
Bitemporal Radiative Transfer Modeling Using Bitemporal 3D-Explicit Forest Reconstruction from Terrestrial Laser Scanning

 ,
,  , , ,
, , ,  , , ,
, , ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
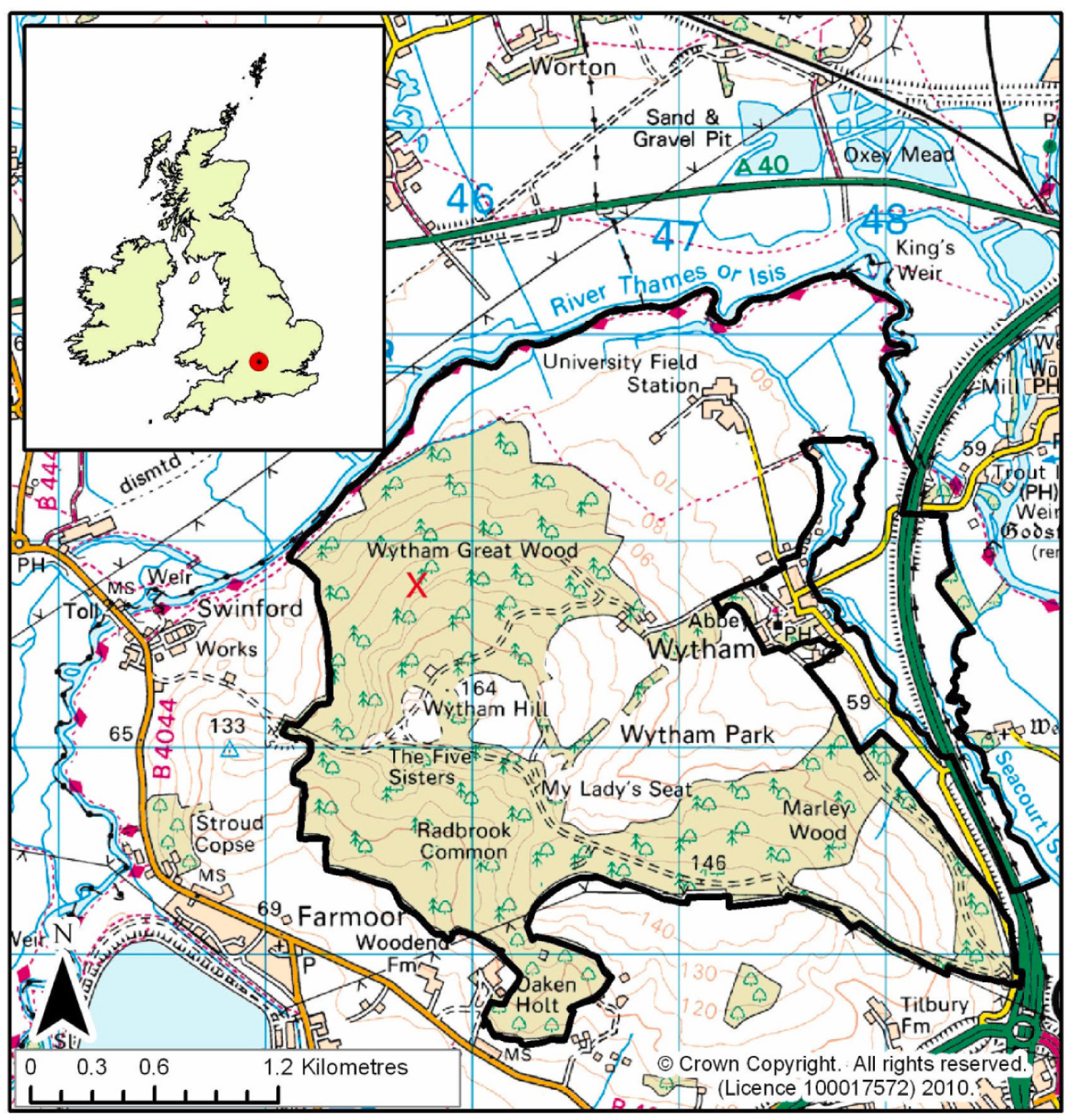
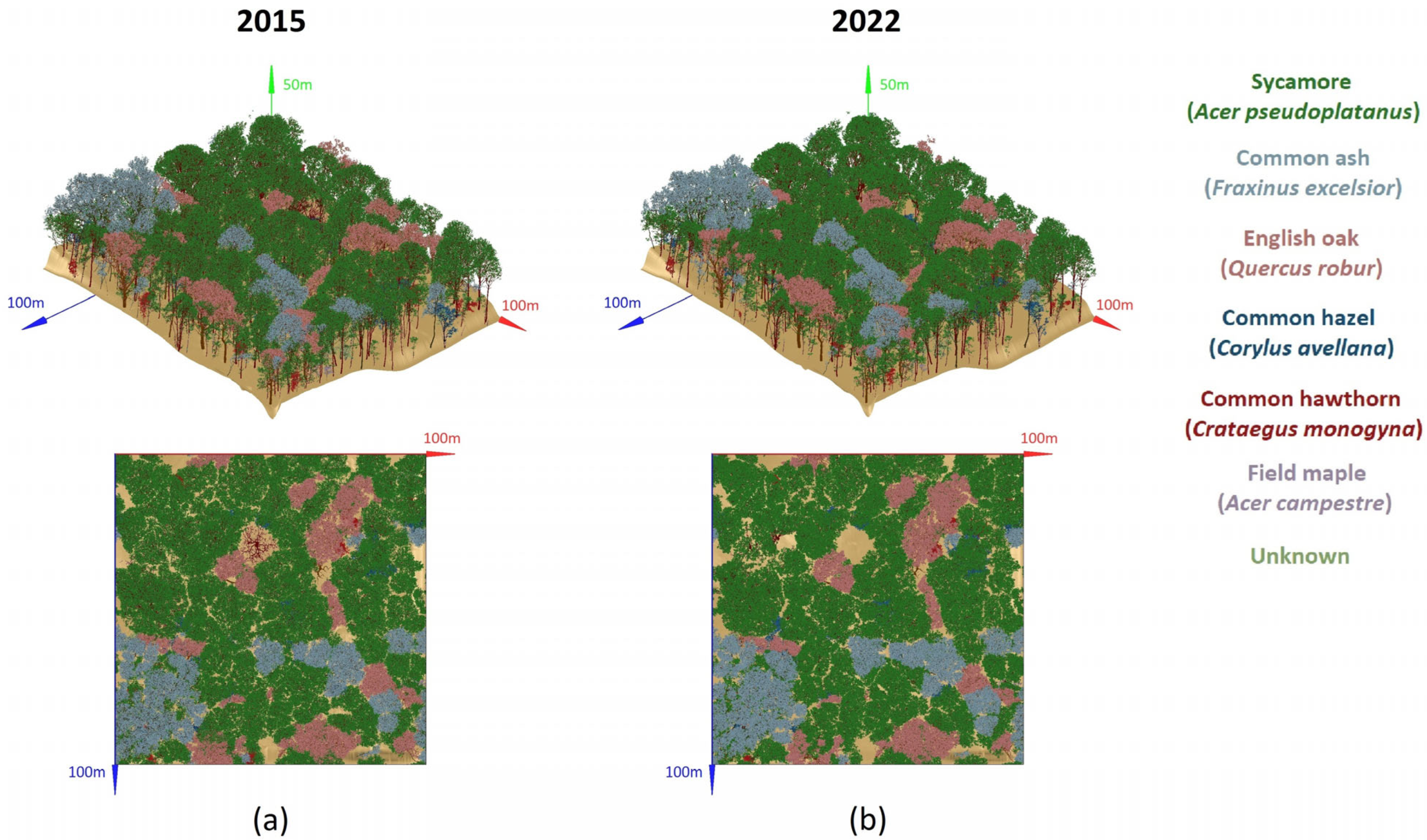
2.1. Study Area and Data Collection
2.2. 3D-Explicit Reconstruction of 2022 Forest Stand Structure
2.2.1. Tree Extraction
2.2.2. Quantitative Modeling of Tree Structure
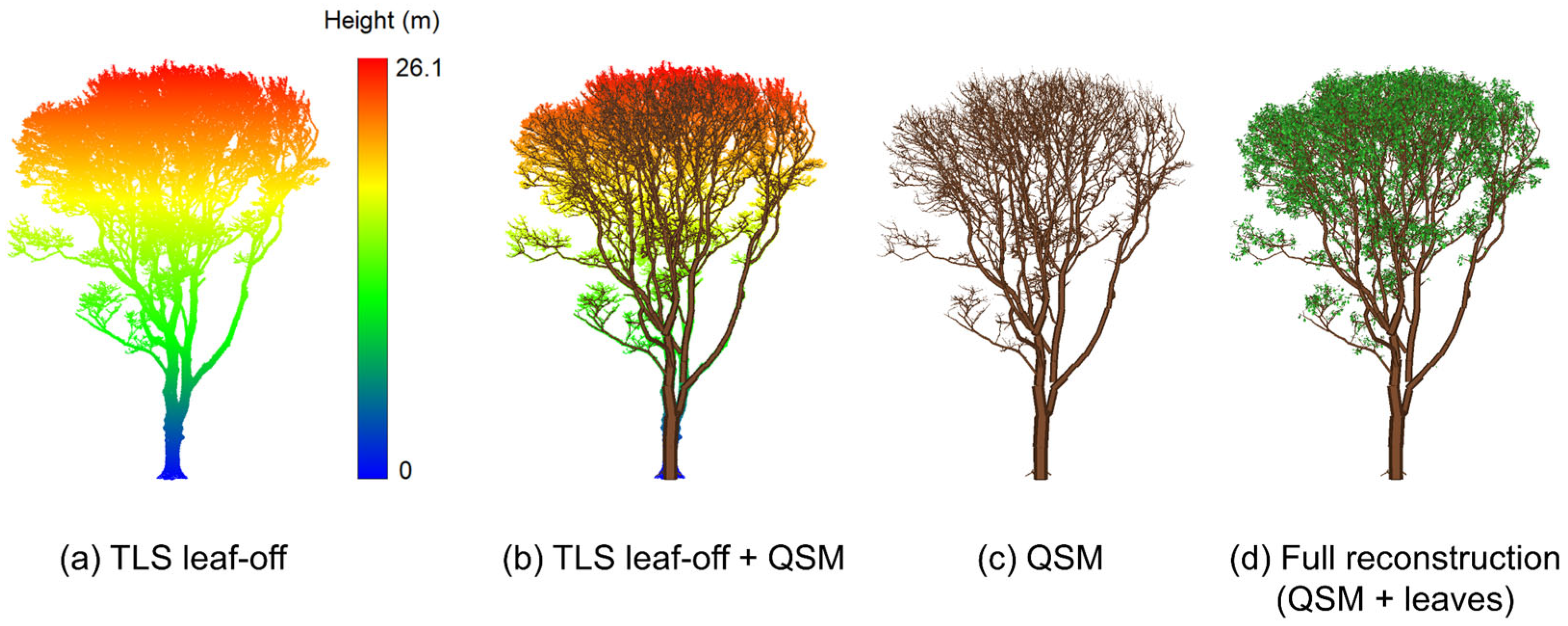
2.2.3. Leaf Addition
2.2.4. DTM Generation and Forest Stand Assembling
2.3. Radiative Transfer Modeling
2.3.1. Canopy Light Regimes at the Stand Scale
2.3.2. Canopy Light Availability at Explicit Locations
2.4. Evaluation of Computational Efficiency of Bitemporal 3D-Explicit Radiative Transfer Modeling
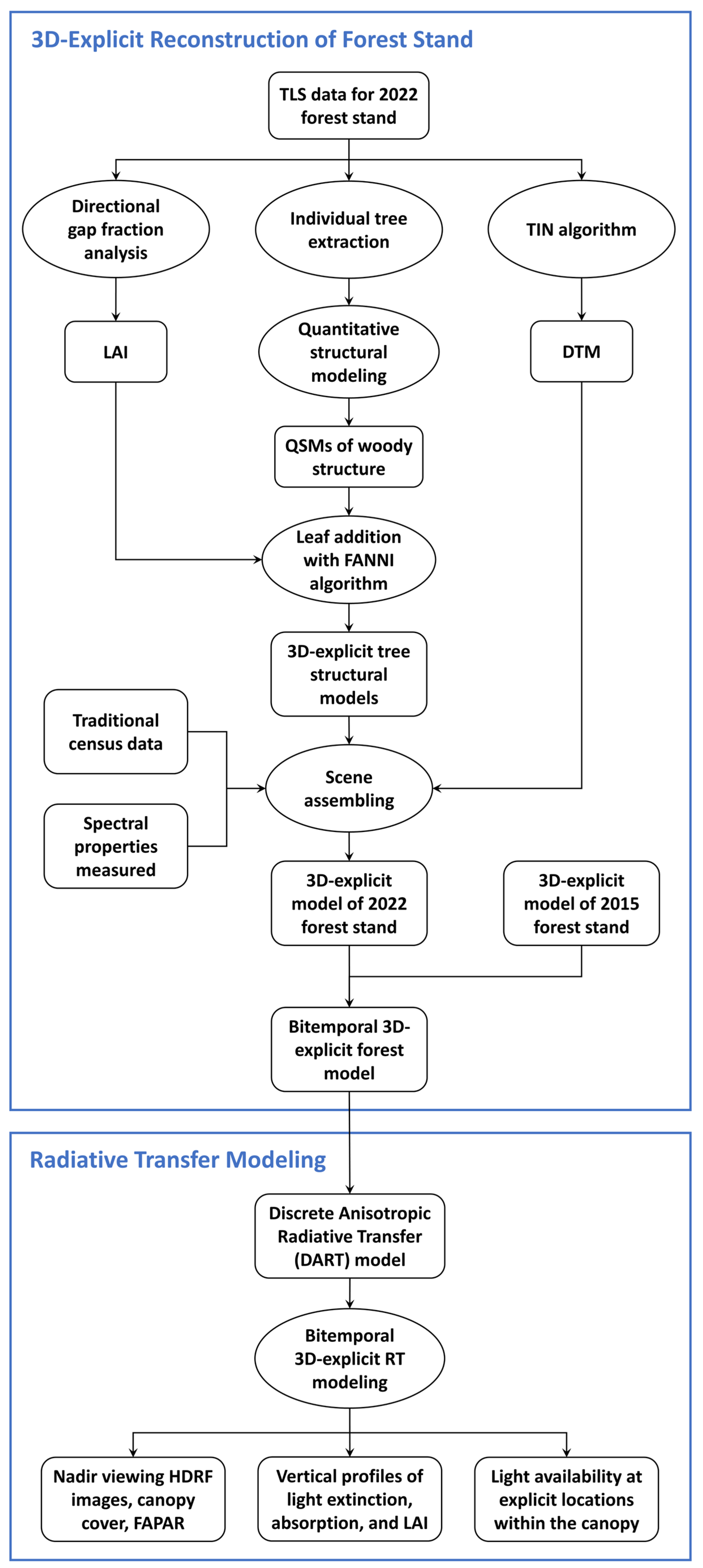
2.5. Flowchart of Research Methodology
3. Results
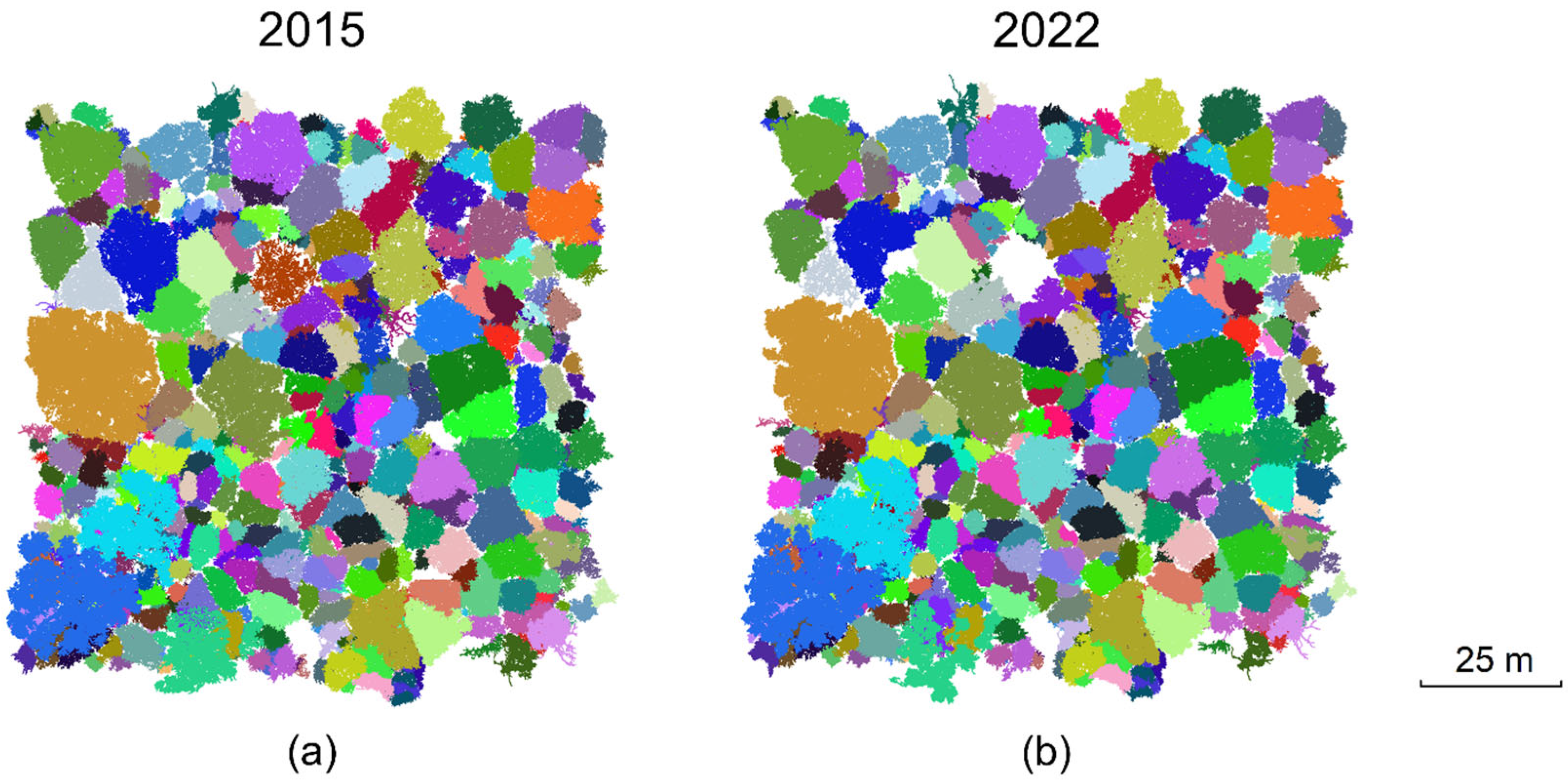
3.1. 3D-Explicit Reconstruction of 2022 Forest Stand
3.2. Bitemporal Radiative Transfer Modeling
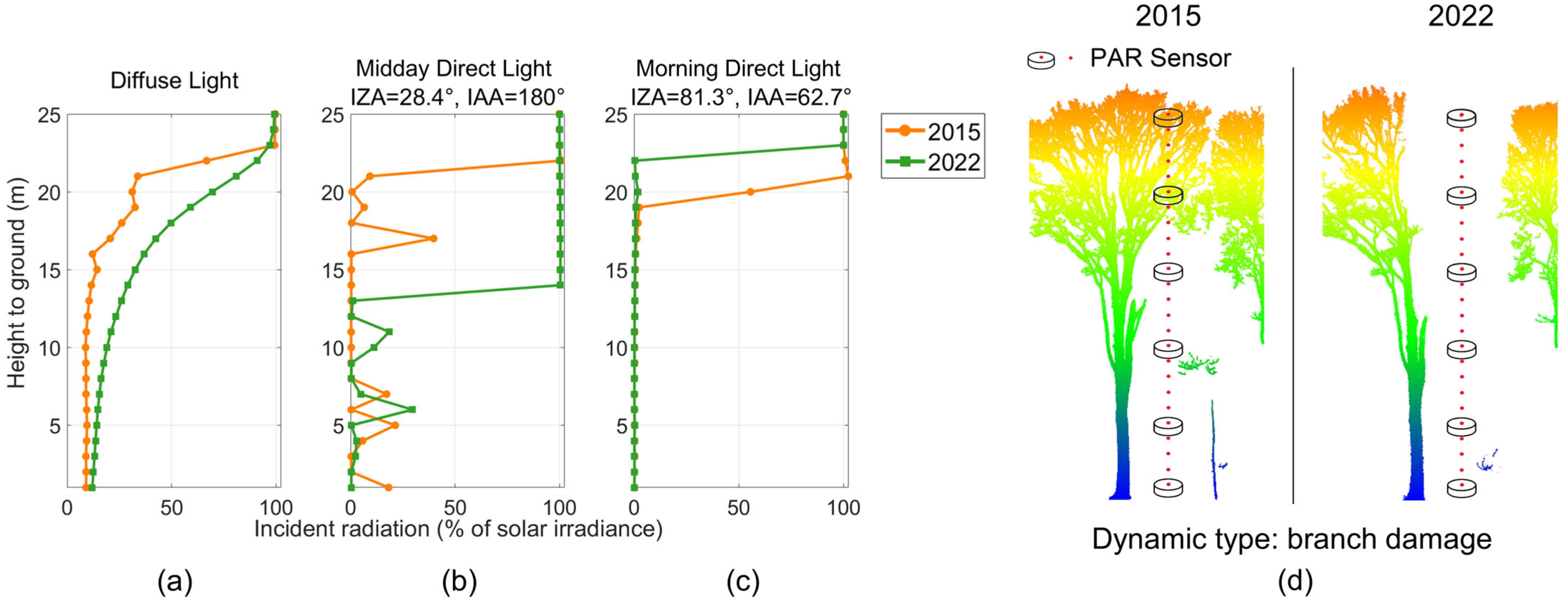
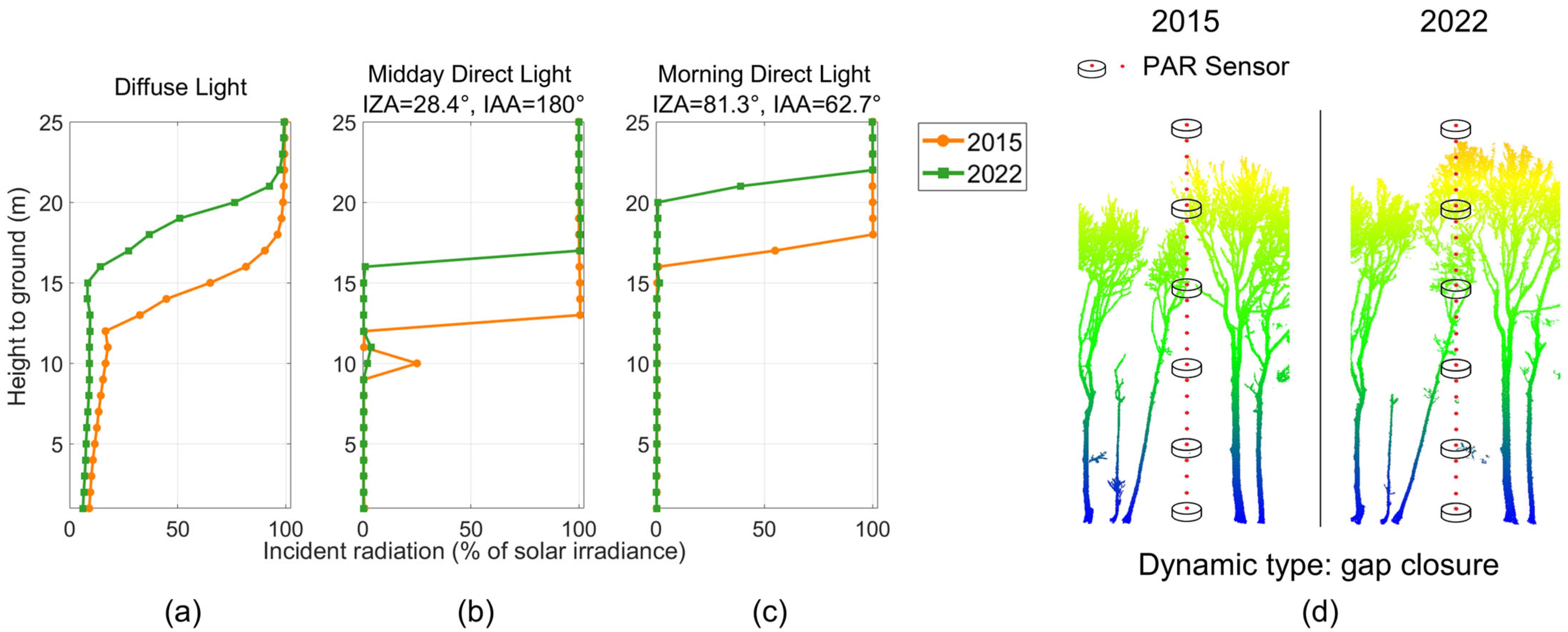
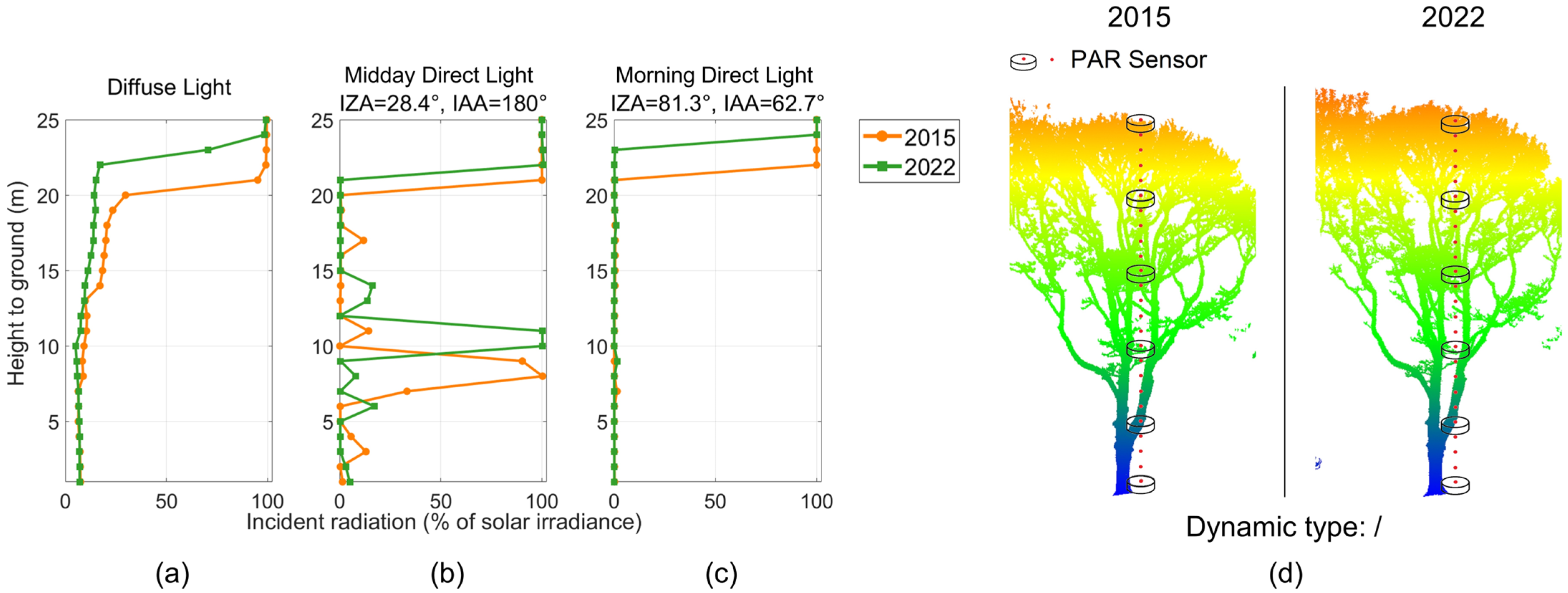
3.2.1. Vertically Resolved Radiative Transfer
3.2.2. Forward Modeling of Hemispherical-Directional Reflectance Factor
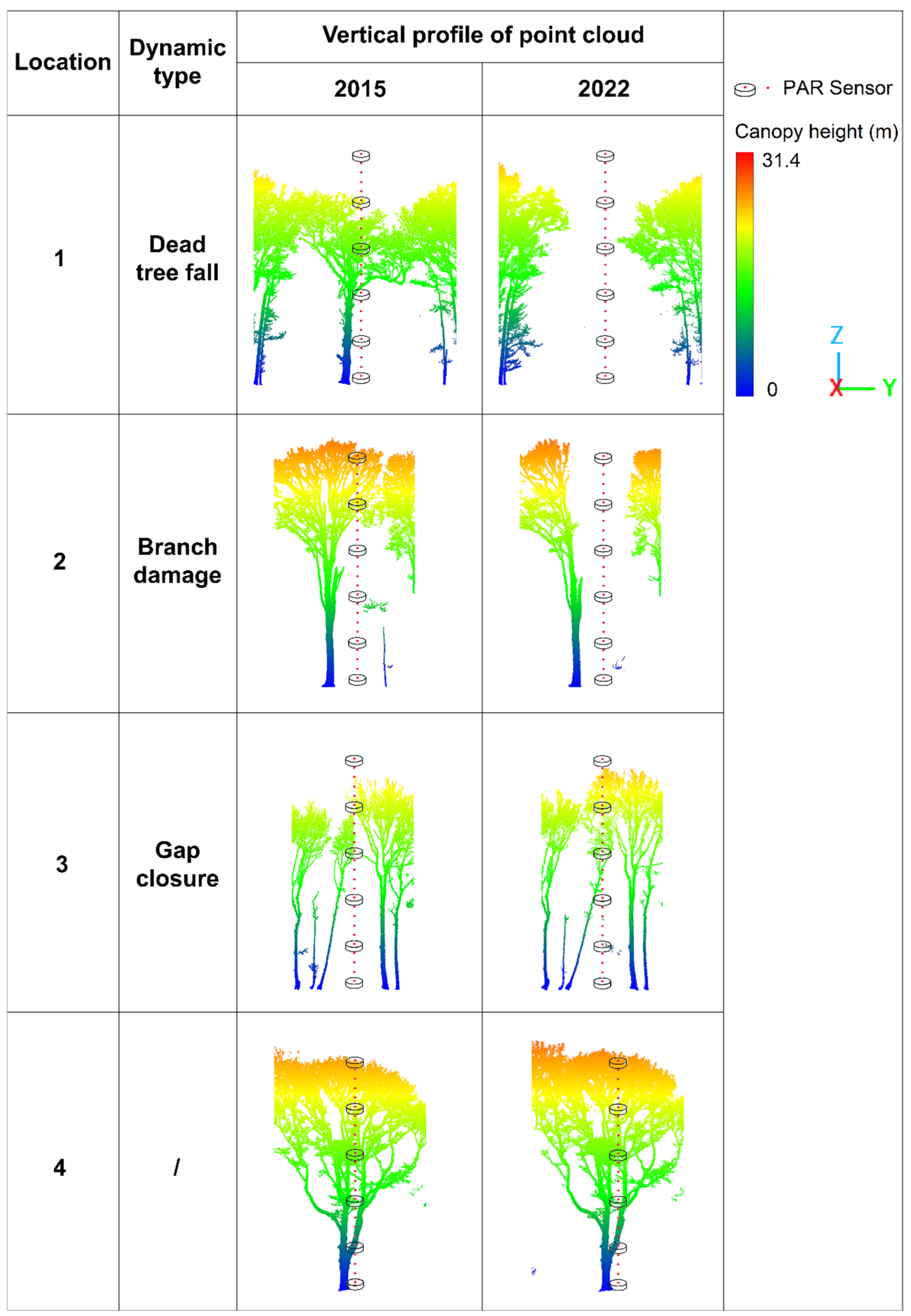
3.2.3. Impacts of Canopy Gap Dynamics on Light Availability
3.2.4. Computational Efficiency of Bitemporal 3D-Explicit Radiative Transfer Modeling
4. Discussion
4.1. Impacts of Forest Structural Dynamics on Radiative Transfer
4.2. Impacts of Canopy Gap Dynamics on Local Light Availability
4.3. Outlook
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bojinski, S.; Verstraete, M.; Peterson, T.C.; Richter, C.; Simmons, A.; Zemp, M. The Concept of Essential Climate Variables in Support of Climate Research, Applications, and Policy. Bull. Am. Meteorol. Soc. 2014, 95, 1431–1443. [Google Scholar] [CrossRef]
- Liu, C.; Calders, K.; Meunier, F.; Gastellu-Etchegorry, J.P.; Nightingale, J.; Disney, M.; Origo, N.; Woodgate, W.; Verbeeck, H. Implications of 3D Forest Stand Reconstruction Methods for Radiative Transfer Modeling: A Case Study in the Temperate Deciduous Forest. J. Geophys. Res. Atmos. 2022, 127, e2021JD036175. [Google Scholar] [CrossRef]
- Gastellu-Etchegorry, J.P.; Grau, E.; Lauret, N. DART: A 3D Model for Remote Sensing Images and Radiative Budget of Earth Surfaces. In Modeling and Simulation in Engineering; IntechOpen: London, UK, 2012. [Google Scholar]
