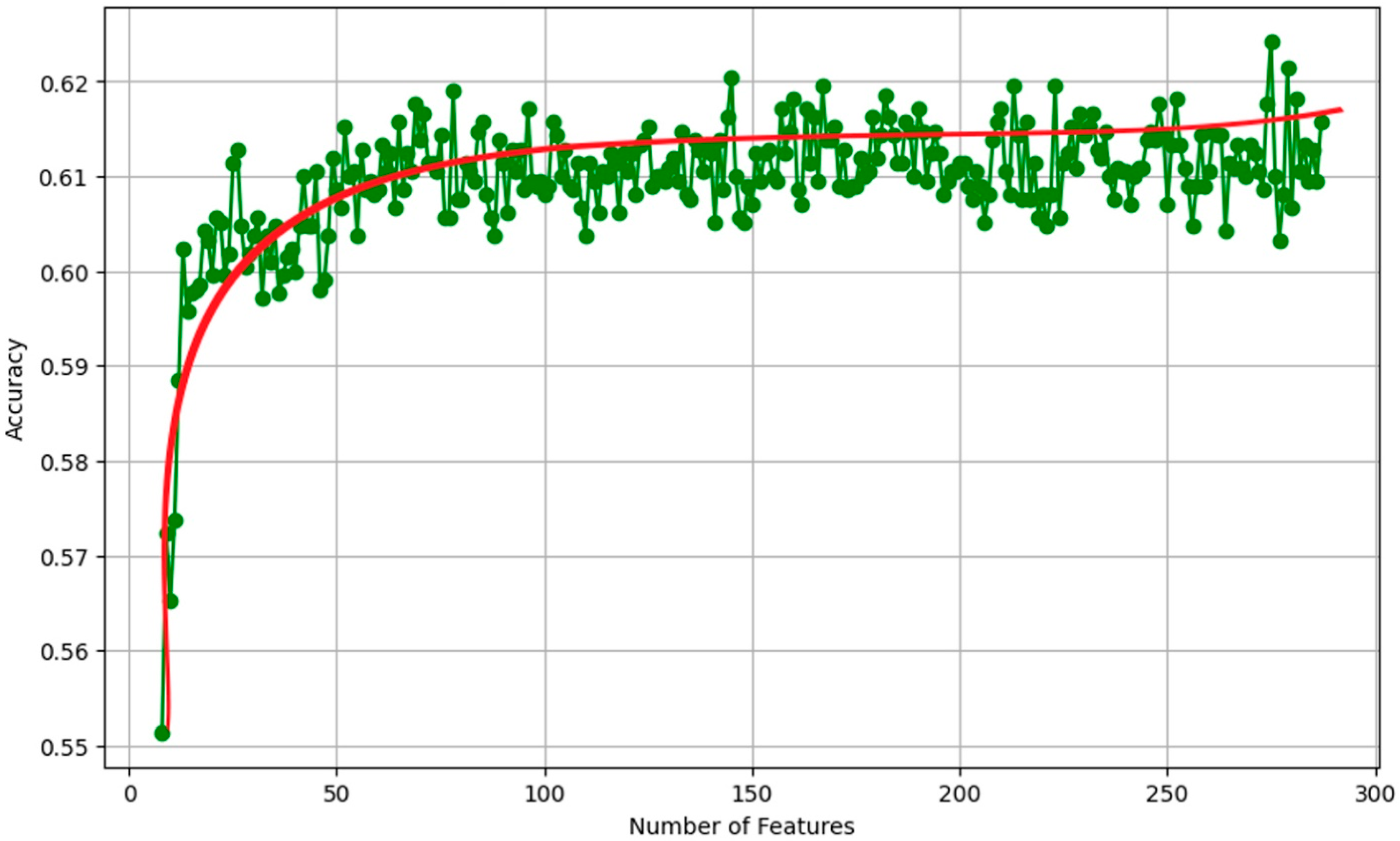
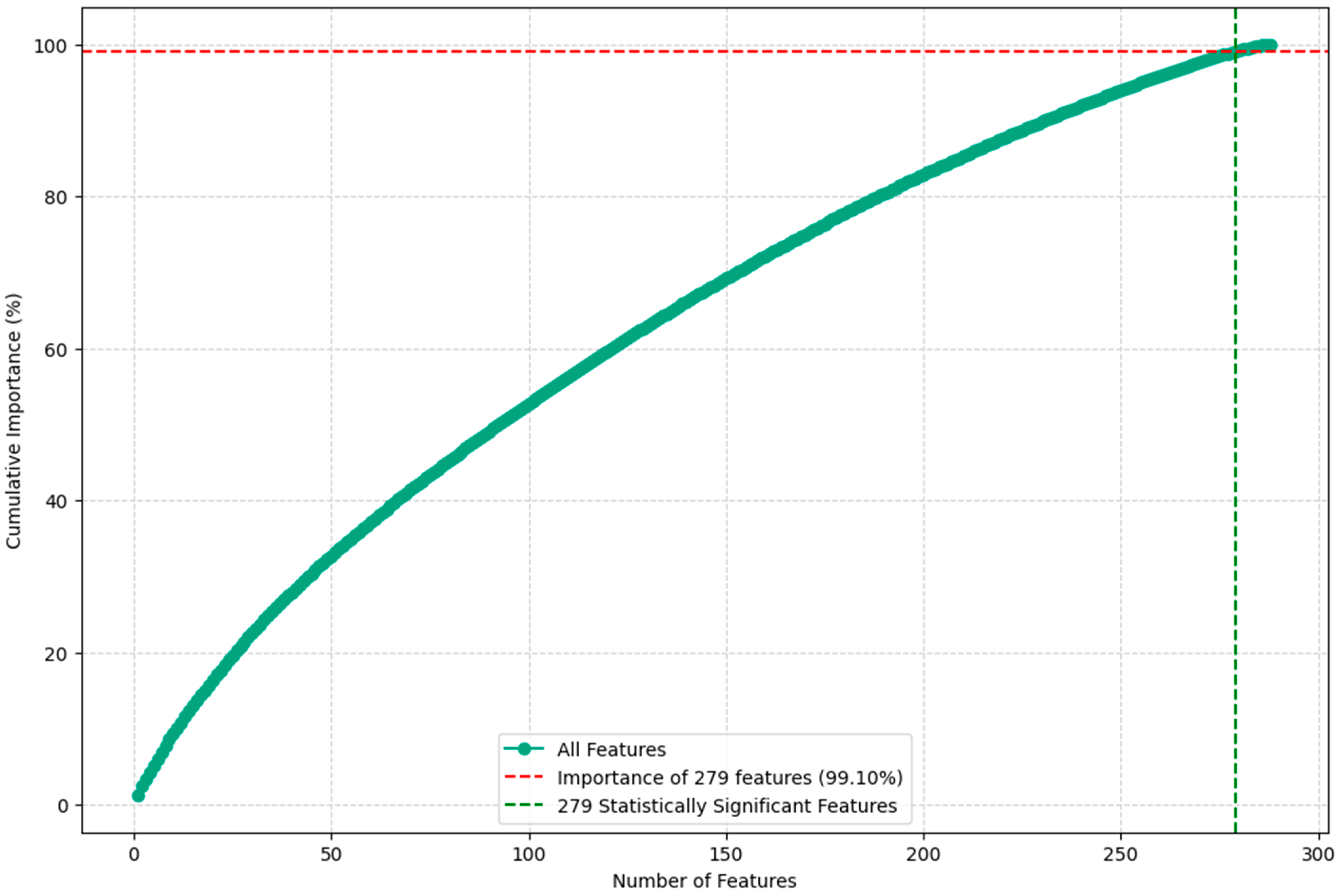
2.5. Validation and Analysis
- Data Preparation: The input data were partitioned into a training and testing set with a stratified sampling approach to ensure proportional representation of each vegetation class in both subsets.
