
Comparing the Use of Red-Edge and Near-Infrared Wavelength Ranges for Detecting Submerged Kelp Canopy

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
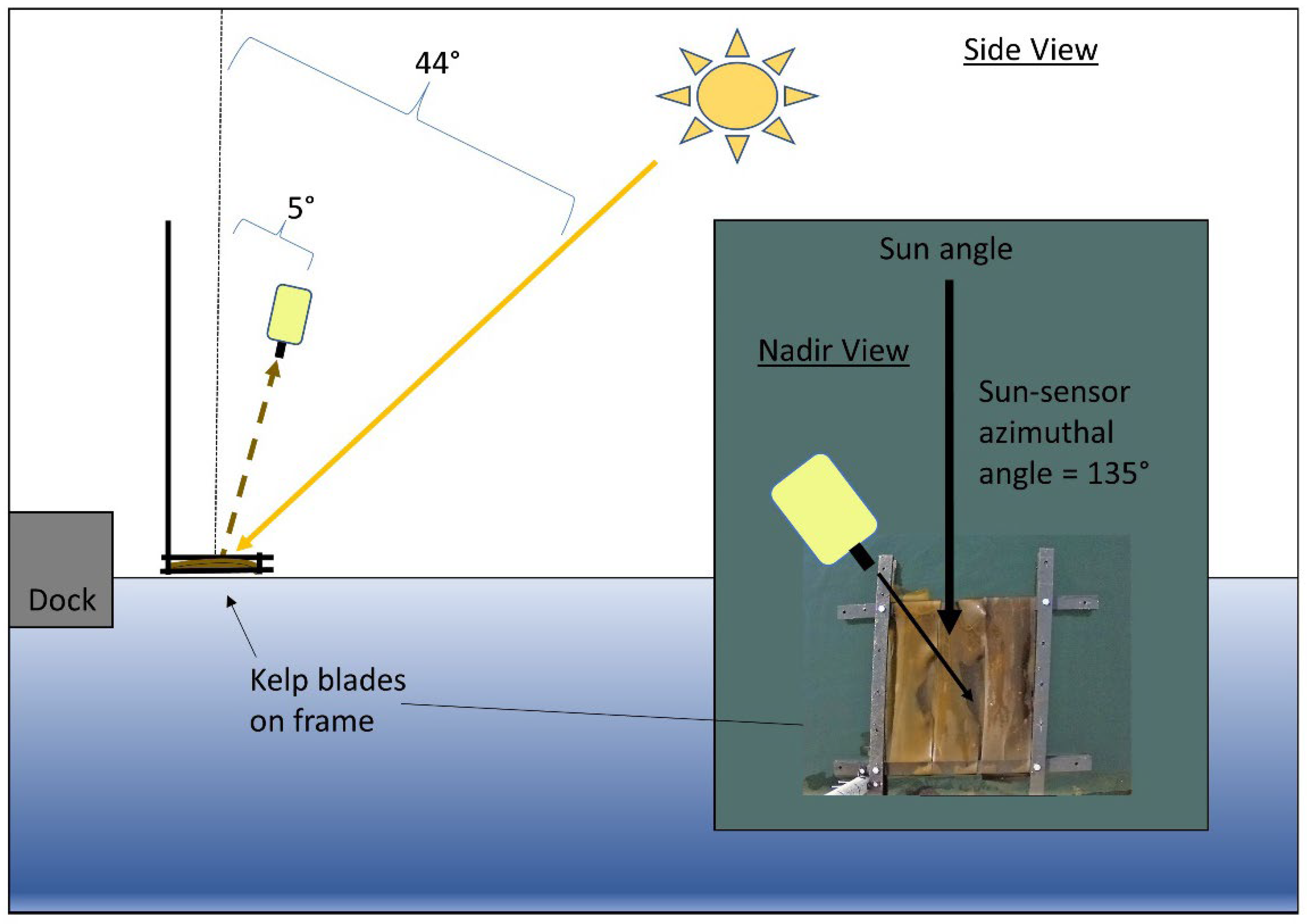
2.1. Spectral Data Acquisition and Processing
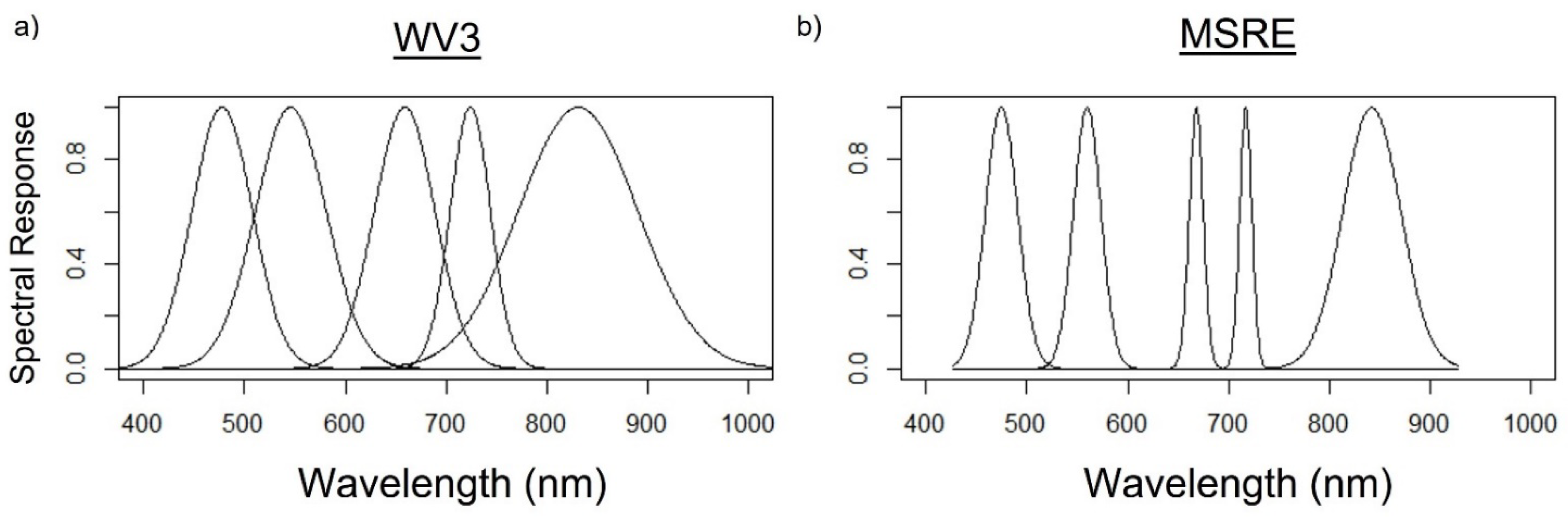
2.2. Simulation of Micasense and WorldView Band R0+ and Indices
2.3. Normalized Vegetation Indices
2.4. Threshold Selection and Depth Limits for Kelp Detection
3. Results
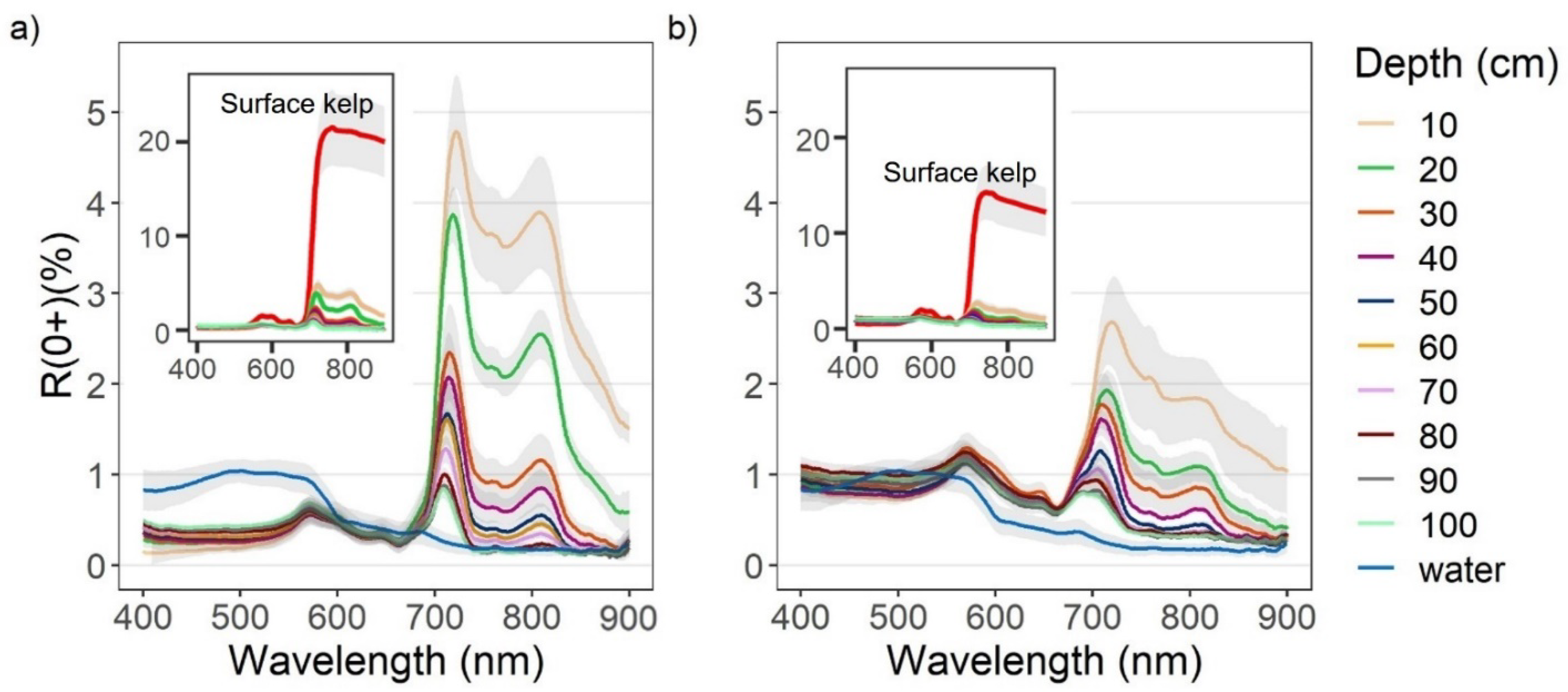
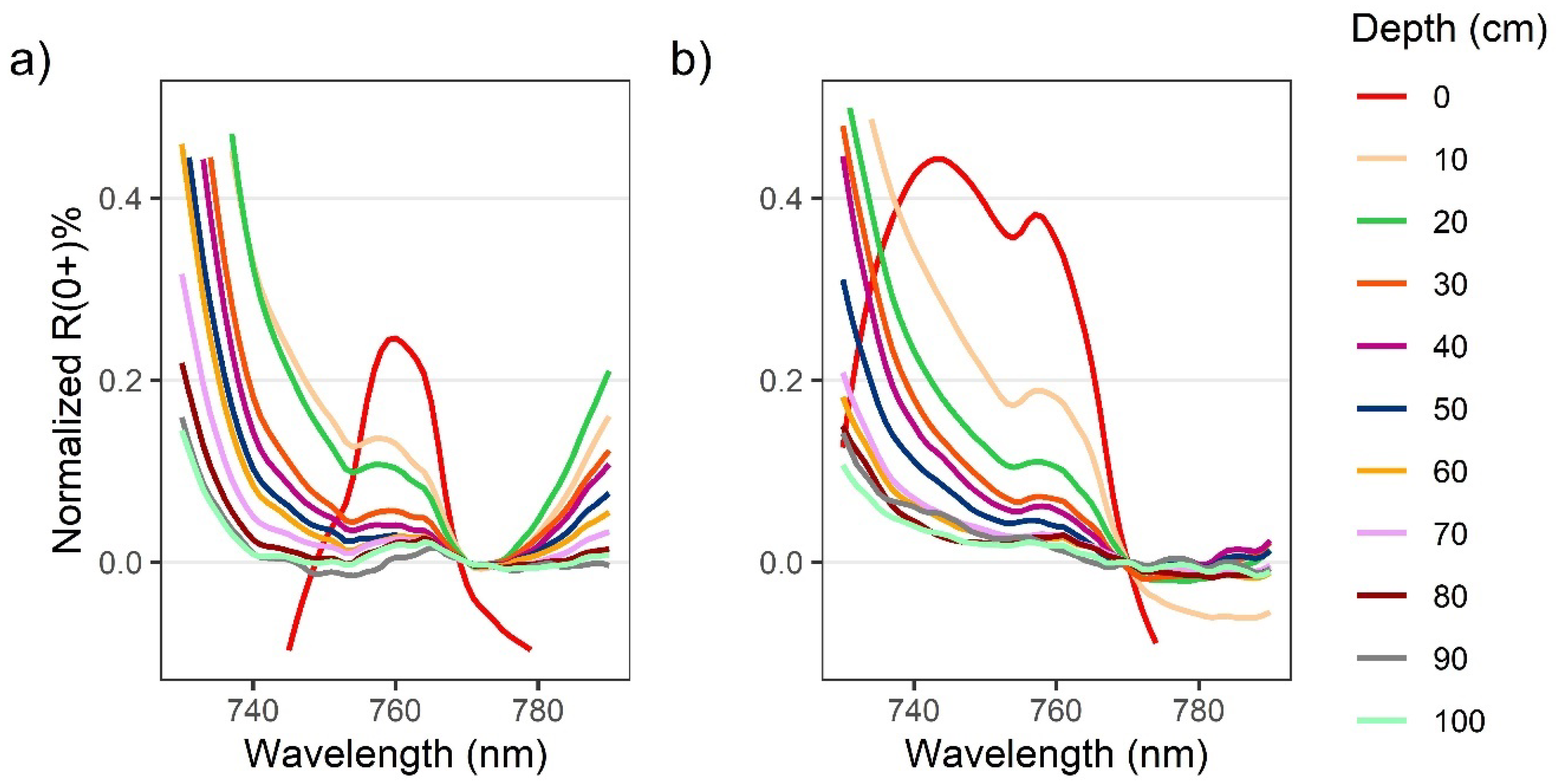
3.1. Spectral Characteristics of Surface and Submerged Kelp
3.2. Vegetation Indices: Signal Strength and Depth-Detection Limits of Submerged Kelp
4. Discussion
4.1. Spectral Characteristics of Kelp as It Is Submerged
4.2. NIR Differences between Nereocystis Bulbs and Blades
4.3. The Implications of VIn Saturation for Detection of Floating and Submerged Kelp
4.4. Depth Detection Limits and Separability between Kelp and Water
4.5. Implications for Mixed Pixels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Druehl, L.D.; Wheeler, W.N. Population Biology of Macrocystis Integrifolia from British Columbia, Canada. Mar. Biol. 1986, 90, 173–179. [Google Scholar] [CrossRef]
- Kain, J.M. Patterns of Relative Growth in Nereocystis Luetkeana (Phaeophyta). J. Phycol. 1987, 23, 181–187. [Google Scholar] [CrossRef]
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global Patterns of Kelp Forest Change over the Past Half-Century. Proc. Natl. Acad. Sci. USA 2016, 113, 13785–13790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druehl, L.D. The Pattern of Laminariales Distribution in the Northeast Pacific. Phycologia 1970, 9, 237–247. [Google Scholar] [CrossRef]
- Jackson, G.A. Internal Wave Attenuation by Coastal Kelp Stands. J. Phys. Oceanogr. 1984, 14, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Mork, M. The Effect of Kelp in Wave Damping. Sarsia 1996, 80, 323–327. [Google Scholar] [CrossRef]
- Krumhansl, K.A.; Scheibling, R. Production and Fate of Kelp Detritus. Mar. Ecol. Prog. Ser. 2012, 467, 281–302. [Google Scholar] [CrossRef] [Green Version]