- Widlowski, J.L.; Pinty, B.; Lopatka, M.; Atzberger, C.; Buzica, D.; Chelle, M.; Disney, M.; Gastellu-Etchegorry, J.P.; Gerboles, M.; Gobron, N.; et al. The Fourth Radiation Transfer Model Intercomparison (RAMI-IV): Proficiency Testing of Canopy Reflectance Models with ISO-13528. J. Geophys. Res. Atmos. 2013, 118, 6869–6890. [Google Scholar] [CrossRef]
- Widlowski, J.L.; CÔté, J.F.; Béland, M. Abstract Tree Crowns in 3D Radiative Transfer Models: Impact on Simulated Open-Canopy Reflectances. Remote Sens. Environ. 2014, 142, 155–175. [Google Scholar] [CrossRef]
- Modest, M.F.; Mazumder, S. Radiative Heat Transfer; Academic Press: Cambridge, MA, USA, 2021; ISBN 032398407X. [Google Scholar]
- Verhoef, W. Theory of Radiative Transfer Models Applied in Optical Remote Sensing of Vegetation Canopies. Ph.D. Thesis, Wageningen University and Research, Wageningen, The Netherlands, 1998. [Google Scholar]
- Ross, J. The Radiation Regime and Architecture of Plant Stands, 3rd ed.; Springer Science & Business Media: Berlin, Germany, 1981. [Google Scholar]
- Myneni, R.B.; Ross, J.; Asrar, G. A Review on the Theory of Photon Transport in Leaf Canopies. Agric. For. Meteorol. 1989, 45, 1–153. [Google Scholar] [CrossRef]
- Liang, S. Quantitative Remote Sensing of Land Surfaces; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 0471723711. [Google Scholar]
- Monsi, M. Uber Den Lichtfaktor in Den Pflanzengesellschaften Und Seine Bedeutung Fur Die Stoffproduktion. Jap. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Bailey, B.N.; Fu, K. The Probability Distribution of Absorbed Direct, Diffuse, and Scattered Radiation in Plant Canopies with Varying Structure. Agric. For. Meteorol. 2022, 322, 109009. [Google Scholar] [CrossRef]
- Goudriaan, J. Crop Micrometeorology a Simulation Study. Ph.D. Thesis, Wageningen University and Research, Wageningen, The Netherlands, 1977. [Google Scholar]
- Norman, J.M.M. Modeling the Complete Crop Canopy. In Modification of the Aerial Environment of Plants; American Society of Agricultural Engineers: St. Joseph, MO, USA, 1979. [Google Scholar]
- Bailey, B.N.; Ponce de León, M.A.; Scott Krayenhoff, E. One-Dimensional Models of Radiation Transfer in Heterogeneous Canopies: A Review, Re-Evaluation, and Improved Model. Geosci. Model Dev. 2020, 13, 4789–4808. [Google Scholar] [CrossRef]
- Smith, N.J.; Chen, J.M.; Black, T.A. Effects of Clumping on Estimates of Stand Leaf Area Index Using the LI-COR LAI-2000. Can. J. For. Res. 1993, 23, 1940–1943. [Google Scholar] [CrossRef]