- Model Training: The Random Forest algorithm was implemented using the scikit-learn library in Python. Hyperparameters such as the number of trees, maximum depth, and minimum samples per leaf were optimized through a grid search and cross-validation process.
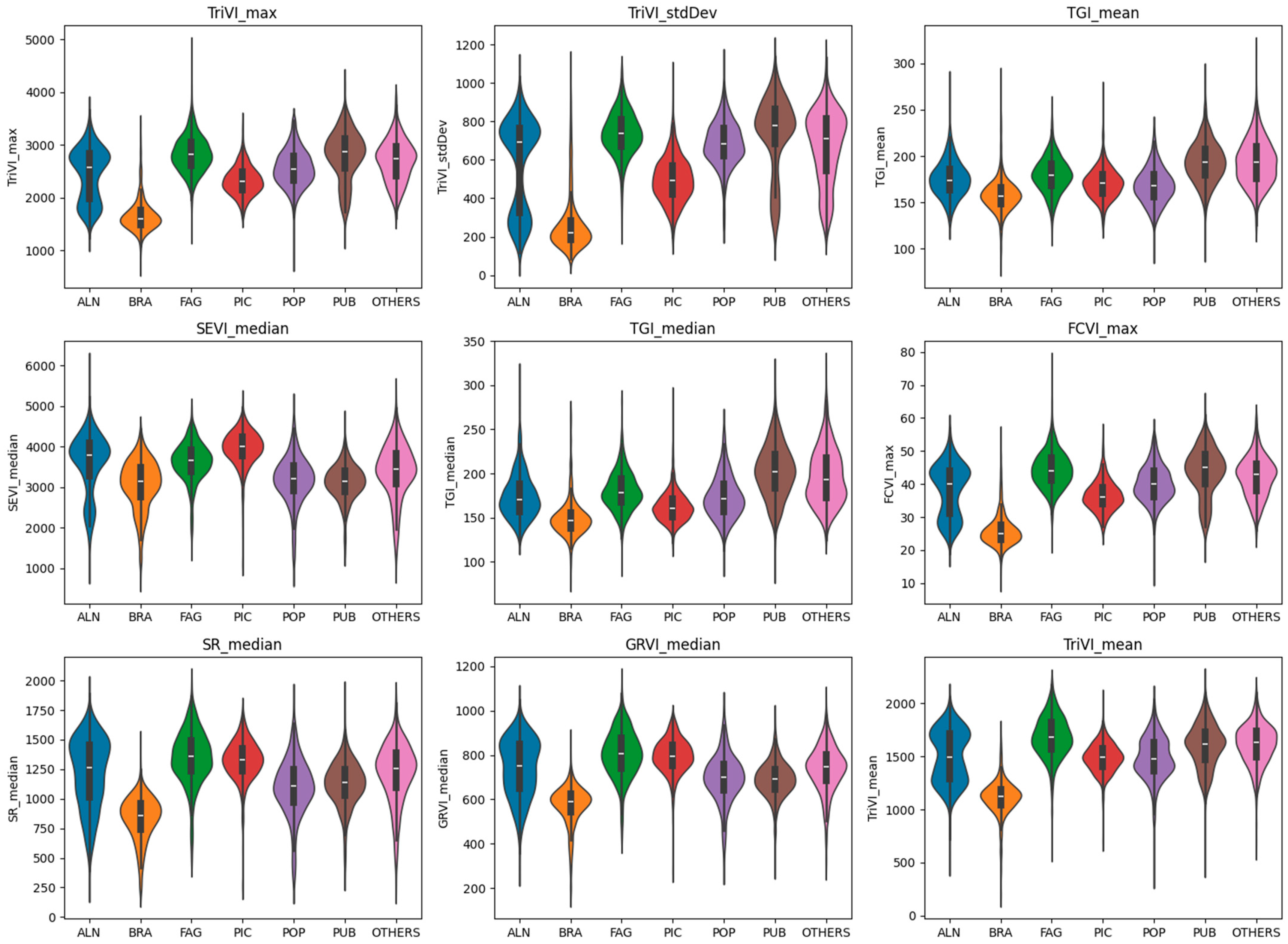
- Feature Importance Evaluation: The trained Random Forest model provides a measure of feature importance, which quantifies the contribution of each predictor variable to the model’s overall predictive performance. This information was used to assess the relative importance of the vegetation indices and their statistical metrics in distinguishing between the different vegetation classes.
