
Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production
 , , ,
, , , 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Carbon Flux
2.2. Remotely Sensed Data
2.3. Clumping Index (CI)
2.4. GPP Models
2.5. Statistical Analyses
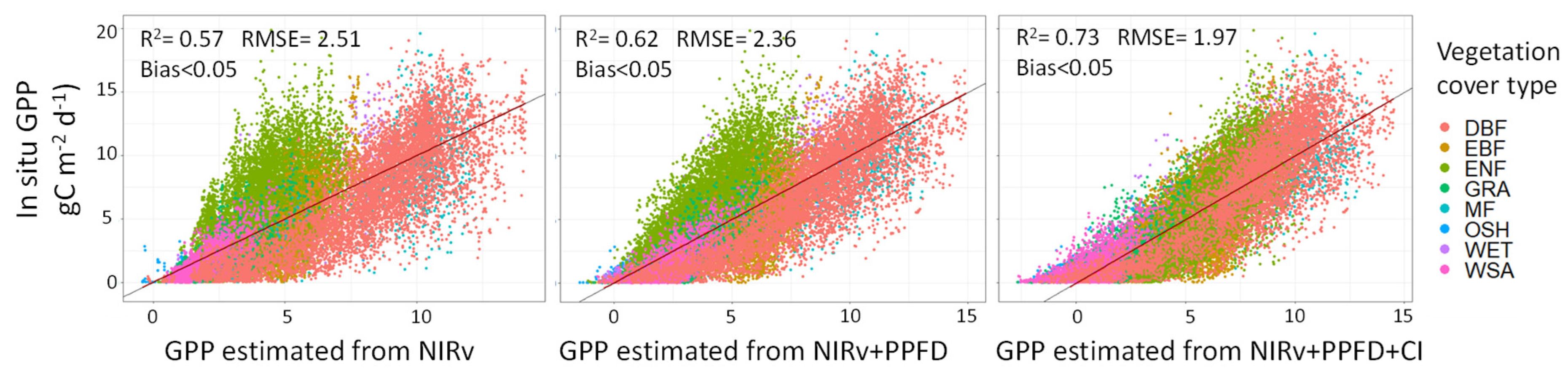
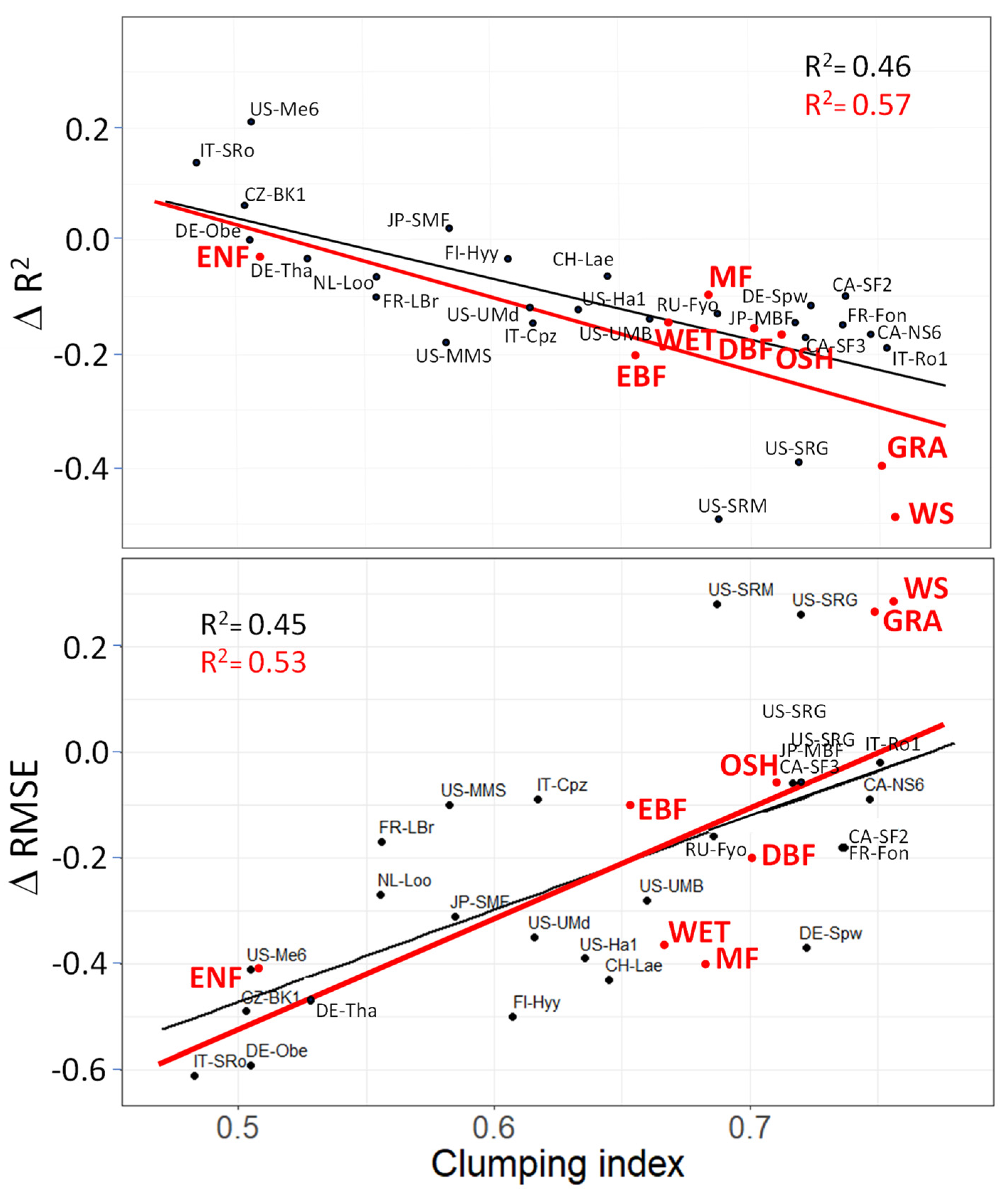
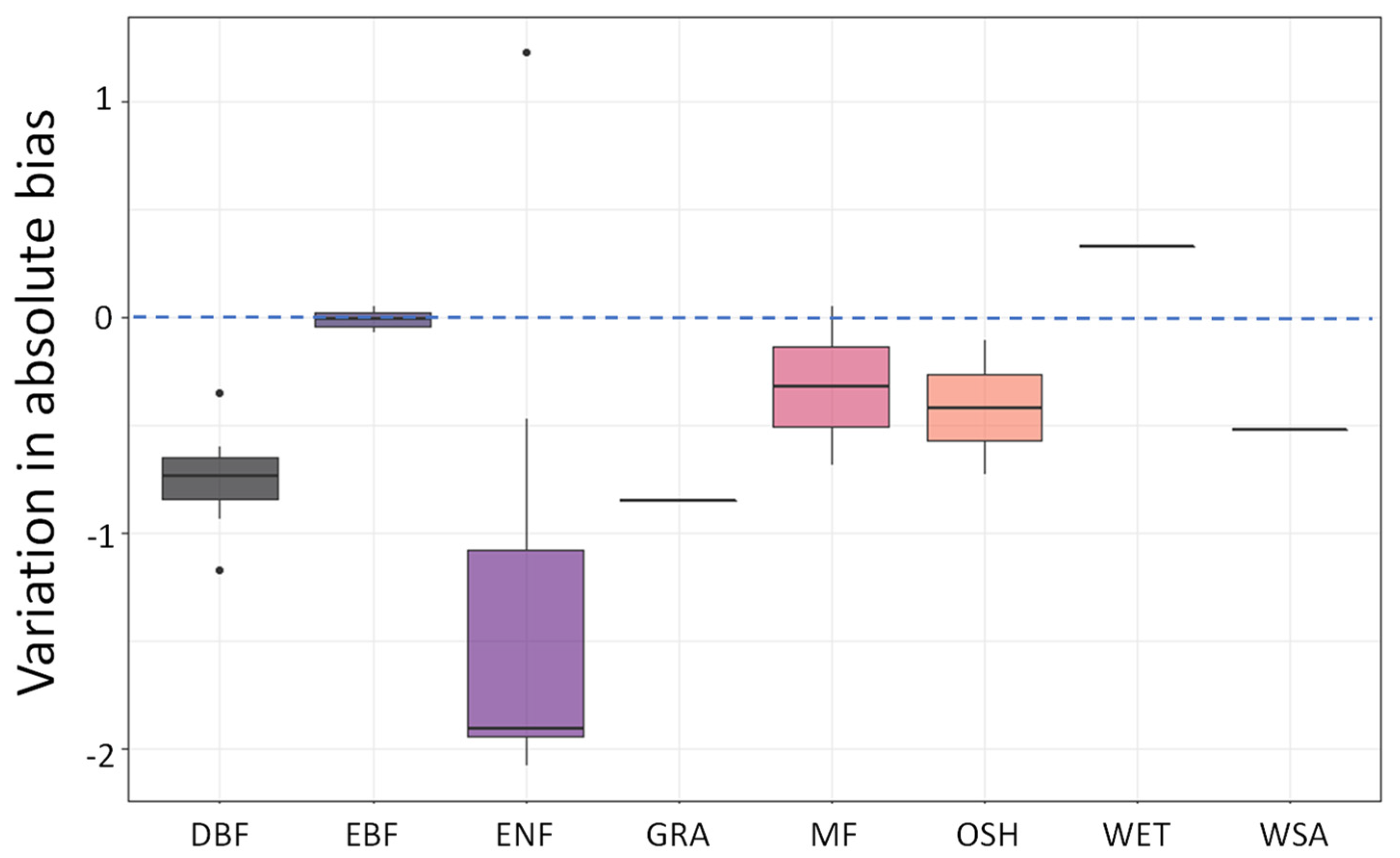
3. Results
4. Discussion
4.1. Adding PPFD and CI to NIRv to Remotely Assess GPP
4.2. Vegetation Cover Types
4.3. Latitudinal Ranges
4.4. Seasonality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Zhang, H.; Liu, Z.; Che, M.; Chen, B. Evaluating Parameter Adjustment in the MODIS Gross Primary Production Algorithm Based on Eddy Covariance Tower Measurements. Remote Sens. 2014, 6, 3321–3348. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tonelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Tian, H.; Dangal, S.R.S.; Ouyang, Z.; Tao, B.; Ren, W.; Lu, C.; Running, S. Modeling and Monitoring Terrestrial Primary Production in a Changing Global Environment: Toward a Multiscale Synthesis of Observation and Simulation. Adv. Meteorol. 2014, 2014, 965936. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.-O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; van Diemen, R.; et al. (Eds.) IPCC Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; 2019. Available online: https://www.ipcc.ch/srccl/ (accessed on 25 February 2023).
- Badgley, G.; Field, C.B.; Berry, J.A. Canopy Near-Infrared Reflectance and Terrestrial Photosynthesis. Sci. Adv. 2017, 3, e1602244. [Google Scholar] [CrossRef]
- Badgley, G.; Anderegg, L.D.L.; Berry, J.A.; Field, C.B. Terrestrial Gross Primary Production: Using NIRV to Scale from Site to Globe. Glob. Chang. Biol. 2019, 25, 3731–3740. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Ryu, Y.; Dechant, B.; Eichelmann, E.; Hemes, K.; Ma, S.; Sanchez, C.R.; Shortt, R.; Szutu, D.; Valach, A.; et al. Outgoing Near-Infrared Radiation From Vegetation Scales With Canopy Photosynthesis Across a Spectrum of Function, Structure, Physiological Capacity, and Weather. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005534. [Google Scholar] [CrossRef]