- Olson, A.M.; Hessing-Lewis, M.; Haggarty, D.; Juanes, F. Nearshore Seascape Connectivity Enhances Seagrass Meadow Nursery Function. Ecol. Appl. 2019, 29, e01897. [Google Scholar] [CrossRef]
- Springer, Y.; Hays, C.; Carr, M.H.; Mackey, M. Ecology and Management of the Bull Kelp, Nereocystis Luetkeana: A Synthesis with Recommendations for Future Research; Lenfest Ocean Program: Washington, DC, USA, 2007; pp. 1–53. [Google Scholar]
- Stekoll, M.S.; Deysher, L.E.; Hess, M. A Remote Sensing Approach to Estimating Harvestable Kelp Biomass. J. Appl. Phycol. 2006, 18, 323–334. [Google Scholar] [CrossRef]
- Bell, T.W.; Allen, J.G.; Cavanaugh, K.C.; Siegel, D.A. Three Decades of Variability in California’s Giant Kelp Forests from the Landsat Satellites. Remote Sens. Environ. 2020, 238, 110811. [Google Scholar] [CrossRef]
- Pfister, C.A.; Berry, H.D.; Mumford, T. The Dynamics of Kelp Forests in the Northeast Pacific Ocean and the Relationship with Environmental Drivers. J. Ecol. 2017, 106, 1520–1533. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Bell, T.; Costa, M.; Eddy, N.E.; Gendall, L.; Gleason, M.G.; Hessing-Lewis, M.; Martone, R.; McPherson, M.; Pontier, O.; et al. A Review of the Opportunities and Challenges for Using Remote Sensing for Management of Surface-Canopy Forming Kelps. Front. Mar. Sci. 2021, 8, 1536. [Google Scholar] [CrossRef]
- Jensen, J.R. Remote Sensing Techniques for Kelp Surveys. Photogramm. Eng. Remote Sens. 1980, 46, 743–755. [Google Scholar]
- Cavanaugh, K.; Siegel, D.; Kinlan, B.; Reed, D. Scaling Giant Kelp Field Measurements to Regional Scales Using Satellite Observations. Mar. Ecol. Prog. Ser. 2010, 403, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, S.B.; Dupont, C.; Boyer, L.; Juanes, F.; Costa, M. Passive Remote Sensing Technology for Mapping Bull Kelp (Nereocystis Luetkeana): A Review of Techniques and Regional Case Study. Glob. Ecol. Conserv. 2019, 19, e00683. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Cavanaugh, K.C.; Bell, T.W.; Hockridge, E.G. An Automated Method for Mapping Giant Kelp Canopy Dynamics from UAV. Front. Environ. Sci. 2021, 8, 587354. [Google Scholar] [CrossRef]
