
Multi-Temporal and Time-Lag Responses of Terrestrial Net Ecosystem Productivity to Extreme Climate from 1981 to 2019 in China

Abstract
:1. Introduction
2. Materials and Methods
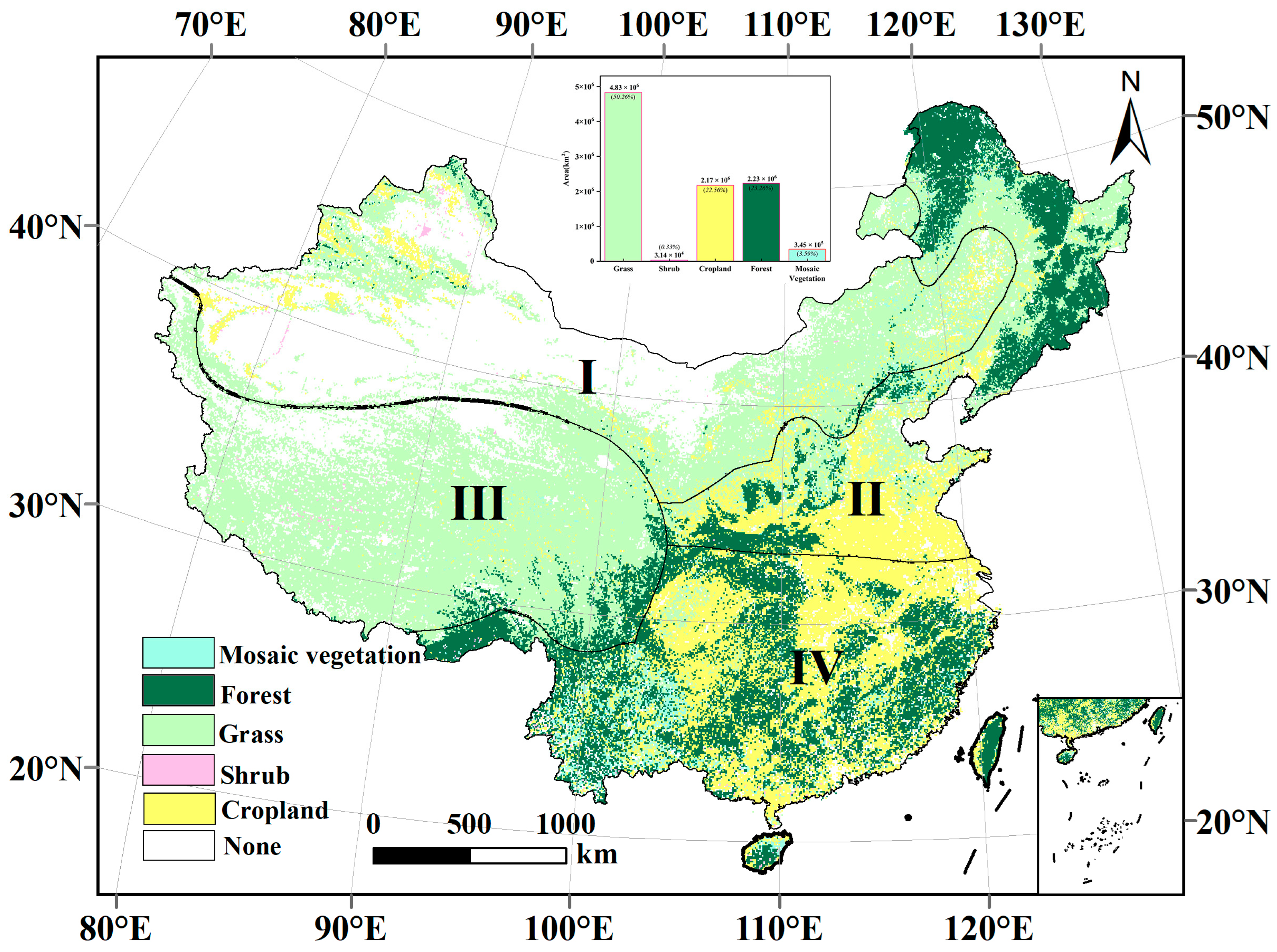
2.1. Study Area
2.2. Material
2.3. Methods
2.3.1. Selection of Climate Extreme Indices
2.3.2. The Linear Regression Method
2.3.3. Partial Correlation between NEP and Extreme Climate Indices
2.3.4. Lagged Response of NEP to Extreme Climate Indices
3. Results
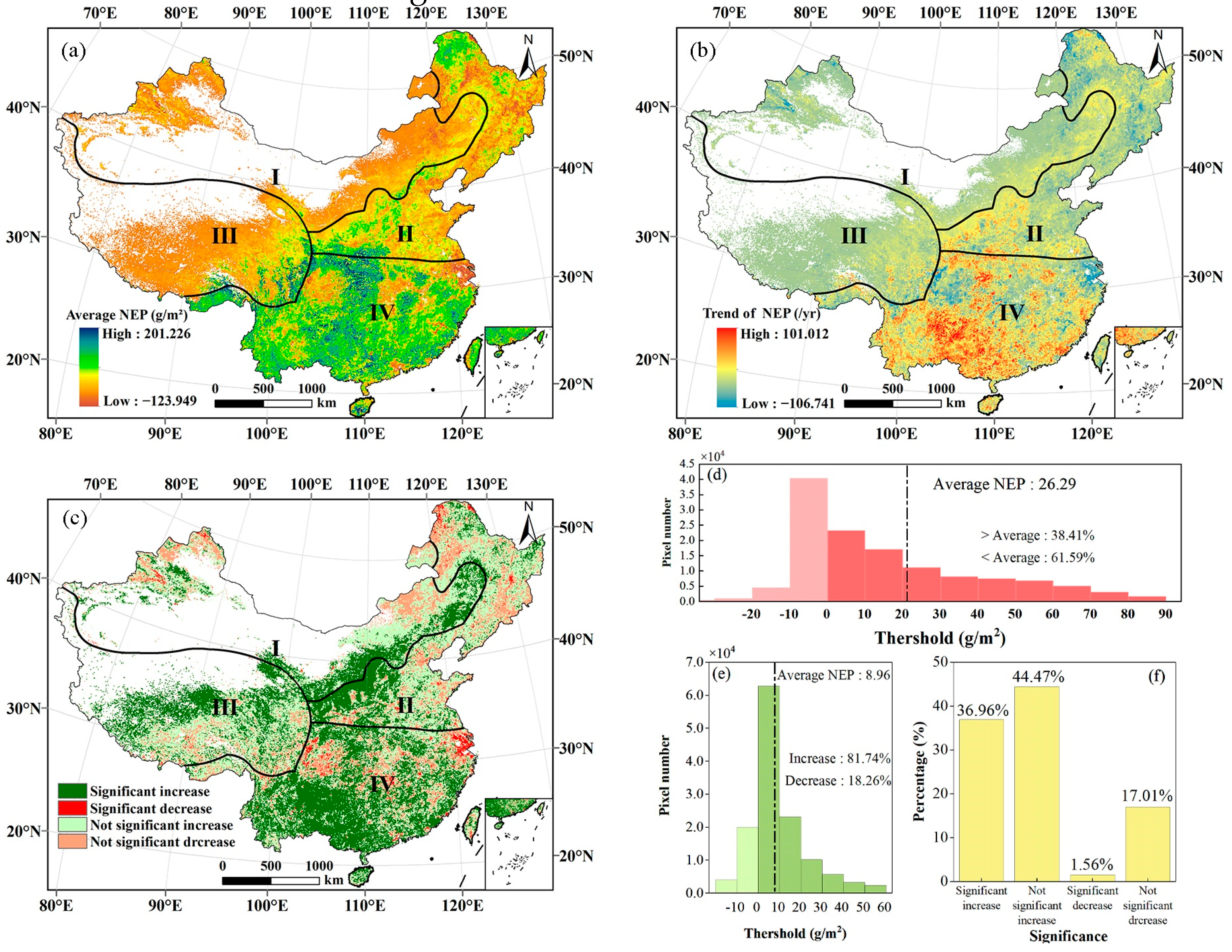
3.1. NEP Dynamics during 1981–2019
3.1.1. Spatiotemporal Dynamics of China’s NEP from 1981 to 2019
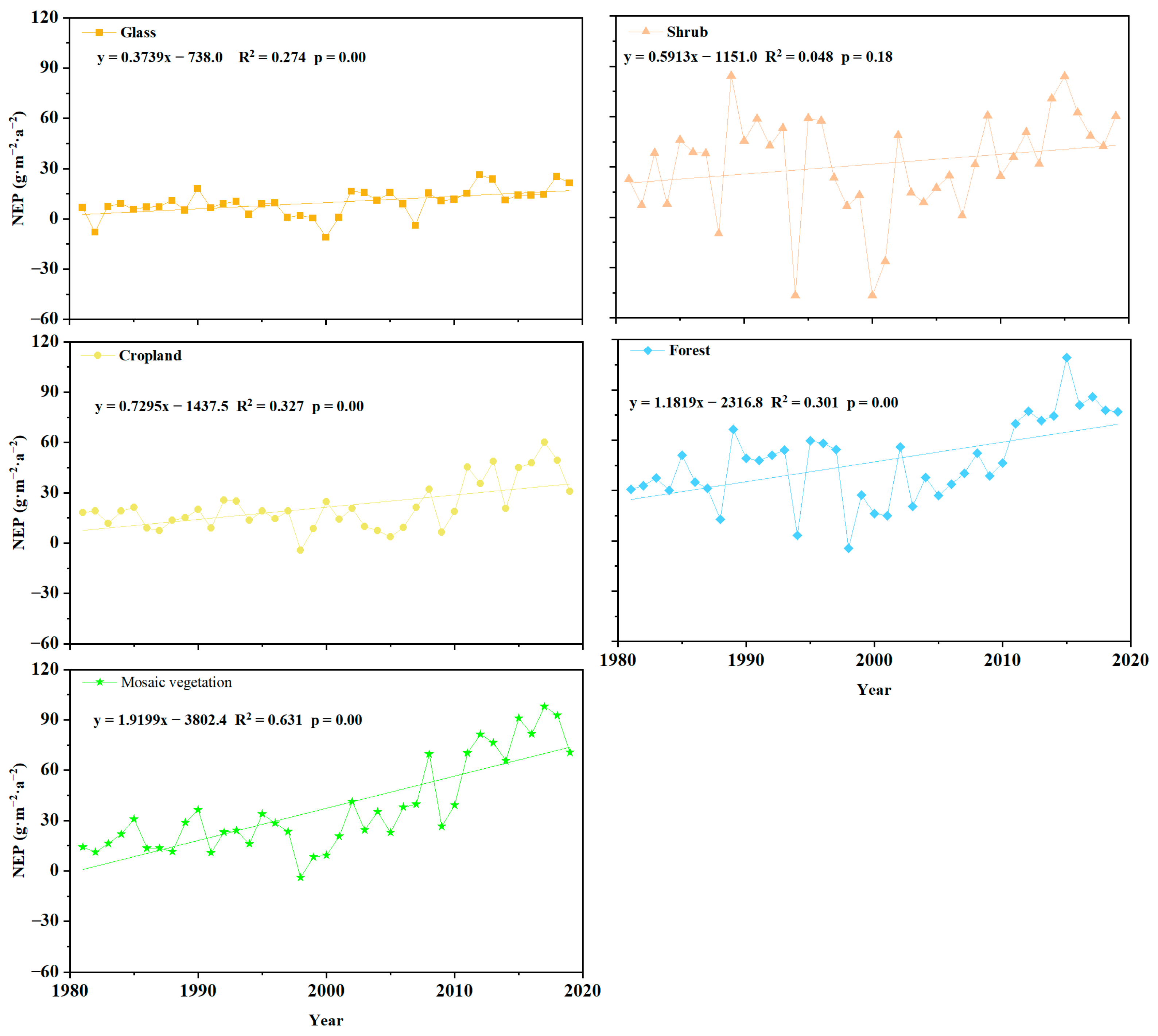
3.1.2. Spatiotemporal Variations in Vegetation NEP from 1981 to 2019
3.2. Annual Variation of Extreme Climate Indices during 1981–2019
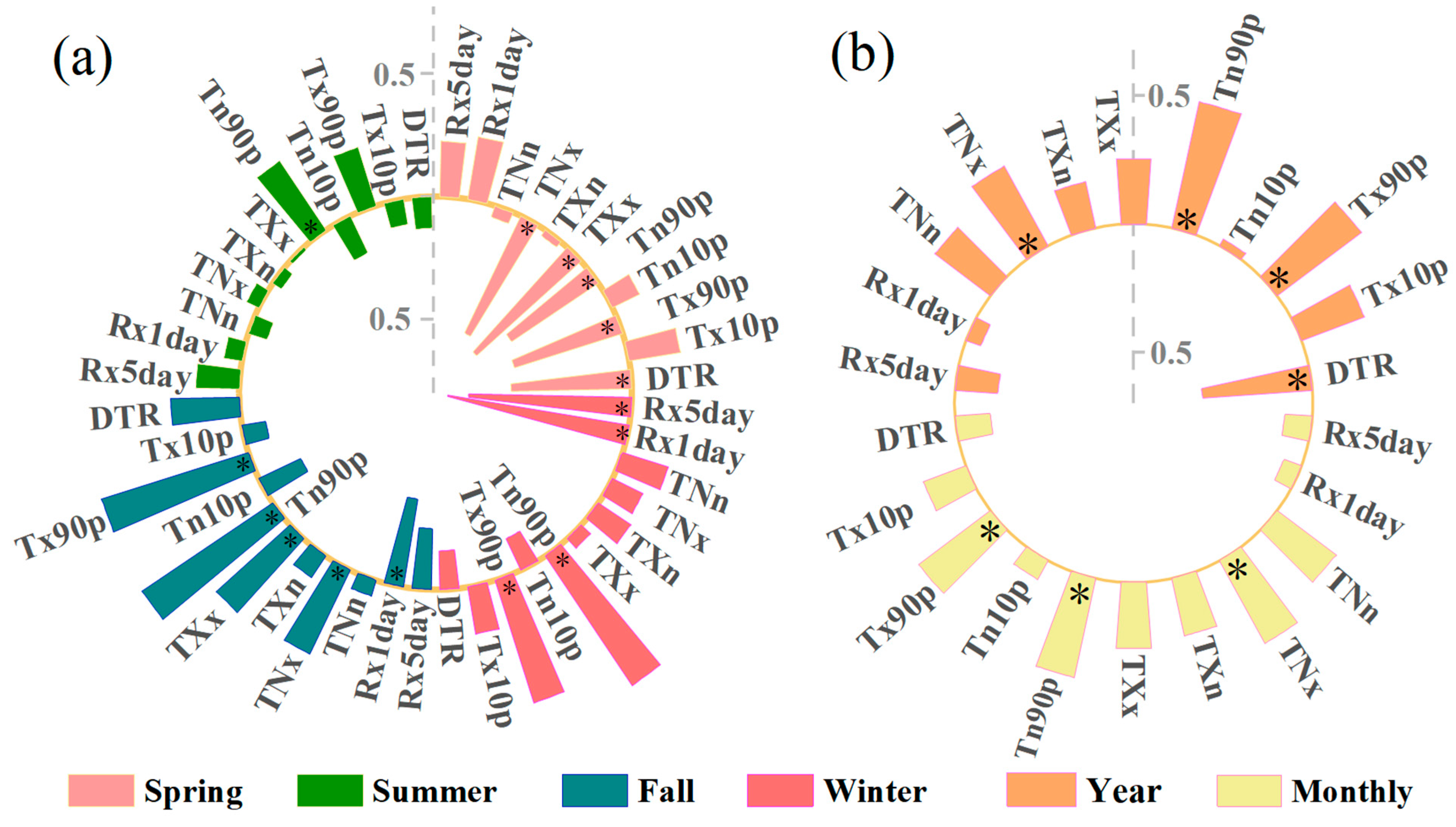
3.3. The Response of NEP to Extreme Climate Indices at Annual, Seasonal, and Monthly Time Scales
3.4. Seasonal Response of NEP to Extreme Climate Indices across Different Vegetation
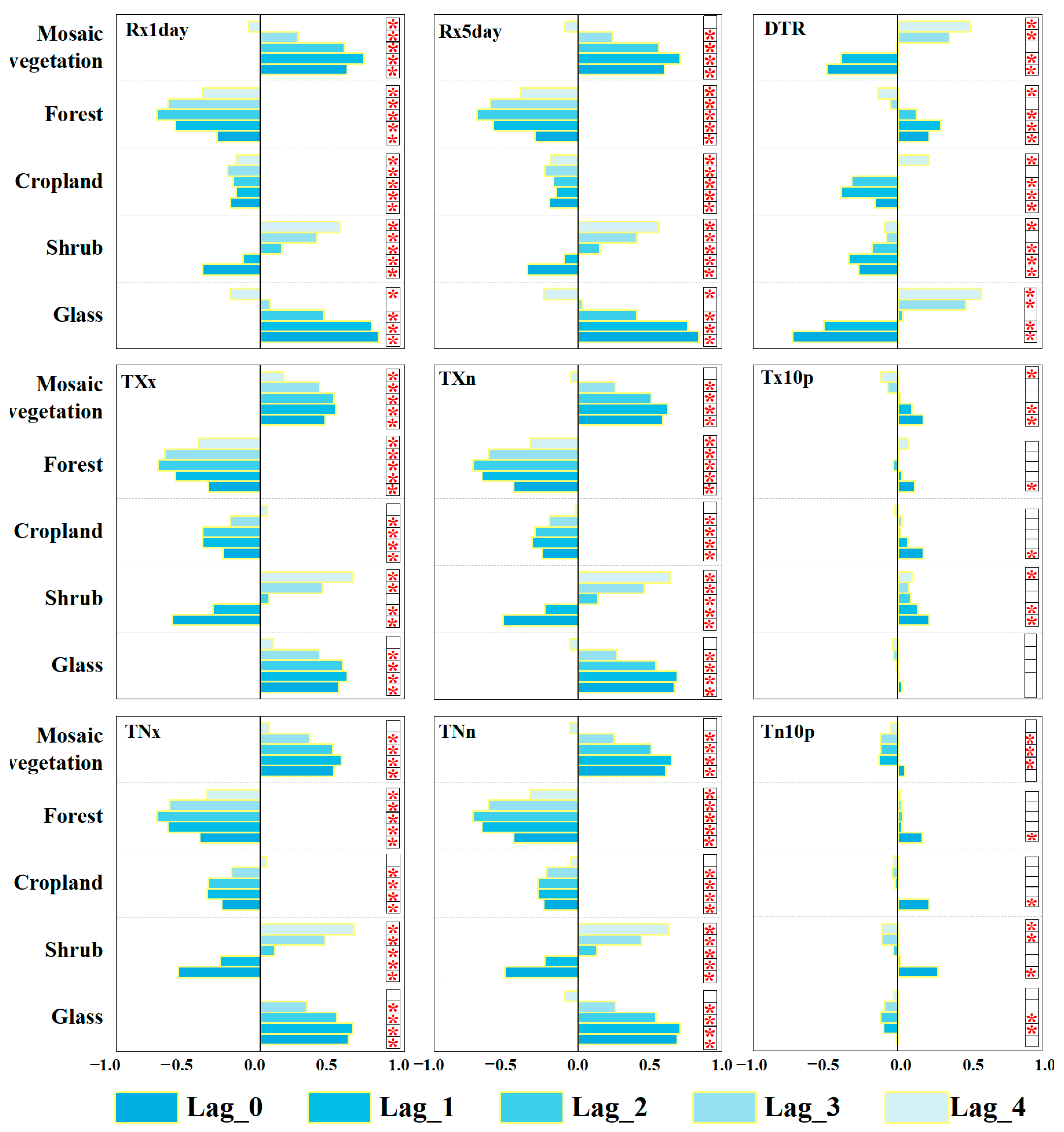
3.5. Time-Lag Response of NEP to Extreme Climate Indices
4. Discussion
4.1. Dynamic of NEP and Extreme Climate Indices in Mainland China
4.2. Multi-Temporal Response Mechanisms of Net Ecosystem Productivity to Extreme Climate in Different Vegetation
4.3. Seasonal Response of Net Ecosystem Productivity to Extreme Climate in Different Vegetation
4.4. The Lag-Time Effect of Net Ecosystem Productivity Response to Extreme Climate by Different Vegetation Types
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qian, Y.; Chakraborty, T.C.; Li, J.; Li, D.; He, C.; Sarangi, C.; Chen, F.; Yang, X.; Leung, L.R. Ruby. Urbanization Impact on Regional Climate and Extreme Weather: Current Understanding, Uncertainties, and Future Research Directions. Adv. Atmos. Sci. 2022, 39, 819–860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, X.; Jiang, H.; Qiao, S.; Guan, M.; Huang, Y.; Gong, R. Widespread decline in winds promoted the growth of vegetation. Sci. Total Environ. 2022, 825, 153682. [Google Scholar] [CrossRef] [PubMed]
