
Detection of Multidecadal Changes in Vegetation Dynamics and Association with Intra-Annual Climate Variability in the Columbia River Basin


Abstract
:1. Introduction
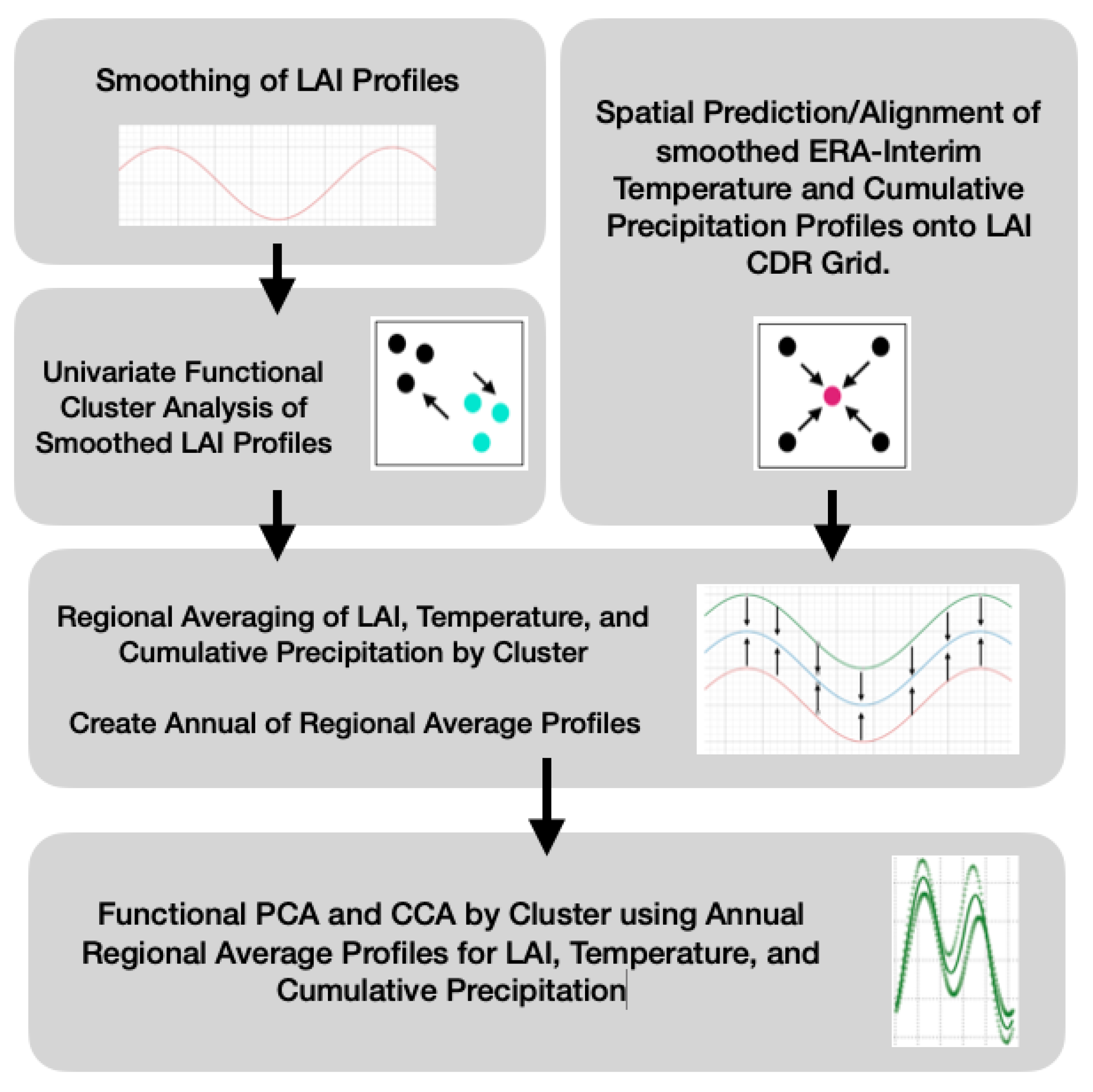
2. Materials and Methods
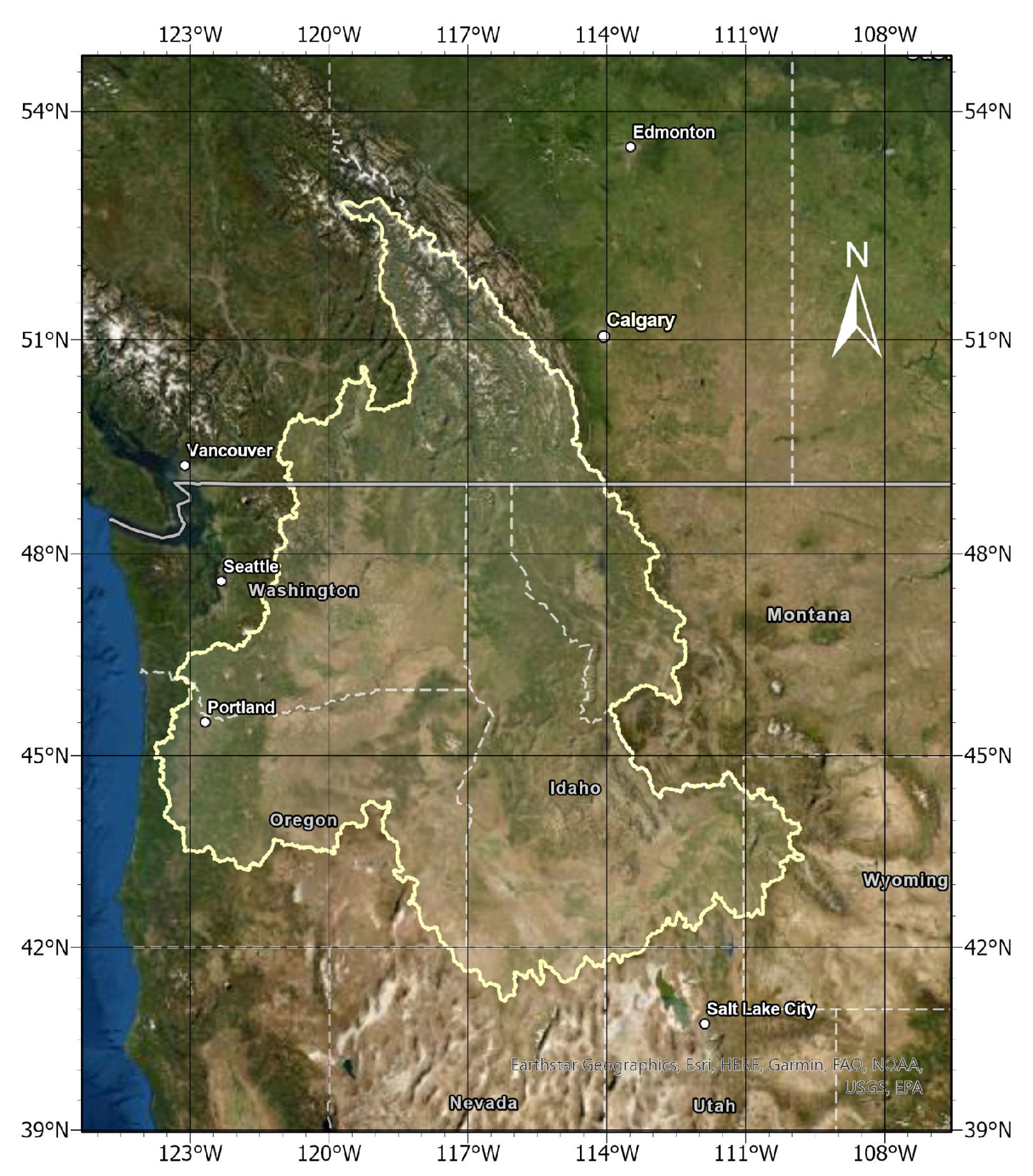
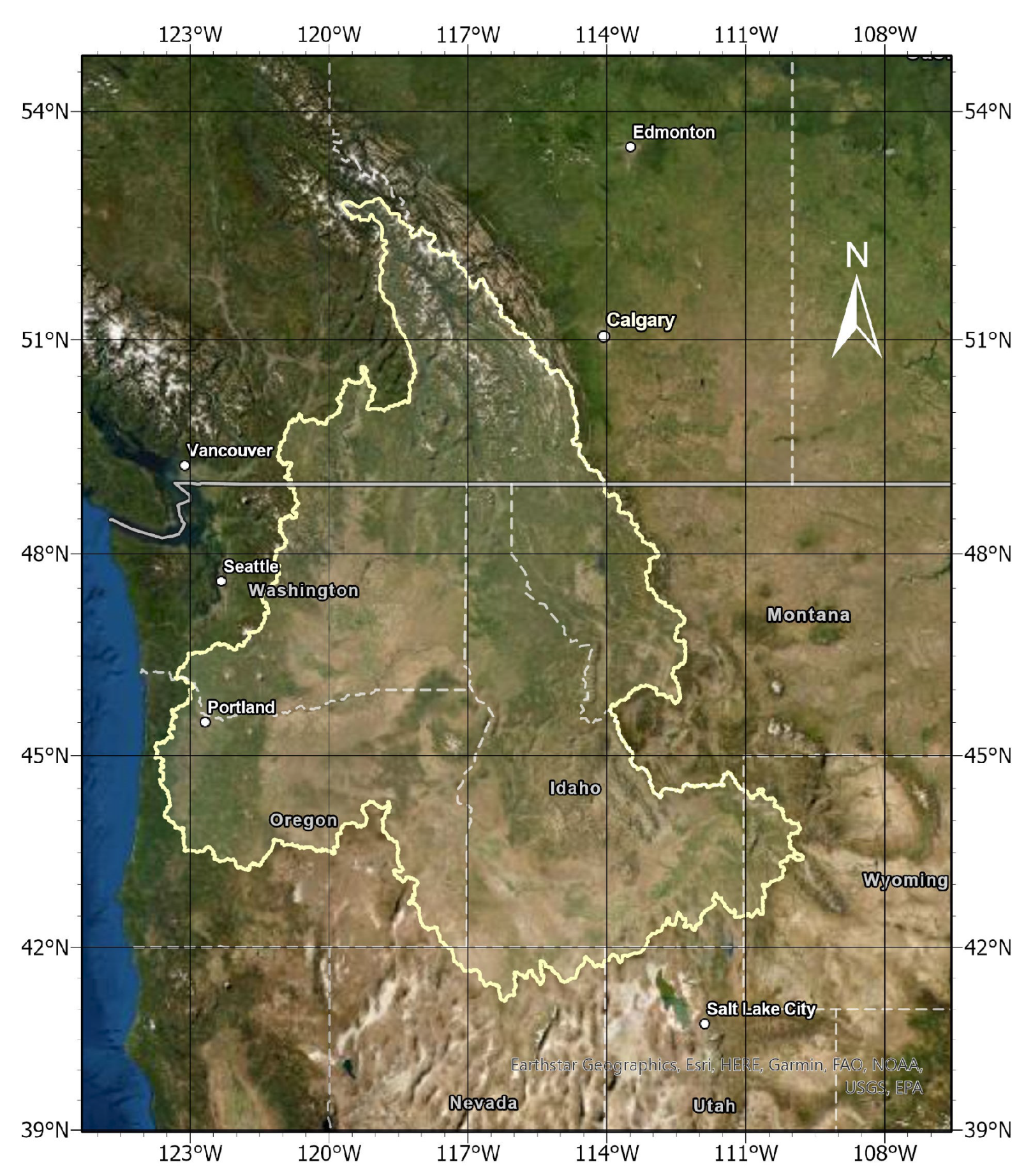
2.1. Study Region
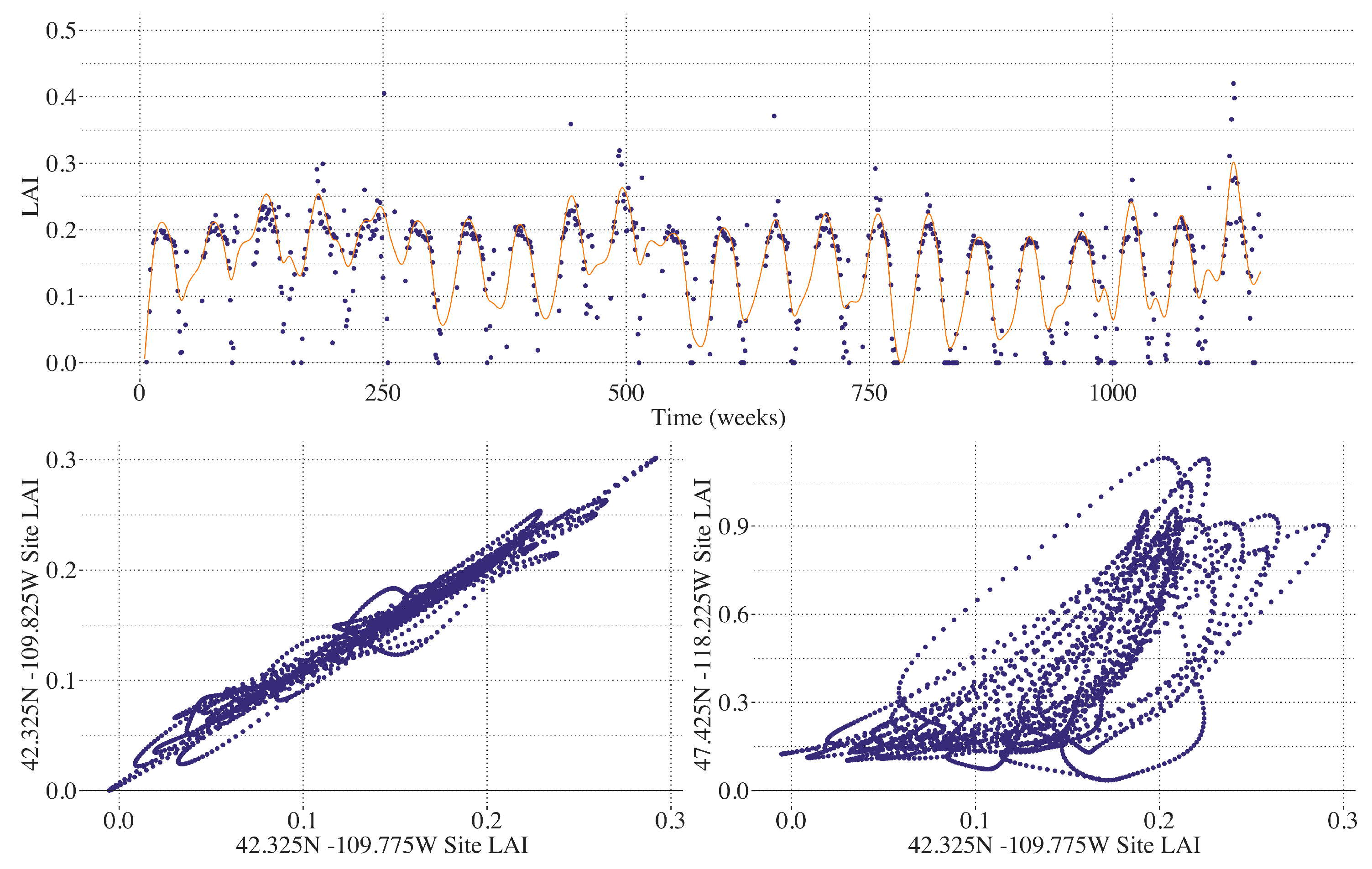
2.2. LAI AVHRR Climate Data Record
2.3. ERA-Interim Reanalysis
2.4. BaseVue 2013 Land Cover and USGS National Elevation Products
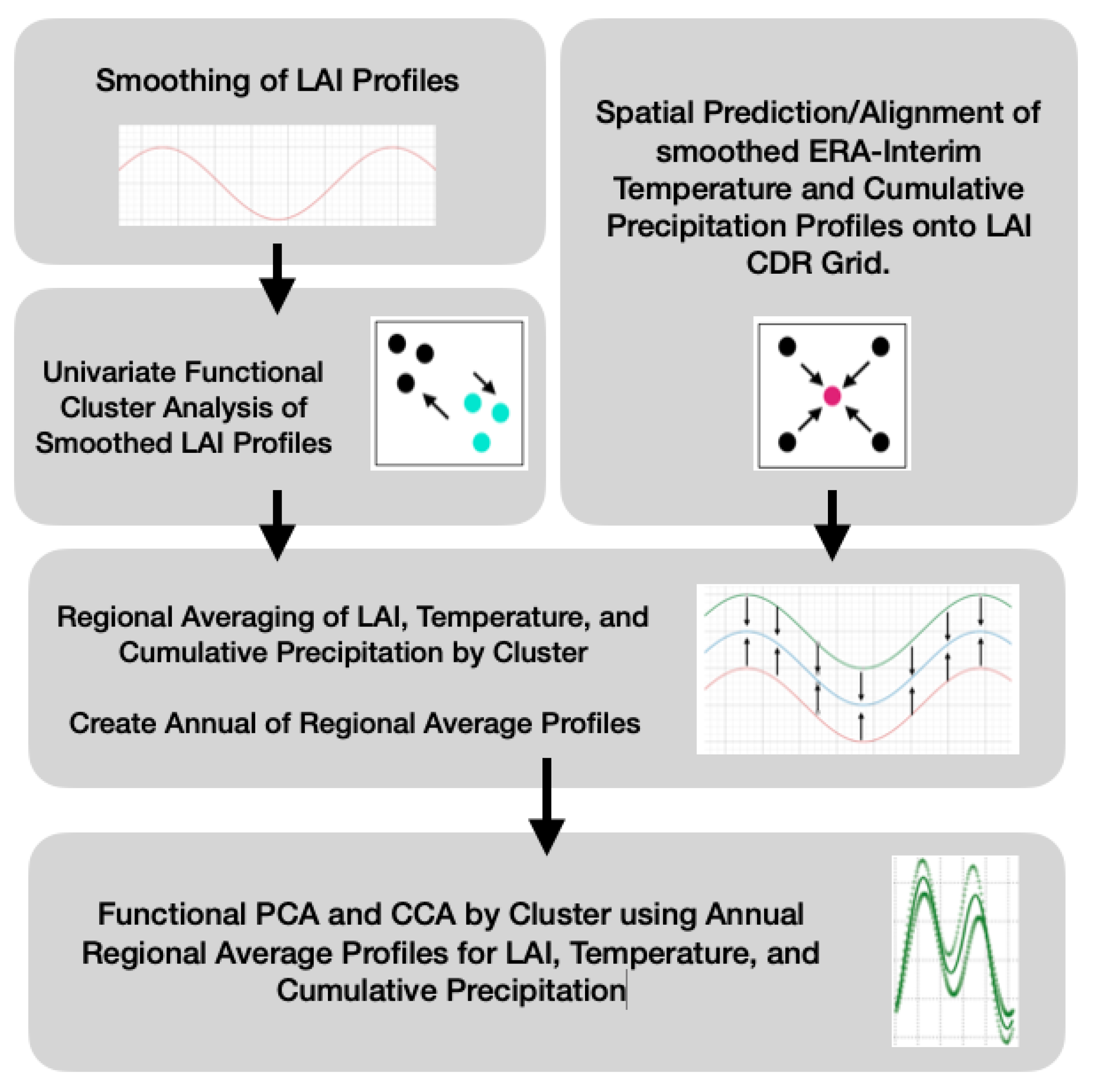
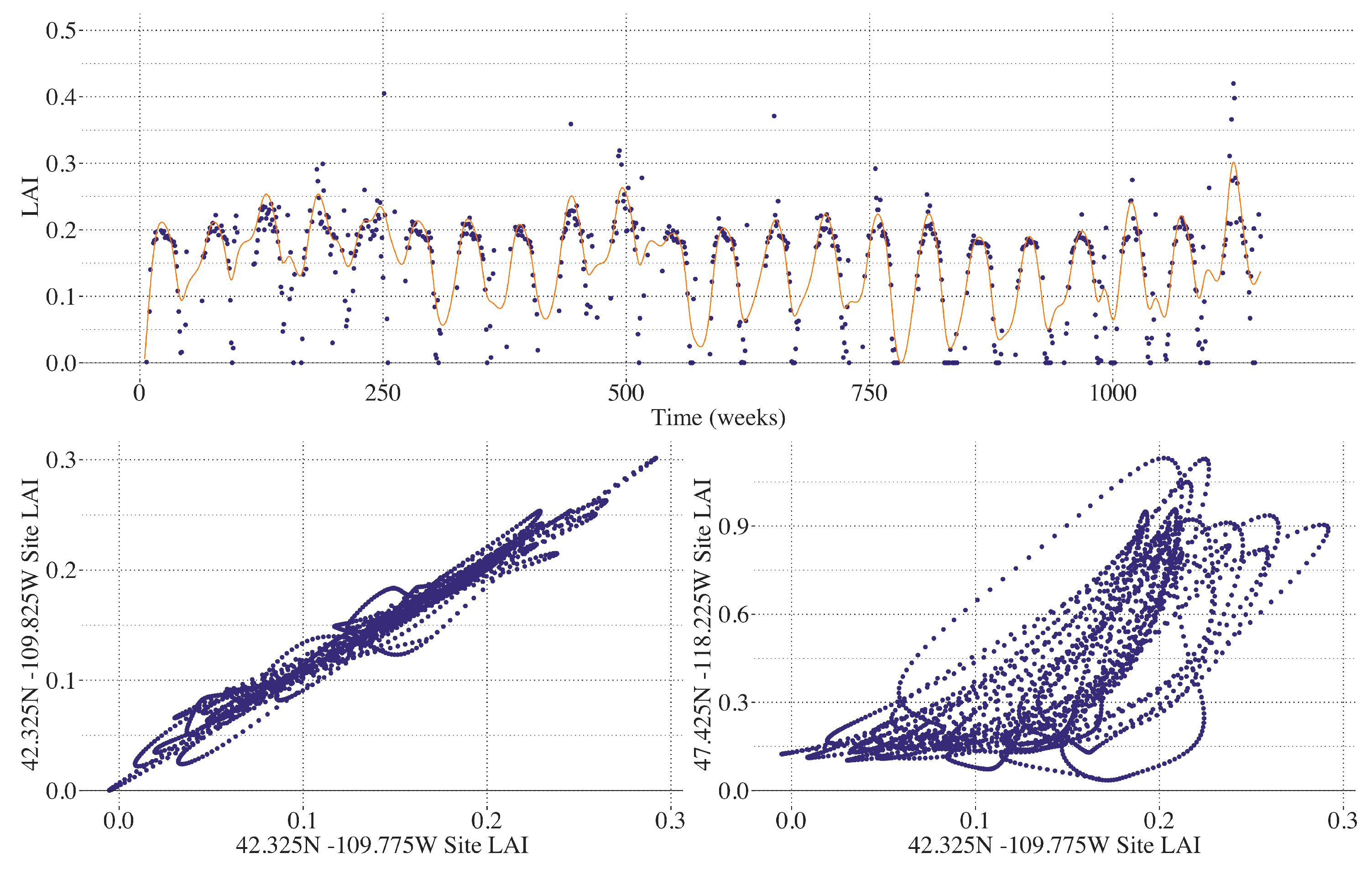
2.5. Smoothing LAI
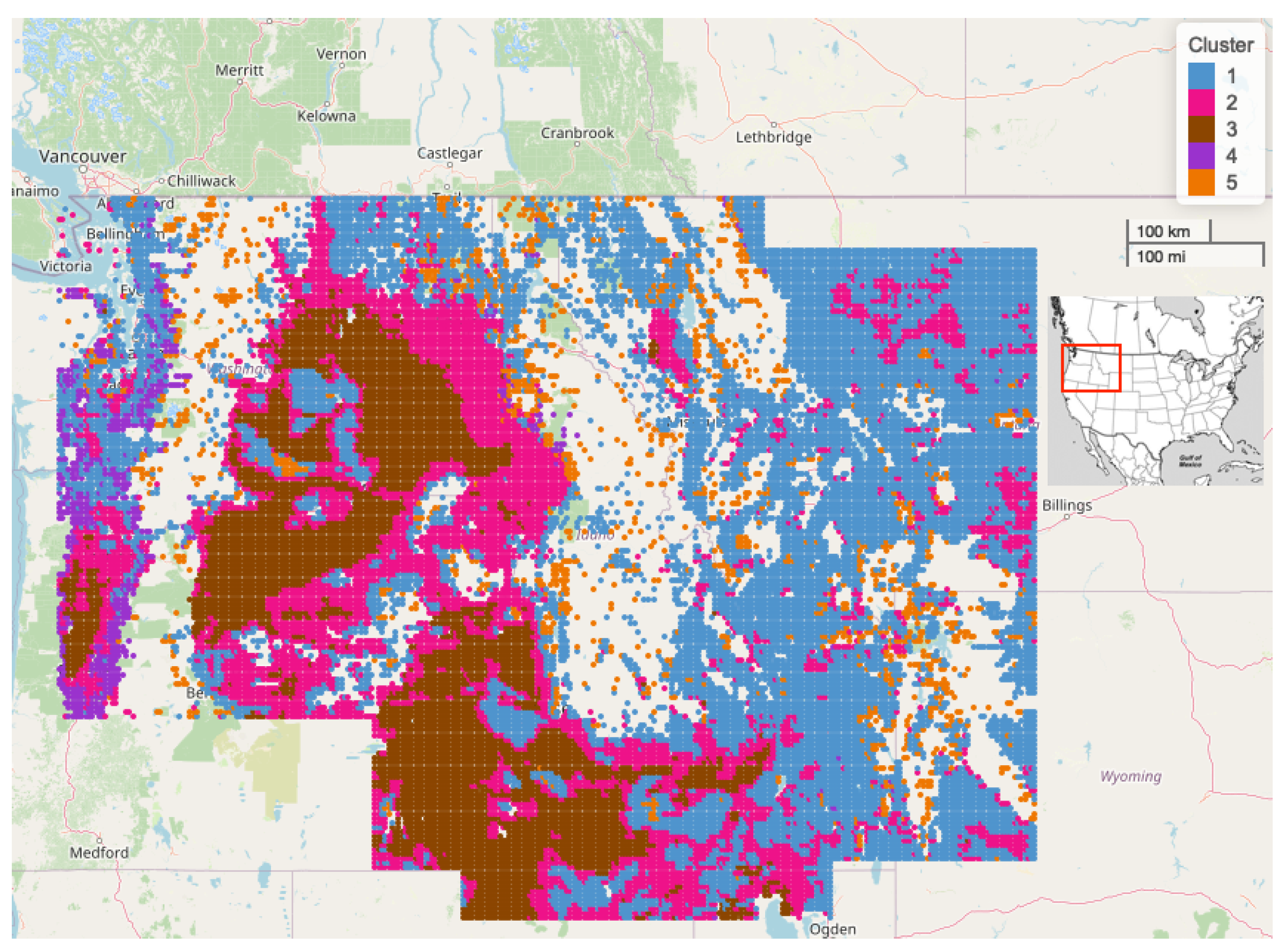
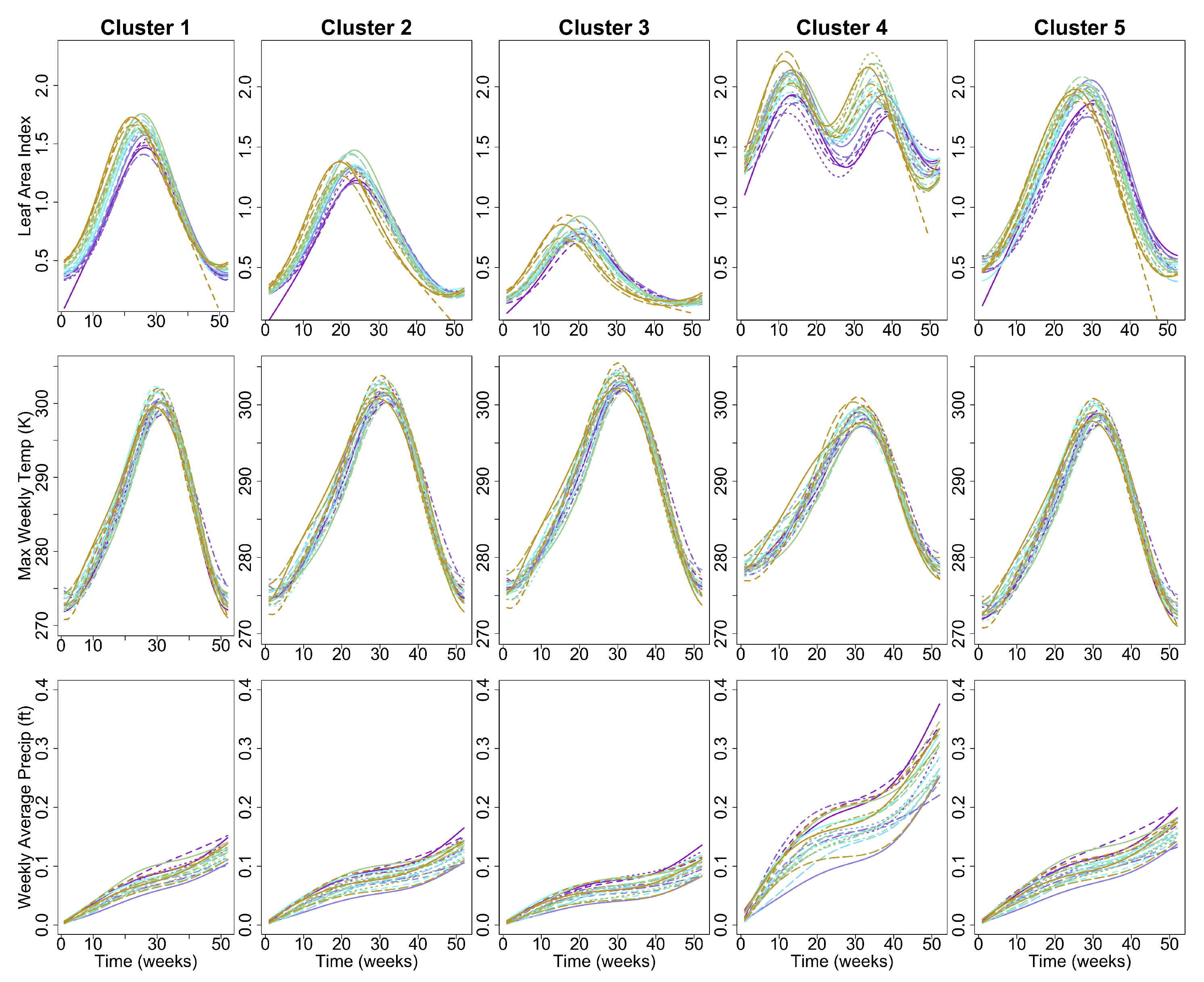
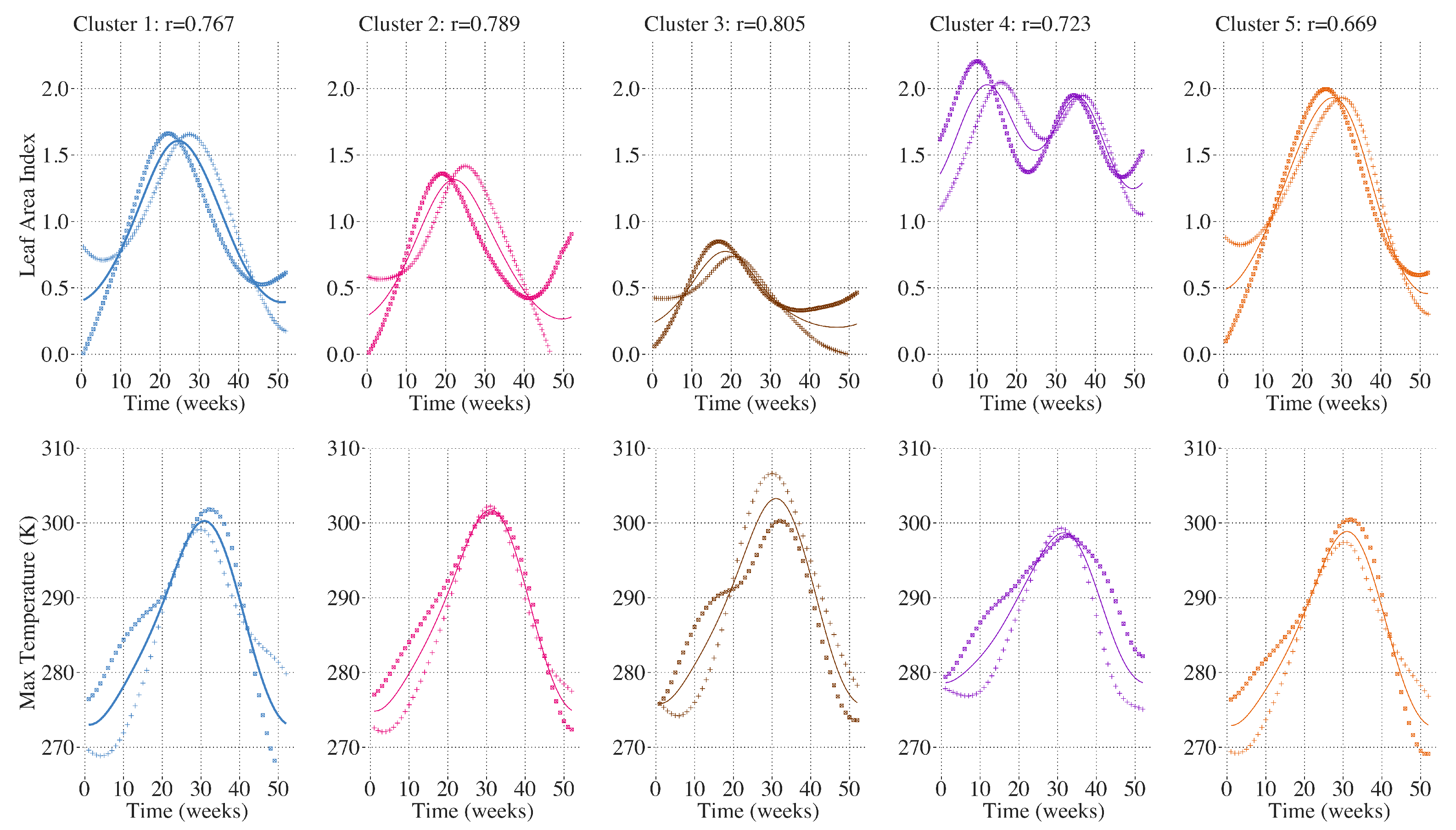
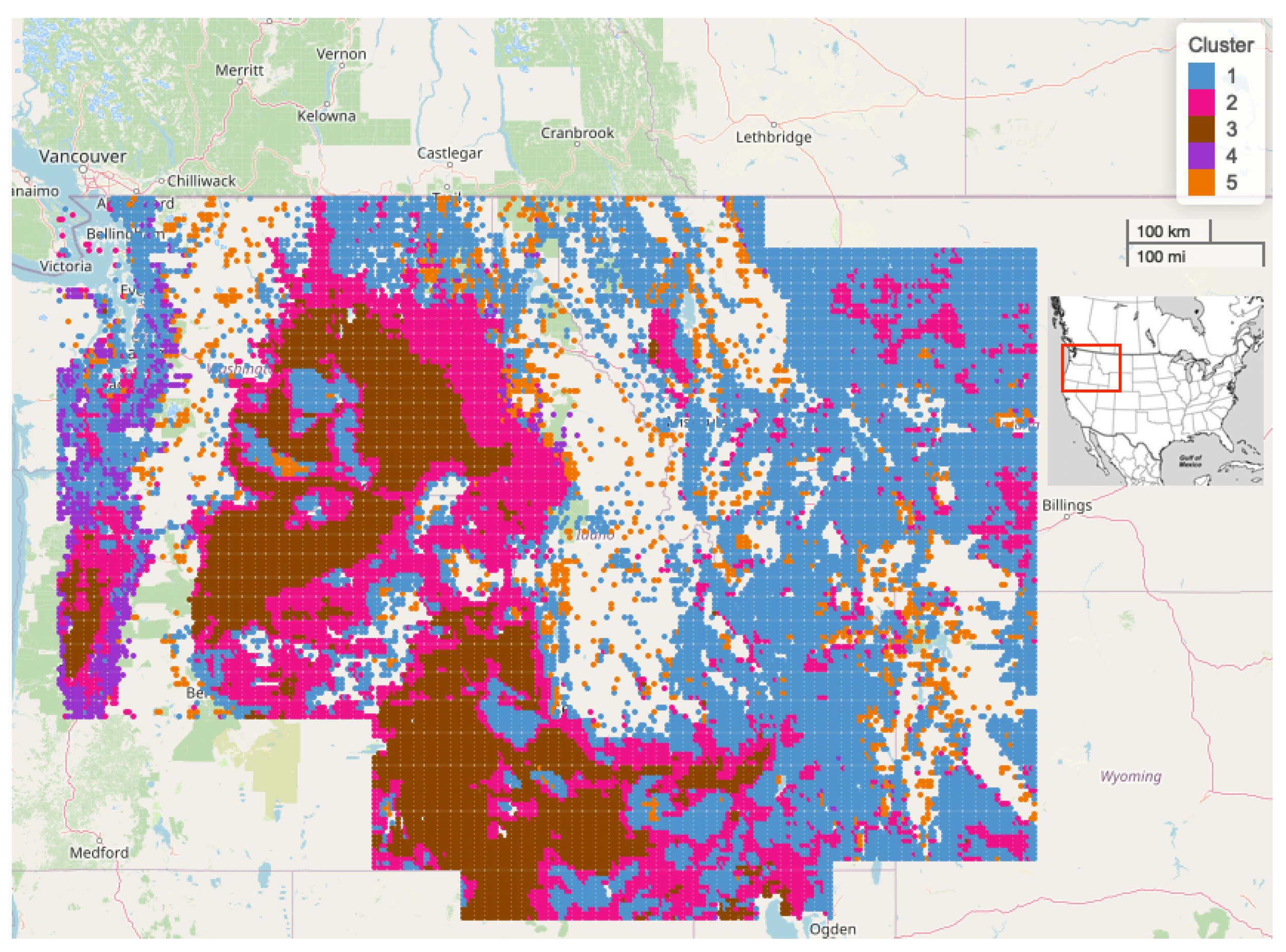
2.6. Spatial Clustering of Univariate High-Dimensional Functional Data
2.7. Ordinary Functional Kriging of the ERA-Interim
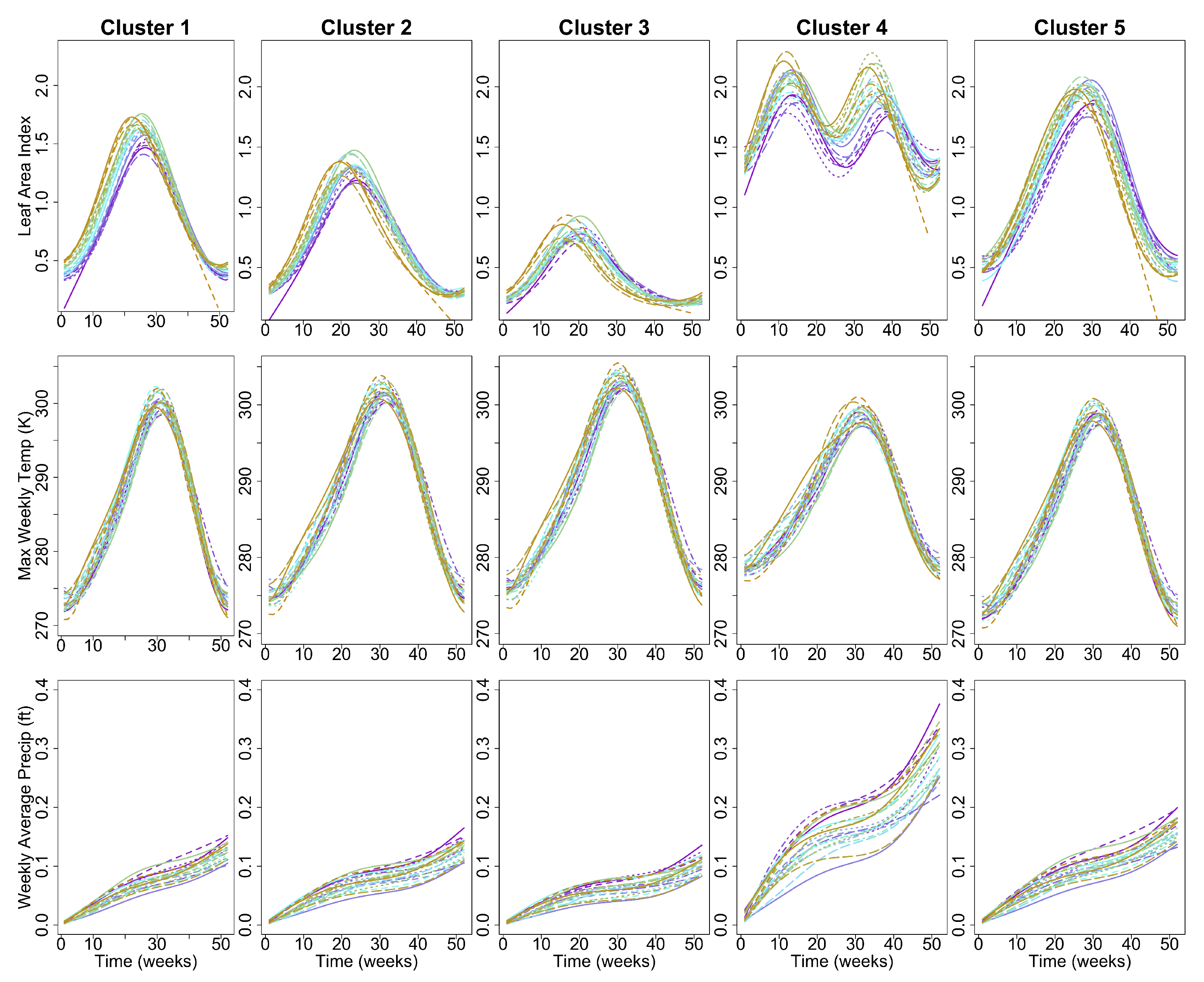
2.8. Deriving Regional Average Profiles
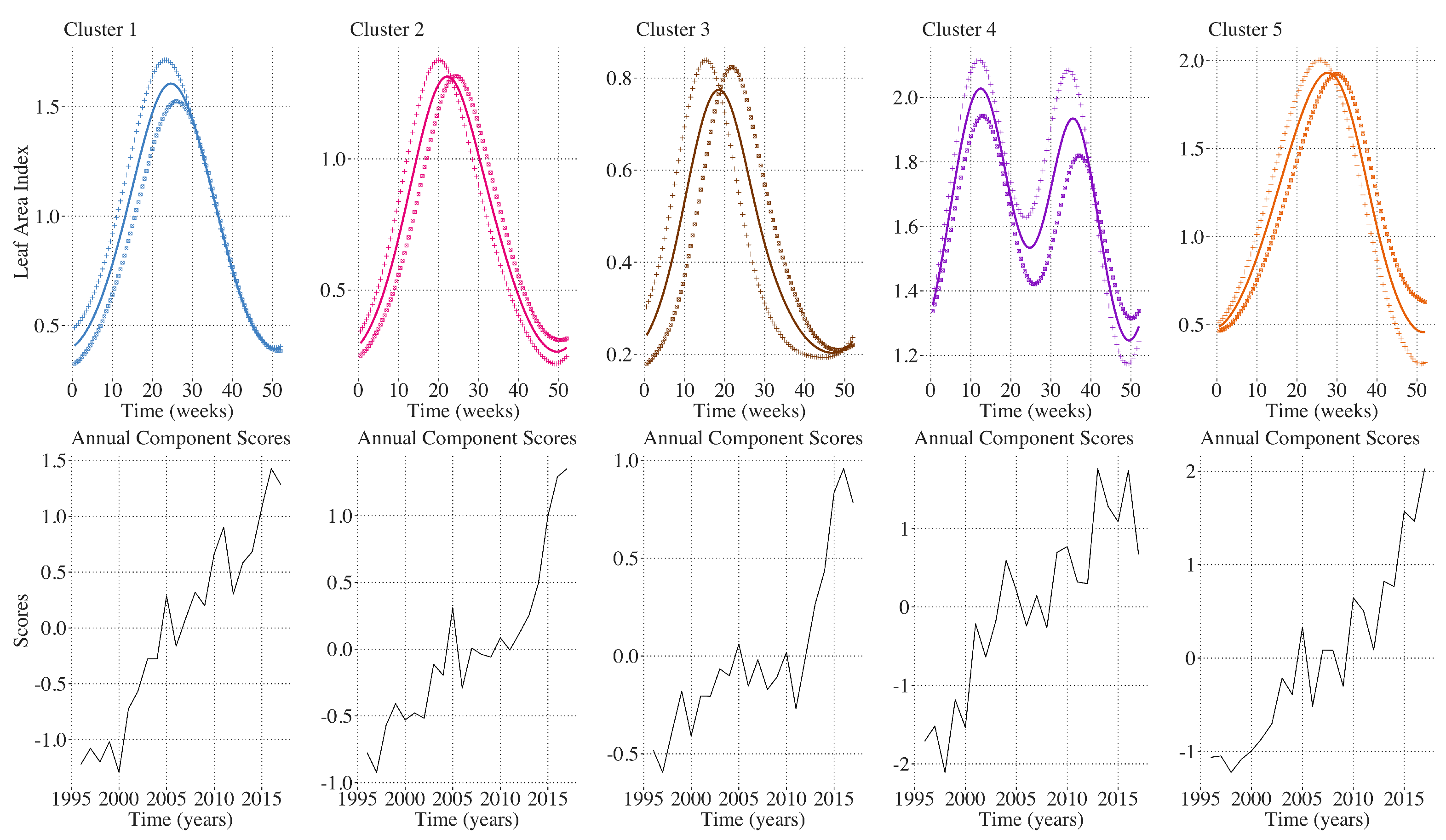
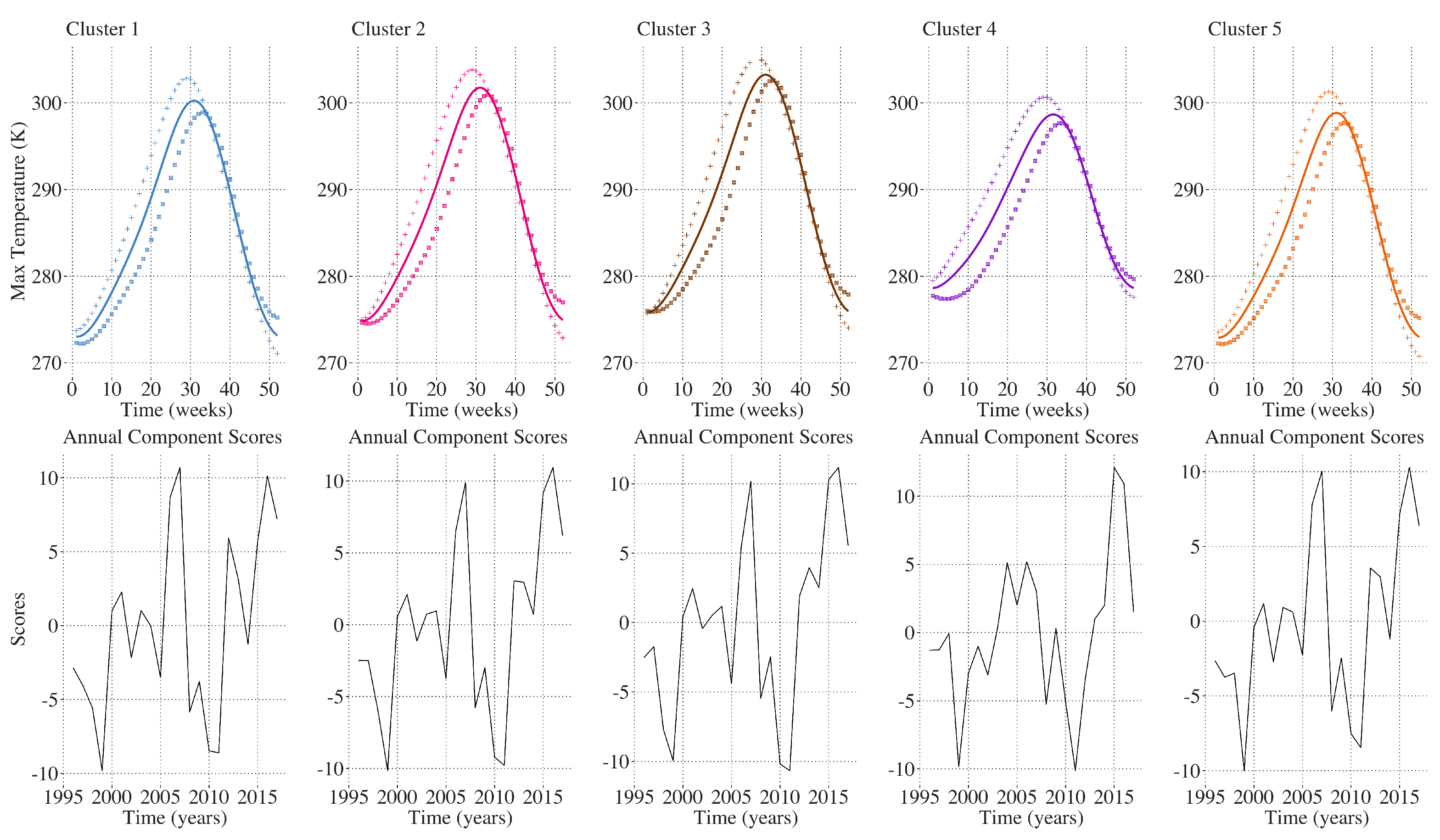
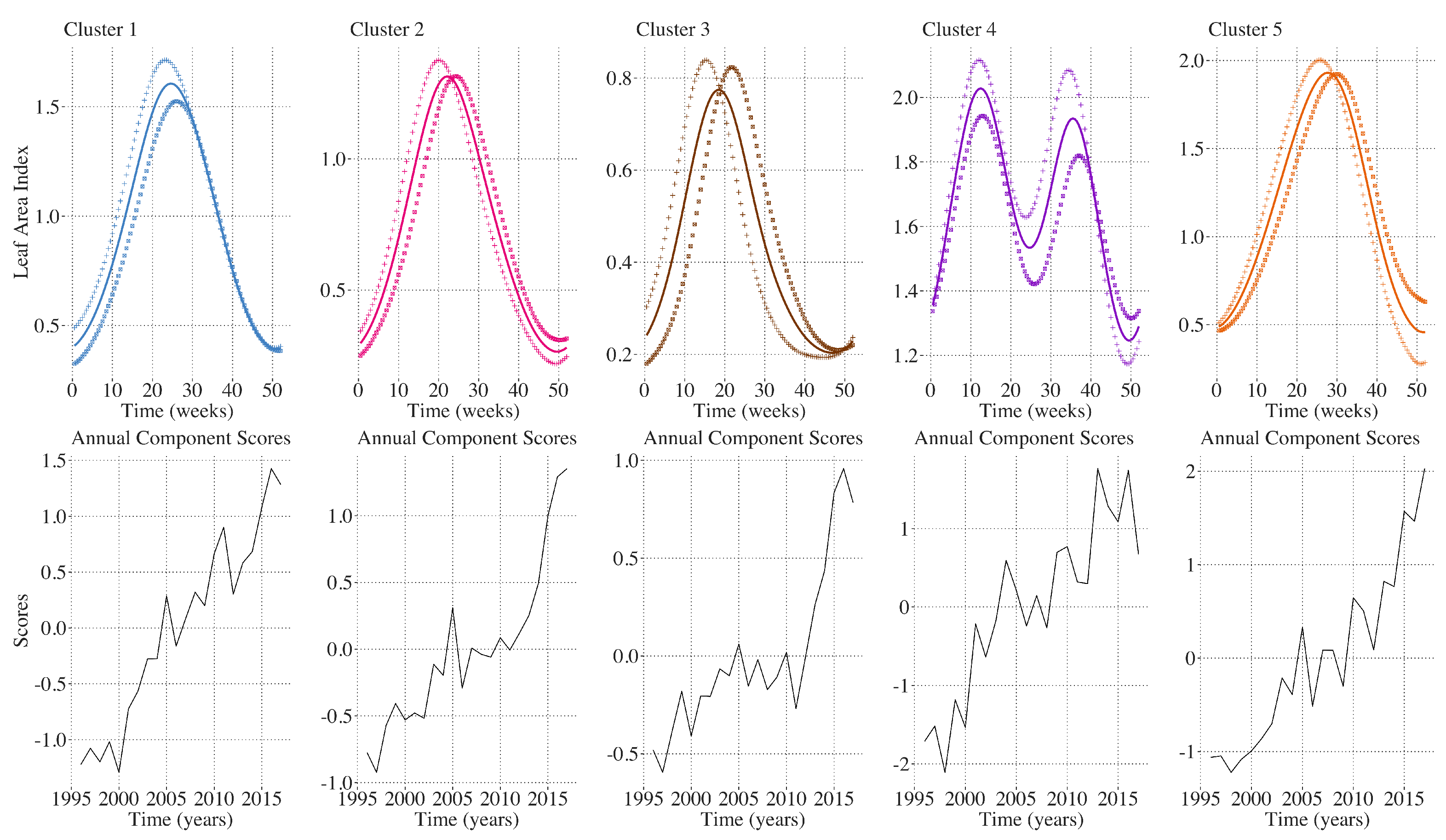
