1. Introduction
Indigenous communities throughout California, USA have long cultural histories of practicing fire stewardship in diverse ecosystems [
1]. This practice of fire stewardship (often referred to as cultural burning) varies across the state and commonly aims to restore culturally important ecosystems, improve food and basketry materials, open waterways, and reduce the risk and severity of wildfire, among other objectives [
2,
3,
4,
5]. The significance of cultural burning has gained attention as wildfires have become increasingly more severe and frequent across California in just the past decade, a deadly pattern that is expected to continue [
6]. This upward trend in wildfire severity and frequency is caused by a confluence of factors including anthropogenic climate change and sociopolitical changes instituted during the European colonization of California in the 1700s that prohibited the practice of cultural burning and decimated Indigenous nations [
1,
6]. Nonetheless, many Indigenous communities have persisted in preserving the knowledge and practice of cultural fire; however, many continue to meet significant barriers to cultural burning such as difficulties obtaining the required permits, equipment, and certifications [
1,
7,
8]. In the absence of this deliberate application of frequent, low-severity fire, many of California’s disturbance-dependent ecosystems have declined, and fuel loads have accumulated to dangerous levels [
1,
5,
9].
As a result of the advocacy of Indigenous communities and increasing evidence of the ecological importance of fire, the California state legislature has invested millions of dollars into the restoration of cultural burning and prescribed fire in an effort to reduce the occurrence and severity of wildfires [
10]. Prescribed fire, also referred to as controlled burning, is the practice of lighting low-severity fires to reduce fuel loads and support the growth of fire-adapted plants. Cultural burning practices, which vary between Indigenous communities, tend to be distinct from typical state-led controlled burning practices in a number of ways. In addition to the variety of ecological and cultural purposes described above, cultural burning is a spiritual practice deeply rooted in generations of place-based knowledge [
8]. While there are other important differences in the purposes and practices of controlled burning and cultural burning that we do not discuss in depth here, we use the term “intentional burn” to refer to the broader category of deliberately set, low-severity fires that encompasses both of these categories.
Given the size of this investment and the sea change in fire management policy, research that analyzes the impact of these intentional burns is critical. In particular, post-fire studies help to determine whether these intentional burns are meeting their stated goals, which may include a reduction in certain wildfire fuels, the recovery of important native plants, or hydrological restoration. While field-based surveys provide valuable and direct observations of these effects, the extent of fire restoration in California demands a suite of methods that are more scalable, affordable, and efficient. Remote sensing—or more specifically, the analysis of satellite imagery—enables the study of ecosystem health and function over large areas and long periods and is therefore critical in measuring how fire impacts different ecosystems.
Many remote sensing analyses of fire impacts on vegetation have focused primarily on wildfires (rather than intentional burns) and have only evaluated a limited number of points in time. Common methodologies use a pre- and post-fire image to calculate wildfire severity, or use single images captured many years after the wildfire to measure the extent of vegetation recovery [
11]. Many of these methods are specifically tailored to conifer-dominated forest ecosystems, and therefore best capture the mortality of high-biomass vegetation or recoveries that endure many years [
11,
12]. There are fewer remote sensing methods developed for the study of other types of fire—in particular, lower severity fires such as intentional burns—and for the study of fire in non-forest ecosystems such as grasslands and shrublands [
12].
The study of how different types of fire impact grasslands is of particular importance. Grasslands are simultaneously one of California’s most biodiverse and most threatened ecosystems and are home to many plant and animal species that are significant to Indigenous communities across the state [
13,
14]. California’s grassland ecosystems are dependent on frequent, low-severity fires for renewal and to prevent the encroachment of shrub and tree species [
15,
16]. Specifically, low-severity fires increase soil nutrient availability and stimulate germination and seed production in many native perennial grasses [
16]. Conversely, high-severity fires (such as wildfires) and the complete suppression of fire have both been shown to reduce nutrient availability and may favor non-native annual grasses over native perennial grasses [
16]. However, because grasses regrow much faster and exhibit greater seasonal variation than trees, annually derived metrics of vegetation recovery (which work well for tracking the impacts of fires on forests) may not capture meaningful metrics of recovery in grasslands.