- Le Quere, C.; Andrew, R.M.; Friedlingstein, P.; Sitch, S.; Hauck, J.; Pongratz, J.; Pickers, P.A.; Ivar Korsbakken, J.; Peters, G.P.; Canadell, J.G.; et al. Global Carbon Budget 2018. Earth Syst. Sci. Data 2018, 10, 2141–2194. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Sun, W.; Chang, J.; Zhu, J.; Chen, L.; Wang, X.; Guo, Y.; Zhang, H.; Yu, L.; et al. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality. Sci. China Life Sci. 2022, 65, 861–895. [Google Scholar] [CrossRef]
- Niu, S.; Fu, Z.; Luo, Y.; Stoy, P.C.; Keenan, T.F.; Poulter, B.; Zhang, L.; Piao, S.; Zhou, X.; Zheng, H.; et al. Interannual variability of ecosystem carbon exchange: From observation to prediction. Glob. Ecol. Biogeogr. 2017, 26, 1225–1237. [Google Scholar] [CrossRef]
- Piao, S.; Yue, C.; Ding, J.; Guo, Z. Perspectives on the role of terrestrial ecosystems in the ‘carbon neutrality’ strategy. Sci. China Earth Sci. 2022, 65, 1178–1186. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, X.; Wang, J.; He, H.; Wang, S.; Wang, M.; Piao, S.; Yan, H.; Ju, W.; Gu, F.; et al. Interannual variability of terrestrial net ecosystem productivity over China: Regional contributions and climate attribution. Environ. Res. Lett. 2019, 9, 014003. [Google Scholar] [CrossRef]
- Zheng, J.; Fan, J.L.; Zhang, F.C. Spatiotemporal trends of temperature and precipitation extremes across contrasting climatic zones of China during 1956–2015. Theor. Appl. Climatol. 2019, 138, 1877–1897. [Google Scholar] [CrossRef]
- Gao, T.; Wang, H.X.J.; Zhou, T.J. Changes of extreme precipitation and nonlinear influence of climate variables over monsoon region in China. Atmos. Res. 2017, 197, 379–389. [Google Scholar] [CrossRef]
- Sun, G.; Wang, Z.; Zhu-Barker, X.; Zhang, N.; Wu, N.; Liu, L.; Lei, Y. Biotic and abiotic controls in determining exceedingly variable responses of ecosystem functions to extreme seasonal precipitation in a mesophytic alpine grassland. Agric. For. Meteorol. 2016, 228, 180–190. [Google Scholar] [CrossRef]