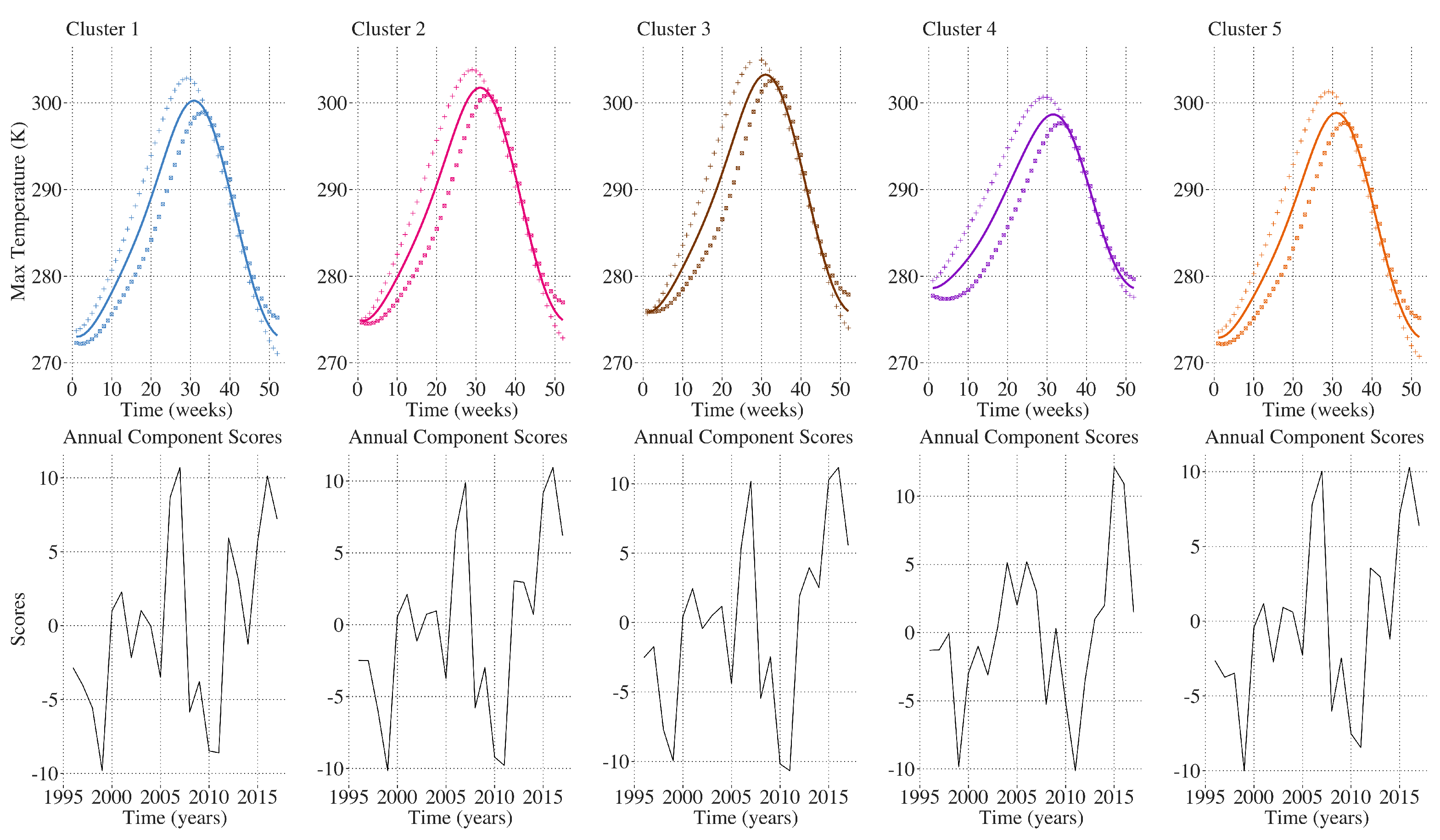
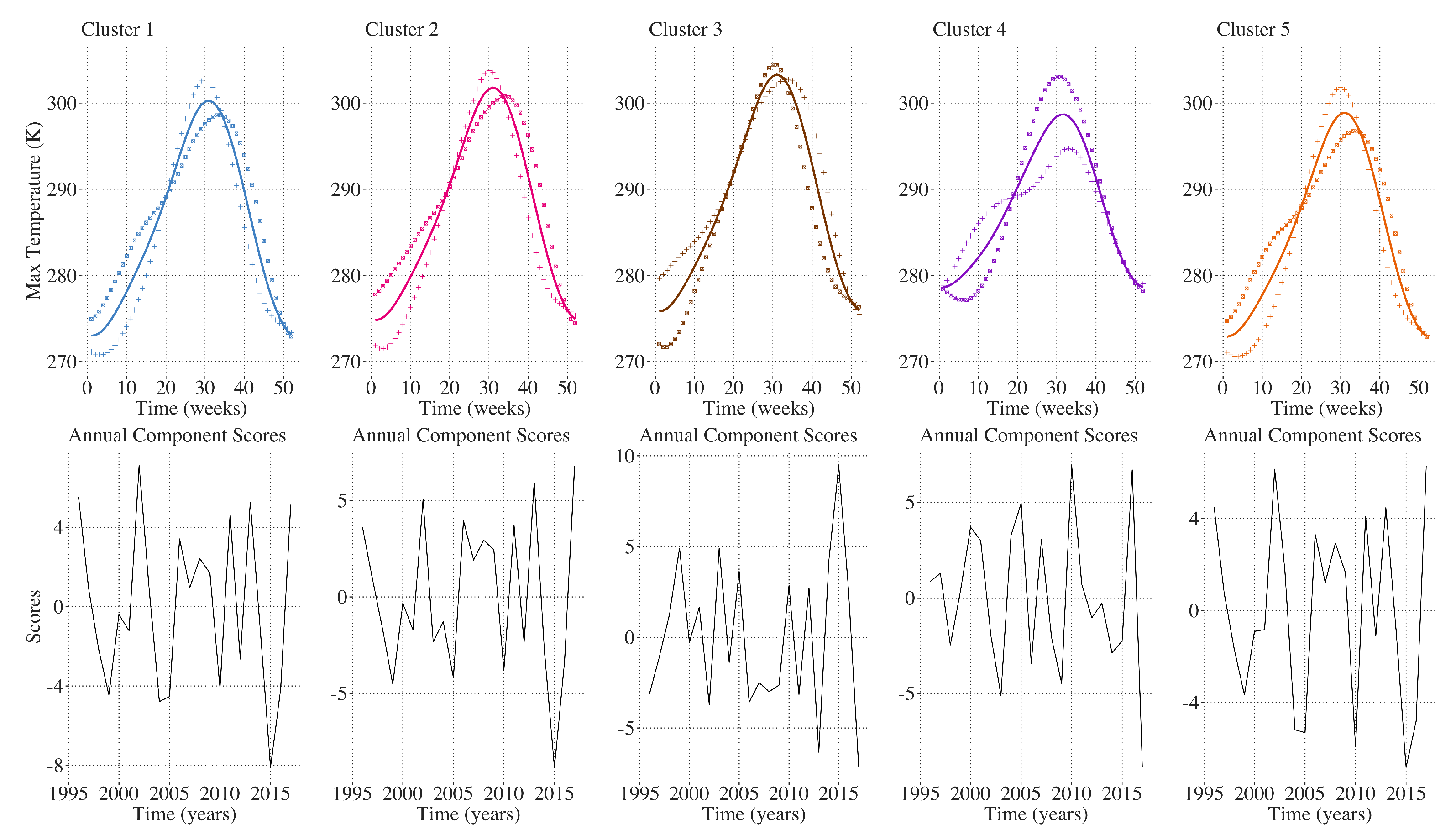
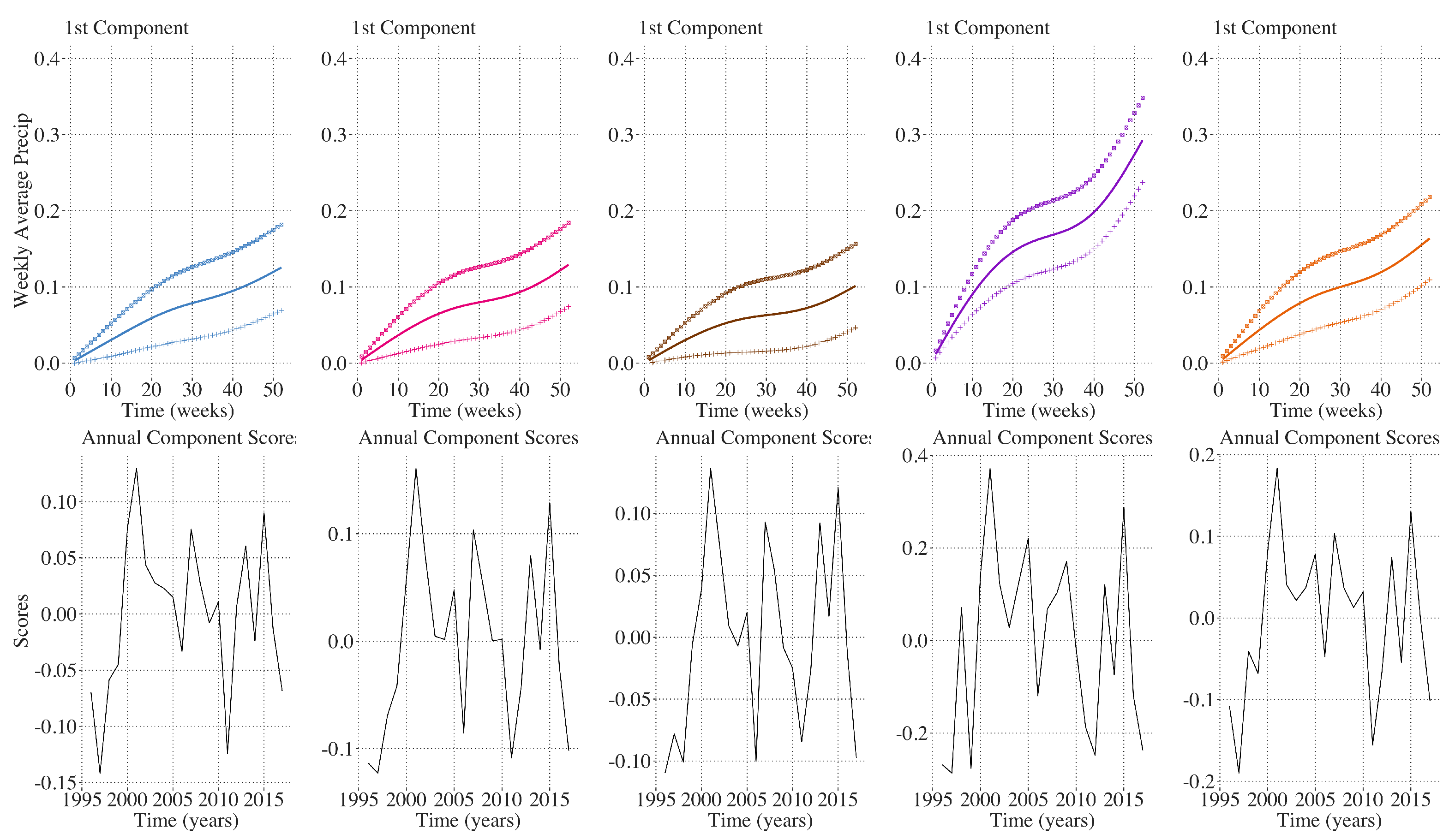
2.9. Inter-Annual Regional LAI and Climate Variation Monitoring
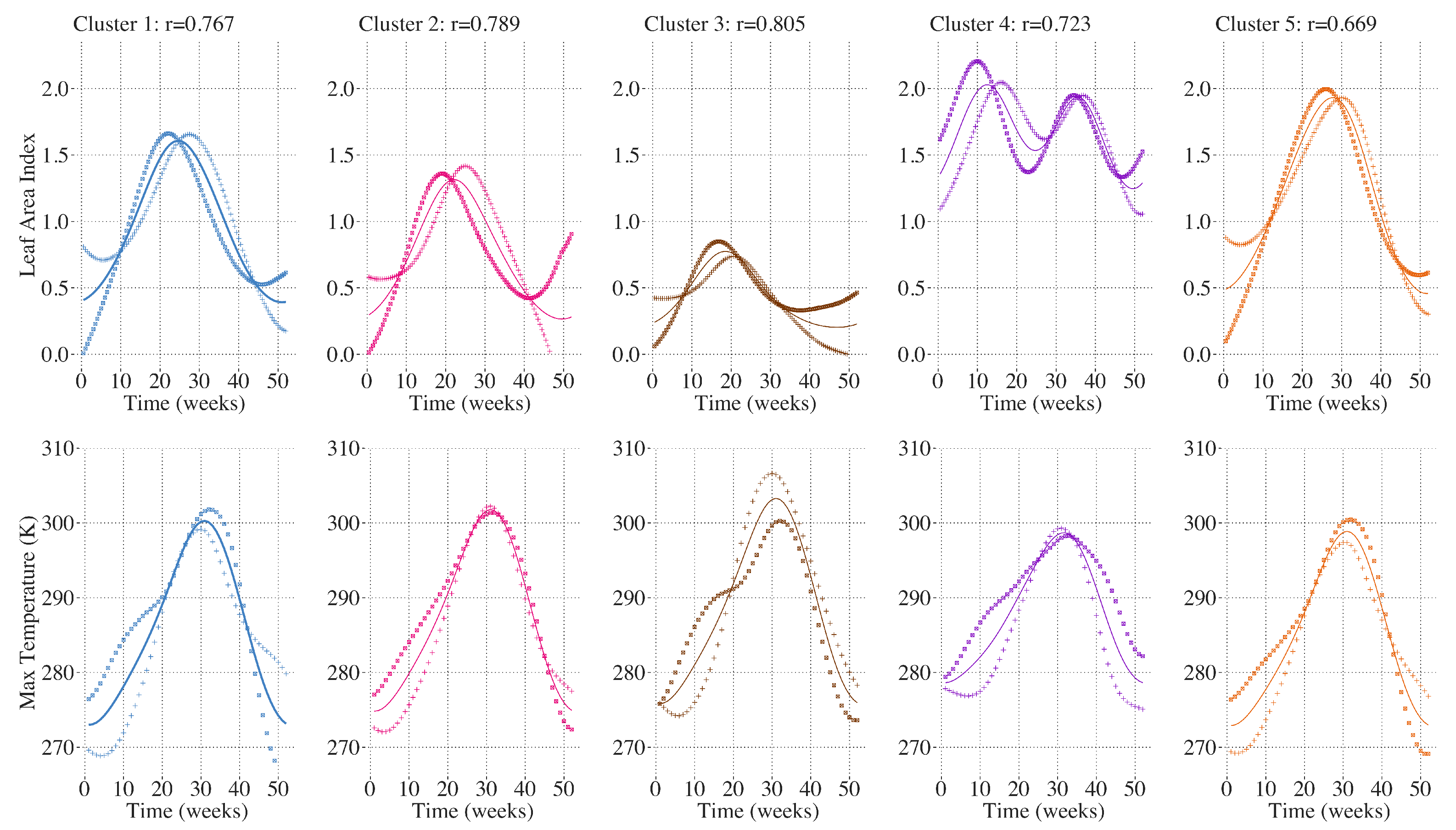
2.10. Inter-Annual Canonical Correlation Analysis between LAI and Climate Attributes
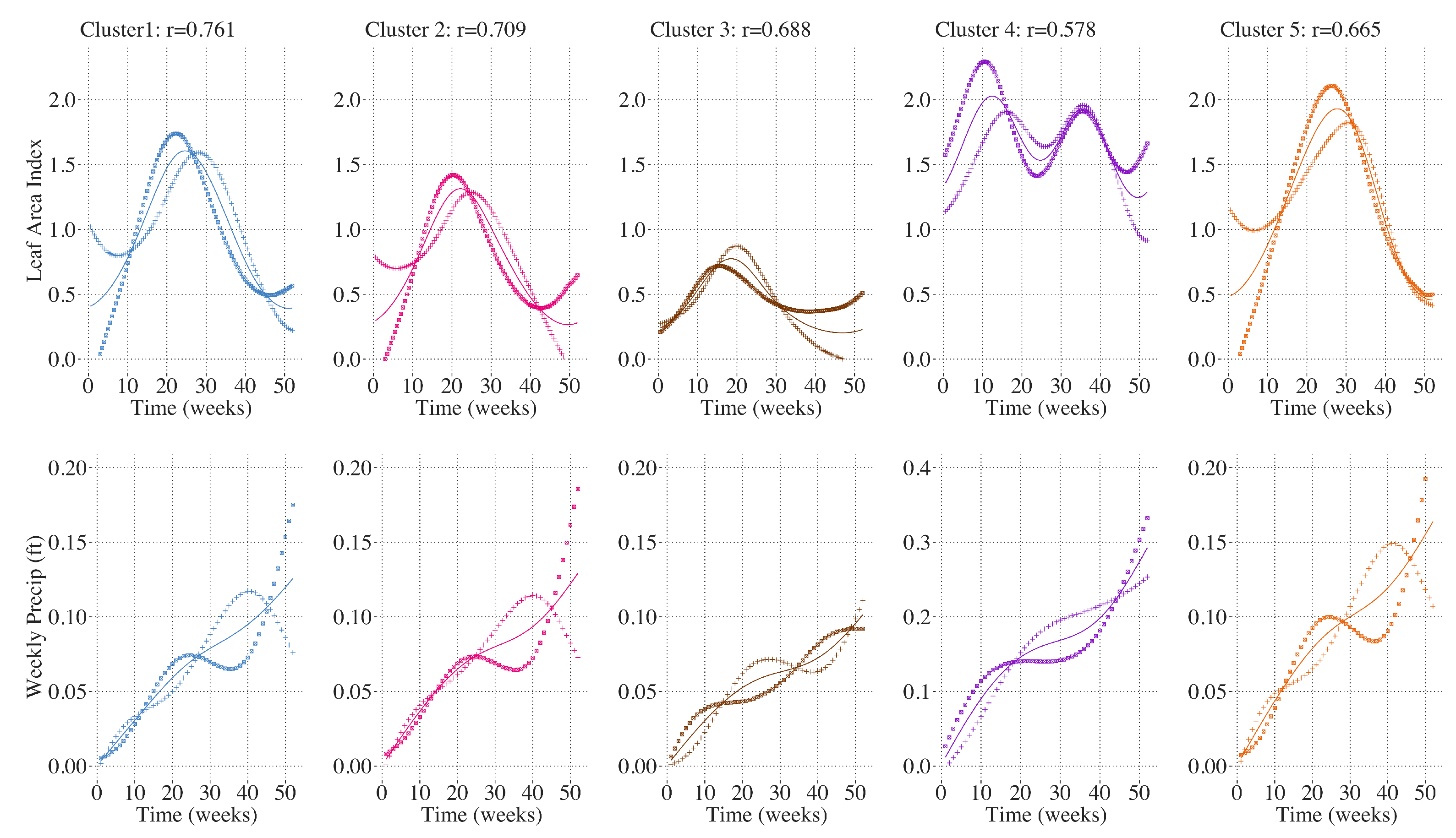
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AVHRR | Advanced Very High-Resolution Radiometer |
| CCA | Canonical Correlation Analysis |
| CDR | Climate Data Record |
| CRB | Columbia River Basin |
| DTM | Digital Terrain Model |
| DTW | Dynamic Time Warping |
| ECMWF | European Centre for Medium-Range Weather Forecasts |
| ECV | Essential Climate Variable |
| ENVR | Section on Statistics and the Environment |
| ERA | ECMWF Re-Analysis |
| FCCA | Functional Canonical Correlation Analysis |
| FDA | Functional Data Analysis |
| FPC | Functional Principal Component |
| FPCA | Funcitonal Principal Component Analysis |
| GCOS | Global Climate Observing System |
| GCV | Generalized Cross-Validation |
