
Multitemporal Hyperspectral Characterization of Wheat Infested by Wheat Stem Sawfly, Cephus cinctus Norton


Abstract
1. Introduction
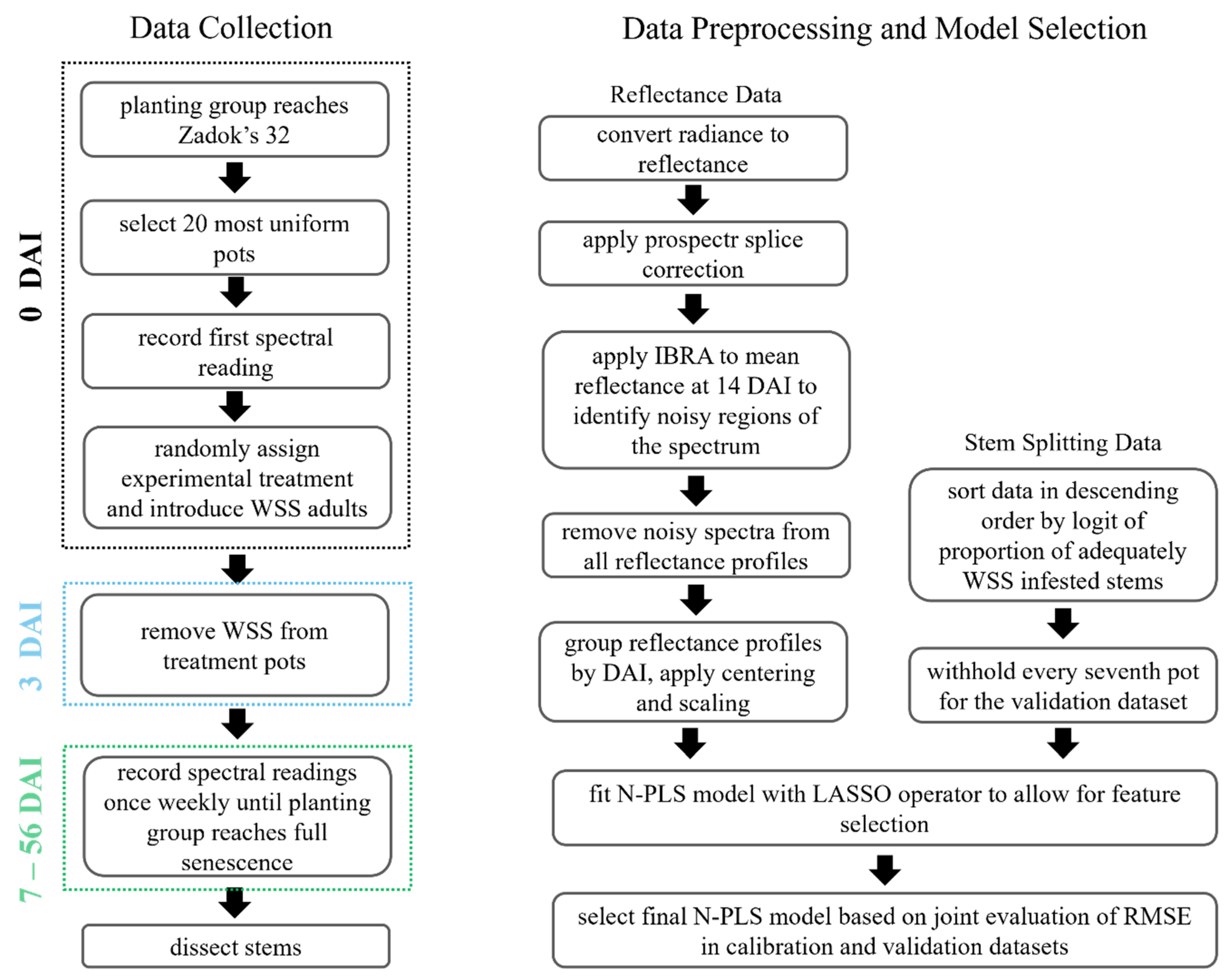
2. Materials and Methods
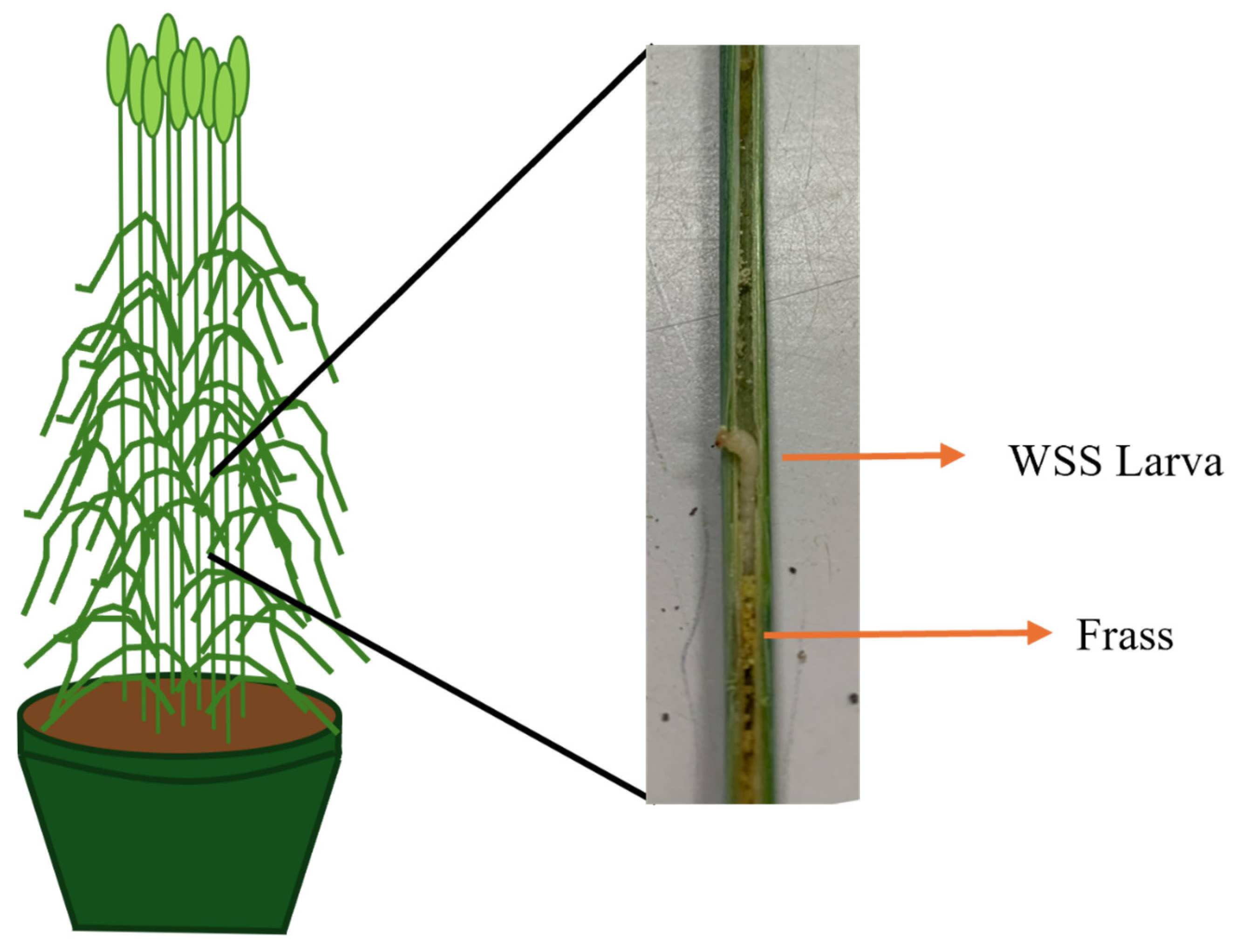