- Zhang, Y.; Commane, R.; Zhou, S.; Williams, A.P.; Gentine, P. Light Limitation Regulates the Response of Autumn Terrestrial Carbon Uptake to Warming. Nat. Clim. Chang. 2020, 10, 739–743. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-Driven Increases in Global Terrestrial Net Primary Production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef]
- Monteith, J.L. Solar Radiation and Productivity in Tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747. [Google Scholar] [CrossRef]
- Dechant, B.; Ryu, Y.; Badgley, G.; Köhler, P.; Rascher, U.; Migliavacca, M.; Zhang, Y.; Tagliabue, G.; Guan, K.; Rossini, M.; et al. NIRVP: A Robust Structural Proxy for Sun-Induced Chlorophyll Fluorescence and Photosynthesis across Scales. Remote Sens. Environ. 2022, 268, 112763. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgley, G.; Dechant, B.; Ryu, Y.; Chen, M.; Berry, J.A. A Practical Approach for Estimating the Escape Ratio of Near-Infrared Solar-Induced Chlorophyll Fluorescence. Remote Sens. Environ. 2019, 232, 111209. [Google Scholar] [CrossRef]
- Khan, A.M.; Stoy, P.C.; Joiner, J.; Baldocchi, D.; Verfaillie, J.; Chen, M.; Otkin, J.A. The Diurnal Dynamics of Gross Primary Productivity Using Observations From the Advanced Baseline Imager on the Geostationary Operational Environmental Satellite-R Series at an Oak Savanna Ecosystem. J. Geophys. Res. Biogeosci. 2022, 127, e2021JG006701. [Google Scholar] [CrossRef]
- Kong, J.; Ryu, Y.; Liu, J.; Dechant, B.; Rey-Sanchez, C.; Shortt, R.; Szutu, D.; Verfaillie, J.; Houborg, R.; Baldocchi, D.D. Matching High Resolution Satellite Data and Flux Tower Footprints Improves Their Agreement in Photosynthesis Estimates. Agric. For. Meteorol. 2022, 316, 108878. [Google Scholar] [CrossRef]
- Jiang, C.; Guan, K.; Wu, G.; Peng, B.; Wang, S. A Daily, 250 m and Real-Time Gross Primary Productivity Product (2000–Present) Covering the Contiguous United States. Earth Syst. Sci. Data 2021, 13, 281–298. [Google Scholar] [CrossRef]