- Nilson, T. A Theoretical Analysis of the Frequency of Gaps in Plant Stands. Agric. Meteorol. 1971, 8, 25–38. [Google Scholar] [CrossRef]
- Pinty, B.; Lavergne, T.; Dickinson, R.E.; Widlowski, J.L.; Gobron, N.; Verstraete, M.M. Simplifying the Interaction of Land Surfaces with Radiation for Relating Remote Sensing Products to Climate Models. J. Geophys. Res. Atmos. 2006, 111, D2. [Google Scholar] [CrossRef]
- Verstraete, M.M.; Pinty, B.; Dickinson, R.E. A Physical Model of the Bidirectional Reflectance of Vegetation Canopies: 1. Theory. J. Geophys. Res. 1990, 95, 11755–11765. [Google Scholar] [CrossRef]
- Kuusk, A. The Hot Spot Effect of a Uniform Vegetative Cover. Sov. J. Remote Sens. 1985, 3, 645–658. [Google Scholar]
- Kuusk, A. The Hot Spot Effect in Plant Canopy Reflectance. In Photon-Vegetation Interactions; Springer: Berlin, Germany, 1991. [Google Scholar]
- Gastellu-Etchegorry, J.P.; Yin, T.; Lauret, N.; Cajgfinger, T.; Gregoire, T.; Grau, E.; Feret, J.B.; Lopes, M.; Guilleux, J.; Dedieu, G.; et al. Discrete Anisotropic Radiative Transfer (DART 5) for Modeling Airborne and Satellite Spectroradiometer and LIDAR Acquisitions of Natural and Urban Landscapes. Remote Sens. 2015, 7, 1667–1701. [Google Scholar] [CrossRef]
- Verhoef, W. Light Scattering by Leaf Layers with Application to Canopy Reflectance Modeling: The SAIL Model. Remote Sens. Environ. 1984, 16, 125–141. [Google Scholar] [CrossRef]
- Borel, C.C.; Gerstl, S.A.W.; Powers, B.J. The Radiosity Method in Optical Remote Sensing of Structured 3-D Surfaces. Remote Sens. Environ. 1991, 36, 13–44. [Google Scholar] [CrossRef]
- Duan, M.; Min, Q.; Lü, D. A Polarized Radiative Transfer Model Based on Successive Order of Scattering. Adv. Atmos. Sci. 2010, 27, 891–900. [Google Scholar] [CrossRef]
- Glassner, A.S. An Introduction to Ray Tracing; Morgan Kaufmann: Burlington, MA, USA, 1989; ISBN 0122861604. [Google Scholar]
- Govaerts, Y.M.; Verstraete, M.M. Raytran: A Monte Carlo Ray-Tracing Model to Compute Light Scattering in Three-Dimensional Heterogeneous Media. IEEE Trans. Geosci. Remote Sens. 1998, 36, 662732. [Google Scholar] [CrossRef]
- Lewis, P. Three-Dimensional Plant Modelling for Remote Sensing Simulation Studies Using the Botanical Plant Modelling System. Agronomie 1999, 19, 185–210. [Google Scholar] [CrossRef]
- North, P.R.J. Three-Dimensional Forest Light Interaction Model Using a Monte Carlo Method. IEEE Trans. Geosci. Remote Sens. 1996, 34, 946–956. [Google Scholar] [CrossRef]