- Schroeder, S.B.; Boyer, L.; Juanes, F.; Costa, M. Spatial and Temporal Persistence of Nearshore Kelp Beds on the West Coast of British Columbia, Canada Using Satellite Remote Sensing. Remote Sens. Ecol. Conserv. 2019, 6, 327–343. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, S.L.; Bell, T.W.; Watson, J.R.; Grorud-Colvert, K.A.; Menge, B.A. Remote Sensing: Generation of Long-Term Kelp Bed Data Sets for Evaluation of Impacts of Climatic Variation. Ecology 2020, 101, e03031. [Google Scholar] [CrossRef]
- Britton-Simmons, K.; Eckman, J.E.; Duggins, D.O. Effect of Tidal Currents and Tidal Stage on Estimates of Bed Size in the Kelp Nereocystis Luetkeana. Mar. Ecol. Prog. Ser. 2008, 355, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Filella, I.; Penuelas, J. The Red Edge Position and Shape as Indicators of Plant Chlorophyll Content, Biomass and Hydric Status. Int. J. Remote Sens. 1994, 15, 1459–1470. [Google Scholar] [CrossRef]
- Pegau, W.S.; Gray, D.; Zaneveld, J.R.V. Absorption and Attenuation of Visible and Near-Infrared Light in Water: Dependence on Temperature and Salinity. Appl. Opt. 1997, 36, 6035–6046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Rundquist, D.C. The Spectral Responses of Ceratophyllum Demersum at Varying Depths in an Experimental Tank. Int. J. Remote Sens. 2003, 24, 859–864. [Google Scholar] [CrossRef]
- Kearney, M.S.; Stutzer, D.; Turpie, K.; Stevenson, J.C. The Effects of Tidal Inundation on the Reflectance Characteristics of Coastal Marsh Vegetation. J. Coast. Res. 2009, 256, 1177–1186. [Google Scholar] [CrossRef]
- Turpie, K.R. Explaining the Spectral Red-Edge Features of Inundated Marsh Vegetation. J. Coast. Res. 2013, 290, 1111–1117. [Google Scholar] [CrossRef]
- Song, B.; Park, K. Detection of Aquatic Plants Using Multispectral UAV Imagery and Vegetation Index. Remote Sens. 2020, 12, 387. [Google Scholar] [CrossRef] [Green Version]