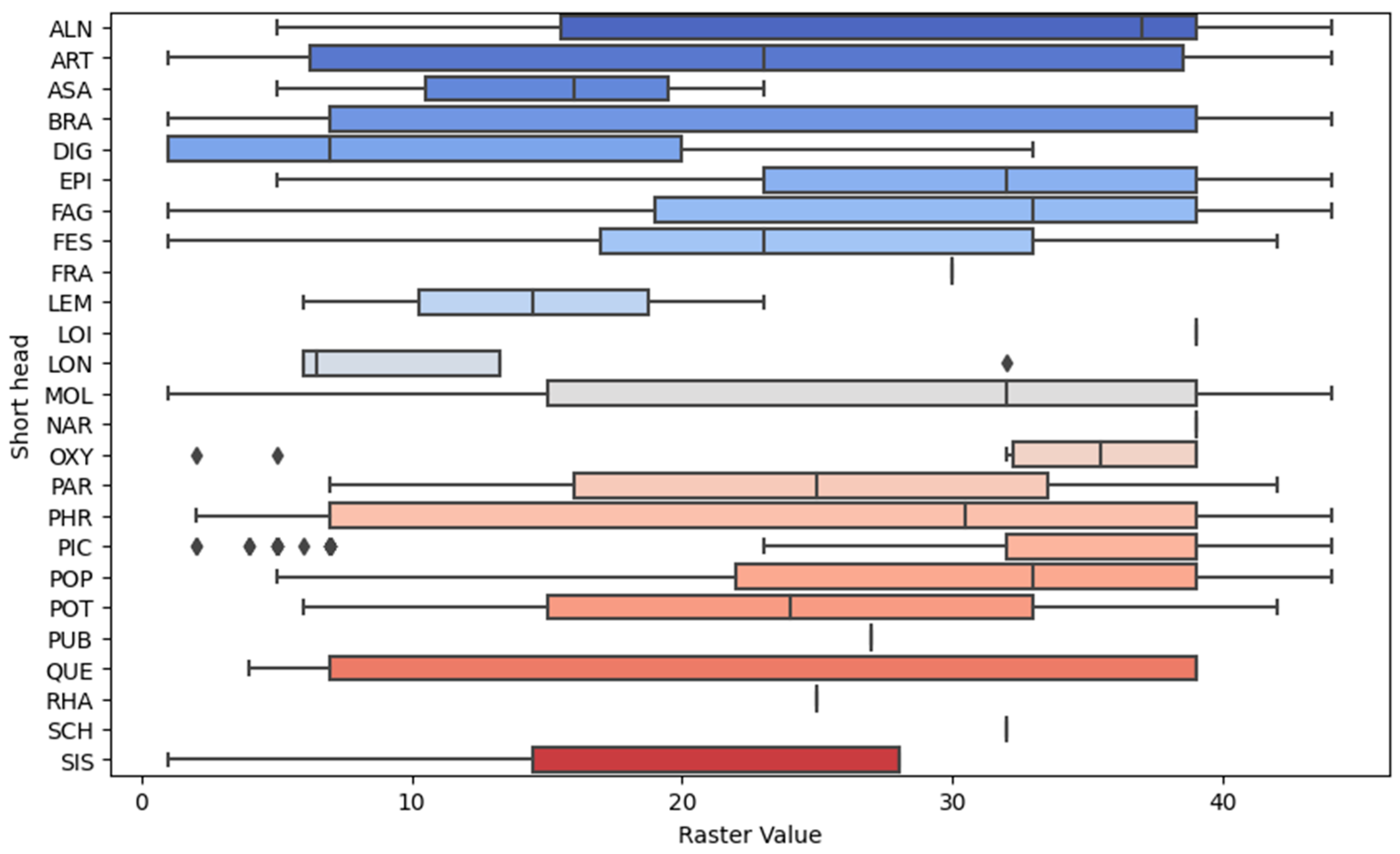
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Executive Steering Committee for Australian Vegetation Information. Australian Vegetation Attribute Manual: National Vegetation Information System, Version 6.0; Department of Environment and Heritage: Canberra, Australia, 2003.
- Faber-Langendoen, D.; Keeler-Wolf, T.; Meidinger, D.; Tart, D.; Hoagland, B.; Josse, C.; Navarro, G.; Ponomarenko, S.; Saucier, J.-P.; Weakley, A.; et al. EcoVeg: A New Approach to Vegetation Description and Classification. Ecol. Monogr. 2014, 84, 533–561. [Google Scholar] [CrossRef]
- De Cáceres, M.; Chytrý, M.; Agrillo, E.; Attorre, F.; Botta-Dukát, Z.; Capelo, J.; Czúcz, B.; Dengler, J.; Ewald, J.; Faber-Langendoen, D.; et al. A Comparative Framework for Broad-scale Plot-based Vegetation Classification. Appl. Veg. Sci. 2015, 18, 543–560. [Google Scholar] [CrossRef]
- Kozhevnikova, M.; Prokhorov, V. Syntaxonomy of the Xero-Mesophytic Oak Forests in the Republic of Tatarstan (Eastern Europe). VCS 2021, 2, 47–58. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical Floristic Classification System of Vascular Plant, Bryophyte, Lichen, and Algal Communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M. Remote Sensing Imagery in Vegetation Mapping: A Review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Tierney, D.A. An Assessment of Vegetation Mapping Scale for Reserve Management: Does Scale of Assessment Dominate Assessment Outcomes? Biodivers. Conserv. 2023, 32, 2731–2745. [Google Scholar] [CrossRef]
- Bunting, P.; Lucas, R.M.; Jones, K.; Bean, A.R. Characterisation and Mapping of Forest Communities by Clustering Individual Tree Crowns. Remote Sens. Environ. 2010, 114, 2536–2547. [Google Scholar] [CrossRef]
- Ganz, S.; Adler, P.; Kändler, G. Forest Cover Mapping Based on a Combination of Aerial Images and Sentinel-2 Satellite Data Compared to National Forest Inventory Data. Forests 2020, 11, 1322. [Google Scholar] [CrossRef]
- De Cáceres, M.; Font, X.; Oliva, F. The Management of Vegetation Classifications with Fuzzy Clustering: Fuzzy Clustering in Vegetation Classifications. J. Veg. Sci. 2010, 21, 1138–1151. [Google Scholar] [CrossRef]
- Gafurov, A. The Methodological Aspects of Constructing a High-Resolution DEM of Large Territories Using Low-Cost UAVs on the Example of the Sarycum Aeolian Complex, Dagestan, Russia. Drones 2021, 5, 7. [Google Scholar] [CrossRef]
- Duda, T.; Canty, M. Unsupervised Classification of Satellite Imagery: Choosing a Good Algorithm. Int. J. Remote Sens. 2002, 23, 2193–2212. [Google Scholar] [CrossRef]
- Liu, X. Supervised Classification and Unsupervised Classification. ATS 670 Class Project, pp. 1–12. Available online: https://lweb.cfa.harvard.edu/~xliu/presentations/SRS1_project_report.PDF (accessed on 27 November 2023).