| GEODE | Global Earth Observing and Dynamics of Ecosystems |
| HPC | High-Performance Computing |
| LAI | Leaf Area Index |
| MODIS | Moderate-Resolution Imaging Spectroradiometer |
Appendix A
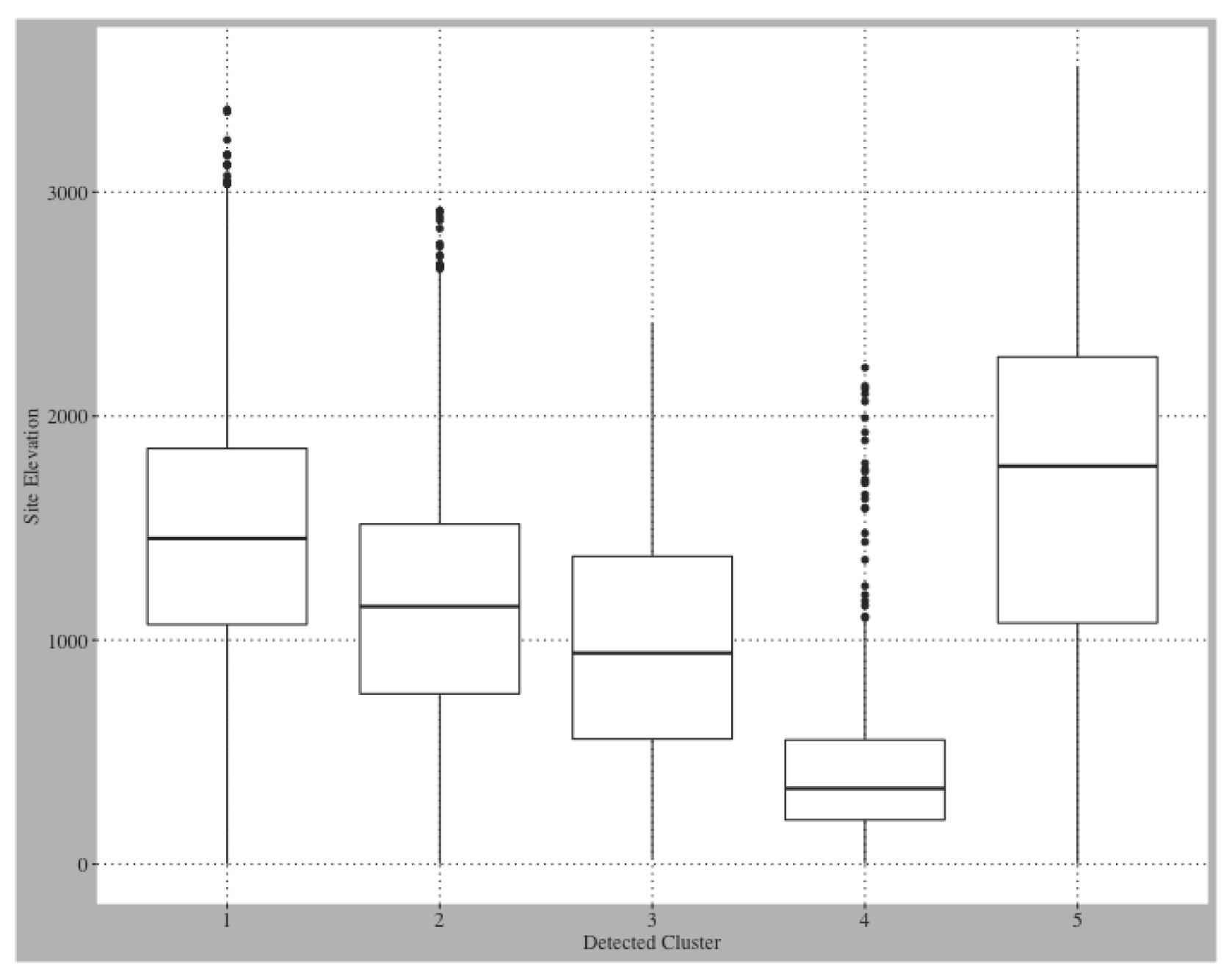
Appendix A.1. Preliminary Elevation Distribution Assessment by Cluster

Appendix A.2. Exploratory Applet Access
References
- Ustin, S.L.; Middleton, E.M. Current and near-term advances in Earth observation for ecological applications. Ecol. Proce. 2021, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X.; et al. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Bojinski, S.; Verstraete, M.; Peterson, T.C.; Richter, C.; Simmons, A.; Zemp, M. The Concept of Essential Climate Variables in Support of Climate Research, Applications, and Policy. Bull. Am. Meteorol. Soc. 2014, 95, 1431–1443. [Google Scholar] [CrossRef]
- Wu, M.; Schurgers, G.; Rummukainen, M.; Smith, B.; Samuelsson, P.; Jansson, C.; Siltberg, J.; May, W. Vegetation–climate feedbacks modulate rainfall patterns in Africa underfuture climate change. Earth Syst. Dyn. 2016, 7, 627–647. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Piao, S.; Li, L.Z.X.; Zhou, L.; Ciais, P.; Wang, T.; Li, Y.; Lian, X.; Wood, E.F.; Friedlingstein, P.; et al. Climate mitigation from vegetation biophysical feedbacks during the past three decades. Nat. Clim. Chang. 2017, 7, 432–436. [Google Scholar] [CrossRef]
- Schnase, J.L.; Lee, T.J.; Mattmann, C.A.; Lynnes, C.S.; Cinquini, L.; Ramirez, P.M.; Hart, A.F.; Williams, D.N.; Waliser, D.; Rinsl, P.; et al. Big Data Challenges in Climate Science: Improving the next-generation cyberinfrastructure. IEEE Geosci. Remote Sens. 2016, 4, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Berner, L.T.; Massey, R.; Jantz, P.; Forbes, B.C.; Macias-Fauria, M.; Myers-Smith, I.; Kumpula, T.; Gauthier, G.; Andreu-Hayles, L.; Gaglioti, B.V.; et al. Summer warming explains widespread but not uniform greening in the Arctic tundra biome. Nat. Commun. 2020, 11, 4621. [Google Scholar] [CrossRef]
- Sumida, A.; Watanabe, T.; Miyaura, T. Interannual variability of leaf area index of an evergreen conifer stand was affected by carry-over effects from recent climate conditions. Sci. Rep. 2018, 8, 13590. [Google Scholar] [CrossRef] [Green Version]
- Forzieri, G.; Duveiller, G. Evaluating the Interplay Between Biophysical Processes and Leaf Area Changes in Land Surface Models. J. Adv. Model. Earth Syst. 2018, 10, 1102–1126. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Fang, J.; Zhou, L.; Guo, Q.; Henderson, M.; Ji, W.; Li, Y.; Tao, S. Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. J. Geophys. Res. Atmos. 2003, 48. [Google Scholar] [CrossRef]
- Ramsay, J.O.; Silverman, B.W. Functional Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Ramsay, J.; Dalzell, C. Some Tools for Functional Data Analysis. J. R. Stat. Soc. Ser. (Methodol.) 1991, 53, 539–572. [Google Scholar] [CrossRef]
- Ullah, S.; Finch, C.F. Applications of functional data analysis: A systematic review. BMC Med. Res. Methodol. 2013, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesaresi, S.; Mancini, A.; Quattrini, G.; Casavecchia, S. Mapping Mediterranean Forest Plant Associations and Habitats with Functional Principal Component Analysis Using Landsat 8 NDVI Time Series. Remote Sens. 2020, 12, 1132. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Huang, S.; Huang, Q.; Wang, H.; Leng, G.; Fang, W. Time-Lagged Response of Vegetation Dynamics to Climatic and Teleconnection Factors. Catena 2020, 189, 104474. [Google Scholar] [CrossRef]
- Sebastian, D.E.; Ganguly, S.; Krishnaswam, J.; Duffy, K.; Nemani, R.; Ghosh, S. Multi-Scale Association between Vegetation Growth and Climate in India: A Wavelet Analysis Approach. Remote Sens. 2019, 11, 2703. [Google Scholar] [CrossRef] [Green Version]
- Ghaderpour, E.; Vujadinovic, T.; Hassan, Q.K. Application of the Least-Squares Wavelet Software in Hydrology: Athabasca River Basin. J. Hydrol. Reg. Stud. 2021, 36, 100847. [Google Scholar] [CrossRef]
- Claverie, M.; Matthews, J.L.; Vermote, E.F.; Justice, C.O. A 30+ Year AVHRR LAI and FAPAR Climate Data Record: Algorithm Description and Validation. Remote Sens. 2016, 8, 263. [Google Scholar] [CrossRef] [Green Version]
- Claverie, M.; Vermote, E. NOAA Climate Data Record (CDR) of Leaf Area Index (LAI) and Fraction of Absorbed Photosynthetically Active Radiation (FAPAR), Version 4; NOAA National Centers for Environmental Information: Boulder, CO, USA, 2014. [CrossRef]
- Dee, D.P.; Uppala, S.M.; Simmons, A.J.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.A.; Balsamo, G.; Bauer, P. The ERA-Interim reanalysis: Configuration and performance of the data assimilation system. QJR Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- MacDonald, Dettwiler and Associates Ltd. (MDA). BaseVue 2013. 2014. Available online: http://www.arcgis.com/home/item.html?id=1770449f11df418db482a14df4ac26eb (accessed on 15 March 2021).
- Gesch, D.; Oimoen, M.; Greenlee, S.; Nelson, C.; Steuck, M.; Tyler, D. The National Elevation Data Set. Photogramm. Eng. Remote Sens. 2002, 68, 5–11. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 10 December 2021).
- Baptiste, A. gridExtra: Miscellaneous Functions for “Grid” Graphics. 2015. R Package Version 2.0.0. Available online: http://CRAN.R-project.org/package=gridExtra (accessed on 10 December 2021).