Many remote sensing studies use annually derived vegetation index (VI) values (e.g., an annual mean or median) to track the disturbance recovery over many years [
11]. For fast-changing grasslands, these annually integrated VI values may fail to capture important ecological changes that could be captured by measures of intra-annual VI variation. Given the importance of seasonal variation and recent increases in computational capacity, remote sensing studies of disturbance are increasingly calculating VI-derived phenological curves from dense time series of satellite imagery. These VI-derived phenological curves yield detailed metrics that describe these intra-annual changes in growth and senescence [
17]. Examples of these phenological metrics include the timing of the start and end of the growing season, the rate of growth or senescence, and the amplitude of the annual growth curve, among many others [
17]. Computation of these phenological metrics typically relies on imagery with high temporal resolution such as imagery from the MODIS satellites [
18,
19]. Though useful over large areas, MODIS imagery has a 1 km spatial resolution, which makes it infeasible to reliably study phenological phenomena occurring at smaller scales, and more generally introduces greater uncertainty in the phenological interpretation of spatially heterogeneous ecosystems [
20]. From 2019 to 2022, 91.44% of the intentional burns in California were smaller than 1 km
2 and have therefore not been the subject of many phenological studies, despite the post-fire recovery of these ecosystems having immense ecological and societal importance [
21]. Therefore, in order to study phenological changes caused by these smaller intentional burns, we need to develop methodologies that utilize imagery with higher spatial resolution and adequate temporal resolution.
To address this, our study used Sentinel-2 satellite imagery, which has a considerably higher spatial resolution than MODIS imagery (10 m to 60 m) and a temporal resolution of 5 days. Combined with its appropriate spectral sensitivity, Sentinel-2 imagery enables the phenological analysis of smaller areas, and is therefore more applicable to the study of intentional burning. However, its lower temporal resolution means that Sentinel-2-derived metrics of phenological timing (such as the timing of the start, peak, and end of season) are often delayed [
22,
23]. Therefore, we propose a methodology that focuses instead on the shape of the VI-derived phenological curve. Specifically, we analyzed five years of Sentinel-2 imagery and summarize the annual trends using the VI mean, fifth percentile, and amplitude values over time and space.
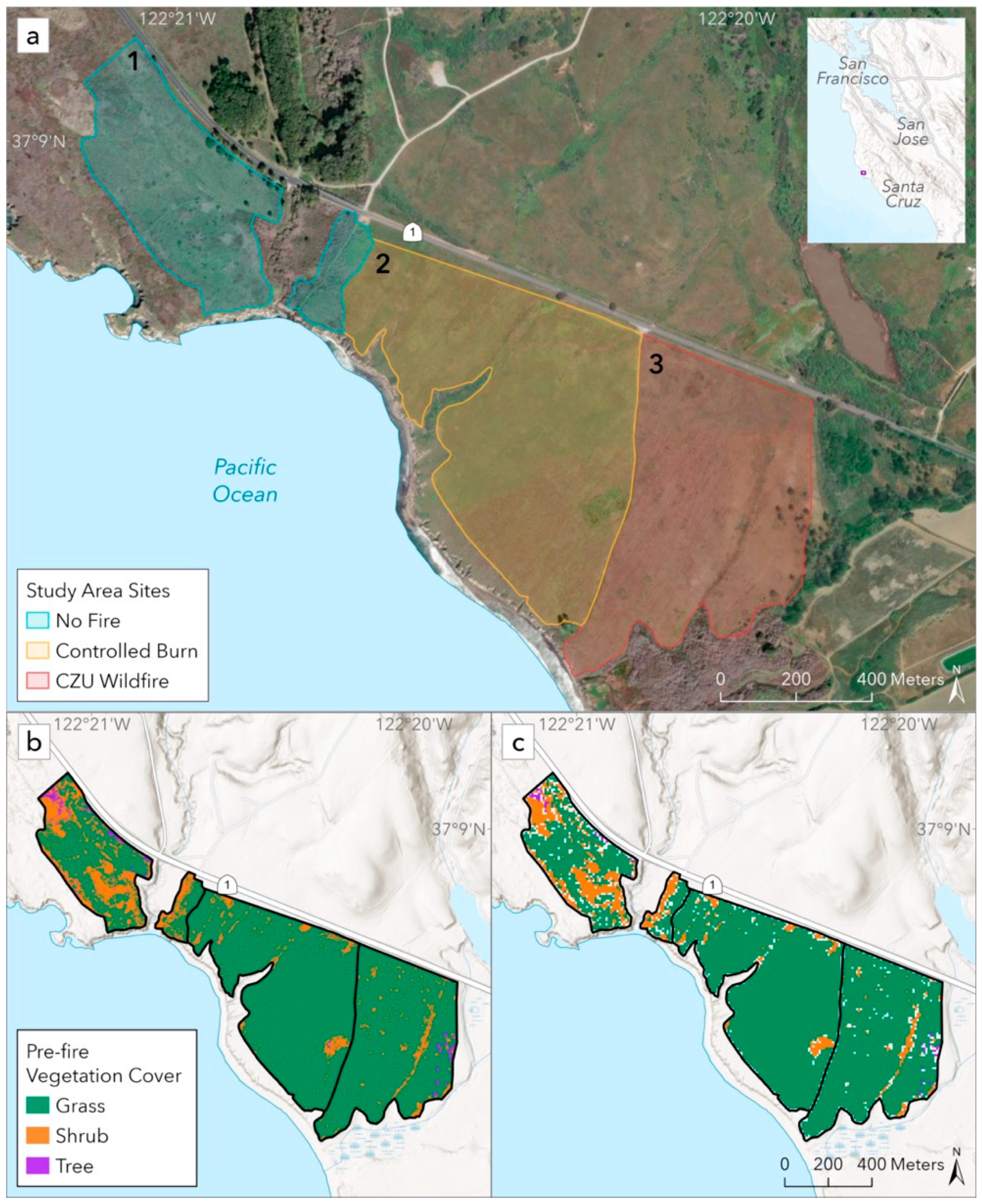
To test our proposed methodology, we chose a unique contiguous grassland along the Central Coast of California with three diverging fire histories: one site that has undergone low-severity controlled burns every two to three years, a second site that experienced a high-severity wildfire, and a third adjacent site with no recorded fire in over 100 years (
Figure 1a). These sites also represent a natural comparison between three emblematic land management approaches in California: frequent intentional burning, fire suppression followed by high-severity wildfire, and fire suppression, respectively. Given the logistical challenges associated with researching fire, few studies have examined sites with a history of repeated intentional burns. Fewer still have been able to compare a repeatedly burned grassland to a grassland burned by a wildfire, or studied the recovery of those sites over many years.
These three sites had different proportions of grass, shrub, and tree cover prior to both fire events, which we expected to respond differently to the low and high-severity fires. We therefore stratified our analysis based on these vegetation cover types to further interrogate the ecological impacts of the fires. We predicted that the grasses and shrubs on the controlled burn site would benefit from the influx of nutrients and sunlight provided by the low-severity fire, leading to a temporary increase in productivity [