- Kozak, M.; Scaman, C.H. Unsupervised Classification Methods in Food Sciences: Discussion and Outlook. J. Sci. Food Agric. 2008, 88, 1115–1127. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Saha, S. Unsupervised Classification: Similarity Measures, Classical and Metaheuristic Approaches, and Applications; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-32450-5. [Google Scholar]
- Olaode, A.; Naghdy, G.; Todd, C. Unsupervised Classification of Images: A Review. Int. J. Image Process. 2014, 8, 325–342. [Google Scholar]
- Ma, Z.; Liu, Z.; Zhao, Y.; Zhang, L.; Liu, D.; Ren, T.; Zhang, X.; Li, S. An Unsupervised Crop Classification Method Based on Principal Components Isometric Binning. ISPRS Int. J. Geo-Inf. 2020, 9, 648. [Google Scholar] [CrossRef]
- Anchang, J.Y.; Ananga, E.O.; Pu, R. An Efficient Unsupervised Index Based Approach for Mapping Urban Vegetation from IKONOS Imagery. Int. J. Appl. Earth Obs. Geoinf. 2016, 50, 211–220. [Google Scholar] [CrossRef]
- Ragettli, S.; Herberz, T.; Siegfried, T. An Unsupervised Classification Algorithm for Multi-Temporal Irrigated Area Mapping in Central Asia. Remote Sens. 2018, 10, 1823. [Google Scholar] [CrossRef]
- Landucci, F.; Tichý, L.; Šumberová, K.; Chytrý, M. Formalized Classification of Species-poor Vegetation: A Proposal of a Consistent Protocol for Aquatic Vegetation. J Veg. Sci. 2015, 26, 791–803. [Google Scholar] [CrossRef]
- Alvarez, M.; Luebert, F. Chilean Vegetation in the Context of the Braun-Blanquet Approach and a Comparison with EcoVeg Formations. Veg. Classif. Surv. 2022, 3, 45–52. [Google Scholar] [CrossRef]
- Jennings, M.D.; Faber-Langendoen, D.; Loucks, O.L.; Peet, R.K.; Roberts, D. Standards for Associations and Alliances of the U.S. National Vegetation Classification. Ecol. Monogr. 2009, 79, 173–199. [Google Scholar] [CrossRef]
- Yermolaev, O.; Usmanov, B.; Gafurov, A.; Poesen, J.; Vedeneeva, E.; Lisetskii, F.; Nicu, I.C. Assessment of Shoreline Transformation Rates and Landslide Monitoring on the Bank of Kuibyshev Reservoir (Russia) Using Multi-Source Data. Remote Sens. 2021, 13, 4214. [Google Scholar] [CrossRef]
- Perevedentsev, Y.P.; Shantalinskii, K.M.; Guryanov, V.V.; Aukhadeev, T.R. Climatic Changes on the Territory of the Volga Federal District. IOP Conf. Ser. Earth Environ. Sci. 2020, 606, 012045. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933. [Google Scholar] [CrossRef]
- Yermolaev, O.; Usmanov, B. The Basin Approach to the Anthropogenic Impact Assessment in Oil-Producing Region. In Proceedings of the International Multidisciplinary Scientific GeoConference Surveying Geology and Mining Ecology Management, SGEM, Albena, Bulgaria, 17 June 2014; Volume 2, pp. 681–688. [Google Scholar]
- Prokhorov, V.; Rogova, T.; Kozhevnikova, M. Vegetation Database of Tatarstan. Phytocoenologia 2017, 47, 309–313. [Google Scholar] [CrossRef]
- Page, R.D.M. The Plant List with Literature 2016. Available online: https://www.gbif.org/dataset/d9a4eedb-e985-4456-ad46-3df8472e00e8 (accessed on 27 November 2023). [CrossRef]
- The WFO Plant List|World Flora Online. Available online: https://wfoplantlist.org/plant-list (accessed on 27 November 2023).
