
Assessing the Impacts of Tidal Creeks on the Spatial Patterns of Coastal Salt Marsh Vegetation and Its Aboveground Biomass


Abstract
1. Introduction
2. Materials and Methods
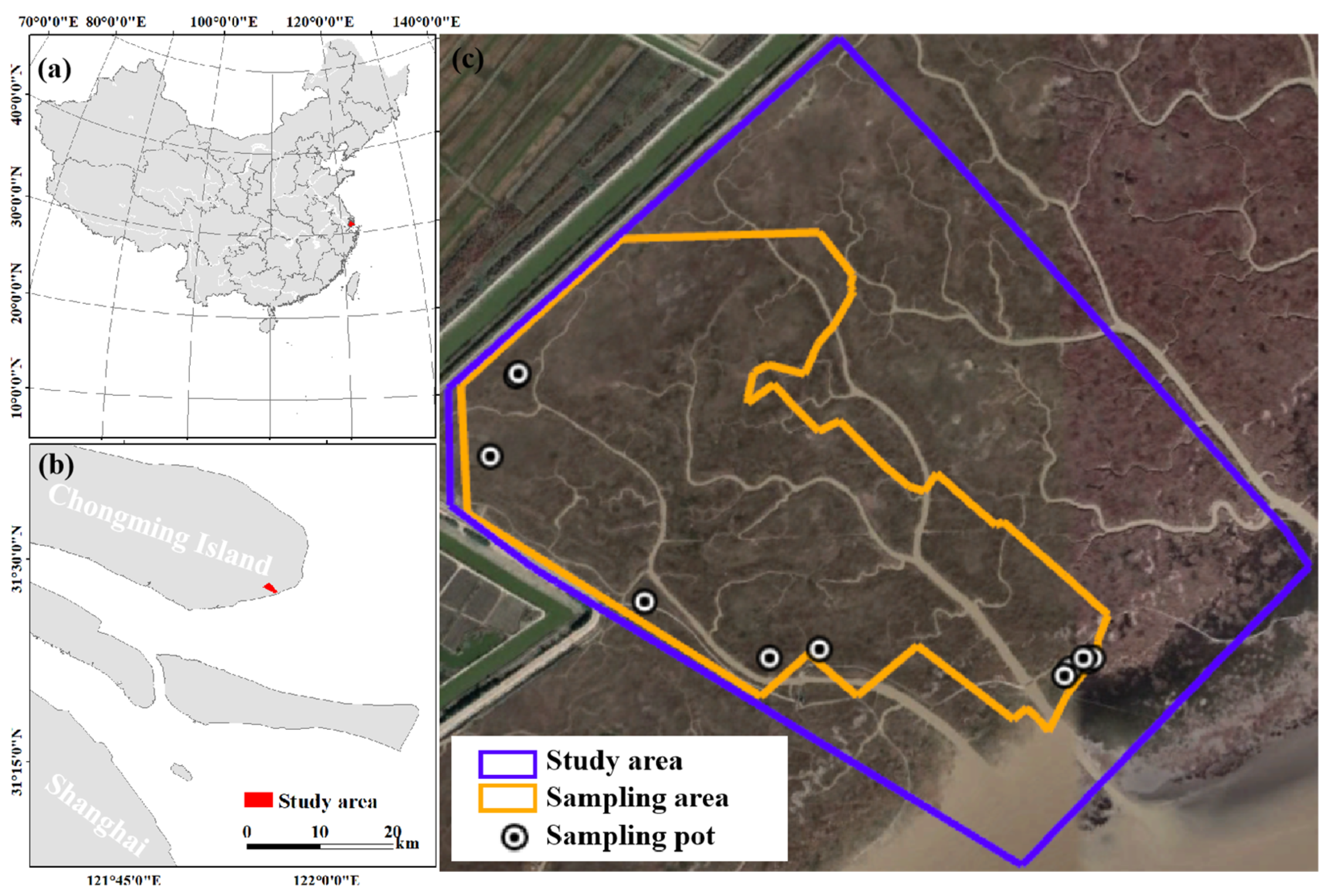
2.1. Study Area
2.2. Data Collection
2.2.1. Vegetation Sampling Data Acquisition
2.2.2. LiDAR and Optical Image Data Acquisition
2.2.3. Soil Analysis
2.3. Data Preprocessing
2.3.1. UAV Image Processing and Vegetation Community Mapping
2.3.2. LiDAR Data Processing
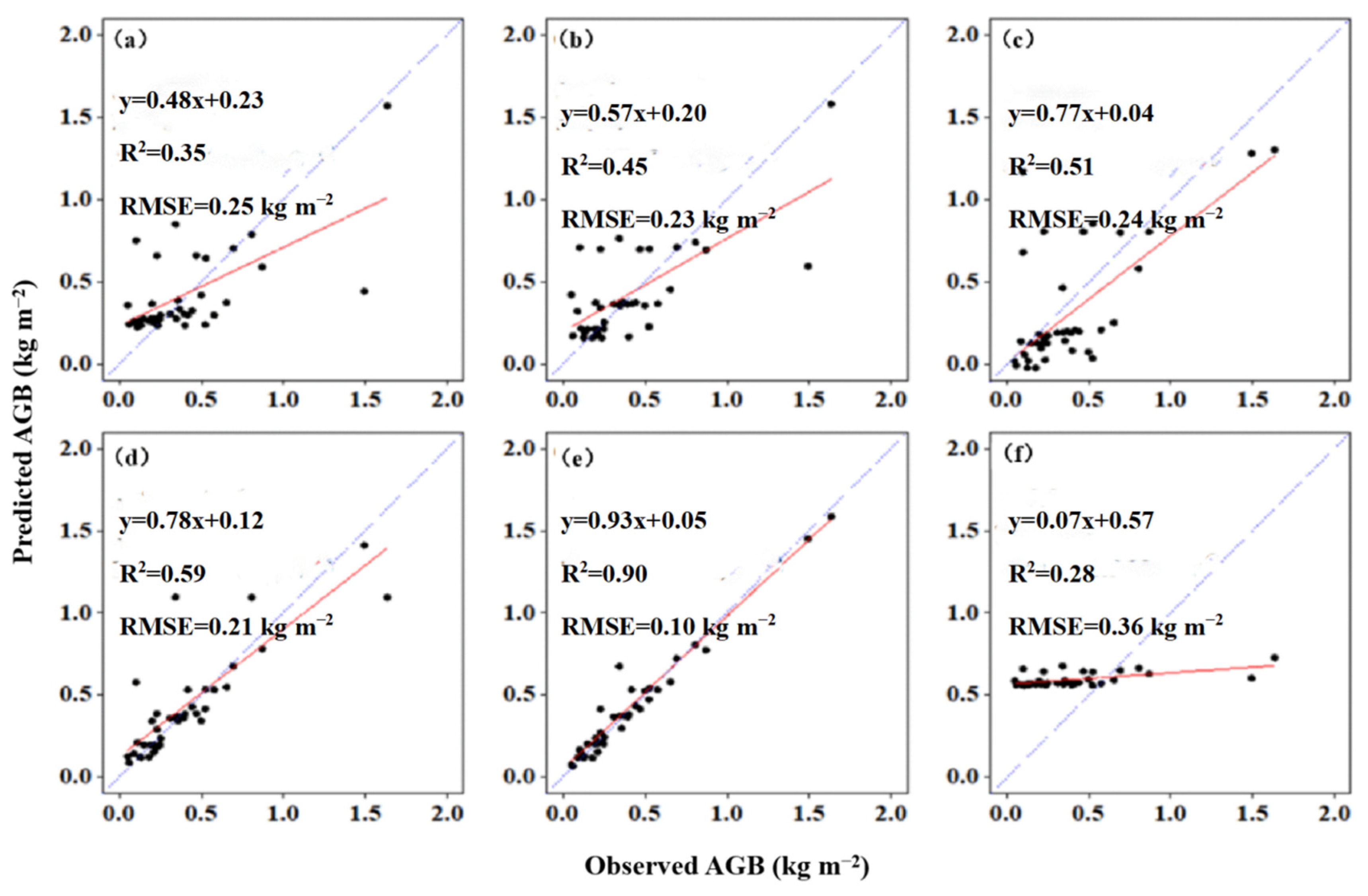
2.4. Establishment of AGB Prediction Model
2.5. Data Analysis
3. Results
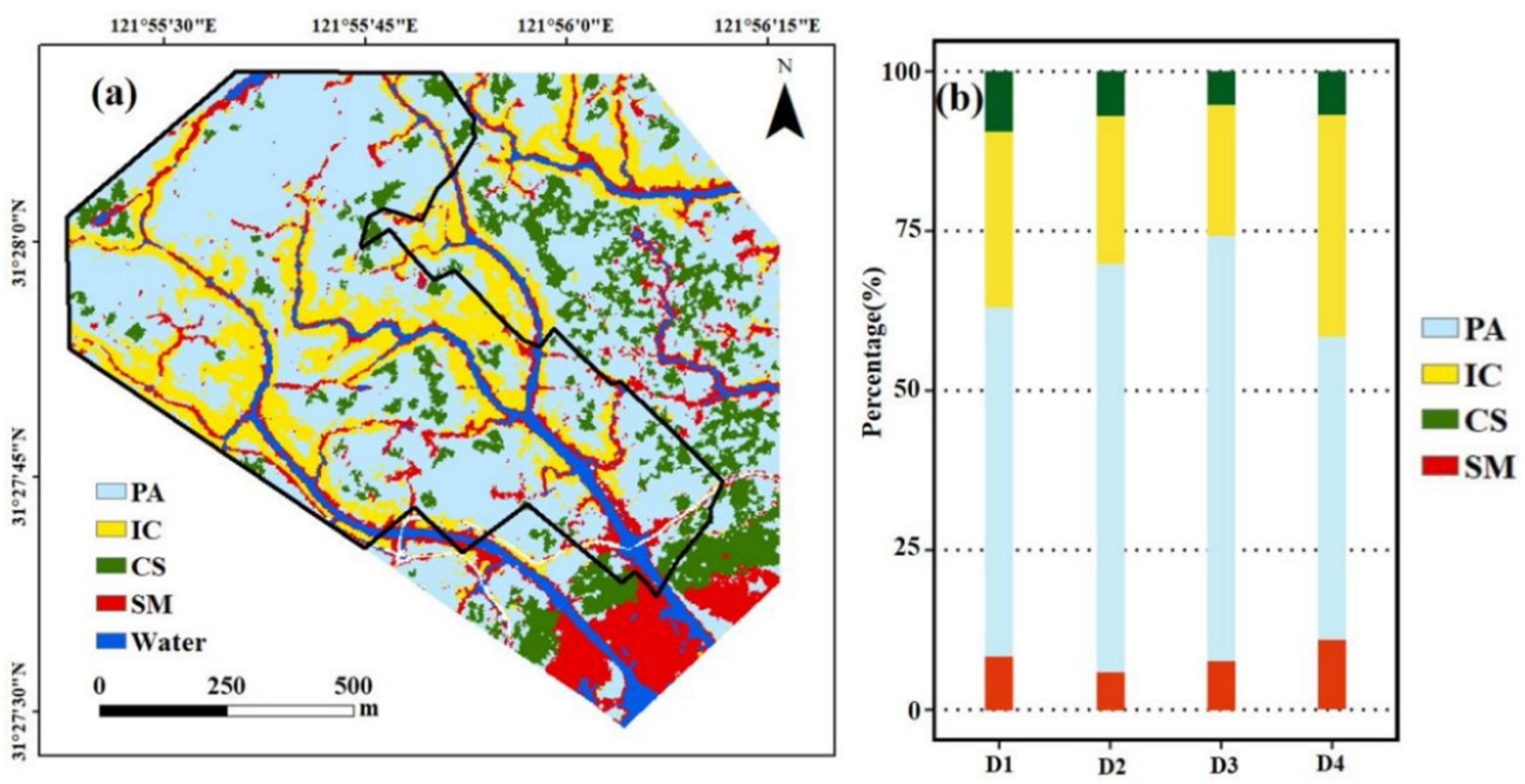
3.1. Accuracy Assessment of the Spatial Distribution of Salt Marsh Communities
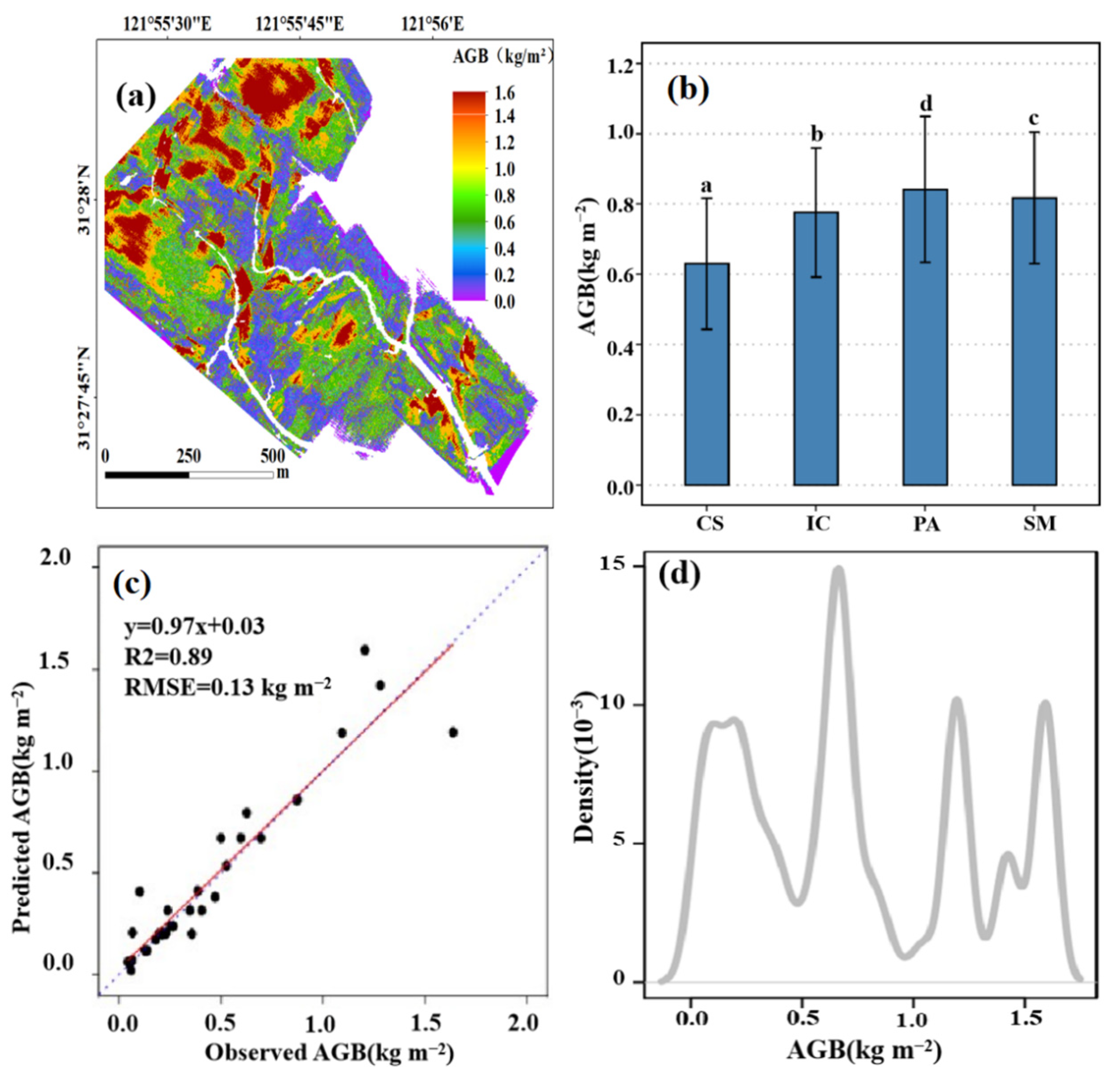
3.2. Estimation of Salt Marsh AGB and Its Spatial Pattern
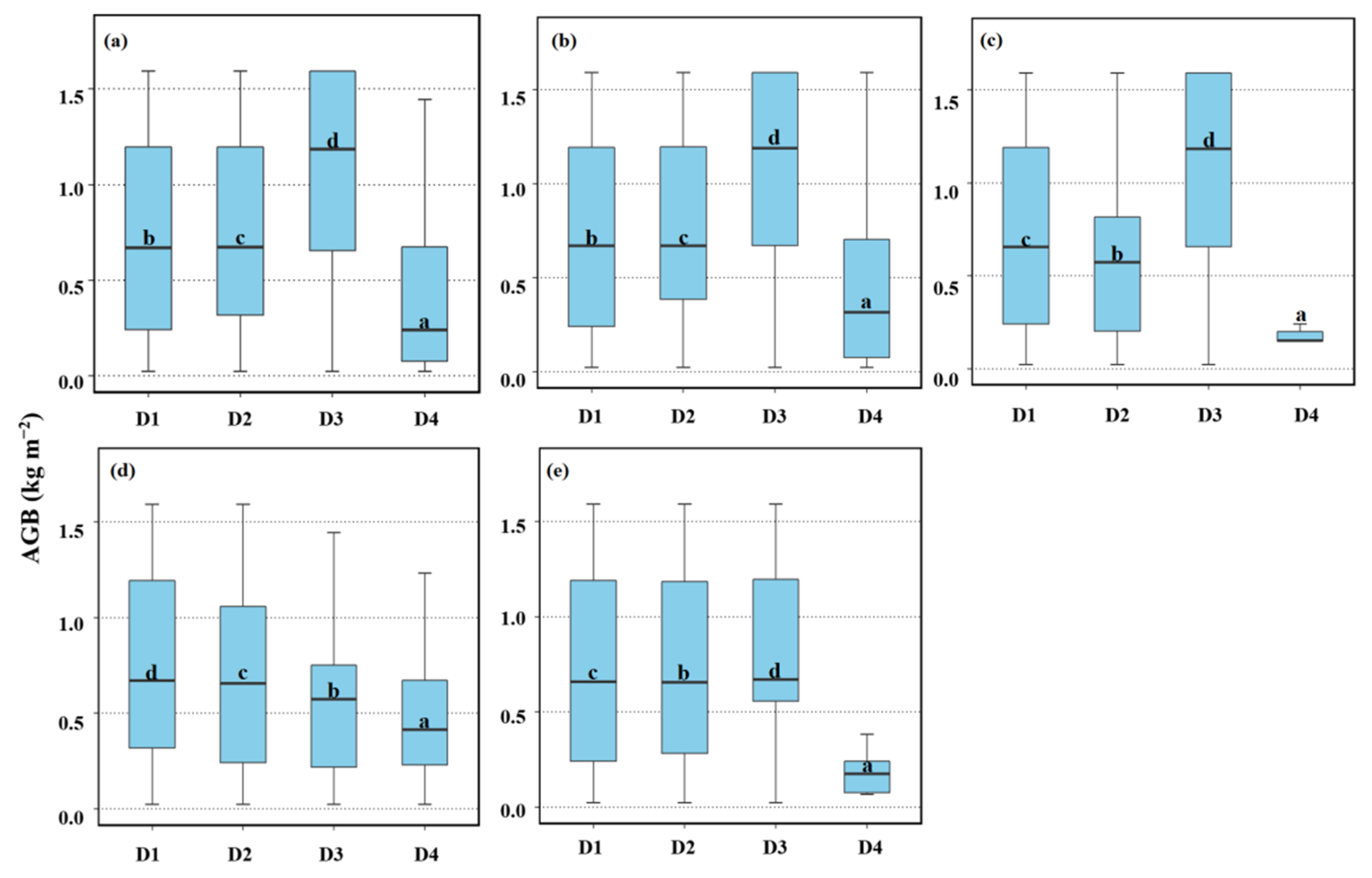
3.3. Impacts of Tidal Creeks on the AGB of Different Salt Marsh Communities
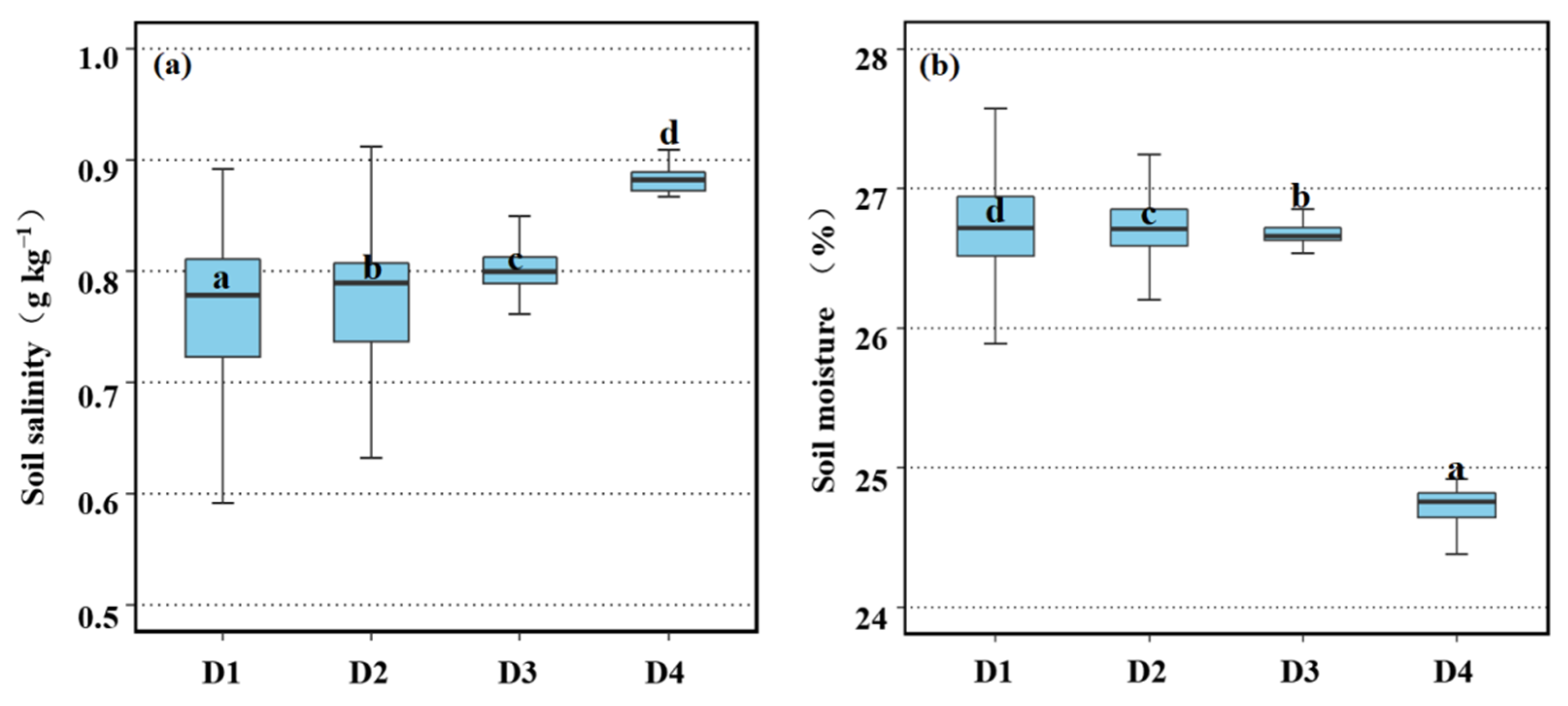
3.4. Spatial Variation in Soil Salinity and Moisture
4. Discussion
4.1. Impacts of Tidal Creeks on the Spatial Distribution of Salt Marsh Communities and Their AGB
4.2. Advantages of AGB Estimation Using UAV-LiDAR Data and Machine Learning Approaches
4.3. Management of Coastal Salt Marsh
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edge, R.S.; Sullivan, M.J.; Pedley, S.M.; Mossman, H.L. Species interactions modulate the response of saltmarsh plants to flooding. Ann. Bot. 2020, 125, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Boscutti, F.; De Nobili, M.; Casolo, V. Plant traits shape the effects of tidal flooding on soil and plant communities in saltmarshes. Plant Ecol. 2018, 219, 823–835. [Google Scholar] [CrossRef]
- Kelleway, J.J.; Cavanaugh, K.; Rogers, K.; Feller, I.C.; Ens, E.; Doughty, C.; Saintilan, N. Review of the ecosystem service implications of mangrove encroachment into salt marshes. Glob. Change Biol. 2017, 23, 3967–3983. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, W.J.; Bernal, B.; Hernandez, M.E. Ecosystem services of wetlands. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 1–4. [Google Scholar] [CrossRef]