- Nijland, W.; Reshitnyk, L.; Rubidge, E. Satellite Remote Sensing of Canopy-Forming Kelp on a Complex Coastline: A Novel Procedure Using the Landsat Image Archive. Remote Sens. Environ. 2019, 220, 41–50. [Google Scholar] [CrossRef]
- Phillips, S.R.; Costa, M. Spatial-Temporal Bio-Optical Classification of Dynamic Semi-Estuarine Waters in Western North America. Estuar. Coast. Shelf Sci. 2017, 199, 35–48. [Google Scholar] [CrossRef]
- Mobley, C.D. Light and Water: Radiative Transfer in Natural Waters; Academic Press: San Diego, CA, USA, 1994; ISBN 978-0-12-502750-2. [Google Scholar]
- Dierssen, H.M.; Chlus, A.; Russell, B. Hyperspectral Discrimination of Floating Mats of Seagrass Wrack and the Macroalgae Sargassum in Coastal Waters of Greater Florida Bay Using Airborne Remote Sensing. Remote Sens. Environ. 2015, 167, 247–258. [Google Scholar] [CrossRef]
- Hu, L.; Hu, C.; Ming-Xia, H. Remote Estimation of Biomass of Ulva Prolifera Macroalgae in the Yellow Sea. Remote Sens. Environ. 2017, 192, 217–227. [Google Scholar] [CrossRef]
- ASD Inc. ASD Light Theory and Measurement Using the FieldSpec HandHeld 2 Portable Spectroradiometer; ASD Inc.: Longmont, CO, USA, 2017. [Google Scholar]
- Mobley, C.D. Estimation of the Remote-Sensing Reflectance from above-Surface Measurements. Appl. Opt. 1999, 38, 7442–7455. [Google Scholar] [CrossRef]
- Mount, R. Acquisition of Through-Water Aerial Survey Images. Available online: https://www.ingentaconnect.com/content/asprs/pers/2005/00000071/00000012/art00005 (accessed on 8 April 2020).
- Cavanaugh, K.; Siegel, D.; Reed, D.; Dennison, P. Environmental Controls of Giant-Kelp Biomass in the Santa Barbara Channel, California. Mar. Ecol. Prog. Ser. 2011, 429, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Micasense RedEdge-MX—MicaSense. Available online: https://micasense.com/rededge-mx/ (accessed on 17 January 2022).
- Maxar WorldView-3. Available online: https://resources.maxar.com/data-sheets/worldview-3 (accessed on 17 January 2022).