- Kozhevnikova, M.V.; Prokhorov, V.E.; Rogova, T.V. Xeromesophytic Broad-Leaved Forest Communities of the Republic of Tatarstan in the Hierarchy of Syntaxa within the Braun-Blanquet System. Uchenye Zap. Kazan. Universiteta. Seriya Estestv. Nauk. 2018, 160, 445–458. [Google Scholar]
- Kozhevnikova, M.V.; Prokhorov, V.E.; Saveliev, A.A. Predictive Modeling for the Distribution of Plant Communities of the Order Quercetalia Pubescenti-Petraeae Klika 1933. Tomsk State Univ. J. Biol. 2019, 47, 59–73. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, Software for Vegetation Classification. J Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Pickens, A.H.; Tyukavina, A.; Hernandez-Serna, A.; Zalles, V.; Turubanova, S.; Kommareddy, I.; Stehman, S.V.; Song, X.-P.; et al. Global Land Use Extent and Dispersion within Natural Land Cover Using Landsat Data. Environ. Res. Lett. 2022, 17, 034050. [Google Scholar] [CrossRef]
- Roujean, J.-L.; Breon, F.-M. Estimating PAR Absorbed by Vegetation from Bidirectional Reflectance Measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Yang, W.; Kobayashi, H.; Wang, C.; Shen, M.; Chen, J.; Matsushita, B.; Tang, Y.; Kim, Y.; Bret-Harte, M.S.; Zona, D.; et al. A Semi-Analytical Snow-Free Vegetation Index for Improving Estimation of Plant Phenology in Tundra and Grassland Ecosystems. Remote Sens. Environ. 2019, 228, 31–44. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a Green Channel in Remote Sensing of Global Vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Wang, F.; Huang, J.; Tang, Y.; Wang, X. New Vegetation Index and Its Application in Estimating Leaf Area Index of Rice. Rice Sci. 2007, 14, 195–203. [Google Scholar] [CrossRef]
- Jurgens, C. The Modified Normalized Difference Vegetation Index (mNDVI) a New Index to Determine Frost Damages in Agriculture Based on Landsat TM Data. Int. J. Remote Sens. 1997, 18, 3583–3594. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Bannari, A.; Asalhi, H.; Teillet, P.M. Transformed Difference Vegetation Index (TDVI) for Vegetation Cover Mapping. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Toronto, ON, Canada, 24–28 June 2002; IEEE: Toronto, ON, Canada, 2002; Volume 5, pp. 3053–3055. [Google Scholar]
- Gitelson, A.A. Wide Dynamic Range Vegetation Index for Remote Quantification of Biophysical Characteristics of Vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Clevers, J.G.P.W. Application of a Weighted Infrared-Red Vegetation Index for Estimating Leaf Area Index by Correcting for Soil Moisture. Remote Sens. Environ. 1989, 29, 25–37. [Google Scholar] [CrossRef]
- Apan, A.; Held, A.; Phinn, S.; Markley, J. Detecting Sugarcane ‘Orange Rust’ Disease Using EO-1 Hyperion Hyperspectral Imagery. Int. J. Remote Sens. 2004, 25, 489–498. [Google Scholar] [CrossRef]
- Sripada, R.P.; Heiniger, R.W.; White, J.G.; Weisz, R. Aerial Color Infrared Photography for Determining Late-Season Nitrogen Requirements in Corn. Agron. J. 2005, 97, 1443–1451. [Google Scholar] [CrossRef]
- Badgley, G.; Field, C.B.; Berry, J.A. Canopy Near-Infrared Reflectance and Terrestrial Photosynthesis. Sci. Adv. 2017, 3, e1602244. [Google Scholar] [CrossRef]
- Zeng, Y.; Hao, D.; Badgley, G.; Damm, A.; Rascher, U.; Ryu, Y.; Johnson, J.; Krieger, V.; Wu, S.; Qiu, H.; et al. Estimating Near-Infrared Reflectance of Vegetation from Hyperspectral Data. Remote Sens. Environ. 2021, 267, 112723. [Google Scholar] [CrossRef]
- Klemas, V.; Smart, R. The Influence of Soil Salinity, Growth Form, and Leaf Moisture on-the Spectral Radiance of. Photogramm. Eng. Remote Sens. 1983, 49, 77–83. [Google Scholar]
- Wilson, E.H.; Sader, S.A. Detection of Forest Harvest Type Using Multiple Dates of Landsat TM Imagery. Remote Sens. Environ. 2002, 80, 385–396. [Google Scholar] [CrossRef]
- Wang, L.; Qu, J.J. NMDI: A Normalized Multi-Band Drought Index for Monitoring Soil and Vegetation Moisture with Satellite Remote Sensing. Geophys. Res. Lett. 2007, 34, L20405. [Google Scholar] [CrossRef]
- Huntjr, E.; Rock, B. Detection of Changes in Leaf Water Content Using Near- and Middle-Infrared Reflectances☆. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar] [CrossRef]
- Wu, W. The Generalized Difference Vegetation Index (GDVI) for Dryland Characterization. Remote Sens. 2014, 6, 1211–1233. [Google Scholar] [CrossRef]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A Modified Soil Adjusted Vegetation Index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of Soil-Adjusted Vegetation Indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Kaufman, Y.J.; Tanre, D. Atmospherically Resistant Vegetation Index (ARVI) for EOS-MODIS. IEEE Trans. Geosci. Remote Sens. 1992, 30, 261–270. [Google Scholar] [CrossRef]
- Huete, A.R. A Soil-Adjusted Vegetation Index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G.; Major, D.J. TSAVI: A Vegetation Index which Minimizes Soil Brightness Effects on LAI and APAR Estimation. In Proceedings of the 12th Canadian Symposium on Remote Sensing Geoscience and Remote Sensing Symposium, Vancouver, CA, Canada, 10–14 July 1989; IEEE: Vancouver, Canada, 1989; Volume 3, pp. 1355–1358. [Google Scholar]