- Beaumont, N.J.; Jones, L.; Garbutt, A.; Hansom, J.; Toberman, M. The value of carbon sequestration and storage in coastal habitats. Estuar. Coast. Shelf Sci. 2014, 137, 32–40. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marbà, N. Estimating global “blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetl. Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Chen, J.; Gu, S.; Shen, M.; Tang, Y.; Matsushita, B. Estimating aboveground biomass of grassland having a high canopy cover: An exploratory analysis of in situ hyperspectral data. Int. J. Remote Sens. 2009, 30, 6497–6517. [Google Scholar] [CrossRef]
- Du, Y.; Wang, J.; Liu, Z.; Yu, H.; Li, Z.; Cheng, H. Evaluation on spaceborne multispectral images, airborne hyperspectral, and LiDAR data for extracting spatial distribution and estimating aboveground biomass of wetland vegetation suaeda salsa. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 12, 200–209. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Yu, H.; Li, F. Mapping Spartina alterniflora biomass using LiDAR and hyperspectral data. Remote Sens. 2017, 9, 589. [Google Scholar] [CrossRef]
- Bertness, M.D.; Pennings, S.C. Spatial Variation in Process and Pattern in Salt Marsh Plant Communities in Eastern North America. In Concepts and Controversies in Tidal Marsh Ecology; Springer: New, York, NY, USA, 2002; pp. 39–57. [Google Scholar]
- Crain, C.M.; Silliman, B.R.; Bertness, S.L.; Bertness, M.D. Physical and biotic drivers of plant distribution across estuarine salinity gradients. Ecology 2004, 85, 2539–2549. [Google Scholar] [CrossRef]
- Xue, L.; Jiang, J.; Li, X.; Yan, Z.; Zhang, Q.; Ge, Z.; Tian, B.; Craft, C. Salinity Affects Topsoil Organic Carbon Concentrations Through Regulating Vegetation Structure and Productivity. J. Geophys. Res. Biogeosciences 2020, 125, e2019JG005217. [Google Scholar] [CrossRef]
- Pennings, S.C.; Grant, M.-B.; Bertness, M.D. Plant zonation in low-latitude salt marshes: Disentangling the roles of flooding, salinity and competition. J. Ecol. 2005, 93, 159–167. [Google Scholar] [CrossRef]
- Kearney, W.S.; Fagherazzi, S. Salt marsh vegetation promotes efficient tidal channel networks. Nat. Commun. 2016, 7, 12287. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Meng, H.; Dong, H.; Guo, Y.; Tong, S. Distribution pattern of plant community in new-born coastal wetland in the Yellow River Delta. Sci. Geogr. Sin. 2015, 35, 1021–1026. [Google Scholar]
- Zhao, X.; Cui, B.; Sun, T.; He, Q. The relationship between the spatial distribution of vegetation and soil environmental factors in the tidal creek areas of the Yellow River Delta. Ecol. Environ. Sci. 2010, 19, 1855–1861. [Google Scholar]