- van Dijk, A.I.J.M.; Dolman, A.J.; Schulze, E.D. Radiation, Temperature, and Leaf Area Explain Ecosystem Carbon Fluxes in Boreal and Temperate European Forests. Glob. Biogeochem. Cycles 2005, 19, 1–15. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Yamamoto, H.; Tani, H.; Zhong, G.; Yin, S.; Guo, E. Spatial Pattern of GPP Variations in Terrestrial Ecosystems and Its Drivers: Climatic Factors, CO2 Concentration and Land-Cover Change, 1982–2015. Ecol. Inform. 2018, 46, 156–165. [Google Scholar] [CrossRef]
- Mäkelä, A.; Kolari, P.; Karimäki, J.; Nikinmaa, E.; Perämäki, M.; Hari, P. Modelling Five Years of Weather-Driven Variation of GPP in a Boreal Forest. Agric. For. Meteorol. 2006, 139, 382–398. [Google Scholar] [CrossRef]
- Vesala, T.; Launiainen, S.; Kolari, P.; Pumpanen, J.; Sevanto, S.; Hari, P.; Nikinmaa, E.; Kaski, P.; Mannila, H.; Ukkonen, E.; et al. Autumn Temperature and Carbon Balance of a Boreal Scots Pine Forest in Southern Finland. Biogeosciences 2010, 7, 163–176. [Google Scholar] [CrossRef]
- Descals, A.; Verger, A.; Yin, G.; Filella, I.; Fu, Y.H.; Piao, S.; Janssens, I.A.; Peñuelas, J. Radiation-constrained Boundaries Cause Nonuniform Responses of the Carbon Uptake Phenology to Climatic Warming in the Northern Hemisphere. Glob. Chang. Biol. 2022, 29, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Stoy, P.C.; Luo, Y.; Chen, J.; Sun, J.; Montagnani, L.; Wohlfahrt, G.; Rahman, A.F.; Rambal, S.; Bernhofer, C.; et al. Climate Controls over the Net Carbon Uptake Period and Amplitude of Net Ecosystem Production in Temperate and Boreal Ecosystems. Agric. For. Meteorol. 2017, 243, 9–18. [Google Scholar] [CrossRef]