- Gillespie, A.R.; Kahle, A.B.; Walker, R.E. Color Enhancement of Highly Correlated Images. II. Channel Ratio and “Chromaticity” Transformation Techniques. Remote Sens. Environ. 1987, 22, 343–365. [Google Scholar] [CrossRef]
- Bendig, J.; Yu, K.; Aasen, H.; Bolten, A.; Bennertz, S.; Broscheit, J.; Gnyp, M.L.; Bareth, G. Combining UAV-Based Plant Height from Crop Surface Models, Visible, and near Infrared Vegetation Indices for Biomass Monitoring in Barley. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 79–87. [Google Scholar] [CrossRef]
- Haboudane, D. Hyperspectral Vegetation Indices and Novel Algorithms for Predicting Green LAI of Crop Canopies: Modeling and Validation in the Context of Precision Agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Chen, J.M. Evaluation of Vegetation Indices and a Modified Simple Ratio for Boreal Applications. Can. J. Remote Sens. 1996, 22, 229–242. [Google Scholar] [CrossRef]
- Vincini, M.; Frazzi, E.; D’Alessio, P. A Broad-Band Leaf Chlorophyll Vegetation Index at the Canopy Scale. Precis. Agric. 2008, 9, 303–319. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel Algorithms for Remote Estimation of Vegetation Fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef]
- Vincini, M.; Frazzi, E. Comparing Narrow and Broad-Band Vegetation Indices to Estimate Leaf Chlorophyll Content in Planophile Crop Canopies. Precis. Agric 2011, 12, 334–344. [Google Scholar] [CrossRef]
- Ill, J.E.P.; McLeod, K.W. Indications of Relative Drought Stress in Longleaf Pine from Thematic Mapper Data. Photogramm. Eng. Remote Sens. 1999, 65, 495–501. [Google Scholar]
- Jordan, C.F. Derivation of Leaf-Area Index from Quality of Light on the Forest Floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.; Didan, K.; Miura, T. Development of a Two-Band Enhanced Vegetation Index without a Blue Band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Louhaichi, M.; Borman, M.M.; Johnson, D.E. Spatially Located Platform and Aerial Photography for Documentation of Grazing Impacts on Wheat. Geocarto Int. 2001, 16, 65–70. [Google Scholar] [CrossRef]
- Pinty, B.; Verstraete, M.M. GEMI: A Non-Linear Index to Monitor Global Vegetation from Satellites. Vegetatio 1992, 101, 15–20. [Google Scholar] [CrossRef]
- Escadafal, R.; Huete, A. Etude Des Propriétés Spectrales Des Sols Arides Appliquée à l’amélioration Des Indices de Végétation Obtenus Par Télédétection. Comptes Rendus de L’académie Des Sci. Série 2 Mécanique Phys. Chim. Sci. L’univers Sci. Terre 1991, 312, 1385–1391. [Google Scholar]
- Rikimaru, A.; Roy, P.S.; Miyatake, S. Tropical Forest Cover Density Mapping. Trop. Ecol. 2002, 43, 39–47. [Google Scholar]
- Jiang, H.; Wang, S.; Cao, X.; Yang, C.; Zhang, Z.; Wang, X. A Shadow- Eliminated Vegetation Index (SEVI) for Removal of Self and Cast Shadow Effects on Vegetation in Rugged Terrains. Int. J. Digit. Earth 2019, 12, 1013–1029. [Google Scholar] [CrossRef]
- Hancock, D.W.; Dougherty, C.T. Relationships between Blue- and Red-based Vegetation Indices and Leaf Area and Yield of Alfalfa. Crops Sci. 2007, 47, 2547–2556. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between Leaf Chlorophyll Content and Spectral Reflectance and Algorithms for Non-Destructive Chlorophyll Assessment in Higher Plant Leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef]
- Yang, P.; Van Der Tol, C.; Campbell, P.K.E.; Middleton, E.M. Fluorescence Correction Vegetation Index (FCVI): A Physically Based Reflectance Index to Separate Physiological and Non-Physiological Information in Far-Red Sun-Induced Chlorophyll Fluorescence. Remote Sens. Environ. 2020, 240, 111676. [Google Scholar] [CrossRef]
- Ren-hua, Z.; Rao, N.X.; Liao, K.N. Approach for a Vegetation Index Resistant to Atmospheric Effect. J. Integr. Plant Biol. 1996, 38, 53–62. [Google Scholar]
- Kawashima, S. An Algorithm for Estimating Chlorophyll Content in Leaves Using a Video Camera. Ann. Bot. 1998, 81, 49–54. [Google Scholar] [CrossRef]
- Crippen, R. Calculating the Vegetation Index Faster. Remote Sens. Environ. 1990, 34, 71–73. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, H.; Wu, Z.; Wang, S.; Sun, H.; Senthilnath, J.; Wang, J.; Robin Bryant, C.; Fu, Y. Modified Red Blue Vegetation Index for Chlorophyll Estimation and Yield Prediction of Maize from Visible Images Captured by UAV. Sensors 2020, 20, 5055. [Google Scholar] [CrossRef]
- Gong, P.; Pu, R.; Biging, G.S.; Larrieu, M.R. Estimation of Forest Leaf Area Index Using Vegetation Indices Derived from Hyperion Hyperspectral Data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1355–1362. [Google Scholar] [CrossRef]
- Gu, Y.; Brown, J.F.; Verdin, J.P.; Wardlow, B. A Five-Year Analysis of MODIS NDVI and NDWI for Grassland Drought Assessment over the Central Great Plains of the United States. Geophys. Res. Lett. 2007, 34, L06407. [Google Scholar] [CrossRef]
- Wang, C.; Chen, J.; Wu, J.; Tang, Y.; Shi, P.; Black, T.A.; Zhu, K. A Snow-Free Vegetation Index for Improved Monitoring of Vegetation Spring Green-up Date in Deciduous Ecosystems. Remote Sens. Environ. 2017, 196, 1–12. [Google Scholar] [CrossRef]
- Sulik, J.J.; Long, D.S. Spectral Considerations for Modeling Yield of Canola. Remote Sens. Environ. 2016, 184, 161–174. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Han, J.; Zhang, Z.; Cao, J. Developing a New Method to Identify Flowering Dynamics of Rapeseed Using Landsat 8 and Sentinel-1/2. Remote Sens. 2020, 13, 105. [Google Scholar] [CrossRef]