16]. In the case of the wildfire, we predicted that the shrubs and trees would experience an immediate decline in productivity in the year following the fire due to direct mortality, with these plants either converting to grass or, in the case of shrubs, resprouting in subsequent years [
15,
24]. We expected productivity to fluctuate on the no fire site due to climatic variation during the study period, but to a much lesser degree than on the sites that experienced fires. While we expected a temporary shock from both fire events, we aimed to investigate whether the wildfire site would appear more phenologically similar to the controlled burn site after three years, or return to a signal more closely resembling the site with no fire.
Recognizing the growing need to monitor the impacts of smaller, low-severity fires and the aforementioned gaps in current methodologies, we propose methods that leverage a dense time series of high spatial resolution imagery to reveal important inter- and intra-annual variation in grasslands before and after two fires. Specifically, we propose three different spatiotemporal analyses that illuminate changes in three key metrics derived from Sentinel-2 imagery. We tested these methods in a unique study area in which adjacent grasslands have diverging fire histories to show how recovery from a low-severity controlled burn and a high-severity wildfire differ from each other and from an adjacent reference site with no recent fire. These methods are intended to be readily scalable and broadly applicable in monitoring the impacts of both high- and low-severity fires in grasslands and other low-biomass ecosystems.
4. Discussion
We used Sentinel-2 imagery to track the phenological indicators of post-fire recovery in adjacent coastal grasslands with varying fire histories: one site burned in a low-severity controlled burn, one site burned in a high-severity wildfire, and one site with no recent fire (
Figure 1a). The methods developed in this paper aimed to monitor post-fire recovery at a temporal resolution relevant to grasslands and at a spatial resolution small enough to capture intentional burns.
In interpreting the results, it is necessary to note that the three sites experienced different disturbance regimes prior to the study period. Both the no fire site and the wildfire site experienced no fires of any kind for more than 100 years prior to the study period, whereas the controlled burn site had been burned at low severity in the fall every two to three years since 1991, and most recently in 2017, 2020, and 2023. Only the 2020 controlled burn was directly included in this study. However, the baseline years included in this study (WYs 2019 and 2020) represented the second and third growing seasons following the fall 2017 controlled burn. While we focused on the effects of the 2020 controlled burn, it is important to keep in mind that the vegetation and seasonality on that site were also influenced by these prior fires.
To characterize differences in the pre-fire vegetation cover across the three sites, we conducted a vegetation classification based on pre-fire imagery captured in May 2020 (WY 2020) and drew on vegetation survey data collected across all three sites in April 2022 as part of related research [
14]. Both of these sources revealed that these three sites had different vegetation composition communities: broadly, the controlled burn site was dominated by grass with sparsely distributed shrubs, while the no fire and wildfire sites had higher shrub cover as well as evergreen trees encroaching along the edges. These differences are likely related in part to their distinct historical and current fire regimes (
Figure 1a) [
14].