- He, L.; Chen, J.M.; Pisek, J.; Schaaf, C.B.; Strahler, A.H. Global Clumping Index Map Derived from the MODIS BRDF Product. Remote Sens. Environ. 2012, 119, 118–130. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A. Defining Leaf Area Index for Non-Flat Leaves. Plant Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Chen, J.M.; Mo, G.; Pisek, J.; Liu, J.; Deng, F.; Ishizawa, M.; Chan, D. Effects of Foliage Clumping on the Estimation of Global Terrestrial Gross Primary Productivity. Glob. Biogeochem. Cycles 2012, 26, 1019. [Google Scholar] [CrossRef]
- Lacaze, R.; Chen, J.M.; Roujean, J.-L.; Leblanc, S.G. Retrieval of Vegetation Clumping Index Using Hot Spot Signatures Measured by POLDER Instrument. Remote Sens. Environ. 2002, 79, 84–95. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Porcar-Castell, A.; Joiner, J.; Guanter, L.; Yang, X.; Migliavacca, M.; Ju, W.; Sun, Z.; Chen, S.; et al. Reduction of Structural Impacts and Distinction of Photosynthetic Pathways in a Global Estimation of GPP from Space-Borne Solar-Induced Chlorophyll Fluorescence. Remote Sens. Environ. 2020, 240, 111722. [Google Scholar] [CrossRef]
- Pastorello, G.; Trotta, C.; Canfora, E.; Chu, H.; Christianson, D.; Cheah, Y.W.; Poindexter, C.; Chen, J.; Elbashandy, A.; Humphrey, M.; et al. The FLUXNET2015 Dataset and the ONEFlux Processing Pipeline for Eddy Covariance Data. Sci. Data 2020, 7, 225. [Google Scholar] [CrossRef]
- Fang, H. Canopy Clumping Index (CI): A Review of Methods, Characteristics, and Applications. Agric. For. Meteorol. 2021, 303, 108374. [Google Scholar] [CrossRef]
- Chen, J.M.; Menges, C.H.; Leblanc, S.G. Global Mapping of Foliage Clumping Index Using Multi-Angular Satellite Data. Remote Sens. Environ. 2005, 97, 447–457. [Google Scholar] [CrossRef]
- Wei, S.; Fang, H. Estimation of Canopy Clumping Index from MISR and MODIS Sensors Using the Normalized Difference Hotspot and Darkspot (NDHD) Method: The Influence of BRDF Models and Solar Zenith Angle. Remote Sens. Environ. 2016, 187, 476–491. [Google Scholar] [CrossRef]