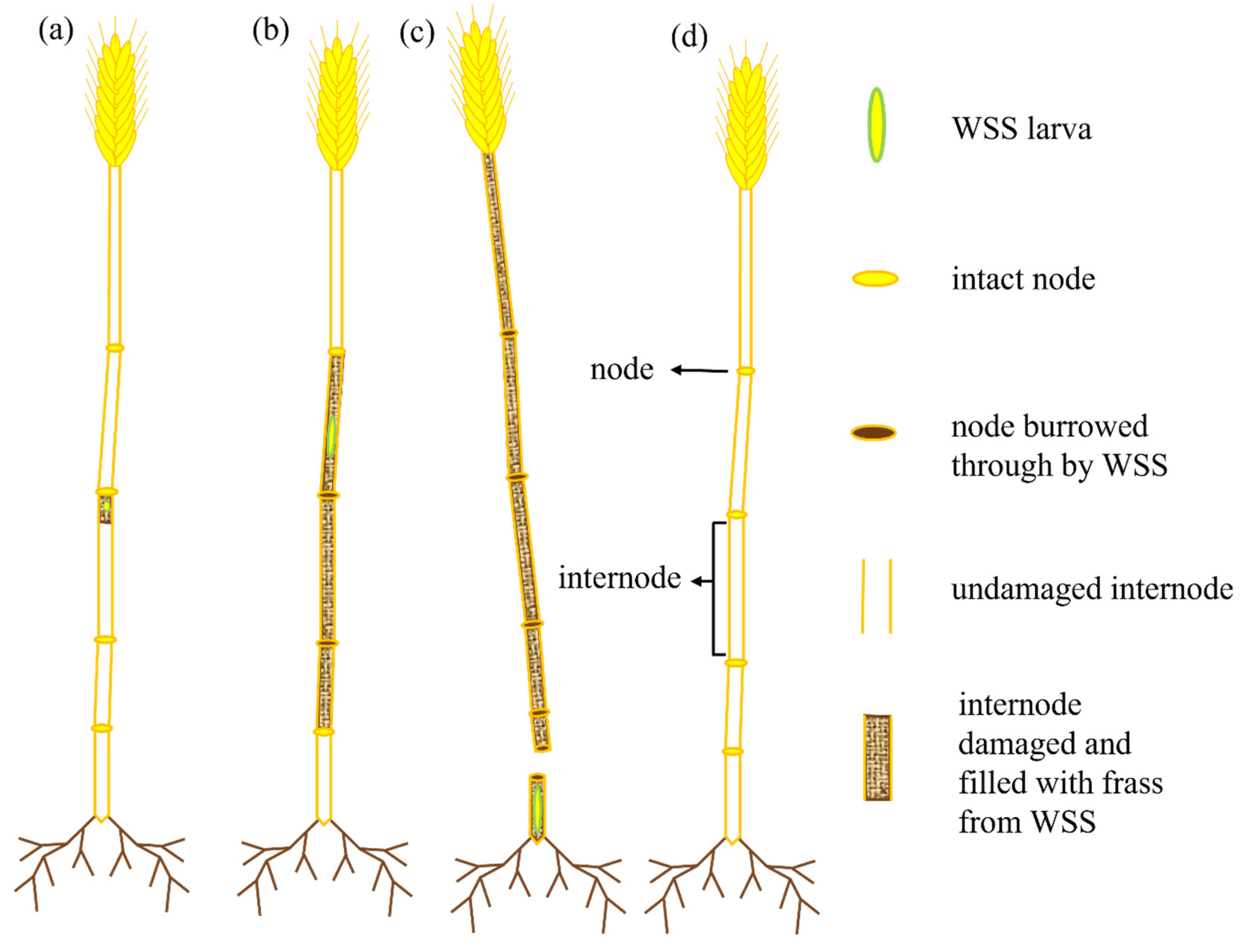
2.1. Plant Cultivation
2.2. Spectral Reflectance Sampling
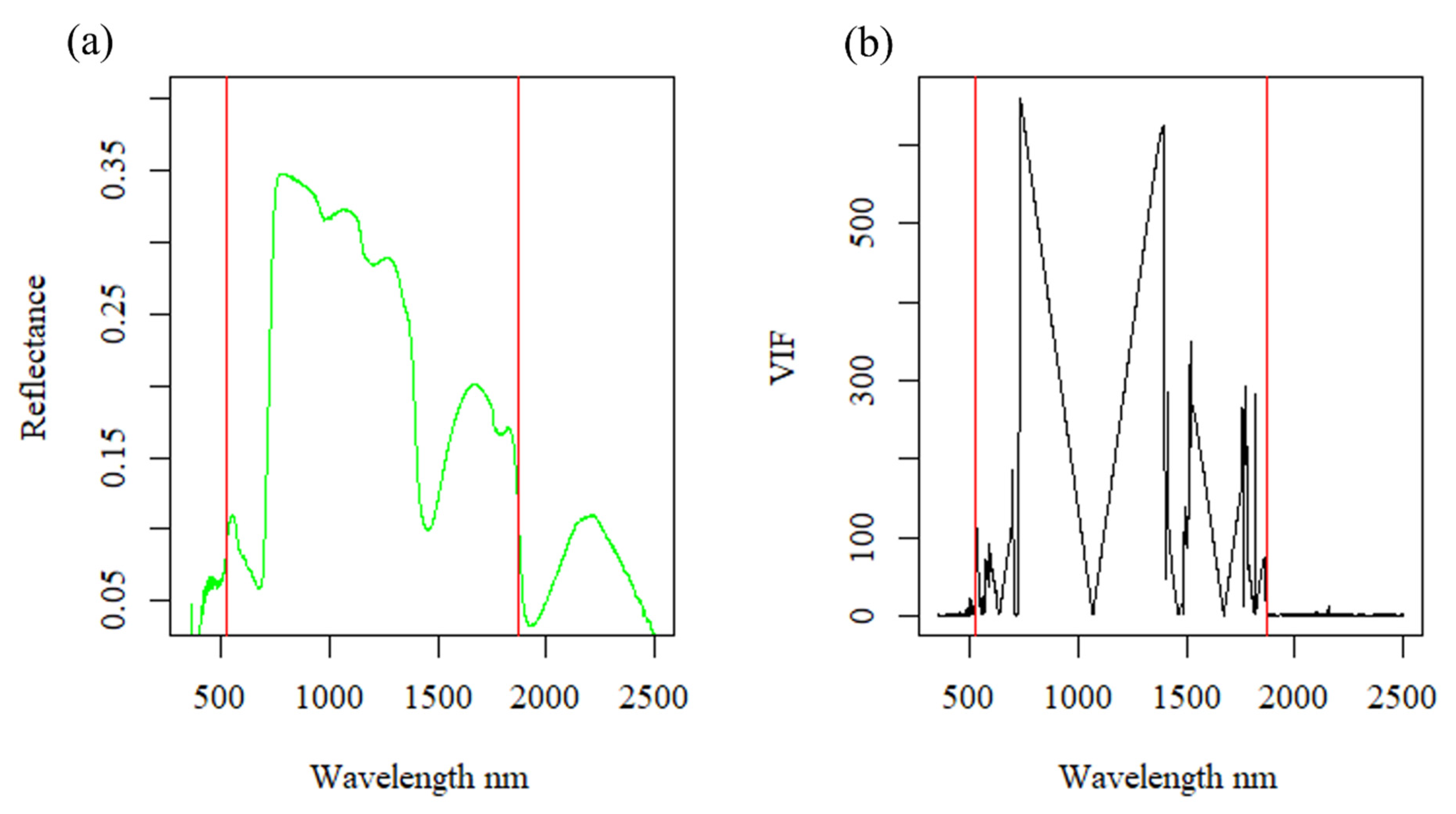
2.3. Data Preprocessing
2.4. Sparse Multiway Partial Least Squares Regression Modeling