- Balling, S.S.; Resh, V.H. The influence of mosquito control recirculation ditches on plant biomass, production and composition in two San Francisco Bay salt marshes. Estuar. Coast. Shelf Sci. 1983, 16, 151–161. [Google Scholar] [CrossRef]
- Byrd, K.B.; O’Connell, J.L.; Di Tommaso, S.; Kelly, M. Evaluation of sensor types and environmental controls on mapping biomass of coastal marsh emergent vegetation. Remote Sens. Environ. 2014, 149, 166–180. [Google Scholar] [CrossRef]
- Ozesmi, S.L.; Bauer, M.E. Satellite remote sensing of wetlands. Wetl. Ecol. Manag. 2002, 10, 381–402. [Google Scholar] [CrossRef]
- Miller, G.J.; Morris, J.T.; Wang, C. Estimating aboveground biomass and its spatial distribution in coastal wetlands utilizing planet multispectral imagery. Remote Sens. 2019, 11, 2020. [Google Scholar] [CrossRef]
- Lumbierres, M.; Méndez, P.F.; Bustamante, J.; Soriguer, R.; Santamaría, L. Modeling biomass production in seasonal wetlands using MODIS NDVI land surface phenology. Remote Sens. 2017, 9, 392. [Google Scholar] [CrossRef]
- Vaghela, B.; Chirakkal, S.; Putrevu, D.; Solanki, H. Modelling above ground biomass of Indian mangrove forest using dual-pol SAR data. Remote Sens. Appl. Soc. Environ. 2021, 21, 100457. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, Q.; Huang, H.; Huang, Y.; Tao, J.; Zhou, G.; Zhang, Y.; Yang, Y.; Lin, J. Aboveground biomass of typical invasive mangroves and its distribution patterns using UAV-LiDAR data in a subtropical estuary: Maoling River estuary, Guangxi, China. Ecol. Indic. 2022, 136, 108694. [Google Scholar] [CrossRef]
- Fatoyinbo, T.; Feliciano, E.A.; Lagomasino, D.; Lee, S.K.; Trettin, C. Estimating mangrove aboveground biomass from airborne LiDAR data: A case study from the Zambezi River delta. Environ. Res. Lett. 2018, 13, 025012. [Google Scholar] [CrossRef]
- Guo, M.; Li, J.; Sheng, C.; Xu, J.; Wu, L. A review of wetland remote sensing. Sensors 2017, 17, 777. [Google Scholar] [CrossRef] [PubMed]
- Dalponte, M.; Coops, N.C.; Bruzzone, L.; Gianelle, D. Analysis on the use of multiple returns LiDAR data for the estimation of tree stems volume. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2009, 2, 310–318. [Google Scholar] [CrossRef]