- Gao, Y.H.; Xiao, L.H.; Chen, D.L.; Xu, J.W.; Zhang, H.W. Comparison between past and future extreme precipitations simulated by global and regional climate models over the Tibetan Plateau. Int. J. Climatol. 2018, 38, 1285–1297. [Google Scholar] [CrossRef]
- Liu, S.; Huang, S.; Xie, Y.; Huang, Q.; Leng, G.; Hou, B.; Zhang, Y.; Wei, X. Spatial-temporal changes of maximum and minimum temperatures in the Wei River Basin, China: Changing patterns, causes and implications. Atmos. Res. 2018, 204, 1–11. [Google Scholar] [CrossRef]
- Jiang, H.L.; Xu, X. Impact of extreme climates on vegetation from multiple scales and perspectives in the Agro-pastural Transitional Zone of Northern China in the past three decades. J. Clean. Prod. 2022, 372, 133459. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Ciais, P.; Friedlingstein, P.; Myneni, R.B.; Cox, P.; Heimann, M.; Miller, J.; Peng, S.; Wang, T.; et al. A two-fold increase of carbon cycle sensitivity to tropical temperature variations. Nature 2014, 506, 212–215. [Google Scholar] [CrossRef]
- Ni, M.; Zhang, X.Y.; Jiang, C.; Wang, H.S. Responses of vegetation to extreme climate events in southwestern China. Chin. J. Plant Ecol. 2021, 45, 626–640. [Google Scholar] [CrossRef]
- Chen, Q.; Mao, F.J.; Du, H.Q.; Li, X.J.; Yin, S.Y.; Yan, M.J. Spatiotemporal variations of subtropical extreme precipitation and its influence on potential net primary productivity in China. Chin. J. Ecol. 2022, 41, 2117–2127. [Google Scholar]
- Islam, A.; Islam, H.; Shahid, S.; Khatun, M.; Almoajel, A. Spatiotemporal nexus between vegetation change and extreme climatic indices and their possible causes of change. J. Environ. Manag. 2021, 289, 112505. [Google Scholar] [CrossRef]
- Ren, J.Y.; Tong, S.Q.; Bao, Y.H.; Guo, X.M. Changes of extreme climate and its effect on net primary productivity in Inner Mongolia. Chin. J. Ecol. 2021, 40, 2410–2420. [Google Scholar]