- España, M.; Baret, F.; Aries, F.; Andrieu, B.; Chelle, M. Radiative Transfer Sensitivity to the Accuracy of Canopy Structure Description. The Case of a Maize Canopy. Agronomie 1999, 19, 241–254. [Google Scholar] [CrossRef]
- Wang, Y.; Kallel, A.; Yang, X.; Regaieg, O.; Lauret, N.; Guilleux, J.; Chavanon, E.; Gastellu-Etchegorry, J.P. DART-Lux: An Unbiased and Rapid Monte Carlo Radiative Transfer Method for Simulating Remote Sensing Images. Remote Sens. Environ. 2022, 274, 112973. [Google Scholar] [CrossRef]
- Widlowski, J.L.; Lavergne, T.; Pinty, B.; Verstraete, M.; Gobron, N. Rayspread: A Virtual Laboratory for Rapid BRF Simulations over 3-D Plant Canopies. Lect. Notes Comput. Sci. Eng. 2006, 211–231. [Google Scholar] [CrossRef]
- Disney, M.; Lewis, P.; Saich, P. 3D Modelling of Forest Canopy Structure for Remote Sensing Simulations in the Optical and Microwave Domains. Remote Sens. Environ. 2006, 100, 114–132. [Google Scholar] [CrossRef]
- Janoutová, R.; Homolová, L.; Malenovskỳ, Z.; Hanuš, J.; Lauret, N.; Gastellu-Etchegorry, J.P. Influence of 3D Spruce Tree Representation on Accuracy of Airborne and Satellite Forest Reflectance Simulated in DART. Forests 2019, 10, 292. [Google Scholar] [CrossRef]
- Li, W.; Guo, Q.; Tao, S.; Su, Y. VBRT: A Novel Voxel-Based Radiative Transfer Model for Heterogeneous Three-Dimensional Forest Scenes. Remote Sens. Environ. 2018, 206, 318–335. [Google Scholar] [CrossRef]
- Van der Zande, D.; Stuckens, J.; Verstraeten, W.W.; Mereu, S.; Muys, B.; Coppin, P. 3D Modeling of Light Interception in Heterogeneous Forest Canopies Using Ground-Based LiDAR Data. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 792–800. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; François, C.; Ustin, S.L. PROSPECT + SAIL Models: A Review of Use for Vegetation Characterization. Remote Sens. Environ. 2009, 113, S56–S66. [Google Scholar] [CrossRef]
- Cescatti, A. Modelling the Radiative Transfer in Discontinuous Canopies of Asymmetric Crowns. I. Model Structure and Algorithms. Ecol. Modell. 1997, 101, 263–274. [Google Scholar] [CrossRef]
- García, M.; North, P.; Viana-Soto, A.; Stavros, N.E.; Rosette, J.; Martín, M.P.; Franquesa, M.; González-Cascón, R.; Riaño, D.; Becerra, J.; et al. Evaluating the Potential of LiDAR Data for Fire Damage Assessment: A Radiative Transfer Model Approach. Remote Sens. Environ. 2020, 247, 111893. [Google Scholar] [CrossRef]