- Hsu, A.J.; Kumagai, J.; Favoretto, F.; Dorian, J.; Guerrero Martinez, B.; Aburto-Oropeza, O. Driven by Drones: Improving Mangrove Extent Maps Using High-Resolution Remote Sensing. Remote Sens. 2020, 12, 3986. [Google Scholar] [CrossRef]
- Cohen, M.C.; de Souza, A.V.; Liu, K.-b.; Rodrigues, E.; Yao, Q.; Ryu, J.; Dietz, M.; Pessenda, L.C.; Rossetti, D. Effects of the 2017–2018 winter freeze on the northern limit of the American mangroves, Mississippi River delta plain. Geomorphology 2021, 394, 107968. [Google Scholar] [CrossRef]
- Laporte-Fauret, Q.; Marieu, V.; Castelle, B.; Michalet, R.; Bujan, S.; Rosebery, D. Low-cost UAV for high-resolution and large-scale coastal dune change monitoring using photogrammetry. J. Mar. Sci. Eng. 2019, 7, 63. [Google Scholar] [CrossRef]
- Fabbri, S.; Grottoli, E.; Armaroli, C.; Ciavola, P. Using High-Spatial Resolution UAV-Derived Data to Evaluate Vegetation and Geomorphological Changes on a Dune Field Involved in a Restoration Endeavour. Remote Sens. 2021, 13, 1987. [Google Scholar] [CrossRef]
- Rende, S.F.; Bosman, A.; Di Mento, R.; Bruno, F.; Lagudi, A.; Irving, A.D.; Dattola, L.; Giambattista, L.D.; Lanera, P.; Proietti, R. Ultra-high-resolution mapping of Posidonia oceanica (L.) delile meadows through acoustic, optical data and object-based image classification. J. Mar. Sci. Eng. 2020, 8, 647. [Google Scholar] [CrossRef]
- Klemas, V.V. Coastal and environmental remote sensing from unmanned aerial vehicles: An overview. J. Coast. Res. 2015, 31, 1260–1267. [Google Scholar] [CrossRef]
- Whitehead, K.; Hugenholtz, C.H. Remote sensing of the environment with small unmanned aircraft systems (UASs), part 1: A review of progress and challenges. J. Unmanned Veh. Syst. 2014, 2, 69–85. [Google Scholar] [CrossRef]
- Xu, Z.; Li, W.; Li, Y.; Shen, X.; Ruan, H. Estimation of secondary forest parameters by integrating image and point cloud-based metrics acquired from unmanned aerial vehicle. J. Appl. Remote Sens. 2019, 14, 022204. [Google Scholar] [CrossRef][Green Version]
- Kalacska, M.; Chmura, G.; Lucanus, O.; Bérubé, D.; Arroyo-Mora, J. Structure from motion will revolutionize analyses of tidal wetland landscapes. Remote Sens. Environ. 2017, 199, 14–24. [Google Scholar] [CrossRef]