- Fang, H.; Wei, S. Global Vegetation Clumping Index Products (8-Days,500 m) (LIS-CI-A1) from 2001–2016; PANGAEA Data Publisher for Earth & Environmental Science: Bremen/Bremerhaven, Germany, 2018. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2015, 1, 1651. [Google Scholar]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating Resource Selection Functions. Ecol. Modell. 2002, 157, 281–300. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP Estimates from NDVI for the Central Grassland Region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Knapp, A.K.; Briggs, J.M.; Koelliker, J.K. Frequency and Extent of Water Limitation to Primary Production in a Mesic Temperate Grassland. Ecosystems 2001, 4, 19–28. [Google Scholar] [CrossRef]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary Production of the Central Grassland Region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Sellers, P.J.; Berry, J.A.; Collatz, G.J.; Field, C.B.; Hall, E.G. Canopy Reflectance, Photosynthesis, and Transpiration. III. A Reanalysis Using Improved Leaf Models and a New Canopy Integration Scheme. Remote Sens. Environ. 1992, 42, 187–216. [Google Scholar] [CrossRef]
- Liu, X.; Liu, L. Influence of the Canopy BRDF Characteristics and Illumination Conditions on the Retrieval of Solar-Induced Chlorophyll Fluorescence. Int. J. Remote Sens. 2018, 39, 1782–1799. [Google Scholar] [CrossRef]
- Romero, J.M.; Cordon, G.B.; Lagorio, M.G. Re-Absorption and Scattering of Chlorophyll Fluorescence in Canopies: A Revised Approach. Remote Sens. Environ. 2020, 246, 111860. [Google Scholar] [CrossRef]
- Churkina, G.; Running, S.W. Contrasting Climatic Controls on the Estimated Productivity of Global Terrestrial Biomes. Ecosystems 1998, 1, 206–215. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Vitousek, P.M. Principles of Terrestrial Ecosystem Ecology; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Asner, G.P.; Scurlock, J.M.O.; Hicke, J.A. Global Synthesis of Leaf Area Index Observations. Glob. Chang. Biol. 2003, 12, 191–205. [Google Scholar] [CrossRef]
- Liu, Z.; Jin, G.; Qi, Y. Estimate of Leaf Area Index in an Old-Growth Mixed Broadleaved-Korean Pine Forest in Northeastern China. PLoS ONE 2012, 7, e32155. [Google Scholar] [CrossRef]