- Kobayashi, H.; Iwabuchi, H. A Coupled 1-D Atmosphere and 3-D Canopy Radiative Transfer Model for Canopy Reflectance, Light Environment, and Photosynthesis Simulation in a Heterogeneous Landscape. Remote Sens. Environ. 2008, 112, 173–185. [Google Scholar] [CrossRef]
- Ni, W.; Li, X.; Woodcock, C.E.; Caetano, M.R.; Strahler, A.H. An Analytical Hybrid GORT Model for Bidirectional Reflectance over Discontinuous Plant Canopies. IEEE Trans. Geosci. Remote Sens. 1999, 37, 987–999. [Google Scholar] [CrossRef]
- Calders, K.; Lewis, P.; Disney, M.; Verbesselt, J.; Herold, M. Investigating Assumptions of Crown Archetypes for Modelling LiDAR Returns. Remote Sens. Environ. 2013, 134, 39–49. [Google Scholar] [CrossRef]
- Schneider, F.D.; Leiterer, R.; Morsdorf, F.; Gastellu-Etchegorry, J.P.; Lauret, N.; Pfeifer, N.; Schaepman, M.E. Simulating Imaging Spectrometer Data: 3D Forest Modeling Based on LiDAR and in Situ Data. Remote Sens. Environ. 2014, 152, 235–250. [Google Scholar] [CrossRef]
- Disney, M.I.; Lewis, P.; Gomez-Dans, J.; Roy, D.; Wooster, M.J.; Lajas, D. 3D Radiative Transfer Modelling of Fire Impacts on a Two-Layer Savanna System. Remote Sens. Environ. 2011, 115, 1866–1881. [Google Scholar] [CrossRef]
- Disney, M.I.; Kalogirou, V.; Lewis, P.; Prieto-Blanco, A.; Hancock, S.; Pfeifer, M. Simulating the Impact of Discrete-Return Lidar System and Survey Characteristics over Young Conifer and Broadleaf Forests. Remote Sens. Environ. 2010, 114, 1546–1560. [Google Scholar] [CrossRef]
- Disney, M.I.; Lewis, P.E.; Bouvet, M.; Prieto-Blanco, A.; Hancock, S. Quantifying Surface Reflectivity for Spaceborne Lidar via Two Independent Methods. IEEE Trans. Geosci. Remote Sens. 2009, 47, 3262–3271. [Google Scholar] [CrossRef]
- Stuckens, J.; Somers, B.; Delalieux, S.; Verstraeten, W.W.; Coppin, P. The Impact of Common Assumptions on Canopy Radiative Transfer Simulations: A Case Study in Citrus Orchards. J. Quant. Spectrosc. Radiat. Transf. 2009, 110, 1–21. [Google Scholar] [CrossRef]
- Widlowski, J.L.; Mio, C.; Disney, M.; Adams, J.; Andredakis, I.; Atzberger, C.; Brennan, J.; Busetto, L.; Chelle, M.; Ceccherini, G.; et al. The Fourth Phase of the Radiative Transfer Model Intercomparison (RAMI) Exercise: Actual Canopy Scenarios and Conformity Testing. Remote Sens. Environ. 2015, 169, 418–437. [Google Scholar] [CrossRef]
- Woodgate, W.; Disney, M.; Armston, J.D.; Jones, S.D.; Suarez, L.; Hill, M.J.; Wilkes, P.; Soto-Berelov, M.; Haywood, A.; Mellor, A. An Improved Theoretical Model of Canopy Gap Probability for Leaf Area Index Estimation in Woody Ecosystems. For. Ecol. Manag. 2015, 358, 303–320. [Google Scholar] [CrossRef]