- Rouse, W.; Haas, R.H.; Deering, W. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; Goddard Space Flight Center: Greenbelt, MD, USA, 1974; p. 87.
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Augenstein, E.W.; Stow, D.; Hope, A. Evaluation of SPOT HRV-XS Data for Kelp Resource Inventories. Photogramm. Eng. Remote Sens. 1991, 57, 501–509. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a Green Channel in Remote Sensing of Global Vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Mora-Soto, A.; Palacios, M.; Macaya, E.C.; Gómez, I.; Huovinen, P.; Pérez-Matus, A.; Young, M.; Golding, N.; Toro, M.; Yaqub, M.; et al. A High-Resolution Global Map of Giant Kelp (Macrocystis Pyrifera) Forests and Intertidal Green Algae (Ulvophyceae) with Sentinel-2 Imagery. Remote Sens. 2020, 12, 694. [Google Scholar] [CrossRef] [Green Version]
- Schultz, B.B. Levene’s Test for Relative Variation. Syst. Biol. 1985, 34, 449–456. [Google Scholar] [CrossRef]
- Algina, J.; Oshima, T.C.; Lin, W.-Y. Type I Error Rates for Welch’s Test and James’s Second-Order Test Under Nonnormality and Inequality of Variance When There Are Two Groups. J. Educ. Stat. 1994, 19, 275–291. [Google Scholar] [CrossRef] [Green Version]
- Shingala, M.C.; Rajyaguru, D.A. Comparison of Post Hoc Tests for Unequal Variance. Int. J. New Technol. Sci. Eng. 2015, 2, 12. [Google Scholar]
- Morel, A.; Prieur, L. Analysis of Variations in Ocean Color. Limnol. Oceanogr. 1977, 22, 709–722. [Google Scholar] [CrossRef]
- Vahtmäe, E.; Paavel, B.; Kutser, T. How Much Benthic Information Can Be Retrieved with Hyperspectral Sensor from the Optically Complex Coastal Waters? J. Appl. Remote Sens. 2020, 14, 016504. [Google Scholar] [CrossRef]
- Welch, B.L. The Generalization of ‘Student’s’ Problem When Several Different Population Variances Are Involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef]
- Wheeler, W.N.; Smith, R.G.; Srivastava, L.M. Seasonal Photosynthetic Performance of Nereocystis Luetkeana. Can. J. Bot. 1984, 62, 664–670. [Google Scholar] [CrossRef]