We expected the inter- and intra-annual NBR signal to vary based on many factors. First, NBR can be an indicator of vegetation health, as photosynthetically active vegetation with a higher moisture content will reflect a higher NBR value [
32]. We expected this to vary between years based on climatic differences (e.g., how much precipitation falls and when in the year it falls), nutrient availability, and disturbance. The no fire site served as a reference to account for relative differences in NBR metrics within and between years that are likely due to climatic variation, as that site experienced no known disturbances during the study period.
Second, vegetation will exhibit higher NBR values when it is photosynthetically most active (during its growing season), and lower values when it is dormant or less photosynthetically active. Therefore, we expected the NBR signal to vary differently for the grasses, shrubs, and trees present within the study area based on differences in their seasonal growth patterns. In general, the growing season of these grasses begins in December or January—soon after California’s wet season begins—and lasts through May or June, senescing during the dry season [
40]. Shrubs in this area begin their growing season in January, but differ from grasses in that they stay photosynthetically active through the dry season, flowering from August to December [
41]. The coniferous trees common in this area are evergreen and therefore exhibit less seasonal variation than the shrubs or grasses, but they do grow new shoots in the spring months. Given these expected differences in seasonal signals and that the three sites had differences in their pre-fire vegetation communities, we first conducted each analysis to compare the three sites, then additionally stratified them by these three types of vegetation cover.
4.1. Full Time Series
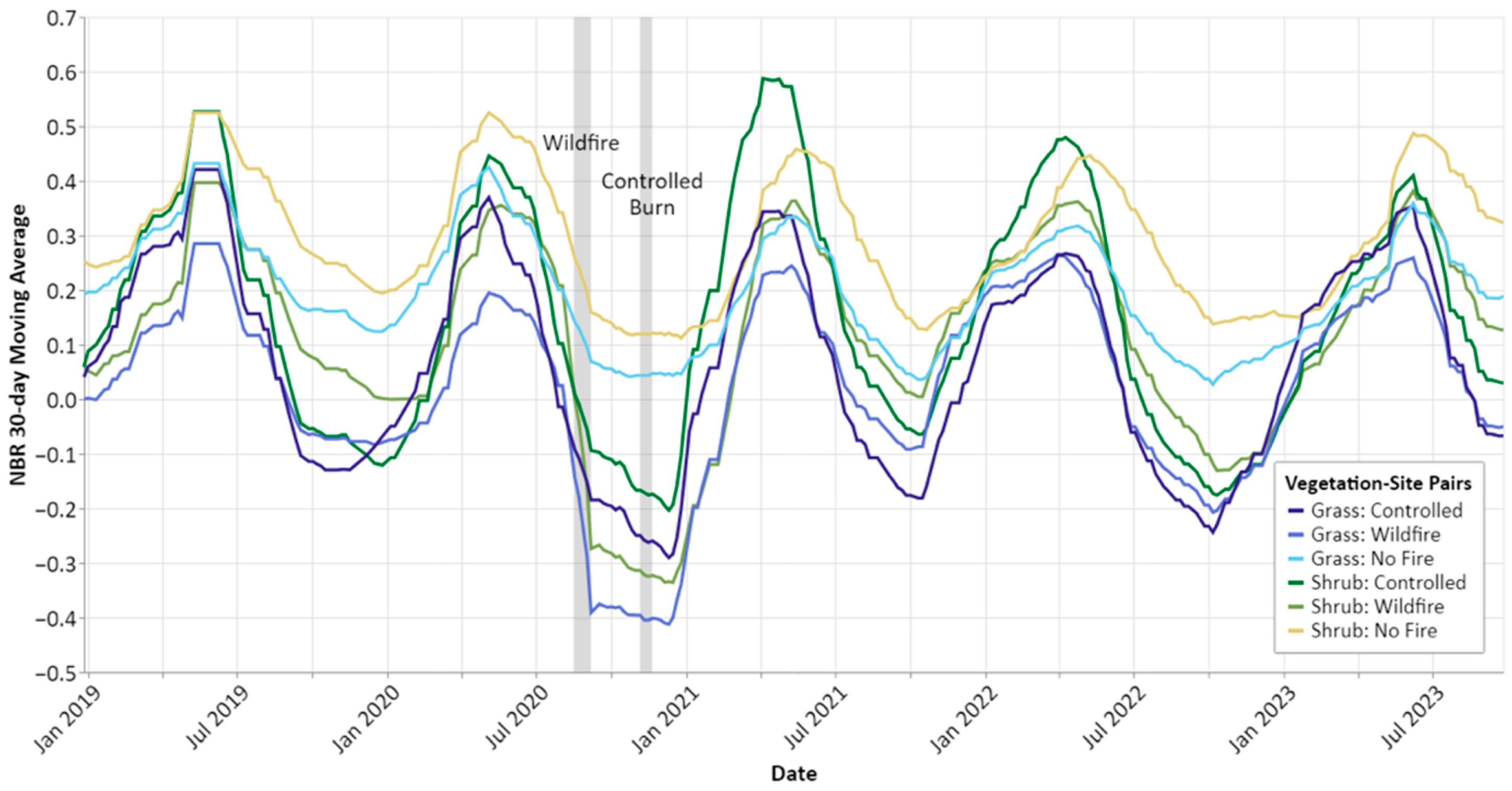
The full time series revealed complex patterns of disturbance and recovery in these grasslands as well as differences in phenological timing between the sites (
Figure 2) and vegetation–site pairs (
Figure 3). First, the full NBR time series revealed important changes in pre- and post-fire seasonality across the three sites (
Figure 2). There was less intra-annual variation in the NBR on the no fire site, which we would expect given its lack of disturbance and higher abundance of shrubs and trees compared to the other two sites. The average NBR was consistently higher on the no fire site except for two periods in which the average NBR on the controlled burn site surpassed it: (1) in the green-up period immediately following the controlled burn (WY 2021), and (2) in the green-up period two years after the controlled burn (WY 2023). One plausible explanation for this considerable change in NBR relative to the reference site could be that the controlled burn resulted in decreased competition for water and light as well as an influx of available nutrients that enabled the grasses and shrubs on that site to grow earlier and at a faster rate than the other two sites [
42]. While we did see a post-fire increase in the average NBR on the wildfire site, it was still consistently lower than that of the controlled burn site, which agrees with other research showing that higher severity fire may not benefit grassland productivity as it can cause soil erosion, soil hydrophobicity, nutrient depletion, and the mortality of belowground biomass [