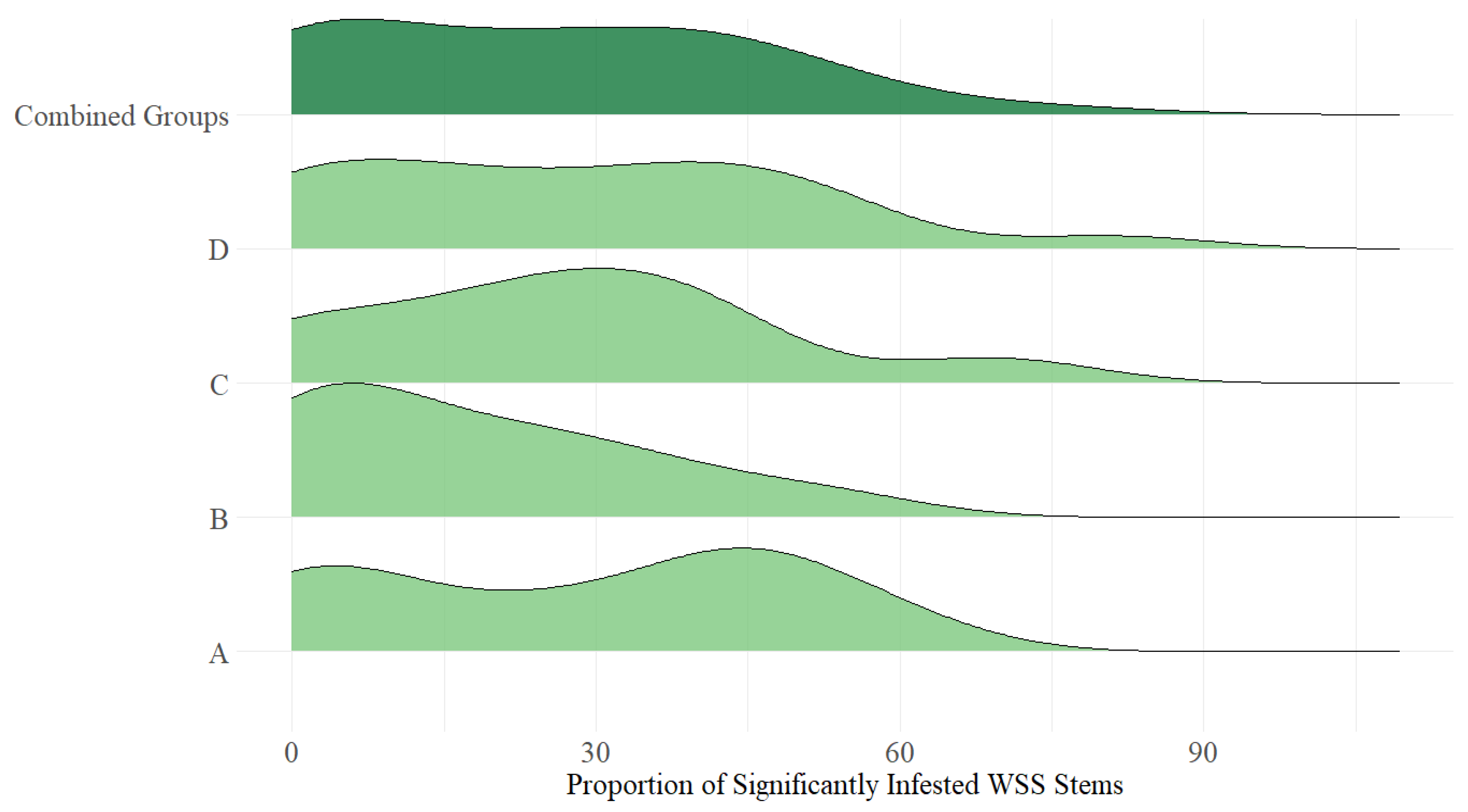
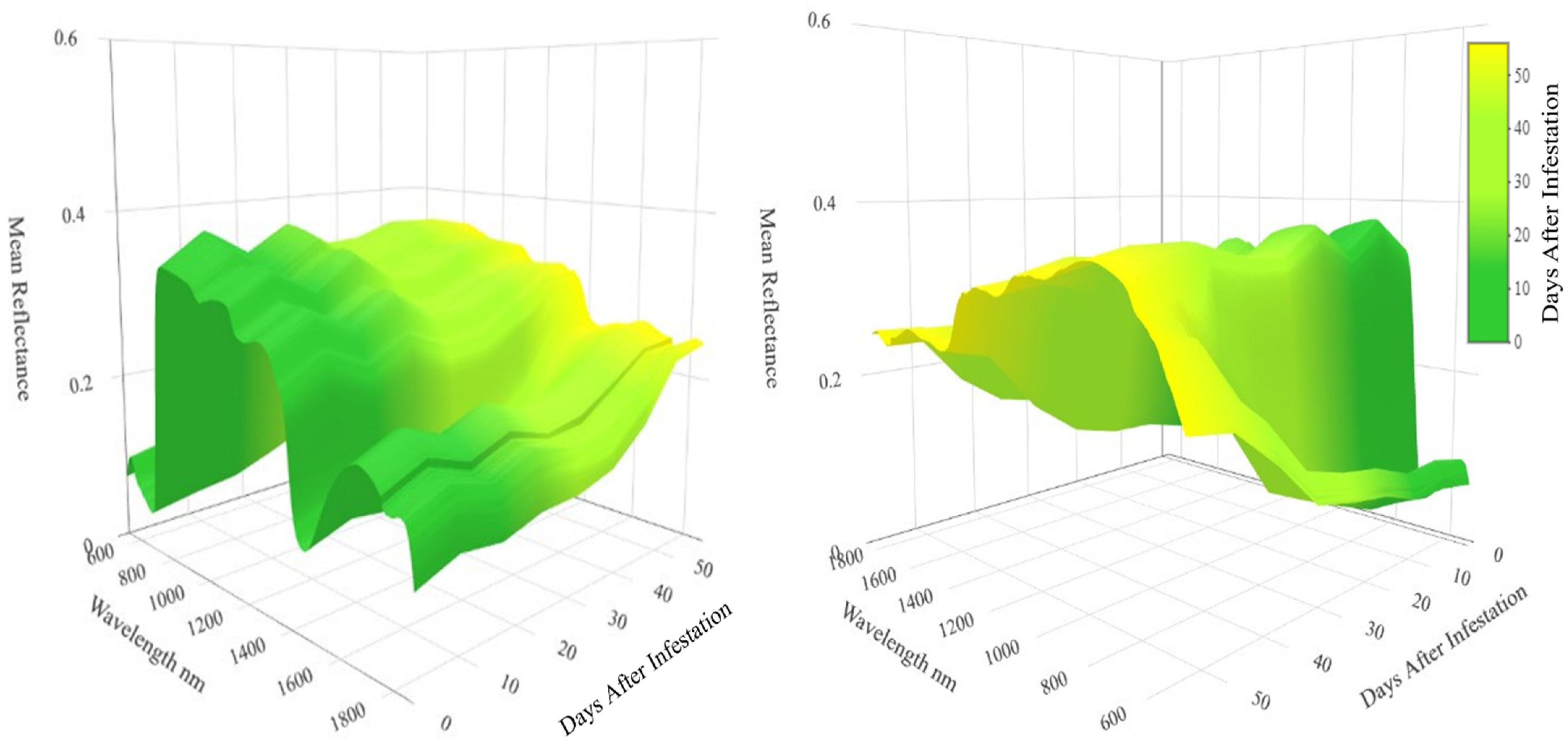
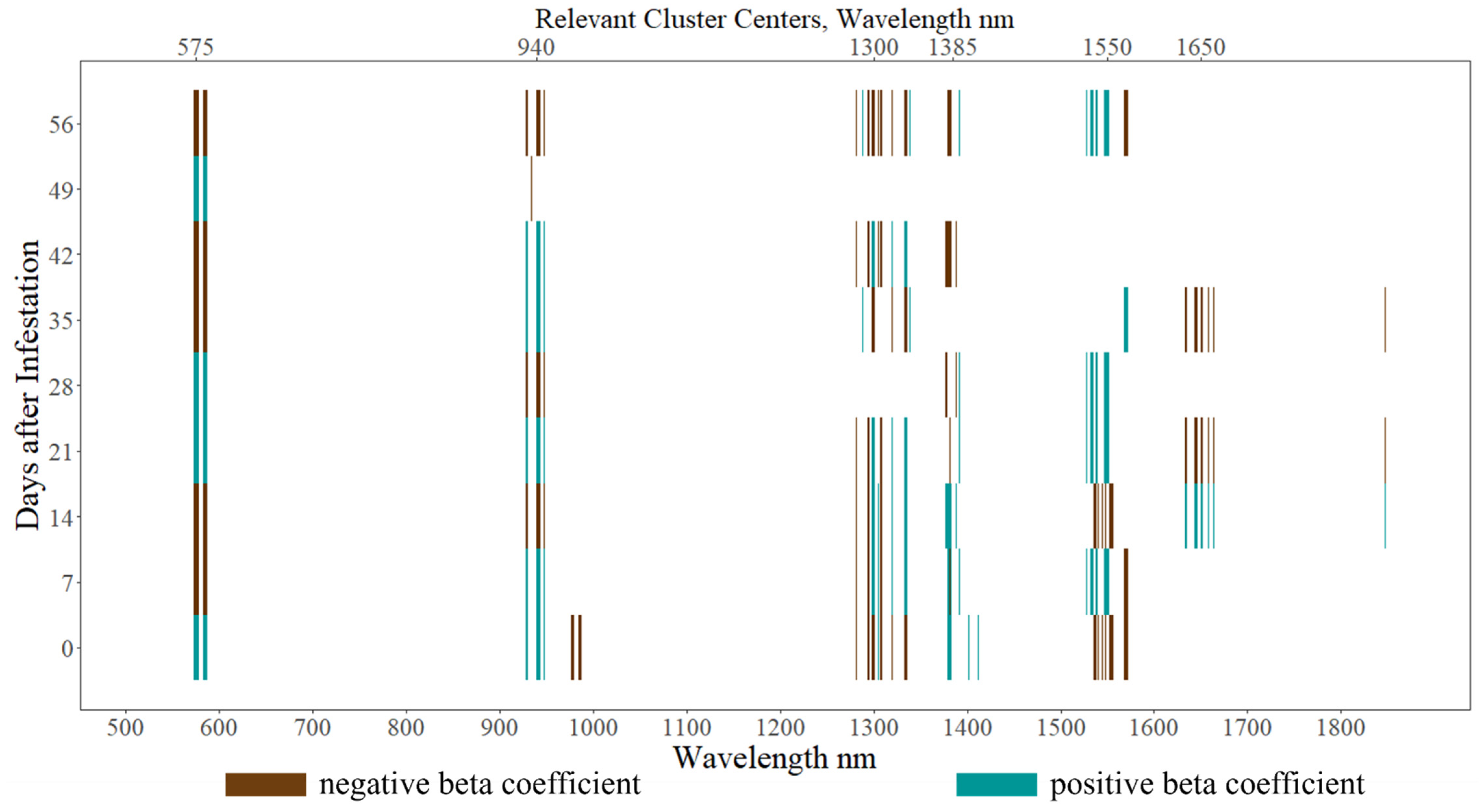
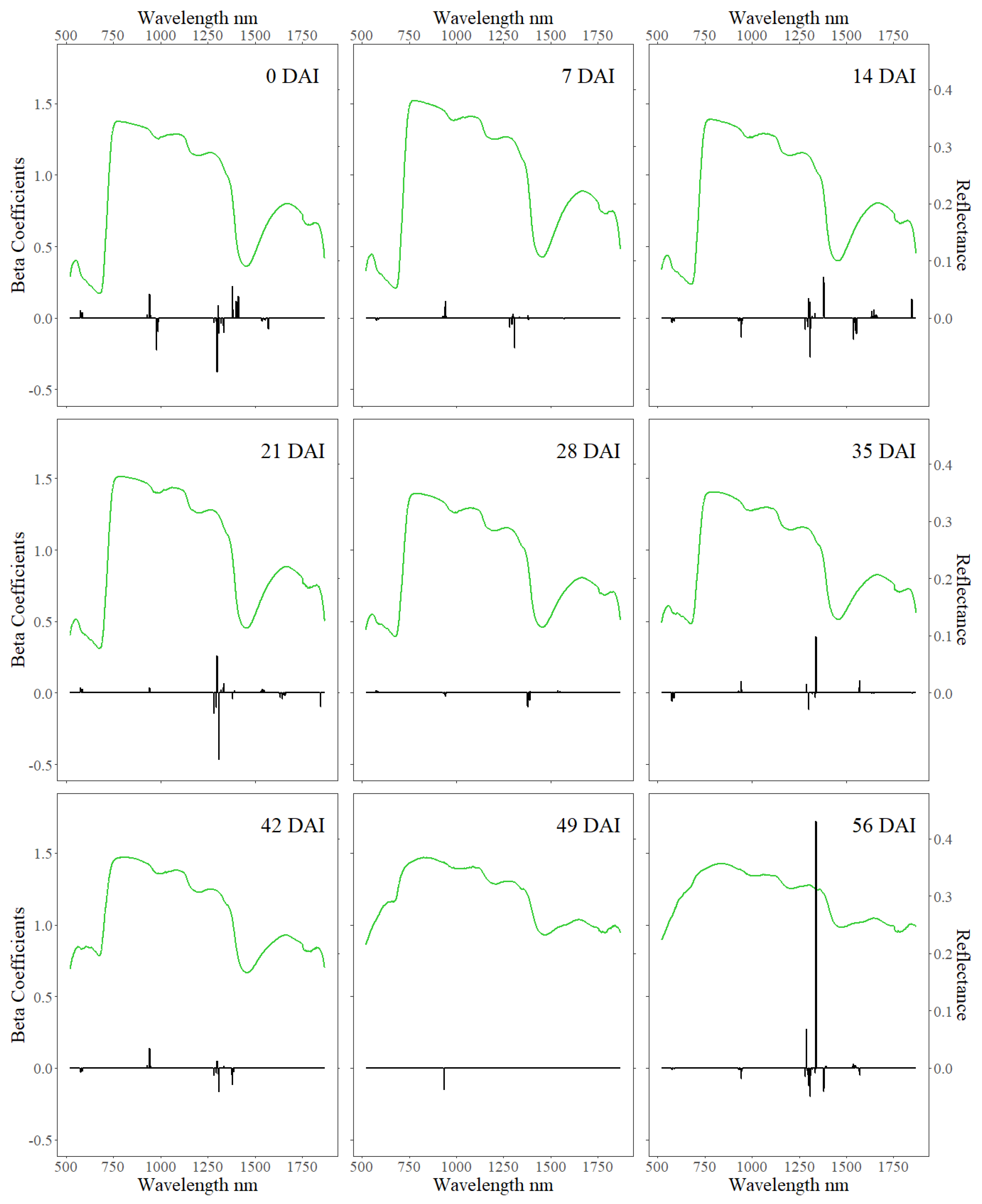
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Beres, B.L.; Dosdall, L.M.; Weaver, D.K.; Cárcamo, H.A.; Spaner, D.M. Biology and Integrated Management of Wheat Stem Sawfly and the Need for Continuing Research. Can. Entomol. 2011, 143, 105–125. [Google Scholar] [CrossRef]
- Criddle, N. The Western-Stem Sawfly and its Control. Dom. Can. Dept. Ag. 1922, 6, 1–8. [Google Scholar]
- Knodel, J.; Shanower, T.; Beauzay, P. Integrated Pest Management of Wheat Stem Sawfly. 2010. Available online: https://library.ndsu.edu/ir/handle/10365/10435 (accessed on 18 February 2024).