- Han, D.D.; Mu, X.M.; Gao, P. Dynamic changes of vegetation in Loess Plateau and its response to extreme climate. Bull. Soil Water Conserv. 2020, 40, 247–254. [Google Scholar]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; Van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef]
- Moustakis, Y.; Papalexiou, S.; Onof, C.; Paschalis, A. Seasonality, intensity, and duration of rainfall extremes change in a warmer climate. Earths Future 2021, 9, 24–32. [Google Scholar] [CrossRef]
- Xie, Y.; Crary, D.; Bai, Y.; Cui, X.; Zhang, A. Modeling Grassland Ecosystem Responses to Coupled Climate and Socioeconomic Influences in Multi-Spatial-And-Temporal Scales. J. Environ. Inform. 2019, 33, 37–46. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Q.; Liu, R.; Zhao, Y.; Zhang, D. Effects of climate change and human activities on vegetation coverage change in northern China considering extreme climate and time-lag and accumulation effects. Sci. Total Environ. 2023, 20, 160527. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Zhang, Y.; Gu, F.; Shi, C.; Miao, N.; Liu, S. Impacts of climate extremes on ecosystem metrics in southwest China. Sci. Total Environ. 2021, 776, 145979. [Google Scholar] [CrossRef]
- Li, L.; Zheng, Z.; Biederman, J.A.; Xu, C.; Xu, Z.; Che, R.; Wang, Y.; Cui, X.; Hao, Y. Ecological responses to heavy rainfall depend on seasonal timing and multi-year recurrence. New Phytol. 2019, 223, 647–660. [Google Scholar] [CrossRef]
- He, H.; Wang, S.; Zhang, L.; Wang, J.; Ren, X.; Zhou, L.; Piao, S.; Yan, H.; Ju, W.; Gu, F. Altered trends in carbon uptake in China’s terrestrial ecosystems under the enhanced summer monsoon and warming hiatus. Natl. Sci. Rev. 2019, 6, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z. Boreal Ecosystem Productivity Simulator (BEPS), Model Item, OpenGMS. 2019. Available online: https://geomodeling.njnu.edu.cn/modelItem/262289db-f24a-4bed-9a42-b2fc7d27c473 (accessed on 20 December 2023).