- Behera, S.K.; Srivastava, P.; Pathre, U.V.; Tuli, R. An Indirect Method of Estimating Leaf Area Index in Jatropha Curcas L. Using LAI-2000 Plant Canopy Analyzer. Agric. For. Meteorol. 2010, 150, 307–311. [Google Scholar] [CrossRef]
- Sonnentag, O.; Chen, J.M.; Roberts, D.A.; Talbot, J.; Halligan, K.Q.; Govind, A. Mapping Tree and Shrub Leaf Area Indices in an Ombrotrophic Peatland through Multiple Endmember Spectral Unmixing. Remote Sens. Environ. 2007, 109, 342–360. [Google Scholar] [CrossRef]
- Sprintsin, M.; Karnieli, A.; Berliner, P.; Rotenberg, E.; Yakir, D.; Cohen, S. The Effect of Spatial Resolution on the Accuracy of Leaf Area Index Estimation for a Forest Planted in the Desert Transition Zone. Remote Sens. Environ. 2007, 109, 416–428. [Google Scholar] [CrossRef]
- Chen, J.M.; Leblanc, S.G. Four-Scale Bidirectional Reflectance Model Based on Canopy Architecture. IEEE Trans. Geosci. Remote Sens. 1997, 35, 1316–1337. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud Cover Limits Net CO2 Uptake and Growth of a Rainforest Tree during Tropical Rainy Seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef]
- Fan, S.-M.; Wofsy, S.C.; Bakwin, P.S.; Jacob, D.J.; Fitzjarrald, D.R. Atmosphere-Biosphere Exchange of CO2 and O3 in the Central Amazon Forest. J. Geophys. Res. 1990, 95, 16851–16864. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef]
- Niu, S.; Luo, Y.; Fei, S.; Montagnani, L.; Bohrer, G.; Janssens, I.A.; Gielen, B.; Rambal, S.; Moors, E.; Matteucci, G. Seasonal Hysteresis of Net Ecosystem Exchange in Response to Temperature Change: Patterns and Causes. Glob. Chang. Biol. 2011, 17, 3102–3114. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
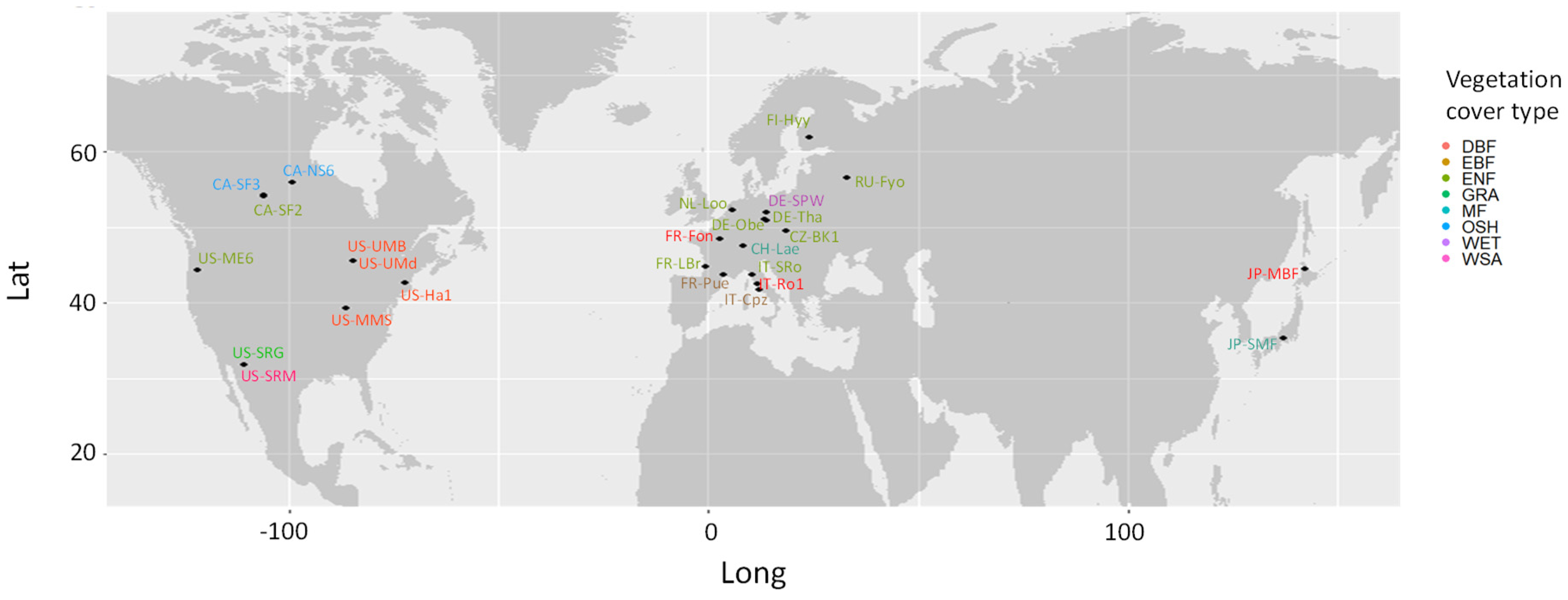
| Site ID | Latitude (°) | Longitude (°) | Vegetation Cover Type | Time Period |
|---|---|---|---|---|
| CA-NS6 | 55.9 | −99.0 | OSH | 2001–2005 |
| CA-SF2 | 54.3 | −105.9 | ENF | 2001–2005 |
| CA-SF3 | 54.1 | −106.0 | OSH | 2001–2006 |