- Olmedo-Masat, O.M.; Raffo, M.P.; Rodríguez-Pérez, D.; Arijón, M.; Sánchez-Carnero, N. How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia). Remote Sens. 2020, 12, 3870. [Google Scholar] [CrossRef]
- Liew, O.W.; Chong, P.C.J.; Li, B.; Asundi, A.K. Signature Optical Cues: Emerging Technologies for Monitoring Plant Health. Sensors 2008, 8, 3205–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruru, D.; Yingqing, H.; Yan, Q.; Qidong, C.; Lei, C. Measuring Pure Water Absorption Coefficient in the Near-Infrared. J. Remote Sens. 2012, 16, 192–206. [Google Scholar]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote Sensing of Solar-Induced Chlorophyll Fluorescence: Review of Methods and Applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Lu, Y.; Li, L.; Hu, C.; Li, L.; Zhang, M.; Sun, S.; Lv, C. Sunlight Induced Chlorophyll Fluorescence in the Near-Infrared Spectral Region in Natural Waters: Interpretation of the Narrow Reflectance Peak around 761 Nm. J. Geophys. Res. Ocean. 2016, 121, 5017–5029. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.B. Physical and Physiological Basis for the Reflectance of Visible and Near-Infrared Radiation from Vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Nicholson, N.L. Field Studies on the Giant Kelp. Nereocystis. J. Phycol. 1970, 6, 177–182. [Google Scholar] [CrossRef]
- Nicholson, N.L. Anatomy of the Medulla of Nereocystis. Bot. Mar. 1976, 19, 23–31. [Google Scholar] [CrossRef]
- Schmitz, K.; Srivastava, L.M. The Fine Structure of Sieve Elements of Nereocystis Lütkeana. Am. J. Bot. 1976, 63, 679–693. [Google Scholar] [CrossRef]
- Nicholson, N.L.; Briggs, W.R. Translocation of Photosynthate in the Brown Alga Nereocystis. Am. J. Bot. 1972, 59, 97–106. [Google Scholar] [CrossRef]
- Slaton, M.R.; Hunt, E.R.; Smith, W.K. Estimating Near-Infrared Leaf Reflectance from Leaf Structural Characteristics. Am. J. Bot. 2001, 88, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Mutanga, O.; Skidmore, A.K. Hyperspectral Band Depth Analysis for a Better Estimation of Grass Biomass (Cenchrus Ciliaris) Measured under Controlled Laboratory Conditions. Int. J. Appl. Earth Obs. Geoinf. 2004, 5, 87–96. [Google Scholar] [CrossRef]