- Doughty, C.L.; Cavanaugh, K.C. Mapping coastal wetland biomass from high resolution unmanned aerial vehicle (UAV) imagery. Remote Sens. 2019, 11, 540. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, X.; Wang, X.; Xu, X.; Chen, B.; Wang, J.; Ma, J.; Zhao, B.; Li, B. Quantifying expansion and removal of Spartina alterniflora on Chongming island, China, using time series Landsat images during 1995–2018. Remote Sens. Environ. 2020, 247, 111916. [Google Scholar] [CrossRef]
- Jiang, Y.; Du, J.; Zhang, J.; Zhang, W.; Zhang, J. The determination of sedimentation rates in various vegetational zones of Chongming tidal flat of the Changjiang Estuary. Acta Oceanol. Sin. 2012, 34, 114–121. [Google Scholar]
- Shilun, Y. A study of coastal morphodynamics on the muddy islands in the Changjiang River estuary. J. Coast. Res. 1999, 15, 32–44. [Google Scholar]
- Ding, W.-H.; Jiang, J.-Y.; Li, X.-Z.; Huang, X.; Li, X.-Z.; Zhou, Y.-X.; Tang, C.-D. Spatial distribution of species and influencing factors across salt marsh in southern Chongming Dongtan. Chin. J. Plant. Ecol. 2015, 39, 704–716. [Google Scholar]
- Gao, Z.; Zhang, L. Multi-seasonal spectral characteristics analysis of coastal salt marsh vegetation in Shanghai, China. Estuar. Coast. Shelf Sci. 2006, 69, 217–224. [Google Scholar] [CrossRef]
- Yan, Q.; Lu, J.; He, W. Succession character of salt marsh vegetations in Chongming Dongtan wetland. J. Appl. Ecol. 2007, 18, 1097–1101. [Google Scholar]
- Foody, G.M. Status of land cover classification accuracy assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Uebersax, J.S. Diversity of decision-making models and the measurement of interrater agreement. Psychol. Bull. 1987, 101, 140. [Google Scholar] [CrossRef]
- Tung, F.; LeDrew, E. The determination of optimal threshold levels for change detection using various accuracy indexes. Photogramm. Eng. Remote Sens. 1988, 54, 1449–1454. [Google Scholar]
- Story, M.; Congalton, R.G. Accuracy assessment: A user’s perspective. Photogramm. Eng. Remote Sens. 1986, 52, 397–399. [Google Scholar]
- Tranmer, M.; Elliot, M. Multiple linear regression. Cathie Marsh Cent. Census Surv. Res. (CCSR) 2008, 5, 1–5. [Google Scholar]
- Guisan, A.; Edwards, T.C., Jr.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z. Artificial Neural Network. In Multivariate Time Series Analysis in Climate and Environmental Research; Springer: New, York, NY, USA, 2018; pp. 1–35. [Google Scholar]