- Cockrell, D.M.; Randolph, T.; Peirce, E.; Peairs, F.B. Survey of Wheat Stem Sawfly (Hymenoptera: Cephidae) Infesting Wheat in Eastern Colorado. J. Econ. Entomol. 2021, 114, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- McCullough, C.T.; Hein, G.L.; Bradshaw, J.D. Phenology and Dispersal of the Wheat Stem Sawfly (Hymenoptera: Cephidae) into Winter Wheat Fields in Nebraska. J. Econ. Entomol. 2020, 113, 1831–1838. [Google Scholar] [CrossRef]
- Holmes, N.D. Food Relations of the Wheat Stem Sawfly, Cephus cinctus Nort. (Hymenoptera: Cephidae). Can. Entomol. 1954, 86, 159–167. [Google Scholar] [CrossRef]
- Morrill, W.L.; Gabor, J.W.; Kushnak, G.D. Wheat Stem Sawfly (Hymenoptera: Cephidae): Damage and Detection. J. Econ. Entomol. 1992, 85, 2413–2417. [Google Scholar] [CrossRef]
- Macedo, T.B.; Weaver, D.K.; Peterson, R.K.D. Characterization of the Impact of Wheat Stem Sawfly, Cephus cinctus Norton, on Pigment Composition and Photosystem II Photochemistry of Wheat Heads. Environ. Entomol. 2006, 35, 1115–1120. [Google Scholar] [CrossRef]
- Delaney, K.J.; Weaver, D.K.; Peterson, R.K.D. Photosynthesis and Yield Reductions from Wheat Stem Sawfly (Hymenoptera: Cephidae): Interactions with Wheat Solidness, Water Stress, and Phosphorus Deficiency. J. Econ. Entomol. 2010, 103, 516–524. [Google Scholar] [CrossRef]
- Seamans, H.L.; Manson, G.F.; Farstad, C.W. The Effect of the Wheat Stem Sawfly (Cephus cinctus Nort.) on the Heads and Grain of Infested Stems. Entomol. Soc. Ontario. 1945, 75, 10–15. [Google Scholar]
- Macedo, T.B.; Weaver, D.K.; Peterson, R.K.D. Photosynthesis in wheat at the grain filling stage is altered by larval wheat stem sawfly (Hymenoptera: Cephidae) injury and reduced water availability. J. Entomol. Sci. 2007, 42, 228–238. [Google Scholar] [CrossRef]
- Rodriguez, D.; Fitzgerald, G.J.; Belford, R.; Christensen, L.K. Detection of Nitrogen Deficiency in Wheat from Spectral Reflectance Indices and Basic Crop Eco-Physiological Concepts. Aust. J. Agric. Res. 2006, 57, 781–789. [Google Scholar] [CrossRef]
- Tilling, A.K.; O’Leary, G.J.; Ferwerda, J.G.; Jones, S.D.; Fitzgerald, G.J.; Rodriguez, D.; Belford, R. Remote Sensing of Nitrogen and Water Stress in Wheat. Field Crops Res. 2007, 104, 77–85. [Google Scholar] [CrossRef]
- Bhattarai, G.P.; Schmid, R.B.; McCornack, B.P. Remote Sensing Data to Detect Hessian Fly Infestation in Commercial Wheat Fields. Sci. Rep. 2019, 9, 6109. [Google Scholar] [CrossRef] [PubMed]
- Mirik, M.; Jones, D.C.; Price, J.A.; Workneh, F.; Ansley, R.J.; Rush, C.M. Satellite Remote Sensing of Wheat Infected by Wheat Streak Mosaic Virus. Plant Dis. 2011, 95, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Bauriegel, E.; Giebel, A.; Geyer, M.; Schmidt, U.; Herppich, W.B. Early Detection of Fusarium Infection in Wheat Using Hyper-Spectral Imaging. Comput. Electron. Agric. 2011, 75, 304–312. [Google Scholar] [CrossRef]
- Ma, H.; Huang, W.; Dong, Y.; Liu, L.; Guo, A. Using UAV-Based Hyperspectral Imagery to Detect Winter Wheat Fusarium Head Blight. Remote Sens. 2021, 13, 3024. [Google Scholar] [CrossRef]
- Collins, W. Remote Sensing of Crop Type and Maturity. Photogramm. Eng. Remote Sens. 1978, 44, 43–55. [Google Scholar]
- Lestina, J.; Cook, M.; Kumar, S.; Morisette, J.; Ode, P.J.; Peairs, F. MODIS Imagery Improves Pest Risk Assessment: A Case Study of Wheat Stem Sawfly (Cephus cinctus, Hymenoptera: Cephidae) in Colorado, USA. Environ. Entomol. 2016, 45, 1343–1351. [Google Scholar] [CrossRef]
- Nansen, C.; Macedo, T.; Swanson, R.; Weaver, D.K. Use of Spatial Structure Analysis of Hyperspectral Data Cubes for Detection of Insect-Induced Stress in Wheat Plants. Int. J. Remote Sens. 2009, 30, 2447–2464. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and Physiological Basis for the Reflectance of Visible and Near-Infrared Radiation from Vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectral Properties of Plants. Appl. Opt. 1965, 4, 11–20. [Google Scholar] [CrossRef]