- Wei, S.S.; Fang, H.L.; Schaaf, C.B.; He, L.M.; Chen, J.M. Global 500 m clumping index product derived from MODIS BRDF data (2001–2017). Remote Sens. Environ. 2019, 232, 111296. [Google Scholar] [CrossRef]
- Feng, X.; Liu, G.; Chen, J.M.; Chen, M.; Liu, J.; Ju, W.M.; Sun, R.; Zhou, W. Net primary productivity of China’s terrestrial ecosystems from a process model driven by remote sensing. J. Environ. Manag. 2007, 85, 563–573. [Google Scholar] [CrossRef]
- He, Q.; Ju, W.; Dai, S.; He, W.; Song, L.; Wang, S.; Li, X.; Mao, G. Drought Risk of Global Terrestrial Gross Primary Productivity Over the Last 40 Years Detected by a Remote Sensing-Driven Process Model. J. Geophys. Res.-Biogeosci. 2021, 126, e2020JG005944. [Google Scholar] [CrossRef]
- Guidigan, M.L.G.; Sanou, C.L.; Ragatoa, D.S.; Fafa, C.O.; Mishra, V.N. Assessing Land Use/Land Cover Dynamic and Its Impact in Benin Republic Using Land Change Model and CCI-LC Products. Earth Syst. Environ. 2018, 3, 127–137. [Google Scholar] [CrossRef]
- Vilar, L. Comparative analysis of CORINE and climate change initiative land cover maps in Europe: Implications for wildfire occurrence estimation at regional and local scales. Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 102–117. [Google Scholar] [CrossRef]
- Karl, T.R.; Nicholls, N.; Ghazi, A. CLIVAR/GCOS/WMO Workshop on Indices and Indicators for Climate Extremes—Workshop summary. Clim. Chang. 1999, 42, 3–7. [Google Scholar] [CrossRef]
- Peterson, T.C.; Manton, M.J. Monitoring changes in climate extremes—A tale of international collaboration. Bull. Am. Meteorol. Soc. 2008, 89, 1266–1271. [Google Scholar] [CrossRef]
- The MathWorks Inc. MATLAB Version: 9.13.0 (R2022b), Natick, Massachusetts: The MathWorks Inc. 2022. Available online: https://www.mathworks.com (accessed on 20 December 2023).
- Anderson, L.O.; Malhi, Y.; Aragao, L.; Ladle, R.; Arai, E.; Barbier, N.; Phillips, O. Remote sensing detection of droughts in Amazonian forest canopies. New Phytol. 2010, 187, 733–750. [Google Scholar] [CrossRef]
- Chen, T.; de Jeu, R.; Liu, Y.; van der Werf, G.; Dolman, A. Using satellite based soil moisture to quantify the water driven variability in NDVI: A case study over mainland Australia. Remote Sens. Environ. 2014, 140, 330–338. [Google Scholar] [CrossRef]
- Ma, Q.; Hänninen, H.; Berninger, F.; Li, X.; Huang, J. Climate warming leads to advanced fruit development period of temperate woody species but divergent changes in its length. Glob. Chang. Biol. 2022, 28, 6021–6032. [Google Scholar] [CrossRef]
- Park, T.; Chen, C.; Macias-Fauria, M.; Tømmervik, H.; Choi, S.; Winkler, A.; Bhatt, U.S.; Walker, D.A.; Piao, S.; Brovkin, V.; et al. Changes in timing of seasonal peak photosynthetic activity in northern ecosystems. Glob. Chang. Biol. 2019, 25, 2382–2395. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, X.; Tian, H.; Wu, X.; Gao, Z.; Feng, Y.; Piao, S.; Lv, N.; Pan, N.; Fu, B. Accelerated increase in vegetation carbon sequestration in China after 2010: A turning point resulting from climate and human interaction. Glob. Chang. Biol. 2021, 27, 5848–5864. [Google Scholar] [CrossRef]
- Wang, J.; Feng, L.; Palmer, P.I.; Liu, Y.; Fang, S.; Bösch, H.; O’Dell, C.W.; Tang, X.; Yang, D.; Liu, L.; et al. Large Chinese land carbon sink estimated from atmospheric carbon dioxide data. Nature 2020, 586, 720–723. [Google Scholar] [CrossRef]
- Lei, X.; Xu, W.; Chen, S.; Yu, T.; Hu, Z.; Zhang, M.; Jiang, L.; Bao, R.; Guan, X.; Ma, M.; et al. How Well Does the ERA5 Reanalysis Capture the Extreme Climate Events Over China? Part I: Extreme Precipitation. Front. Environ. Sci. 2022, 10, 921658. [Google Scholar] [CrossRef]
- Moon, J.-Y.; Choi, Y.; Kim, Y.; Min, S. Subseasonal to Annual Long-Term Trends in Climate Extremes Over East Asia, 1981–2021. Front. Earth Sci. 2022, 10, 9026–9042. [Google Scholar] [CrossRef]
- Xu, X.; Jiang, H.; Guan, M.; Wang, L.; Huang, Y.; Jiang, Y.; Wang, A. Vegetation responses to extreme climatic indices in coastal China from 1986 to 2015. Sci. Total Environ. 2020, 744, 140784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xiao, C.; Li, S. Construction of multi-extreme climate events composite grads index and comprehensive analysis of extreme climate in the Yangtze River Basin from 1961 to 2020. Chin. J. Geophys. 2023, 66, 920–938. [Google Scholar]
- Shen, M.; Piao, S.; Chen, X.; An, S.; Fu, Y.H.; Wang, S.; Cong, N.; Janssens, I.A. Strong impacts of daily minimum temperature on the green-up date and summer greenness of the Tibetan Plateau. Glob. Chang. Biol. 2016, 22, 3057–3066. [Google Scholar] [CrossRef] [PubMed]
- Davy, R.; Esau, I.; Chernokulsky, A.; Outten, S.; Zilitinkevich, S. Diurnal asymmetry to the observed global warming. Int. J. Climatol. 2017, 37, 79–93. [Google Scholar] [CrossRef]