- Åkerblom, M.; Raumonen, P.; Casella, E.; Disney, M.I.; Danson, F.M.; Gaulton, R.; Schofield, L.A.; Kaasalainen, M. Non-Intersecting Leaf Insertion Algorithm for Tree Structure Models. Interface Focus 2018, 8, 20170045. [Google Scholar] [CrossRef] [PubMed]
- Calders, K.; Origo, N.; Burt, A.; Disney, M.; Nightingale, J.; Raumonen, P.; Åkerblom, M.; Malhi, Y.; Lewis, P. Realistic Forest Stand Reconstruction from Terrestrial LiDAR for Radiative Transfer Modelling. Remote Sens. 2018, 10, 933. [Google Scholar] [CrossRef]
- Åkerblom, M.; Kaitaniemi, P. Terrestrial Laser Scanning: A New Standard of Forest Measuring and Modelling? Ann. Bot. 2021, 128, mcab111. [Google Scholar] [CrossRef]
- Disney, M. Terrestrial LiDAR: A Three-Dimensional Revolution in How We Look at Trees. New Phytol. 2019, 222, 1736–1741. [Google Scholar] [CrossRef] [PubMed]
- Côté, J.F.; Widlowski, J.L.; Fournier, R.A.; Verstraete, M.M. The Structural and Radiative Consistency of Three-Dimensional Tree Reconstructions from Terrestrial Lidar. Remote Sens. Environ. 2009, 113, 1067–1081. [Google Scholar] [CrossRef]
- Kükenbrink, D.; Schneider, F.D.; Schmid, B.; Gastellu-Etchegorry, J.P.; Schaepman, M.E.; Morsdorf, F. Modelling of Three-Dimensional, Diurnal Light Extinction in Two Contrasting Forests. Agric. For. Meteorol. 2021, 296, 108230. [Google Scholar] [CrossRef]
- Lukeš, P.; Neuwirthová, E.; Lhotáková, Z.; Janoutová, R.; Albrechtová, J. Upscaling Seasonal Phenological Course of Leaf Dorsiventral Reflectance in Radiative Transfer Model. Remote Sens. Environ. 2020, 246, 111862. [Google Scholar] [CrossRef]
- Kirby, K.J.; Bazely, D.R.; Goldberg, E.A.; Hall, J.E.; Isted, R.; Perry, S.C.; Thomas, R.C. Changes in the Tree and Shrub Layer of WythamWoods (Southern England) 1974-2012: Local and National Trends Compared. Forestry 2014, 87, cpu026. [Google Scholar] [CrossRef]
- Savill, P.; Perrins, C.; Kirby, K.; Fisher, N. Wytham Woods: Oxford’s Ecological Laboratory; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Calders, K.; Verbeeck, H.; Burt, A.; Origo, N.; Nightingale, J.; Malhi, Y.; Wilkes, P.; Raumonen, P.; Bunce, R.G.H.; Disney, M. Laser Scanning Reveals Potential Underestimation of Biomass Carbon in Temperate Forest. Ecol. Solut. Evid. 2022, 3, e12197. [Google Scholar] [CrossRef]
- The ECN Data Centre Site Information: Wytham. Available online: https://ecn.ac.uk/sites/site/terr/wytham (accessed on 1 April 2019).