- Saberioon, M.M.; Amin, M.S.M.; Anuar, A.R.; Gholizadeh, A.; Wayayok, A.; Khairunniza-Bejo, S. Assessment of Rice Leaf Chlorophyll Content Using Visible Bands at Different Growth Stages at Both the Leaf and Canopy Scale. Int. J. Appl. Earth Obs. Geoinf. 2014, 32, 35–45. [Google Scholar] [CrossRef]
- Hunt, E.R.; Doraiswamy, P.C.; McMurtrey, J.E.; Daughtry, C.S.T.; Perry, E.M.; Akhmedov, B. A Visible Band Index for Remote Sensing Leaf Chlorophyll Content at the Canopy Scale. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 103–112. [Google Scholar] [CrossRef]
- Broge, N.H.; Leblanc, E. Comparing Prediction Power and Stability of Broadband and Hyperspectral Vegetation Indices for Estimation of Green Leaf Area Index and Canopy Chlorophyll Density. Remote Sens. Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- McKight, P.E.; Najab, J. Kruskal-Wallis Test. In The Corsini Encyclopedia of Psychology; Weiner, I.B., Craighead, W.E., Eds.; Wiley: Hoboken, NJ, USA, 2010; ISBN 978-0-470-17024-3. [Google Scholar] [CrossRef]
- Larner, A.J. Effect Size (Cohen’s d) of Cognitive Screening Instruments Examined in Pragmatic Diagnostic Accuracy Studies. Dement. Geriatr. Cogn. Disord. Extra 2014, 4, 236–241. [Google Scholar] [CrossRef]
- Dinno, A. Nonparametric Pairwise Multiple Comparisons in Independent Groups Using Dunn’s Test. Stata J. 2015, 15, 292–300. [Google Scholar] [CrossRef]
- McKnight, P.E.; Najab, J. Mann-Whitney U Test. In The Corsini Encyclopedia of Psychology; Weiner, I.B., Craighead, W.E., Eds.; Wiley: Hoboken, NJ, USA, 2010; ISBN 978-0-470-17024-3. [Google Scholar] [CrossRef]
- Prieditis, N. Alnus Glutinosa—Dominated Wetland Forests of the Baltic Region: Community Structure, Syntaxonomy and Conservation. Plant Ecol. 1997, 129, 49–94. [Google Scholar] [CrossRef]
- Jun-hua, C. A Study of Characteristics of Community Structure of the Main Forests in the Guansi River Valley of Mianyang City. J. Sichuan For. Sci. Technol. 2006, 27, 41–46. [Google Scholar]
- Shim, I.; Kim, J.-B.; Jung, Y.-K.; Park, I.; Kim, M.-H.; Shin, H.; Cho, K. Eco-Floristic Characters of Vegetation in Successional Stages of Abandoned Paddy Fields. J. Korea Soc. Environ. Restor. Technol. 2015, 18, 29–41. [Google Scholar] [CrossRef][Green Version]
- Yang, L.P.; Ye, Q.Y.; Yang, S.H.; Wang, B.R. The features of Alnus nepalensis community in phosphate mining wasteland and its role in vegetation restoration. J. Yunnan Univ. 2004, 26, 234–237. [Google Scholar]
- Hoffmann, M.; Ermakov, N. Biogeographical Study of West Siberian Hemiboreal Forest Associations with Species Range Overlay Methods. Flora 2008, 203, 234–242. [Google Scholar] [CrossRef]
- Korolyuk, A.; Tishchenko, M.P.; Yamalov, S. Forest Meadows of the West Siberian Plain and Revision of the Order Carici Macrourae–Crepidetalia Sibiricae. Veg. Russ. 2016, 29, 67–88. [Google Scholar] [CrossRef]
- Pisarenko, O. Mosses in Brachypodio-Betuletea Forests of West Siberia. BIO Web Conf. 2019, 16, 00026. [Google Scholar] [CrossRef]
- Yin, X.; Zhou, G. Climatic suitability of the potential geographic distribution of Fagus longipetiolata in China. Environ. EarthSci. 2014, 73, 1143–1149. [Google Scholar] [CrossRef]
- Packham, J.; Thomas, P.; Atkinson, M.D.; Degen, T. Biological Flora of the British Isles: Fagus Sylvatica. J. Ecol. 2012, 100, 1557–1608. [Google Scholar] [CrossRef]
- Homma, K. Effects of Snow Pressure on Growth Form and Life History of Tree Species in Japanese Beech Forest. J. Veg. Sci. 1997, 8, 781–788. [Google Scholar] [CrossRef]
- Adel, M.; Pourbabaei, H.; Dey, D. Ecological Species Group—Environmental Factors Relationships in Unharvested Beech Forests in the North of Iran. Ecol. Eng. 2014, 69, 1–7. [Google Scholar] [CrossRef]
- Li, Y. Plant Species Diversity of Main Forest Community Types in San Jiangyuan National Nature Reserve. For. Res. 2007, 20, 241–245. [Google Scholar]
- He, S.; Ma, Y.; Liang, M.; Wang, D.; Fei, Y. Study on the Distribution and Community Characteristics of the Endangered Plant Picea Neoveitchii Mast. in Hubei Province. Asian Agric. Res. 2017, 9, 31–35. [Google Scholar] [CrossRef]
- Yang, X.; Shi, C.; Wang, X.; Ma, H.; Yan, H. Species Composition, Structure and Spatial Distribution of Secondary Cold-Temperate Picea Forest in Guandi Mountain, China. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2017, 28, 1421–1430. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Q.; Zhang, L.; Wang, X.; Cao, X.; Yang, X. Modern Pollen Assemblages of the Forest Communities and Their Relationships with Vegetation and Climate in Northern China. J. Geogr. Sci. 2009, 19, 643–659. [Google Scholar] [CrossRef]
- Yang, H.; Ma, Q.-R.; Yang, J.; Zhou, L.; Cao, B.; Zhang, W.-J. Characteristics of Soil Microbial Communities in Different Restoration Models in the Ecological Immigrants’ Emigration Area in Southern Ningxia, China. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2022, 33, 219–228. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Y.; Chen, Y.; Zhang, N.; Li, W. Degradation of Populus Euphratica Community in the Lower Reaches of the Tarim River, Xinjiang, China. J. Environ. Sci. 2005, 17, 740–747. [Google Scholar]