- Ceppi, P.; Nowack, P. Observational evidence that cloud feedback amplifies global warming. Proc. Natl. Acad. Sci. USA 2021, 118, e2026290118. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Maclean, I.M.D.; Gardner, A.S.; Gaston, K.J. Global variation in diurnal asymmetry in temperature, cloud cover, specific humidity and precipitation and its association with leaf area index. Glob. Chang. Biol. 2020, 26, 7099–7111. [Google Scholar] [CrossRef]
- Quang-Van, D.; Chen, F.; Asano, Y.; Gu, Y.; Nishi, A.; Kusaka, H.; Niyogi, D. Causes for Asymmetric Warming of Sub-Diurnal Temperature Responding to Global Warming. Geophys. Res. Lett. 2022, 49, e2022GL100029. [Google Scholar]
- Panwar, A.; Kleidon, A. Evaluating the Response of Diurnal Variations in Surface and Air Temperature to Evaporative Conditions across Vegetation Types in FLUXNET and ERA5. J. Clim. 2022, 35, 2701–2728. [Google Scholar] [CrossRef]
- Ouyang, Z.; Sciusco, P.; Jiao, T.; Feron, S.; Lei, C.; Li, F.; John, R.; Fan, P.; Li, X.; Williams, C.A.; et al. Albedo changes caused by future urbanization contribute to global warming. Nat. Commun. 2022, 13, 3800. [Google Scholar] [CrossRef]
- Kopp, D.; Allen, D. Scaling spatial pattern in river networks: The effects of spatial extent, grain size and thematic resolution. Landsc. Ecol. 2021, 36, 2781–2794. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Xiang, F.; Qin, W.; Jiang, W. Vegetation dynamics and the relations with climate change at multiple time scales in the Yangtze River and Yellow River Basin, China. Ecol. Indicat. 2020, 110, 105892. [Google Scholar] [CrossRef]
- Lian, X.; Jiao, L.; Liu, Z.; Jia, Q.; Zhong, J.; Fang, M.; Wang, W. Multi-spatiotemporal heterogeneous legacy effects of climate on terrestrial vegetation dynamics in China. GISci. Remote Sens. 2022, 59, 164–183. [Google Scholar] [CrossRef]
- Zhou, J.; Jia, L.; Menenti, M.; van Hoek, M.; Lu, J.; Zheng, C.; Wu, H.; Yuan, X. Characterizing vegetation response to rainfall at multiple temporal scales in the Sahel-Sudano-Guinean region using transfer function analysis. Remote Sens. Environ. 2021, 252, 112108. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Li, L.Z.; Li, Y.; Huntingford, C.; Ciais, P.; Cescatti, A.; Janssens, I.A.; Peñuelas, J.; Buermann, W.; et al. Summer soil drying exacerbated by earlier spring greening of northernvegetation. Sci. Adv. 2020, 6, eaax0255. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Geng, X.; Yin, G.; Hänninen, H.; Hao, F.; Zhang, X.; Fu, Y.H. Legacy effect of spring phenology on vegetation growth in temperate China. Agric. For. Meteorol. 2020, 281, 107845. [Google Scholar] [CrossRef]
- Sun, B.; Jiang, M.; Han, G.; Zhang, L.; Zhou, J.; Bian, C.; Du, Y.; Yan, L.; Xia, J. Experimental warming reduces ecosystem resistance and resilience to severe flooding in a wetland. Sci. Adv. 2022, 8, eabl9526. [Google Scholar] [CrossRef]
- Plank, T.; Manning, C.E. Subducting carbon. Nature 2019, 574, 343–352. [Google Scholar] [CrossRef]
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef]
- Cui, Y.; Ouyang, S.; Zhao, Y.; Tie, L.; Shao, C.; Duan, H. Plant responses to high temperature and drought: A bibliometrics analysis. Front. Plant Sci. 2022, 13, 1052660. [Google Scholar] [CrossRef]
- Huang, M.; Piao, S.; Ciais, P.; Peñuelas, J.; Wang, X.; Keenan, T.F.; Peng, S.; Berry, J.A.; Wang, K.; Mao, J.; et al. Air temperature optima of vegetation productivity across global biomes. Nat. Ecol. Evol. 2019, 3, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Piao, S.; Chen, A.; Zeng, Z.; Ciais, P.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Peng, S.; Peñuelas, J.; et al. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Glob. Chang. Biol. 2015, 21, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, Y.; He, B. Spatiotemporal dynamics of grassland pheonology and sensitivity to extreme precipitation in autumn in Qing-hai-Tibetan Plateau. Bull. Soil Water Conserv. 2023, 30, 353–363. [Google Scholar]
- Sun, Y.; Zhang, Y.; Zheng, Z.; Zhao, G.; Zhu, Y.; Gao, J.; Zhang, Y. Strong inhibiting effect of daytime warming but weak promoting effect of nighttime warming on carbon use efficiency in Northern Hemisphere. Glob. Planet Chang. 2023, 228, 104192. [Google Scholar] [CrossRef]
- Chamberlain, C.J.; Cook, B.I.; De Cortazar-Atauri, I.G.; Wolkovich, E.M. Rethinking false spring risk. Glob. Chang. Biol. 2019, 25, 2209–2220. [Google Scholar] [CrossRef]
- Yang, H.; Munson, S.M.; Huntingford, C.; Carvalhais, N.; Knapp, A.K.; Li, X.; Peñuelas, J.; Zscheischler, J.; Chen, A. The detection and attribution of extreme reductions in vegetation growth across the global land surface. Glob. Chang. Biol. 2023, 29, 2351–2362. [Google Scholar] [CrossRef]
- Mahecha, M.D.; Bastos, A.; Bohn, F.J.; Eisenhauer, N.; Feilhauer, H.; Hartmann, H.; Hickler, T.; Kalesse-Los, H.; Migliavacca, M.; Otto, F.E.; et al. Biodiversity loss and climate extremes—Study the feedbacks. Nature 2023, 11, 612. [Google Scholar] [CrossRef]