- Burt, A.; Disney, M.; Calders, K. Extracting Individual Trees from Lidar Point Clouds Using Treeseg. Methods Ecol. Evol. 2019, 10, 438–445. [Google Scholar] [CrossRef]
- Calders, K.; Newnham, G.; Burt, A.; Murphy, S.; Raumonen, P.; Herold, M.; Culvenor, D.; Avitabile, V.; Disney, M.; Armston, J.; et al. Nondestructive Estimates of Above-Ground Biomass Using Terrestrial Laser Scanning. Methods Ecol. Evol. 2015, 6, 198–208. [Google Scholar] [CrossRef]
- Raumonen, P.; Kaasalainen, M.; Markku, Å.; Kaasalainen, S.; Kaartinen, H.; Vastaranta, M.; Holopainen, M.; Disney, M.; Lewis, P. Fast Automatic Precision Tree Models from Terrestrial Laser Scanner Data. Remote Sens. 2013, 5, 491–520. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A. Defining Leaf Area Index for Non-flat Leaves. Plant. Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Calders, K.; Origo, N.; Disney, M.; Nightingale, J.; Woodgate, W.; Armston, J.; Lewis, P. Variability and Bias in Active and Passive Ground-Based Measurements of Effective Plant, Wood and Leaf Area Index. Agric. For. Meteorol. 2018, 252, 231–240. [Google Scholar] [CrossRef]
- Calders, K.; Schenkels, T.; Bartholomeus, H.; Armston, J.; Verbesselt, J.; Herold, M. Monitoring Spring Phenology with High Temporal Resolution Terrestrial LiDAR Measurements. Agric. For. Meteorol. 2015, 203, 158–168. [Google Scholar] [CrossRef]
- Jupp, D.L.B.; Culvenor, D.S.; Lovell, J.L.; Newnham, G.J.; Strahler, A.H.; Woodcock, C.E. Estimating Forest LAI Profiles and Structural Parameters Using a Ground-Based Laser Called ’Echidna®. Tree Physiol. 2009, 29, 171–181. [Google Scholar] [CrossRef]
- Black, T.A.; Chen, J.-M.; Lee, X.; Sagar, R.M. Characteristics of Shortwave and Longwave Irradiances under a Douglas-Fir Forest Stand. Can. J. For. Res. 1991, 21, 1020–1028. [Google Scholar] [CrossRef]
- Ryu, Y.; Nilson, T.; Kobayashi, H.; Sonnentag, O.; Law, B.E.; Baldocchi, D.D. On the Correct Estimation of Effective Leaf Area Index: Does It Reveal Information on Clumping Effects? Agric. For. Meteorol. 2010, 150, 463–472. [Google Scholar] [CrossRef]
- Axelsson, P. DEM Generation from Laser Scanner Data Using Adaptive TIN Models. Int. Arch. Photogramm. Remote Sens. 2000, 23, 110–117. [Google Scholar]
- Gastellu-Etchegorry, J.P.; Demarez, V.; Pinel, V.; Zagolski, F. Modeling Radiative Transfer in Heterogeneous 3-D Vegetation Canopies. Remote Sens. Environ. 1996, 58, 131–156. [Google Scholar] [CrossRef]
- Pinty, B.; Gobron, N.; Widlowski, J.; Gerstl, S.A.W.; Verstraete, M.M.; Antunes, M.; Bacour, C.; Gascon, F.; Gastellu, J.; Goel, N. Radiation Transfer Model Intercomparison (RAMI) Exercise. J. Geophys. Res. Atmos. 2001, 106, 11937–11956. [Google Scholar] [CrossRef]
- Pinty, B.; Widlowski, J.L.; Taberner, M.; Gobron, N.; Verstraete, M.M.; Disney, M.; Gascon, F.; Gastellu, J.P.; Jiang, L.; Kuusk, A.; et al. Radiation Transfer Model Intercomparison (RAMI) Exercise: Results from the Second Phase. J. Geophys. Res. Atmos. 2004, 109, D6. [Google Scholar] [CrossRef]
- Widlowski, J.L.; Taberner, M.; Pinty, B.; Bruniquel-Pinel, V.; Disney, M.; Fernandes, R.; Gastellu-Etchegorry, J.P.; Gobron, N.; Kuusk, A.; Lavergne, T.; et al. Third Radiation Transfer Model Intercomparison (RAMI) Exercise: Documenting Progress in Canopy Reflectance Models. J. Geophys. Res. Atmos. 2007, 112, D9. [Google Scholar] [CrossRef]
- Liu, C.; Calders, K.; Origo, N.; Disney, M.; Meunier, F.; Woodgate, W.; Gastellu-Etchegorry, J.P.; Nightingale, J.; Honkavaara, E.; Hakala, T.; et al. Reconstructing the Digital Twin of Forests from a 3D Library: Quantifying Trade-Offs for Radiative Transfer Modeling. Remote Sens. Environ. 2023, 298, 113832. [Google Scholar] [CrossRef]
- Gastellu-Etchegorry, J.P. DART User’s Manua (5.7.9). Available online: https://dart.omp.eu/Public/documentation/contenu/documentation/DART_User_Manual.pdf (accessed on 11 May 2021).