- Shi, H.; Shi, Q.; Zhou, X.; Imin, B.; Li, H.; Zhang, W.; Kahaer, Y. Effect of the Competition Mechanism of between Co-Dominant Species on the Ecological Characteristics of Populus Euphratica under a Water Gradient in a Desert Oasis. Glob. Ecol. Conserv. 2021, 27, e01611. [Google Scholar] [CrossRef]
- Li, M.; Song, Z.; Li, Z.; Qiao, R.; Zhang, P.; Ding, C.; Xie, J.; Chen, Y.; Guo, H. Populus Root Exudates Are Associated with Rhizosphere Microbial Communities and Symbiotic Patterns. Front. Microbiol. 2022, 13, 1042944. [Google Scholar] [CrossRef]
- Ya, C. Response of Anatomy and Hydraulic Characteristics of Xylem Stem of Populus euphratica Oliv. to Drought Stress. Chin. J. Eco-Agric. 2012, 20, 1059. [Google Scholar] [CrossRef]
- Fortini, P.; Marzio, P.D.; Conte, A.; Antonecchia, G.; Proietti, E.; Pietro, R. Morphological and Molecular Results from a Geographical Transect Focusing on Quercus pubescens/Q. virgiliana Ecological-Altitudinal Vicariance in Peninsular Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2022, 156, 1498–1511. [Google Scholar] [CrossRef]
- Demeter, Z.; Kanalas, P.; Máthé, C.; Cseke, K.; Szőllősi, E.; M-Hamvas, M.; Jámbrik, K.; Kiss, Z.; Mészáros, I. Osmotic Stress Responses of Individual White Oak (Quercus Section, Quercus Subgenus) Genotypes Cultured in Vitro. J. Plant Physiol. 2014, 171, 16–24. [Google Scholar] [CrossRef]
- George, L.O.; Bazzaz, F. The Fern Understory as an Ecological Filter: Growth and Survival of Canopy-Tree Seedlings. Ecology 1999, 80, 846–856. [Google Scholar] [CrossRef]
- And, C.; Rambal, S. Field Study of Leaf Photosynthetic Performance by a Mediterranean Deciduous Oak Tree (Quercus pubescens) during a Severe Summer Drought. New Phytol. 1995, 131, 159–167. [Google Scholar] [CrossRef]
- Nardini, A.; Pitt, F. Drought Resistance of Quercus pubescens as a Function of Root Hydraulic Conductance, Xylem Embolism and Hydraulic Architecture. New Phytol. 1999, 143, 485–493. [Google Scholar] [CrossRef]
- Fardeeva, M.; Lukyanova, Y.; Eskina, A.; Usmanov, B. Distribution and Habitat Features of Rare Orchid Species (Orchidaceae Juss.) in the National Park “Nizhnyaya Kama”. E3S Web Conf. 2023, 420, 01023. [Google Scholar] [CrossRef]
- Mukharamova, S.; Saveliev, A.; Ivanov, M.; Gafurov, A.; Yermolaev, O. Estimating the Soil Erosion Cover-Management Factor at the European Part of Russia. ISPRS Int. J. Geo-Inf. 2021, 10, 645. [Google Scholar] [CrossRef]
- Gafurov, A. Mapping of Rill Erosion of the Middle Volga (Russia) Region Using Deep Neural Network. ISPRS Int. J. Geo-Inf. 2022, 11, 197. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, W.; Lu, Y.; Zhang, W.; Wang, Y. Carbon Storage Dynamics of Secondary Forest Succession in the Central Loess Plateau of China. Forests 2019, 10, 342. [Google Scholar] [CrossRef]
- Kurbatova, A.I. Analytical Review of Modern Studies of Changes in the Biotic Components of the Carbon Cycle. Vestn. Ross. Univ. Družby Nar. Ser. Èkol. Bezop. Žiznedeât. 2020, 28, 428–438. [Google Scholar] [CrossRef]
- Hao, G.; Dong, Z. Vegetation Succession Accelerated the Accumulation of Soil Organic Carbon on Road-Cut Slopes by Changing the Structure of the Bacterial Community. Ecol. Eng. 2023, 197, 107118. [Google Scholar] [CrossRef]









| FC BB * | Raster Model Classes |
|---|---|
| ALN | 4 **, 5 **, 7 **, 17, 28, 42, 43 |
| BRA | 6 **, 36 ** |
| FAG | 11 **, 13, 14, 18, 19 **, 22 **, 23, 24, 29 **, 31, 34, 35, 37, 38, 44 ** |
| PIC | 2 **, 30 ** |
| POP | 1 **, 26, 33 **, 36 |
| PUB | 14 **, 25 **, 32 |
| OTHERS | *** |
| Class | Number of Pixels | Area (km2) | % of Area of RT Forests |
|---|---|---|---|
| ALN | 2,082,534 | 1874 | 16 |
| BRA | 440,568 | 397 | 3 |
| FAG | 6,370,003 | 5733 | 48 |
| PIC | 564,690 | 508 | 4 |
| POP | 876,324 | 789 | 7 |
| PUB | 1,320,184 | 1188 | 10 |
| OTHERS | 1,742,692 | 1568 | 13 |
| Feature | Importance | F-Value | p-Value |
|---|---|---|---|
| TriVI_max | 1.319% | 1093.94 | <0.0000 |
| TriVI_stdDev | 1.175% | 1046.95 | <0.0000 |
| TGI_mean | 0.934% | 435.02 | <0.0000 |
| SEVI_median | 0.919% | 319.74 | <0.0000 |
| TGI_median | 0.871% | 541.51 | <0.0000 |
| FCVI_max | 0.870% | 1075.45 | <0.0000 |
| SR_median | 0.828% | 580.48 | <0.0000 |
| GRVI_median | 0.792% | 607.80 | <0.0000 |
| TriVI_mean | 0.748% | 910.28 | <0.0000 |
| NDYI_median | 0.732% | 202.37 | <0.0000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gafurov, A.; Prokhorov, V.; Kozhevnikova, M.; Usmanov, B. Forest Community Spatial Modeling Using Machine Learning and Remote Sensing Data. Remote Sens. 2024, 16, 1371. https://doi.org/10.3390/rs16081371
Gafurov A, Prokhorov V, Kozhevnikova M, Usmanov B. Forest Community Spatial Modeling Using Machine Learning and Remote Sensing Data. Remote Sensing. 2024; 16(8):1371. https://doi.org/10.3390/rs16081371
Chicago/Turabian StyleGafurov, Artur, Vadim Prokhorov, Maria Kozhevnikova, and Bulat Usmanov. 2024. "Forest Community Spatial Modeling Using Machine Learning and Remote Sensing Data" Remote Sensing 16, no. 8: 1371. https://doi.org/10.3390/rs16081371
APA StyleGafurov, A., Prokhorov, V., Kozhevnikova, M., & Usmanov, B. (2024). Forest Community Spatial Modeling Using Machine Learning and Remote Sensing Data. Remote Sensing, 16(8), 1371. https://doi.org/10.3390/rs16081371