- Vanoni, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Drought and frost contribute to abrupt growth decreases before tree mortality in nine temperate tree species. For. Ecol. Manag. 2016, 382, 51–63. [Google Scholar] [CrossRef]
- Luo, Y.; Gessler, A.; D’Odorico, P.; Hufkens, K.; Stocker, B. Quantifying effects of cold acclimation and delayed springtime photosynthesis resumption in northern ecosystems. New Phytol. 2023, 240, 984–1002. [Google Scholar] [CrossRef]
- Holdrege, M.C.; Kulmatiski, A.; Beard, K.H.; Palmquist, K.A. Precipitation Intensification Increases Shrub Dominance in Arid, Not Mesic, Ecosystems. Ecosystems 2023, 26, 568–584. [Google Scholar] [CrossRef]
- Zang, Y.X.; Xu, W.X.; Wu, K.; Yang, W.K. Effect of Nitrogen Application on the Sensitivity of Desert Shrub Community Productivity to Precipitation in Central Asia. Front. Plant Sci. 2022, 13, 916706. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.H.; Wang, C.K.; Luo, Y.Q. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Liu, H.; Piao, S.; Ciais, P.; Wu, X.; Yin, Y.; Wang, H. Enhanced growth after extreme wetness compensates for post-drought carbon loss in dry forests. Nat. Commun. 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhou, L.; Chi, Y.G. Spring phenology rather than climate dominates the trends in peak of growing season in the Northern Hemisphere. Glob. Chang. Biol. 2023, 29, 4543–4555. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Wang, T.J.; Han, Q.; Dai, Y.B.; Wang, L.C.; Chen, X. Evaluation of Environmental Controls on Terrestrial Net Ecosystem Exchange of CO2: A Global Perspective from the FLUXNET Sites. J. Geophys. Res.-Atmos. 2022, 127, e2022JD037217. [Google Scholar] [CrossRef]
- Wang, L.; Hu, F.; Miao, Y.; Zhang, C.; Zhang, L.; Luo, M. Changes in Vegetation Dynamics and Relations with Extreme Climate on Multiple Time Scales in Guangxi, China. Remote Sens. 2022, 14, 2013. [Google Scholar] [CrossRef]
- Duan, Y.; Ren, Z.; Sun, Y. Time-lay effects of climate on water use efficiency in the Loess Plateau of northern Shaanxi. Acta Ecol. Sin. 2020, 40, 3408–3419. [Google Scholar]
- Zhu, X.J.; Yu, G.R.; Chen, Z.; Zhang, W.K.; Han, L.; Wang, Q.F.; Qi, H.; Yang, M.; Liu, Z.G.; Dou, X.J.; et al. Ecosystem responses dominate the trends of annual gross primary productivity over terrestrial ecosystems of China during 2000–2020. Agric. For. Meteorol. 2023, 343, 109758. [Google Scholar] [CrossRef]
- Xu, M.; Sun, Y.; Zhang, T.; Zhang, Y.; Zhu, J.; He, Y.; Wang, L.; Yu, G. Biotic effects dominate the inter-annual variability in ecosystem carbon exchange in a Tibetan alpine meadow. J. Plant Ecol. 2022, 15, 882–896. [Google Scholar] [CrossRef]
- Teets, A.; Bailey, A.S.; Hufkens, K.; Ollinger, S.; Schädel, C.; Seyednasrollah, B.; Richardson, A.D. Early spring onset increases carbon uptake more than late fall senescence: Modeling future phenological change in a US northern deciduous forest. Oecologia 2023, 201, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, L.M.; Collins, S.L.; Krofcheck, D.J.; Pockman, W.T. Minimal mortality and rapid recovery of the dominant shrub Larrea tridentata following an extreme cold event in the northern Chihuahuan Desert. J. Veg. Sci. 2019, 30, 963–972. [Google Scholar] [CrossRef]
- Werner, C.M.; Harrison, S.P.; Safford, H.D.; Bohlman, G.N.; Serata, R. Extreme pre-fire drought decreases shrub regeneration on fertile soils. Ecol. Appl. 2022, 32, e02464. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Bloor, J.M.; Kreyling, J.; Ransijn, J.C.; Nijs, I.; Jentsch, A.; Zeiter, M. Patterns and drivers of biodiversity-stability relationships under climate extremes. J. Ecol. 2018, 106, 890–902. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Definition | Unit |
|---|---|---|
| Temperature | ||
| TNn | Minimum value of daily minimum temperature | °C |
| TNx | Maximum value of daily minimum temperature | °C |
| TXn | Minimum value of daily maximum temperature | °C |
| TXx | Maximum value of daily maximum temperature | °C |
| Tn10p | Count of days where TN < 10th percentile | days |
| Tn90p | Count of days where TN > 90th percentile | days |
| Tx10p | Count of days where TX < 10th percentile | days |
| Tx90p | Count of days where TX > 90th percentile | days |
| DTR | Mean value of difference between TX (daily maximum temperature) and TN (daily minimum temperature) | °C |
| Precipitation | ||
| Rx1day | Maximum precipitation amount in one-day period | mm |
| Rx5day | Maximum total precipitation amount in consecutive five-day period | mm |
| Average NEP (g·m−2) | Grass | Shrub | Cropland | Forest | Mosaic Vegetation | Total |
| <−20 | 0.13 | 0.00 | 0.06 | 0.06 | 0.01 | 0.26 |
| −20–−10 | 0.36 | 0.00 | 0.19 | 0.11 | 0.02 | 0.68 |
| −10–0 | 1.73 | 0.00 | 0.75 | 0.49 | 0.09 | 3.06 |
| 0–10 | 22.78 | 0.10 | 3.68 | 1.71 | 0.47 | 28.75 |
| 10–20 | 9.41 | 0.01 | 5.72 | 2.47 | 0.64 | 18.25 |