- Tucker, C.J. Remote Sensing of Leaf Water Content in the Near Infrared. Remote Sens. Environ. 1980, 10, 23–32. [Google Scholar] [CrossRef]
- Seelig, H.D.; Hoehn, A.; Stodieck, L.S.; Klaus, D.M.; Adams Iii, W.W.; Emery, W.J. The Assessment of Leaf Water Content Using Leaf Reflectance Ratios in the Visible, Near-, and Short-Wave-Infrared. Int. J. Remote Sens. 2008, 29, 3701–3713. [Google Scholar] [CrossRef]
- Thomas, S.; Kuska, M.T.; Bohnenkamp, D.; Brugger, A.; Alisaac, E.; Wahabzada, M.; Mahlein, A.K. Benefits of Hyperspectral Imaging for Plant Disease Detection and Plant Protection: A Technical Perspective. J. Plant Dis. Prot. 2018, 125, 5–20. [Google Scholar] [CrossRef]
- Hughes, F.G. On the Mean Accuracy of Statistical Pattern Recognizers. IEEE 1986, 14, 55–63. [Google Scholar] [CrossRef]
- Georganos, S.; Grippa, T.; Vanhuysse, S.; Lennert, M.; Shimoni, M.; Kalogirou, S.; Wolff, E. Less is More: Optimizing Classification Performance Through Feature Selection in a Very-High-Resolution Remote Sensing Object-Based Urban Application. GISci. Remote Sens. 2018, 55, 221–242. [Google Scholar] [CrossRef]
- Romero, A.; Gatta, C.; Camps-Valls, G. Unsupervised Deep Feature Extraction for Remote Sensing Image Classification. IEEE Trans. Geosci. Remote Sens. 2015, 54, 1349–1362. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant Spectral Diversity Integrates Functional and Phylogenetic Components of Biodiversity and Predicts Ecosystem Function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef]
- Yu, H.; Guo, L.; Kharbach, M.; Han, W. Multi-Way Analysis Coupled with Near-Infrared Spectroscopy in Food Industry: Models and Applications. Foods 2021, 10, 802. [Google Scholar] [CrossRef]
- Lopez-Fornieles, E.; Brunel, G.; Rancon, F.; Gaci, B.; Metz, M.; Devaux, N.; Taylor, J.; Tisseyre, B.; Roger, J.M. Potential of Multiway PLS (N-PLS) Regression Method to Analyse Time-Series of Multispectral Images: A Case Study in Agriculture. Remote Sens. 2022, 14, 216. [Google Scholar] [CrossRef]
- Bro, R. Multiway Calibration. Multilinear PLS. J. Chemom. 1996, 10, 47–61. [Google Scholar] [CrossRef]
- Wold, S.; Geladi, P.; Esbensen, K.; Öhman, J. Multi-Way Principal Components-and PLS-Analysis. J. Chemom. 1987, 1, 41–56. [Google Scholar] [CrossRef]
- Varella, A.C.; Weaver, D.K.; Blake, N.K.; Hofland, M.L.; Heo, H.Y.; Cook, J.P.; Lamb, P.F.; Jordan, K.W.; Akhunov, E.; Chao, S.; et al. Analysis of Recombinant Inbred Line Populations Derived from Wheat Landraces to Identify New Genes for Wheat Stem Sawfly Resistance. Theor. Appl. Genet. 2019, 132, 2195–2207. [Google Scholar] [CrossRef]
- United States Department of Agriculture, N.A.S.S. Montana 2023 Wheat Varieties. 2023. Available online: https://agr.mt.gov/News/USDA-NASS-Wheat-and-Barley-Survey-Indicates-Top-2023-Varietals (accessed on 16 March 2024).
- Lee, H.; Cho, S.; Lim, J.; Lee, A.; Kim, G.; Song, D.J.; Chun, S.W.; Kim, M.J.; Mo, C. Performance Comparison of Tungsten-Halogen Light and Phosphor-Converted NIR LED in Soluble Solid Content Estimation of Apple. Sensors 2023, 23, 1961. [Google Scholar] [CrossRef]
- Danner, M.; Locherer, M.; Hank, T.; Richter, K. Spectral Sampling with the ASD FieldSpec 4—Theory, Management, Problems, Interpretation. In EnMAP Field Guides Technical Report, GFZ Data Services; EnMAP: Potsdam, Germany, 2015. [Google Scholar] [CrossRef]
- R Core Team. R Language Definition; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Roudier, P.; Lalibert’, E. Asdreader. 2017. Available online: https://cran.r-project.org/web/packages/asdreader/index.html (accessed on 23 January 2024).
- Stevens, A.; Ramirez-Lopez, L. Prospectr. 2024. Available online: https://cran.r-project.org/web/packages/prospectr/index.html (accessed on 23 January 2024).
- Hatchell, D.C. ASD Technical Guide 3rd Ed. Section 0–1 Section 0 Technical Guide. 1999. Available online: https://www.yumpu.com/en/document/view/11346134/analytical-spectral-devices-inc-asd-technical-guide (accessed on 23 January 2024).