- Engel, Y.; Mannor, S.; Meir, R. The kernel recursive least-squares algorithm. IEEE Trans. Signal Process. 2004, 52, 2275–2285. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Shen, W.; Li, M.; Huang, C.; Tao, X.; Wei, A. Annual Forest aboveground biomass changes mapped using ICESat/GLAS measurements, historical inventory data, and time-series optical and radar imagery for Guangdong province, China. Agric. For. Meteorol. 2018, 259, 23–38. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Yan, G.; Zhai, J.; Cong, L.; Dai, L.; Zhang, Z.; Zhang, M. The size and distribution of tidal creeks affects salt marsh restoration. J. Environ. Manag. 2020, 259, 110070. [Google Scholar] [CrossRef]
- Snow, A.A.; Vince, S.W. Plant zonation in an Alaskan salt marsh: II. An experimental study of the role of edaphic conditions. J. Ecol. 1984, 7, 669–684. [Google Scholar] [CrossRef]
- He, Y.; Li, X.; Ma, Z.; Sun, Y.; Jia, Y. Vegetation zonation related to the edaphic factors in the East headland of Chongming Island. Acta Ecol. Sin. 2010, 30, 4919–4927. [Google Scholar]
- Li, W.-Q.; Xiao-Jing, L.; Khan, M.A.; Gul, B. Relationship between soil characteristics and halophytic vegetation in coastal region of North China. Pak. J. Bot. 2008, 40, 1081–1090. [Google Scholar]
- Emery, N.C.; Ewanchuk, P.J.; Bertness, M.D. Competition and salt-marsh plant zonation: Stress tolerators may be dominant competitors. Ecology 2001, 82, 2471–2485. [Google Scholar] [CrossRef]
- Ishikawa, S.-I.; Kachi, N. Shoot population dynamics of Carex kobomugi on a coastal sand dune in relation to its zonal distribution. Aust. J. Bot. 1998, 46, 111–121. [Google Scholar] [CrossRef]
- Janousek, C.N.; Mayo, C. Plant responses to increased inundation and salt exposure: Interactive effects on tidal marsh productivity. Plant Ecol. 2013, 214, 917–928. [Google Scholar] [CrossRef]
- Brown, A.M.; Bledsoe, C. Spatial and temporal dynamics of mycorrhizas in Jaumea carnosa, a tidal saltmarsh halophyte. J. Ecol. 1996, 84, 703–715. [Google Scholar] [CrossRef]
- Valiela, I.; Teal, J.M.; Deuser, W.G. The nature of growth forms in the salt marsh grass Spartina alterniflora. Am. Nat. 1978, 112, 461–470. [Google Scholar] [CrossRef]
- Bockelmann, A.-C.; Bakker, J.P.; Neuhaus, R.; Lage, J. The relation between vegetation zonation, elevation and inundation frequency in a Wadden Sea salt marsh. Aquat. Bot. 2002, 73, 211–221. [Google Scholar] [CrossRef]