- Wang, Y.; Lauret, N.; Gastellu-Etchegorry, J.P. DART Radiative Transfer Modelling for Sloping Landscapes. Remote Sens. Environ. 2020, 247, 111902. [Google Scholar] [CrossRef]
- Berk, A.; Bernstein, L.S.; Robertson, D.C. MODTRAN: A Moderate Resolution Model for LOWTRAN; Geophysics Laboratory: Lakewood, CA, USA, 1989; Volume 7. [Google Scholar]
- Grau, E.; Gastellu-Etchegorry, J.P. Radiative Transfer Modeling in the Earth-Atmosphere System with DART Model. Remote Sens. Environ. 2013, 139, 149–170. [Google Scholar] [CrossRef]
- Zemp, M.; Chao, Q.; Han Dolman, A.J.; Herold, M.; Krug, T.; Speich, S.; Suda, K.; Thorne, P.; Yu, W. GCOS 2022 Implementation Plan; World Meteorological Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Mariscal, M.J.; Martens, S.N.; Ustin, S.L.; Chen, J.; Weiss, S.B.; Roberts, D.A. Light-Transmission Profiles in an Old-Growth Forest Canopy: Simulations of Photosynthetically Active Radiation by Using Spatially Explicit Radiative Transfer Models. Ecosystems 2004, 7, 454–467. [Google Scholar] [CrossRef]
- Schaepman-Strub, G.; Schaepman, M.E.; Painter, T.H.; Dangel, S.; Martonchik, J.V. Reflectance Quantities in Optical Remote Sensing-Definitions and Case Studies. Remote Sens. Environ. 2006, 103, 27–42. [Google Scholar] [CrossRef]
- Zhang, J.; Khoshbakht, M.; Liu, J.; Gou, Z.; Xiong, J.; Jiang, M. A Clustering Review of Vegetation-Indicating Parameters in Urban Thermal Environment Studies towards Various Factors. J. Therm. Biol. 2022, 110, 103340. [Google Scholar] [CrossRef]
- Welles, J.M.; Norman, J.M. Instrument for Indirect Measurement of Canopy Architecture. Agron. J. 1991, 83, 818–825. [Google Scholar] [CrossRef]
- Iqbal, M. An Introduction to Solar Radiation; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 0323151817. [Google Scholar]
- Li, T.; Yang, Q. Advantages of Diffuse Light for Horticultural Production and Perspectives for Further Research. Front. Plant Sci. 2015, 6, 704. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L.; Pearcy, R.W. The Importance of Sunflecks for Forest Understory Plants. Bioscience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Pearcy, R.W. Photosynthetic Utilisation of Lightflecks by Understory Plants. Aust. J. Plant Physiol. 1988, 15, 223–238. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Chazdon, R.L.; Gross, L.J.; Mott, K.A. Photosynthetic Utilization of Sunflecks: A Temporally Patchy Resource on a Time Scale of Seconds to Minutes. In Exploitation of Environmental Heterogeneity by Plants; Academic Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Alton, P.B.; North, P.R.; Los, S.O. The Impact of Diffuse Sunlight on Canopy Light-Use Efficiency, Gross Photosynthetic Product and Net Ecosystem Exchange in Three Forest Biomes. Glob. Chang. Biol. 2007, 13, 776–787. [Google Scholar] [CrossRef]
- Denning, A.S.; Nicholls, M.; Prihodko, L.; Baker, I.; Vidale, P.L.; Davis, K.; Bakwin, P. Simulated Variations in Atmospheric CO2 over a Wisconsin Forest Using a Coupled Ecosystem-Atmosphere Model. Glob. Chang. Biol. 2003, 9, 1241–1250. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of Ecosystem Dynamics, Plant Geography and Terrestrial Carbon Cycling in the LPJ Dynamic Global Vegetation Model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Vicari, M.B.; Disney, M.; Wilkes, P.; Burt, A.; Calders, K.; Woodgate, W. Leaf and Wood Classification Framework for Terrestrial LiDAR Point Clouds. Methods Ecol. Evol. 2019, 10, 680–694. [Google Scholar] [CrossRef]
- Wang, D.; Momo Takoudjou, S.; Casella, E. LeWoS: A Universal Leaf-Wood Classification Method to Facilitate the 3D Modelling of Large Tropical Trees Using Terrestrial LiDAR. Methods Ecol. Evol. 2020, 11, 376–389. [Google Scholar] [CrossRef]
- Du, S.; Lindenbergh, R.; Ledoux, H.; Stoter, J.; Nan, L. AdTree: Accurate, Detailed, and Automatic Modelling of Laser-Scanned Trees. Remote Sens. 2019, 11, 2074. [Google Scholar] [CrossRef]
- Hakala, T.; Suomalainen, J.; Kaasalainen, S.; Chen, Y. Full Waveform Hyperspectral LiDAR for Terrestrial Laser Scanning. Opt. Express 2012, 20, 7119–7127. [Google Scholar] [CrossRef]
- Terryn, L.; Calders, K.; Bartholomeus, H.; Bartolo, R.E.; Brede, B.; D’hont, B.; Disney, M.; Herold, M.; Lau, A.; Shenkin, A.; et al. Quantifying Tropical Forest Structure through Terrestrial and UAV Laser Scanning Fusion in Australian Rainforests. Remote Sens. Environ. 2022, 271, 112912. [Google Scholar] [CrossRef]
- Schneider, F.D.; Kükenbrink, D.; Schaepman, M.E.; Schimel, D.S.; Morsdorf, F. Quantifying 3D Structure and Occlusion in Dense Tropical and Temperate Forests Using Close-Range LiDAR. Agric. For. Meteorol. 2019, 268, 249–257. [Google Scholar] [CrossRef]
- Chavana-Bryant, C.; Malhi, Y.; Anastasiou, A.; Enquist, B.J.; Cosio, E.G.; Keenan, T.F.; Gerard, F.F. Leaf Age Effects on the Spectral Predictability of Leaf Traits in Amazonian Canopy Trees. Sci. Total Environ. 2019, 666, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Ebengo, D.M.; De Boissieu, F.; Vincent, G.; Weber, C.; Féret, J.B. Simulating Imaging Spectroscopy in Tropical Forest with 3d Radiative Transfer Modeling. Remote Sens. 2021, 13, 2120. [Google Scholar] [CrossRef]
- Krishna Moorthy, S.M.; Raumonen, P.; Van den Bulcke, J.; Calders, K.; Verbeeck, H. Terrestrial Laser Scanning for Non-Destructive Estimates of Liana Stem Biomass. For. Ecol. Manag. 2020, 456, 117751. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hardware | 2 × 64-Core AMD EPYC 7773X (Milan-X @ 2.2 GHz) One Node × 18 Processors for Each RT Simulation | ||
|---|---|---|---|
| DART simulation settings | Radiative method | Image and sensor simulations | DART-Lux |
| Radiative budget simulations | DART-FT | ||
| Target sample density per pixel (DART-Lux) | 100 | ||
| Cell dimension (DART-FT) | 1 m | ||
| Transition: TOA <-> BOA | RT modeling with Earth-atmosphere coupling and diffuse transmittance | ||
| Year | 2015 | 2022 | |
|---|---|---|---|
| LAI | 3.8 | 4.7 | |
| Vegetationcover | Leaf cover | 85% | 85% |
| Wood cover | 48% | 43% | |
| Canopy cover (leaf + wood) | 88% | 87% | |
| FAPAR | Leaf | 0.69 | 0.73 |
| Wood | 0.14 | 0.10 | |
| Understorey | 0.06 | 0.06 | |
| Total | 0.89 | 0.90 | |
| RT Modeling Type | Spatial Resolution | Time Consumption (min) | RAM Consumption (GB) |
|---|---|---|---|
| Optical remote sensing observation | 10 m | 7 | 8 |
| 25 cm | 17 | 10.2 | |
| 1 cm | 4867.8 | 785.4 | |
| Canopy light distribution | 1 m | 216.8 | 41.8 |
| Light availability at explicit locations (PAR sensor simulation) | Each sensor | 24.1 | 3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Calders, K.; Origo, N.; Terryn, L.; Adams, J.; Gastellu-Etchegorry, J.-P.; Wang, Y.; Meunier, F.; Armston, J.; Disney, M.; et al. Bitemporal Radiative Transfer Modeling Using Bitemporal 3D-Explicit Forest Reconstruction from Terrestrial Laser Scanning. Remote Sens. 2024, 16, 3639. https://doi.org/10.3390/rs16193639
Liu C, Calders K, Origo N, Terryn L, Adams J, Gastellu-Etchegorry J-P, Wang Y, Meunier F, Armston J, Disney M, et al. Bitemporal Radiative Transfer Modeling Using Bitemporal 3D-Explicit Forest Reconstruction from Terrestrial Laser Scanning. Remote Sensing. 2024; 16(19):3639. https://doi.org/10.3390/rs16193639
Chicago/Turabian StyleLiu, Chang, Kim Calders, Niall Origo, Louise Terryn, Jennifer Adams, Jean-Philippe Gastellu-Etchegorry, Yingjie Wang, Félicien Meunier, John Armston, Mathias Disney, and et al. 2024. "Bitemporal Radiative Transfer Modeling Using Bitemporal 3D-Explicit Forest Reconstruction from Terrestrial Laser Scanning" Remote Sensing 16, no. 19: 3639. https://doi.org/10.3390/rs16193639
APA StyleLiu, C., Calders, K., Origo, N., Terryn, L., Adams, J., Gastellu-Etchegorry, J.-P., Wang, Y., Meunier, F., Armston, J., Disney, M., Woodgate, W., Nightingale, J., & Verbeeck, H. (2024). Bitemporal Radiative Transfer Modeling Using Bitemporal 3D-Explicit Forest Reconstruction from Terrestrial Laser Scanning. Remote Sensing, 16(19), 3639. https://doi.org/10.3390/rs16193639