42,
43]. Importantly, our findings also suggest that the expected benefits of the controlled burn (e.g., increased access to nutrients, water, and sunlight) were still distinguishable up to two years after the burn.
Across all years, the vegetation on the no fire site appeared to senesce at a similar point in time as the fire sites but at a slower rate and to a lesser degree, suggesting that vegetation on that site experienced less seasonal change (
Figure 2). This similarity in senescence timing aligns with other research showing that senescence in grasslands is primarily driven by changes in day length [
44]. The wildfire site exhibited less seasonal change than the controlled burn site prior to the wildfire, but more closely matched the shape of the controlled burn site’s growth curve after the wildfire (
Figure 2). This suggests that this disturbance immediately increased the seasonal variation in that grassland, and that some of that increase persisted even two years after the wildfire.
Next, we stratified the full NBR time series by vegetation type to analyze differences in pre- and post-fire seasonality across the vegetation–site pairs (
Figure 3). This vegetation stratification step allowed us to test various comparisons, for example, whether shrubs responded differently to fires of different severities, and whether grasses and shrubs responded differently to the same fire.
Figure 3 shows that the pattern of earlier and faster green-up on the controlled burn site following the controlled burn was largely driven by the shrub-dominated pixels, both in the green-up period immediately following the burn (WY 2021) and two years later (WY 2023). This indicates that shrub-dominated areas responded more favorably to this low-severity fire than grass-dominated areas, which may be due to differences in direct mortality resulting from the fire (i.e., shrubs did not experience mortality of aboveground biomass to the same extent as grasses in the controlled burn). Grass-dominated areas appeared to respond similarly to both types of fire, which suggests that the wildfire did not burn at high enough severity to cause lasting damage to the soil or grasses [
42,
43].
When comparing the response of shrubs to these two types of fire, we observed that the shrub-dominated areas on the controlled burn site exhibited earlier green-up and a higher magnitude of green-up than the pre-fire values when compared to the wildfire site, while the shrub-dominated areas on the wildfire site appeared to be phenologically similar pre- and post-fire. This suggests that while shrub productivity may be increased following low-severity fire, it may not be significantly impacted by high-severity fire (or at least not within a three year post-fire period). One possible explanation is that the negative NBR signal from shrub mortality following the wildfire was balanced by the regrowth of grasses and the resprouting of fire-adapted shrubs [
14,
15]. It is important to note that while low-severity fire appeared to increase the productivity of individual shrubs, we do not suggest that it increases the abundance of shrubs. In fact, the frequent, low-severity burns conducted at the controlled burn site likely reduce the establishment and encroachment of more shrubs and other woody vegetation, which is one of the goals of this style of fire management [
14,
15].