| 20–30 | 5.35 | 0.01 | 4.68 | 2.93 | 0.60 | 13.58 |
| 30–40 | 2.48 | 0.01 | 2.92 | 3.02 | 0.47 | 8.89 |
| 40–50 | 1.02 | 0.01 | 1.84 | 3.20 | 0.42 | 6.49 |
| 50–60 | 0.52 | 0.02 | 1.54 | 3.34 | 0.45 | 5.86 |
| 60–70 | 0.29 | 0.03 | 1.41 | 3.25 | 0.41 | 5.39 |
| 70–80 | 0.17 | 0.02 | 1.04 | 2.46 | 0.25 | 3.93 |
| 80–90 | 0.09 | 0.01 | 0.62 | 1.59 | 0.13 | 2.44 |
| >90 | 0.11 | 0.01 | 0.54 | 1.67 | 0.11 | 2.43 |
| Trend of NEP (g·m−2·a−2) | Grass | Shrub | Cropland | Forest | Mosaic Vegetation | Total |
| <−20 | 0.11 | 0.00 | 0.20 | 0.25 | 0.01 | 0.56 |
| −20–−10 | 0.32 | 0.00 | 0.59 | 1.08 | 0.03 | 2.03 |
| −10–0 | 5.31 | 0.03 | 2.78 | 4.14 | 0.26 | 12.51 |
| 0–10 | 37.53 | 0.22 | 8.52 | 6.84 | 1.01 | 54.11 |
| 10–20 | 5.48 | 0.04 | 4.96 | 4.66 | 0.83 | 15.98 |
| 20–30 | 1.06 | 0.02 | 2.43 | 2.94 | 0.57 | 7.03 |
| 30–40 | 0.31 | 0.01 | 1.44 | 1.75 | 0.38 | 3.89 |
| 40–50 | 0.09 | 0.01 | 0.92 | 0.96 | 0.27 | 2.25 |
| 50–60 | 0.03 | 0.01 | 0.45 | 0.39 | 0.15 | 1.04 |
| >60 | 0.02 | 0.00 | 0.25 | 0.23 | 0.09 | 0.60 |
| Significance of Trend | Grass | Shrub | Cropland | Forest | Mosaic Vegetation | Total |
| Significant increase | 14.79 | 0.10 | 11.10 | 8.98 | 2.40 | 37.36 |
| Significant decrease | 22.29 | 0.10 | 9.73 | 11.30 | 1.34 | 44.76 |
| Not significant increase | 0.25 | 0.00 | 0.59 | 0.42 | 0.02 | 1.28 |
| Not significant decrease | 6.79 | 0.03 | 3.71 | 5.75 | 0.32 | 16.60 |
| Spring | Grass | Shrub | Cropland | Forest | Mosaic Vegetation | Summer | Grass | Shrub | Cropland | Forest | Mosaic Vegetation |
| DTR | −0.27 | −0.51 * | −0.39 * | −0.51 * | −0.43 * | DTR | 0.02 | −0.11 | −0.36 * | −0.23 | −0.12 |
| TNx | −0.01 | 0.41 * | 0.16 | 0.39 * | 0.12 | TNx | 0.08 | 0.34 * | 0.26 | 0.25 | 0.27 |
| TNn | −0.63 * | −0.26 | −0.47 * | −0.25 | −0.54 * | TNn | −0.67 * | 0.21 | −0.04 | 0.05 | 0.19 |
| TXx | 0.00 | 0.27 | 0.07 | 0.23 | 0.02 | TXx | 0.38 * | 0.25 | 0.13 | 0.12 | 0.22 |
| TXn | −0.65 * | −0.08 | −0.45 * | −0.21 | −0.50 * | TXn | −0.67 * | 0.20 | 0.13 | 0.21 | 0.33 * |
| Tn10p | −0.59 * | −0.39 * | −0.53 * | −0.48 * | −0.62 * | Tn10p | −0.49 * | 0.26 | −0.13 | −0.08 | 0.13 |
| Tx10p | −0.20 | −0.17 | −0.03 | −0.05 | 0.03 | Tx10p | −0.58 * | −0.03 | 0.01 | 0.05 | −0.05 |
| Tx90p | −0.60 * | −0.31 | −0.49 * | −0.46 * | −0.63 * | Tx90p | −0.62 * | 0.27 | −0.17 | −0.09 | 0.11 |
| Tn90p | −0.25 | 0.04 | −0.06 | 0.04 | −0.03 | Tn90p | −0.57 * | 0.00 | 0.03 | 0.10 | −0.02 |
| Rx1day | 0.10 | 0.47 * | 0.17 | 0.32 | 0.21 | Rx1day | 0.59 * | −0.14 | −0.07 | −0.10 | −0.14 |
| Rx5day | 0.07 | 0.46 * | 0.15 | 0.31 | 0.26 | Rx5day | 0.63 * | −0.14 | −0.24 | −0.28 | −0.26 |
| Fall | Grass | Shrub | Cropland | Forest | Mosaic vegetation | Winter | Grass | Shrub | Cropland | Forest | Mosaic vegetation |
| DTR | −0.27 | −0.32 | −0.18 | 0.08 | −0.04 | DTR | −0.27 | −0.65 * | −0.23 | −0.40 * | −0.37 * |
| TNx | 0.46 * | 0.54 * | 0.63 * | 0.65 * | 0.47 * | TNx | 0.34 * | 0.55 * | 0.18 | 0.35 * | 0.17 |
| TNn | 0.46 * | 0.32 | 0.21 | −0.32 | 0.50 * | TNn | 0.20 | −0.09 | −0.06 | 0.27 | 0.34 * |
| TXx | 0.12 | 0.12 | 0.42 * | 0.74 * | 0.12 | TXx | 0.30 | 0.19 | 0.09 | 0.01 | −0.05 |
| TXn | 0.57 * | 0.35 * | 0.18 | −0.45 * | 0.49 * | TXn | 0.22 | 0.16 | 0.03 | 0.29 | 0.44 * |
| Tn10p | 0.23 | 0.20 | 0.19 | −0.26 | 0.41 * | Tn10p | −0.10 | −0.20 | −0.42 * | −0.10 | 0.02 |
| Tx10p | 0.18 | −0.07 | −0.27 | −0.56 * | 0.18 | Tx10p | −0.13 | −0.34 * | 0.10 | 0.08 | 0.24 |
| Tx90p | 0.31 | 0.18 | 0.13 | −0.49 * | 0.36 * | Tx90p | −0.12 | −0.04 | −0.30 | −0.08 | 0.14 |
| Tn90p | 0.35 * | 0.20 | 0.13 | −0.42 * | 0.39 * | Tn90p | −0.10 | −0.15 | 0.03 | 0.12 | 0.23 |
| Rx1day | −0.01 | −0.05 | −0.10 | 0.26 | 0.27 | Rx1day | −0.17 | 0.31 | 0.22 | 0.37 * | 0.14 |
| Rx5day | −0.04 | −0.06 | −0.29 | 0.12 | 0.17 | Rx5day | −0.27 | 0.28 | 0.11 | 0.34 | 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Xu, X.; Zhang, T.; Jiang, H.; Xia, H.; Xu, X.; Xu, J. Multi-Temporal and Time-Lag Responses of Terrestrial Net Ecosystem Productivity to Extreme Climate from 1981 to 2019 in China. Remote Sens. 2024, 16, 163. https://doi.org/10.3390/rs16010163
Huang Y, Xu X, Zhang T, Jiang H, Xia H, Xu X, Xu J. Multi-Temporal and Time-Lag Responses of Terrestrial Net Ecosystem Productivity to Extreme Climate from 1981 to 2019 in China. Remote Sensing. 2024; 16(1):163. https://doi.org/10.3390/rs16010163
Chicago/Turabian StyleHuang, Yiqin, Xia Xu, Tong Zhang, Honglei Jiang, Haoyu Xia, Xiaoqing Xu, and Jiayu Xu. 2024. "Multi-Temporal and Time-Lag Responses of Terrestrial Net Ecosystem Productivity to Extreme Climate from 1981 to 2019 in China" Remote Sensing 16, no. 1: 163. https://doi.org/10.3390/rs16010163
APA StyleHuang, Y., Xu, X., Zhang, T., Jiang, H., Xia, H., Xu, X., & Xu, J. (2024). Multi-Temporal and Time-Lag Responses of Terrestrial Net Ecosystem Productivity to Extreme Climate from 1981 to 2019 in China. Remote Sensing, 16(1), 163. https://doi.org/10.3390/rs16010163