| CH-Lae | 47.5 | 8.4 | MF | 2004–2014 |
| CZ-BK1 | 49.5 | 18.5 | ENF | 2004–2014 |
| DE-Obe | 50.8 | 13.7 | ENF | 2009–2014 |
| DE-Spw | 51.9 | 14.0 | WET | 2010–2014 |
| DE-Tha | 51.0 | 13.6 | ENF | 2000–2014 |
| FI-Hyy | 61.8 | 24.3 | ENF | 2000–2014 |
| FR-Fon | 48.5 | 2.8 | DBF | 2005–2014 |
| FR-LBr | 44.7 | −0.8 | ENF | 2000–2008 |
| FR-Pue | 43.7 | 3.6 | EBF | 2000–2014 |
| IT-Cpz | 41.7 | 12.4 | EBF | 2000–2008 |
| IT-Ro1 | 42.4 | 11.9 | DBF | 2000–2008 |
| IT-SRo | 43.7 | 10.3 | ENF | 2000–2012 |
| JP-MBF | 44.4 | 142.3 | DBF | 2004–2005 |
| JP-SMF | 35.3 | 137.1 | MF | 2005–2006 |
| NL-Loo | 52.2 | 5.7 | ENF | 2005–2014 |
| RU-Fyo | 56.5 | 32.9 | ENF | 2000–2014 |
| US-Ha1 | 42.5 | −72.2 | DBF | 2007–2009 |
| US-Me6 | 44.3 | −121.6 | ENF | 2000–2012 |
| US-MMS | 39.3 | −86.4 | DBF | 2011–2014 |
| US-SRG | 31.8 | −110.8 | GRA | 2000–2014 |
| US-SRM | 31.8 | −110.9 | WSA | 2008–2014 |
| US-UMB | 45.6 | −84.7 | DBF | 2004–2014 |
| US-UMd | 45.6 | −84.7 | DBF | 2000–2014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filella, I.; Descals, A.; Balzarolo, M.; Yin, G.; Verger, A.; Fang, H.; Peñuelas, J. Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production. Remote Sens. 2023, 15, 2207. https://doi.org/10.3390/rs15082207
Filella I, Descals A, Balzarolo M, Yin G, Verger A, Fang H, Peñuelas J. Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production. Remote Sensing. 2023; 15(8):2207. https://doi.org/10.3390/rs15082207
Chicago/Turabian StyleFilella, Iolanda, Adrià Descals, Manuela Balzarolo, Gaofei Yin, Aleixandre Verger, Hongliang Fang, and Josep Peñuelas. 2023. "Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production" Remote Sensing 15, no. 8: 2207. https://doi.org/10.3390/rs15082207
APA StyleFilella, I., Descals, A., Balzarolo, M., Yin, G., Verger, A., Fang, H., & Peñuelas, J. (2023). Photosynthetically Active Radiation and Foliage Clumping Improve Satellite-Based NIRv Estimates of Gross Primary Production. Remote Sensing, 15(8), 2207. https://doi.org/10.3390/rs15082207