4.2. Annual Metrics
While the full time series are vital in understanding differences in intra-annual patterns between the sites (
Figure 2) and vegetation–site pairs (
Figure 3), it can be difficult to distinguish longer-term patterns occurring across multiple water years. To summarize and facilitate the interpretation of these complex time series, we focused on three key annual metrics: mean, fifth percentile, and amplitude (
Figure 4 and
Figure 5). Each of these three metrics summarized ecologically important information about the growth patterns on each site (
Figure 4) or vegetation–site pair (
Figure 5). Briefly, the annual mean values of VIs such as NBR are commonly used to track changes in the overall vegetation condition and are correlated with annual gross primary productivity [
45]. The fifth percentile NBR value is useful in tracking disturbance (the minimum values of NBR are designed to track damage from fire) and discerning the extent of senescence during the dry season, which can help to distinguish shrub- or tree-dominated areas from grass-dominated ones (
Figure 3) [
31]. Finally, the amplitude of the NBR curve (here defined as the 5th percentile value subtracted from the 95th percentile value) is an efficient measure of both seasonality and annual gross primary productivity as it is directly correlated to the area under the annual phenological curve [
46].
Changes in these three annual metrics revealed important differences across the three sites (
Figure 4). First, it is important to note that changes in the NBR signal on the no fire site are likely due primarily to year-to-year differences in climate. California experienced lower than average precipitation in WYs 2020 and 2021, and we saw a general trend of decreasing mean and fifth percentile NBR over the study period on the no fire site (
Figure 4). Therefore, when measuring this ecosystem’s resilience, it is best not to test whether these metrics recovered to the pre-fire values because climatic differences between these years may have resulted in a gradual decline. For that reason, we employed a pairwise offset method to facilitate the analysis of changes in each metric relative to the reference (no fire site), thereby accounting for climatic differences between years (similar to [
18]). These pairwise offset charts (
Figure 4, columns 2 and 3) highlight how these metrics changed on the fire sites relative to the no fire site, which experienced the same climatic changes between water years but no disturbance.
While their overall trajectories were similar, there were clear differences in the magnitude of response to disturbance between the controlled burn and wildfire sites (
Figure 4). Specifically, the wildfire site exhibited a greater relative decline in fifth percentile NBR during the year of the fire and a much greater relative increase in NBR amplitude following the fire. Interestingly, our relative comparison between the controlled burn site and the reference site indicated that the annual NBR metrics recovered to pre-fire values within one water year following the low-severity burn, and was potentially more productive than the no fire site in WY 2023 (
Figure 4c,f,i). This aligns with our expectations that grasslands recover quickly and benefit from low-severity fire, and that high-severity fire may lead to larger ecological shocks [
16].
The amplitude of the phenological curve is fairly under-utilized compared to other annual metrics and our results showed it to be important in the analysis of grassland disturbance and recovery. The controlled burn site had a considerably higher amplitude than either the wildfire or no fire site across all years (
Figure 4g), indicating that this site is more seasonally variable. As predicted based on their similar pre-fire disturbance regimes, the amplitude on the wildfire site was very comparable to the no fire site for the two pre-fire years of our study (
Figure 4g). The wildfire site’s amplitude increased dramatically in the year of the wildfire and while it decreased in the years since, it remained significantly higher than the no fire site in WY 2023 (
Figure 4g,h). Therefore, it will be important to study whether the amplitude and seasonality on the wildfire site either diverges permanently from the no fire site, or if it eventually converges back in the absence of repeated disturbance.
Next, we analyzed changes in these three annual metrics across the eight vegetation–site pairs (
Figure 5). While the separation of vegetation types was imperfect—Sentinel-2 pixels cover an approximate area of 100 m
2 and therefore almost always include a mix of vegetation types—our analysis revealed important differences between the phenological signals of grass-, tree-, and shrub-dominated pixels both before and after the fire events. As we hypothesized, the mean and fifth percentile NBR values were higher for the shrub- and tree-dominated pixels than for the grass-dominated pixels on all three sites. On the no fire site, the difference between the average mean and fifth percentile NBR values of these two groups of vegetation stayed consistent across all five water years (
Figure 5a,d). This indicates that the signals on the reference site were acting as we expected, with all vegetation types experiencing slight differences each year due to variations in climate.
Vegetation changes over the course of the study (WYs 2019 to 2023) were not observed on the no fire and controlled burn sites [
14]. However, it was more difficult to interpret the post-fire signals on the basis of vegetation on the wildfire site—the CZU wildfire burned at very high severity and caused significant vegetation changes. Specifically, we found that the majority of shrubs on that site were top-killed by the wildfire (meaning that they exhibited partial or total mortality of aboveground biomass) and had just begun to resprout in April 2022 [
14]. In this case, a decline in fifth percentile NBR in the following years could be due to seasonally variable grasses replacing these dead shrubs, rather than a decline in vegetation health. Therefore, in the case of high-severity fire, it is important to field-verify whether a change in VI values is due to shifts in vegetation cover or changes in vegetation health.