- Gower, J.F.R.; Doerffer, R.; Borstad, G.A. Interpretation of the 685nm Peak in Water-Leaving Radiance Spectra in Terms of Fluorescence, Absorption and Scattering, and Its Observation by MERIS. Int. J. Remote Sens. 1999, 20, 1771–1786. [Google Scholar] [CrossRef]
- O’Neill, J.D.; Costa, M.; Sharma, T. Remote Sensing of Shallow Coastal Benthic Substrates: In Situ Spectra and Mapping of Eelgrass (Zostera Marina) in the Gulf Islands National Park Reserve of Canada. Remote Sens. 2011, 3, 975–1005. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J.D.; Costa, M. Mapping Eelgrass (Zostera Marina) in the Gulf Islands National Park Reserve of Canada Using High Spatial Resolution Satellite and Airborne Imagery. Remote Sens. Environ. 2013, 133, 152–167. [Google Scholar] [CrossRef]
- Finger, D.J.I.; McPherson, M.L.; Houskeeper, H.F.; Kudela, R.M. Mapping Bull Kelp Canopy in Northern California Using Landsat to Enable Long-Term Monitoring. Remote Sens. Environ. 2021, 254, 112243. [Google Scholar] [CrossRef]
- Sutherland, I.R. Kelp Inventory, 1989 The Vancouver Island and Malcolm Island Shores of Queen Charlotte Strait Including a Summary of Historical Inventory Information for the Area; Kelp Inventory; Ministry of Environment: Vancouver, BC, Canada, 1990; p. 46.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Name | Units | Angle from Nadir | Sun-Sensor Azimuthal Angle |
|---|---|---|---|---|
| λ | Wavelength | nm | - | - |
| LT | Above-water radiance | μW cm−2sr−1nm−1 | 5° | 135° |
| Lspec | White panel radiance | μW cm−2sr−1nm−1 | 5° | 135° |
| Lsky | Sky radiance | μW cm−2sr−1nm−1 | 175° | 135° |
| Proportionality factor | - | - | - |
| Spectral Sample Type | Samples Collected | Samples Removed during Quality Control | Samples Used in Analysis |
|---|---|---|---|
| Bulbs (surface-100 cm) | 220 | 0 | 220 |
| Blades (surface-100 cm) | 220 | 51 | 169 |
| Water | 60 | 0 | 60 |