- Mossman, H.L.; Grant, A.; Davy, A.J. Manipulating saltmarsh microtopography modulates the effects of elevation on sediment redox potential and halophyte distribution. J. Ecol. 2020, 108, 94–106. [Google Scholar] [CrossRef]
- Cao, H.; Zhu, Z.; van Belzen, J.; Gourgue, O.; van de Koppel, J.; Temmerman, O.S.; Herman, P.M.; Zhang, L.; Yuan, L.; Bouma, T.J. Salt marsh establishment in poorly consolidated muddy systems: Effects of surface drainage, elevation, and plant age. Ecosphere 2021, 12, e03755. [Google Scholar] [CrossRef]
- Wang, D.; Wan, B.; Liu, J.; Su, Y.; Guo, Q.; Qiu, P.; Wu, X. Estimating aboveground biomass of the mangrove forests on northeast Hainan Island in China using an upscaling method from field plots, UAV-LiDAR data and Sentinel-2 imagery. Int. J. Appl. Earth Obs. Geoinf. 2020, 85, 101986. [Google Scholar] [CrossRef]
- Janowski, L.; Tylmann, K.; Trzcinska, K.; Rudowski, S.; Tegowski, J. Exploration of glacial landforms by object-based image analysis and spectral parameters of digital elevation model. IEEE Trans. Geosci. Remote Sens. 2021, 60, 1–17. [Google Scholar] [CrossRef]
- Guo, Q.; Su, Y.; Hu, T.; Zhao, X.; Wu, F.; Li, Y.; Liu, J.; Chen, L.; Xu, G.; Lin, G. An integrated UAV-borne lidar system for 3D habitat mapping in three forest ecosystems across China. Int. J. Remote Sens. 2017, 38, 2954–2972. [Google Scholar] [CrossRef]
- Maimaitijiang, M.; Sagan, V.; Sidike, P.; Hartling, S.; Esposito, F.; Fritschi, F.B. Soybean yield prediction from UAV using multimodal data fusion and deep learning. Remote Sens. Environ. 2020, 237, 111599. [Google Scholar] [CrossRef]
- Xu, J.-X.; Ma, J.; Tang, Y.-N.; Wu, W.-X.; Shao, J.-H.; Wu, W.-B.; Wei, S.-Y.; Liu, Y.-F.; Wang, Y.-C.; Guo, H.-Q. Estimation of Sugarcane Yield Using a Machine Learning Approach Based on UAV-LiDAR Data. Remote Sens. 2020, 12, 2823. [Google Scholar] [CrossRef]
- Gleason, C.J.; Im, J. Forest biomass estimation from airborne LiDAR data using machine learning approaches. Remote Sens. Environ. 2012, 125, 80–91. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, C.; Yang, H.; Yang, G.; Han, L.; Li, Z.; Feng, H.; Xu, B.; Wu, J.; Lei, L. Estimation of maize above-ground biomass based on stem-leaf separation strategy integrated with LiDAR and optical remote sensing data. PeerJ 2019, 7, e7593. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, X.; Jiang, J.; Xue, L.; Craft, C.B. Distribution of organic carbon storage in different salt-marsh plant communities: A case study at the Yangtze estuary. Estuar. Coast. Shelf Sci. 2020, 243, 106900. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Foin, T.C.; Ustin, S.L. A simple empirical model of salt marsh plant spatial distributions with respect to a tidal channel network. Ecol. Model. 2001, 139, 293–307. [Google Scholar] [CrossRef]