On the controlled burn site, the shrub-dominated areas exhibited greater mean NBR values for two growing seasons post-fire, confirming our earlier finding that shrub-dominated areas appeared to benefit from this low-severity fire (
Figure 5c). The grass-dominated areas on the controlled burn site exhibited a decline in mean and fifth percentile NBR in the year of the fire but appeared to recover in one or two years relative to pre-fire values on that site and corresponding values on the no fire site, which is consistent with other literature showing that these grasses are highly resilient to low-severity fire (
Figure 5c,f) [
16]. The trajectories on the wildfire site appeared to be similar again to the controlled burn site but greater in magnitude, with the exception of shrubs: the mean NBR of shrub-dominated areas on the wildfire site sharply declined in the year of the wildfire, whereas those on the controlled burn site exhibited no decline in the year of that fire (
Figure 5b,c). This is likely due to the fact that the high-severity wildfire top-killed many shrubs on that site, while the low-severity controlled burn resulted in minimal canopy mortality of the shrubs [
14]. However, the mean NBR of the shrub-dominated areas on the wildfire site quickly rebounded in WYs 2022 and 2023, which ran contrary to our expectation that the signal of shrub re-sprouting would occur over three or four years based on other studies that have measured the rates of resprouting in coyote brush (
Baccharis pilularis), the dominant shrub within the wildfire site [
15,
24]. Therefore, it is likely that this signal of fast recovery is due to grass regrowing beneath top-killed shrubs, and the same is true for trees [
14].
The amplitude of the NBR curve varied significantly between the three sites and between vegetation types (
Figure 5g–i). On the no fire site, the amplitude of NBR was relatively constant between the water years and for all three vegetation types (
Figure 5g). It was surprising that the amplitude of the grass- and shrub-dominated areas on the no fire site—and wildfire site prior to the fire—were much lower than the controlled burn site (
Figure 5g–i). The lower seasonality and lower annual growth on these sites could be due to a greater competition for light, water, and nutrients in the absence of fire or other disturbances, and potentially the build-up of dead biomass. It is also possible that the observed differences in amplitude on the three sites can be explained partially by differences in species composition, given that the amplitude on the wildfire site increased in all post-fire water years, and likely experienced at least temporary shifts in vegetation composition due to the canopy and total mortality of the shrubs and trees. However, given that the amplitude of shrub-dominated areas was higher than that of the grass-dominated areas across all water years and sites (
Figure 5g–i), it is more likely that increased amplitude values post-fire were due not to shrub mortality, but rather to higher annual productivity following the fire caused by a release from competition for light, nutrients, and water [
16]. Given the relatively lower amplitude on the wildfire site two years post-fire, our findings suggest that the regular controlled burning on the site (roughly every three years) is promoting greater annual growth of both grass and shrub species, even in the third growing season following a burn, and to a greater extent than infrequent high-severity fire. This fits with our expectation, as many of the plants in California’s coastal grasslands are adapted to this disturbance regime [
16].
4.3. Spatial Variation
In addition to these temporal variations, all three metrics displayed varying degrees of spatial heterogeneity across the study period that likely correspond to spatial variation in abiotic conditions, fire severity, and other factors (
Figure 6). Across both fire sites, we found that the areas with higher mean and fifth percentile NBR values across all years corresponded with depressions and drainages where we would expect water to gather based on the area’s topography. Interestingly, the NBR amplitude also appeared to follow this pattern. This suggests that the areas likely to have higher soil moisture farther into the dry season are more productive. The increase in spatial variation of NBR amplitude on the wildfire site following the wildfire is an interesting result that may be explained by variation in fire severity, resulting in differences in the availability of nutrients, light, and water [
42,
43]. Investigating the spatial variation in these temporal changes is a necessary step as it checks for additional factors that may be contributing to the signals in the full time series and the annual NBR metrics.
4.4. Methodological Strengths
This is a strategic approach to the remote sensing of fire recovery in grasslands that analyzes intra- and inter-annual changes over the spatial and temporal dimensions. Unlike in forests where intra-annual changes may be less significant, the health of a grassland may be signaled by changes in the timing and magnitude of green-up and senescence [
17]. While all of our results revealed important details about the ecosystem’s response to fire, we found that the full time series stratified by vegetation type (
Figure 3) was valuable for discerning overall patterns. Our methods overcome some of the key barriers that make full time series charts like this one less accessible to scientists and land managers. Performed on a local computer, preparing and analyzing a dense time series like this one would require a potentially prohibitive amount of computational power and storage space. Our analysis instead uses the GEE Python API, which stores and analyzes all of the imagery on external servers, enabling the analysis of hundreds of images on a small laptop. Another important barrier to full time series analysis is the exclusion of clouds and noisy images, which we carried out by using an automated algorithm that is directly transferable to other areas. Finally, by calculating the 30-day moving average as we did here, scientists and land managers are able to study these important temporal patterns without relying on a perfect cloud filtering algorithm.