| Sky | 40 | 0 | 40 |
| Band | WV3 | MSRE |
|---|---|---|
| Blue | 445–517 nm | 459–491 nm |
| Green | 507–586 nm | 546.5–573.5 nm |
| Red | 626–696 nm | 661–675 nm |
| Red-edge | 698–749 nm | 711–723 nm |
| Near-infrared | 765–899 nm | 813.5–870.5 nm |
| Vegetation Index (VIn) | VIn Equation |
|---|---|
| RE_R | |
| RE_G | |
| RE_B | |
| NIR_R | |
| NIR_G | |
| NIR_B |
| Index | RE_B | RE_G | RE_R | NIR_B | NIR_G | NIR_R | ||
|---|---|---|---|---|---|---|---|---|
| MSRE | Bulb | Conservative (0.0) | >100 | 90 | >100 | 40 | 30 | 30 |
| Dynamic (max.) | >100 | >100 | >100 | >100 | 90 | 50 | ||
| Blade | Conservative (0.0) | 50 | 40 | 90 | 10 | 10 | 10 | |
| Dynamic (max.) | >100 | >100 | >100 | >100 | >100 | 30 | ||
| WV3 | Bulb | Conservative (0.0) | 90 | 80 | 100 | 50 | 40 | 40 |
| Dynamic (max.) | >100 | >100 | >100 | >100 | >100 | 80 | ||
| Blade | Conservative (0.0) | 40 | 40 | 60 | 20 | 10 | 20 | |
| Dynamic (max.) | >100 | >100 | >100 | >100 | >100 | 40 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timmer, B.; Reshitnyk, L.Y.; Hessing-Lewis, M.; Juanes, F.; Costa, M. Comparing the Use of Red-Edge and Near-Infrared Wavelength Ranges for Detecting Submerged Kelp Canopy. Remote Sens. 2022, 14, 2241. https://doi.org/10.3390/rs14092241
Timmer B, Reshitnyk LY, Hessing-Lewis M, Juanes F, Costa M. Comparing the Use of Red-Edge and Near-Infrared Wavelength Ranges for Detecting Submerged Kelp Canopy. Remote Sensing. 2022; 14(9):2241. https://doi.org/10.3390/rs14092241
Chicago/Turabian StyleTimmer, Brian, Luba Y. Reshitnyk, Margot Hessing-Lewis, Francis Juanes, and Maycira Costa. 2022. "Comparing the Use of Red-Edge and Near-Infrared Wavelength Ranges for Detecting Submerged Kelp Canopy" Remote Sensing 14, no. 9: 2241. https://doi.org/10.3390/rs14092241
APA StyleTimmer, B., Reshitnyk, L. Y., Hessing-Lewis, M., Juanes, F., & Costa, M. (2022). Comparing the Use of Red-Edge and Near-Infrared Wavelength Ranges for Detecting Submerged Kelp Canopy. Remote Sensing, 14(9), 2241. https://doi.org/10.3390/rs14092241