- Call, M.; Sanders, C.J.; Macklin, P.A.; Santos, I.R.; Maher, D.T. Carbon outwelling and emissions from two contrasting mangrove creeks during the monsoon storm season in Palau, Micronesia. Estuar. Coast. Shelf Sci. 2019, 218, 340–348. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Vegetation Community | Species Composition | Number of Samples | Average Value of an AGB (kg m−2) |
|---|---|---|---|---|
| PA | Phragmites australis | Phragmites australis, Carex scabrifolia, Imperata cylindrica | 21 | 0.56 |
| IC | Imperata cylindrica | Imperata cylindrica, Phragmites australis | 9 | 0.21 |
| SM | Scirpus mariqueter | Scirpus mariqueter | 9 | 0.29 |
| CS | Carex scabrifolia | Carex scabrifolia, Phragmites australis | 21 | 0.21 |
| Community Type | Producer Accuracy (%) | User Accuracy (%) |
|---|---|---|
| PA | 97.07 | 96.89 |
| IC | 79.17 | 98.88 |
| SM | 99.88 | 95.15 |
| CS | 96.46 | 98.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.-N.; Ma, J.; Xu, J.-X.; Wu, W.-B.; Wang, Y.-C.; Guo, H.-Q. Assessing the Impacts of Tidal Creeks on the Spatial Patterns of Coastal Salt Marsh Vegetation and Its Aboveground Biomass. Remote Sens. 2022, 14, 1839. https://doi.org/10.3390/rs14081839
Tang Y-N, Ma J, Xu J-X, Wu W-B, Wang Y-C, Guo H-Q. Assessing the Impacts of Tidal Creeks on the Spatial Patterns of Coastal Salt Marsh Vegetation and Its Aboveground Biomass. Remote Sensing. 2022; 14(8):1839. https://doi.org/10.3390/rs14081839
Chicago/Turabian StyleTang, Ya-Nan, Jun Ma, Jing-Xian Xu, Wan-Ben Wu, Yuan-Chen Wang, and Hai-Qiang Guo. 2022. "Assessing the Impacts of Tidal Creeks on the Spatial Patterns of Coastal Salt Marsh Vegetation and Its Aboveground Biomass" Remote Sensing 14, no. 8: 1839. https://doi.org/10.3390/rs14081839
APA StyleTang, Y.-N., Ma, J., Xu, J.-X., Wu, W.-B., Wang, Y.-C., & Guo, H.-Q. (2022). Assessing the Impacts of Tidal Creeks on the Spatial Patterns of Coastal Salt Marsh Vegetation and Its Aboveground Biomass. Remote Sensing, 14(8), 1839. https://doi.org/10.3390/rs14081839