Annual temporal summaries helped make these dense datasets more readily legible and understandable. Furthermore, the pairwise offsets comparing relative changes on the fire sites to the no fire site (
Figure 4) allowed us to investigate the post-fire recovery time while accounting for climatic variation, similar to other works that have used temporal changes in the mean VI values to measure the rates of recovery [
18]. This analysis of post-fire recovery time may help managers determine the appropriate return interval for intentional burning, which is relatively understudied in California’s grasslands [
12,
15]. In the case of the controlled burn site, our findings suggest that the ecological benefits of this intentional burning (e.g., increased vegetation productivity due to greater access to water, nutrients, and sunlight) may last for at least three growing seasons following the burn. In this ecosystem, the optimal return interval of intentional burning may be instead driven by the desire to reduce the encroachment of shrubs and trees, which, if given more than three growing seasons to establish, may be difficult to kill with low-severity fire [
15,
47].
Depending on the application, maps showing the variation in each of these metrics over space (such as
Figure 6) may reveal critical spatial patterns that are meaningful for land managers. We would expect this spatial evaluation to be most important on sites with variable soil types, microclimates, or other abiotic factors. Taken together, our three methodological approaches—analyzing the full time series, summarizing it through annual metrics, and evaluating spatial variation—successfully condensed complex spatiotemporal data to reveal ecologically relevant information.
The methods we propose here are designed to be accessible, transferable, and computationally lightweight. First, we utilized free data and open source software whenever possible. The geemap package (v. 0.32.1) is open source, and Sentinel-2 imagery, which is reliably orthorectified and atmospherically corrected, is free to use. The main exception to this is the GEE Python API, which is not open source but is free to researchers and nonprofit organizations. Second, in contrast to many phenological studies of disturbance, we designed methods that do not rely on curve-fitting models. While curve-fitting models can be very useful in summarizing phenological patterns, they typically rely on user-inputted assumptions that are tailored to the seasonal patterns of a given study area or ecosystem, making methods that rely on them less broadly applicable or replicable [
20,
48]. We instead made use of parameters related to the shape of the phenological curve, namely the mean, fifth percentile, and amplitude, which are efficient to calculate, do not require user-inputted assumptions, and are not negatively affected by the temporal lag caused by the coarser temporal resolution of Sentinel-2 imagery [
22,
23,
45].
4.5. Limitations and Future Work
Our study utilized field data from only one post-fire growing season, which was not sufficient to thoroughly validate our remotely sensed findings. As discussed previously, fluctuations in NBR can be the result of changes in vegetation health, density, or type (e.g., shrubs converting to grasses after a high-severity fire), and data collected in the field can help to differentiate among these possible causes. In particular, field monitoring may be especially useful in the case of high-severity wildfire, when vegetation type conversions are more common. Research that conducts multiple vegetation surveys over many years is now needed to validate which of these metrics is most helpful in capturing important vegetation changes, and whether the amplitude or other metrics can reliably differentiate between more specific vegetation types (i.e., annual and perennial grasses). However, the stochastic nature of wildfire means that pre-fire field data are not always available to scientists and land managers. Therefore, methods such as ours that do not rely on comprehensive field data may offer the only pre-fire view available. Future work could also take advantage of the full Sentinel-2 imagery archive to study relative changes in VIs extending as far back as 2015, which would enable a comparison of the impacts of an earlier controlled burn at the site (2017) to the two more recent burns (2020 and 2023). In addition, these methods are readily applicable to other vegetation indices, and future work could explore the relative strengths and weaknesses of other indices in the spatiotemporal analysis of grassland recovery from fire.
More research is now needed to test how these methods perform for other intentional burns in other grasslands worldwide as well compare how these methods perform in forest ecosystems. In addition, comparative analyses across multiple fires would help to identify the most informative pre- and post-fire time frames for such assessments like the minimum time needed to reliably determine differences between sites or to declare the recovery of the ecosystem parameters targeted by managers. As previously mentioned, longer-term studies of intentional burn recovery could also help to better illuminate the optimal fire return interval in different grasslands. Finally, these methods represent only one piece of evaluating intentional burning in grasslands. In particular, some of the goals of intentional burns such as supporting the growth of certain native grasses or the removal of invasive species are not easily captured with satellite imagery analysis.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}