- Morales, G.; Sheppard, J.W.; Logan, R.D.; Shaw, J.A. Hyperspectral Dimensionality Reduction Based on Inter-Band Redundancy Analysis and Greedy Spectral Selection. Remote Sens. 2021, 13, 3649. [Google Scholar] [CrossRef]
- Thoms, L.J.; Girwidz, R. Training and Assessment of Experimental Competencies from a Distance: Optical Spectrometry via the Internet. Il Nuovo C. C 2015, 38, 59–63. [Google Scholar] [CrossRef]
- Hervas, D. sNPLS. 2022. Available online: https://cran.r-project.org/web/packages/sNPLS/index.html (accessed on 1 May 2024).
- Hervás, D.; Prats-Montalbán, J.M.; Lahoz, A.; Ferrer, A. Sparse N-way partial least squares with R package sNPLS. Chemom. Intell. Lab. Syst. 2018, 179, 54–63. [Google Scholar] [CrossRef]
- Tibshirani, R. Regression Shrinkage and Selection via the Lasso. J. R. Stat. Soc. Ser. B 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Goodarzi, M.; Freitas, M.P. On the use of PLS and N-PLS in MIA-QSAR: Azole Antifungals. Intell. Lab. Syst. 2009, 96, 59–62. [Google Scholar] [CrossRef]
- Bidinger, F.; Musgrave, R.B.; Fischer, R.A. Contribution of Stored Pre-Anthesis Assimilate to Grain Yield in Wheat and Barley. Nature 1977, 270, 431–433. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Rock, B.N. Detection of Changes in Leaf Water Content Using Near-and Middle-Infrared Reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar] [CrossRef]
- Zhang, W.; Li, X.; Zhao, L. Discovering the Representative Subset with Low Redundancy for Hyperspectral Feature Selection. Remote Sens. 2019, 11, 1341. [Google Scholar] [CrossRef]
- Woolley, J.T. Reflectance and Transmittance of Light by Leaves. Plant Physiol. 1971, 47, 656–662. [Google Scholar] [CrossRef]
- Macedo, T.B.; Peterson, R.K.; Weaver, D.K.; Morrill, W.L. Wheat Stem Sawfly, Cephus cinctus Norton, Impact on Wheat Primary Metabolism: An Ecophysiological Approach. Environ. Entomol. 2005, 34, 719–726. [Google Scholar] [CrossRef]
- Liu, J.; Van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 619987. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Serrano, L.; Penuelas, J.; Ustin, S.L. Remote Sensing of Nitrogen and Lignin in Mediterranean Vegetation from AVIRIS Data: Decomposing Biochemical from Structural Signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water from Space. Remote Sens Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Buteler, M.; Weaver, D.K. Host Selection by the Wheat Stem Sawfly in Winter Wheat and the Role of Semiochemicals Mediating Oviposition Preference. Èntomol. Exp. Appl. 2012, 143, 138–147. [Google Scholar] [CrossRef]
- Xiao, G.; Zhang, X.; Niu, Q.; Li, X.; Li, X.; Zhong, L.; Huang, J. Winter Wheat Yield Estimation at the Field Scale using Sentinel-2 Data and Deep Learning. Comput. Electron. Agric. 2024, 216, 108555. [Google Scholar] [CrossRef]
- Ma, H.; Jing, Y.; Huang, W.; Shi, Y.; Dong, Y.; Zhang, J.; Liu, L. Integrating Early Growth Information to Monitor Winter Wheat Powdery Mildew Using Multi-Temporal Landsat-8 Imagery. Sensors 2018, 18, 3290. [Google Scholar] [CrossRef] [PubMed]
- Crema, A.; Boschetti, M.; Nutini, F.; Cillis, D.; Casa, R. Influence of Soil Properties on Maize and Wheat Nitrogen Status Assessment from Sentinel-2 Data. Remote Sens. 2020, 12, 2175. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavelength | Days after Infestation | Beta Coefficient |
|---|---|---|
| 1339 | 56 | 1.72 |
| 1308 | 21 | −0.46 |
| 1339 | 35 | 0.39 |
| 1300 | 0 | −0.37 |
| 1380 | 14 | 0.28 |
| 1308 | 14 | −0.27 |
| 1289 | 56 | 0.27 |
| 1300 | 21 | 0.26 |
| 1381 | 14 | 0.25 |
| 1301 | 0 | −0.24 |
| 979 | 0 | −0.22 |
| 1380 | 0 | 0.22 |
| 1308 | 7 | −0.21 |
| 1308 | 56 | −0.19 |
| 1381 | 0 | 0.19 |
| 978 | 0 | −0.17 |
| 1380 | 56 | −0.16 |
| 1301 | 21 | 0.16 |
| 942 | 0 | 0.16 |
| 1308 | 42 | −0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermatinger, L.S.; Powell, S.L.; Peterson, R.K.D.; Weaver, D.K. Multitemporal Hyperspectral Characterization of Wheat Infested by Wheat Stem Sawfly, Cephus cinctus Norton. Remote Sens. 2024, 16, 3505. https://doi.org/10.3390/rs16183505
Ermatinger LS, Powell SL, Peterson RKD, Weaver DK. Multitemporal Hyperspectral Characterization of Wheat Infested by Wheat Stem Sawfly, Cephus cinctus Norton. Remote Sensing. 2024; 16(18):3505. https://doi.org/10.3390/rs16183505
Chicago/Turabian StyleErmatinger, Lochlin S., Scott L. Powell, Robert K. D. Peterson, and David K. Weaver. 2024. "Multitemporal Hyperspectral Characterization of Wheat Infested by Wheat Stem Sawfly, Cephus cinctus Norton" Remote Sensing 16, no. 18: 3505. https://doi.org/10.3390/rs16183505
APA StyleErmatinger, L. S., Powell, S. L., Peterson, R. K. D., & Weaver, D. K. (2024). Multitemporal Hyperspectral Characterization of Wheat Infested by Wheat Stem Sawfly, Cephus cinctus Norton. Remote Sensing, 16(18), 3505. https://doi.org/10.3390/rs16183505