- Gervini, D.; Khanal, M. Exploring patterns of demand in bike sharing systems via replicated point process models. J. R. Stat. Soc. Ser. Appl. Stat. 2019, 68, 585–602. [Google Scholar] [CrossRef]
- Maechler, M. Cluster Analysis Basics and Extensions. 2021. R Package Version 2.1.1. Available online: https://CRAN.R-project.org/package=cluster (accessed on 1 February 2021).
- Froese, V.; Jain, B.; Niedermeier, R.; Renken, M. Comparing temporal graphs using dynamic time warping. Soc. Netw. Anal. Min. 2020, 10, 50. [Google Scholar] [CrossRef]
- Gold, O.; Sharir, M. Dynamic Time Warping and Geometric Edit Distance. ACM Trans. Algorithms 2018, 14, 50. [Google Scholar] [CrossRef]
- Whetten, A.B. Localized Mutual Information Monitoring of Pairwise Associations in Animal Movement. arXiv 2021, arXiv:2111.10628. [Google Scholar]
- Ramon, G. Spatial Prediction for Function Value Data. 2020. R Package Version 2.0. Available online: https://CRAN.R-project.org/package=geofd (accessed on 20 March 2021).
- Ramon, G. An R Package for Function-Valued Geostatistical Prediction. Rev. Colomb. EstadíStica Diciembre 2012, 35, 385–407. [Google Scholar]
- Cheng, J. Leaflet: Create Interactive Web Maps with the JavaScript ’Leaflet’ Library. 2019. R Package Version 2.0.3. Available online: https://CRAN.R-project.org/package=leaflet (accessed on 1 October 2021).
- Whetten, A.B. Characterizing Clustering Models of High-dimensional Remotely Sensed Data Using Subsampled Field-subfield Spatial Cross-validated Random Forests. Int. J. Geospat. Environ. Res. 2021, 8, 4. Available online: https://dc.uwm.edu/ijger/vol8/iss3/4 (accessed on 11 November 2021).
- Fensholt, R.; Sandholt, I. Evaluation of MODIS and NOAA AVHRR vegetation indices with in situ measurements in a semi-arid environment. Int. J. Remote Sens. 2005, 26, 2561–2594. [Google Scholar] [CrossRef]
- Steven, M.; Malthus, T.J.; Baret, F.; Xu, H.; Chopping, M.J. Intercalibration of vegetation indices from different sensor systems. Remote Sens. Environ. 2003, 88, 412–422. [Google Scholar] [CrossRef]
- Hansen, M.C.; DeFries, R.S.; Townshend, J.R.G.; Sohlberg, R.; Dimiceli, C.; Carroll, M. Towards an operational MODIS continuous field of percent tree cover algorithm: Examples using AVHRR and MODIS data. Remote Sens. Environ. 2002, 83, 303–319. [Google Scholar] [CrossRef]
- Fensholt, R.; Rasmussen, K.; Nielsen, T.T.; Mbow, C. Evaluation of earth observation based long term vegetation trends—Intercomparing NDVI time series trend analysis consistency of Sahel from AVHRR GIMMS, Terra MODIS and SPOT VGT data. Remote Sens. Environ. 2009, 113, 1886–1898. [Google Scholar] [CrossRef]
- Cihlar, J.; Tcherednichenko, I.; Latifovic, R.; Li, Z.; Chen, J. Impact of variable atmospheric water vapor content on AVHRR data corrections over land. IEEE Trans. Geosci. Remote Sens. 2001, 39, 173–180. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Fensholt, R.; Langanke, T.; Rasmussen, K.; Reenberg, A.; Prince, S.D.; Tucker, C.; Scholes, R.J.; Lee, Q.B.; Bondeau, A.; Eastman, R.; et al. Greenness in semi-arid areas across the globe 1981–2007—An Earth Observing Satellite based analysis of trends and drivers. Remote Sens. Environ. 2012, 121, 144–158. [Google Scholar] [CrossRef]
- Mao, J.; Ribes, A.; Yan, B.; Shi, X.; Thortin, P.E.; Seferian, R.; Ciais, P.; Myneni, R.B.; Douville, H. Human-induced greening of the northern extratropical land surface. Nat. Clim. Chang. 2016, 6, 959–963. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Fitchett, J.M.; Grab, S.W.; Thompson, D.I. Plant phenology and climate change: Progress in methodological approaches and application. Prog. Phys. Geogr. Earth Environ. 2015, 39, 460–482. [Google Scholar] [CrossRef]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.H.; Ciasia, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z. Leaf onset in the northern hemisphere triggered by daytime temperature. Nat. Commun. 2015, 6, 6911. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Du, J.; Zhao, W.; Yang, J.; Chen, L.; Zhu, X.; Chang, X.; Liu, H. Assessing temperature sensitivity of subalpine shrub phenology in semi-arid mountain regions of China. Agric. For. Meteorol. 2015, 213, 42–52. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Alm-Kubler, K.; Bissolli, P.; Braslavska, O.; Briede, A. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Harrington, C.A.; Gould, P.J.; St Clair, B. Modeling the effects of winter environment on dormancy release of Douglas-fir. For. Ecol. Manag. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Sherry, R.A.; Zhou, X.; Gu, S.; Arnone, J.A., III; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y. Divergence of reproductive phenology under climate warming. Proc. Natl. Acad. Sci. USA 2007, 104, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ry, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, X.; Roy, J.; Sonie, L.; Chuine, I. Changes in leaf phenology of three European oak species in response to experimental climate change. New Phytol. 2010, 186, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W.; Medina-Elizade, M. Global temperature change. Proc. Natl. Acad. Sci. USA 2006, 103, 14288–14293. Available online: www.pnas.orgcgidoi10.1073pnas.0606291103 (accessed on 11 November 2021). [CrossRef] [Green Version]
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Chang. Biol. 2006, 12, 343–351. [Google Scholar] [CrossRef]
- Robeson, S.M. Trends in time-varying percentiles of daily minimum and maximum temperature over North America. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef] [Green Version]
- Adole, T.; Dash, J.; Rodriguez-Galiano, V.; Atkinson, P.M. Photoperiod controls vegetation phenology across Africa. Commun. Biol. 2019, 2, 391. [Google Scholar] [CrossRef] [Green Version]
- Ghelardini, L.; Santini, A.; Black-Samuelsson, S.; Myking, T.; Falusi, M. Bud dormancy release in elm (Ulmus spp.) clones—A case study of photoperiod and temperature responses. Tree Physiol. 2010, 30, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Jolly, W.M.; Nemani, R.; Running, S.W. A generalized, bioclimatic index to predict foliar phenology in response to climate. Glob. Chang. Biol. 2005, 11, 619–632. [Google Scholar] [CrossRef]
- Garonna, I.; de Jong, R.; Stockli, R.; Schmind, B.; Schenkel, D.; Schimel, D.; Schaepman, M.E. Shifting relative importance of climatic constraints on land surface phenology. Environ. Res. Lett. 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Shutova, E.; Wielgolaski, F.E.; Karlsen Makarova, O.; Berlina, N.; Filimonova, T.; Haraldson, E.; Aspholm, P.E.; Flo, L.; Hogda, K.A. Growing seasons of Nordic mountain birch in northernmost Europe as indicated by long-term field studies and analyses of satellite images. Int. J. Biometeorol. 2006, 51, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.M.; Miller-Rushing, A.J.; Inouye, W. Changes in snowmelt date and summer precipitation affect the flowering phenology of Erythronium grandiflorum (glacier lily; Liliaceae). Am. J. Bot. 2010, 97, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Hänninen, H.; Slaney, M.; Linder, S. Dormancy release of Norway spruce under climatic warming: Testing ecophysiological models of bud burst with a whole-tree chamber experiment. Tree Physiol. 2007, 27, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohue, R.J.; Roderick, R.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef] [Green Version]
- Haverd, V.; Smith, B.; Canadell, J.G.; Cuntz, M.; Mikaloff-Fletcher, S.; Farquhar, G.; Woodgate, W.; Briggs, P.R.; Trudinger, C.M. Higher than expected CO2 fertilization inferred from leaf to global observations. Glob. Chang. Biol. 2020, 26, 2390–2402. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
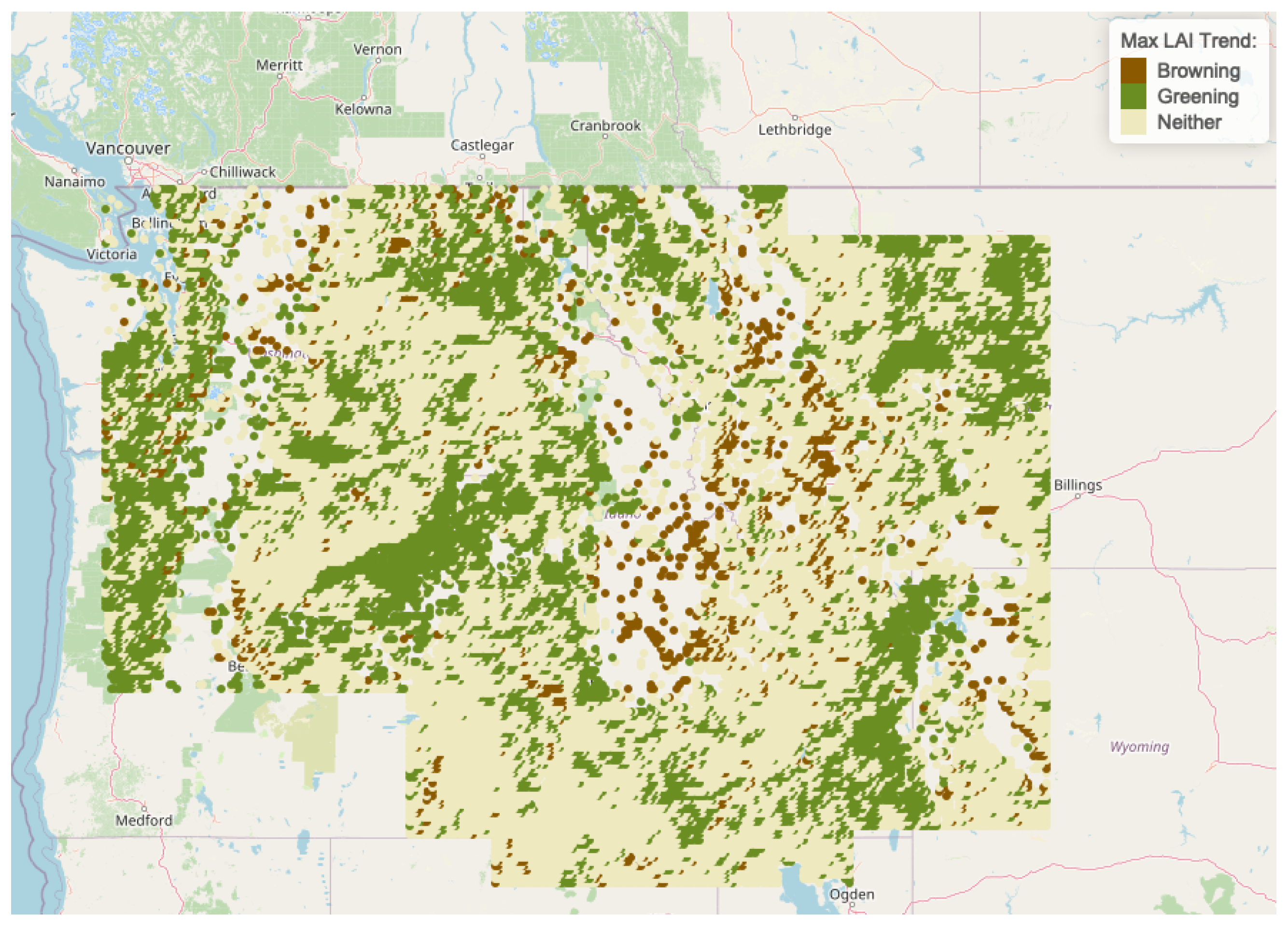
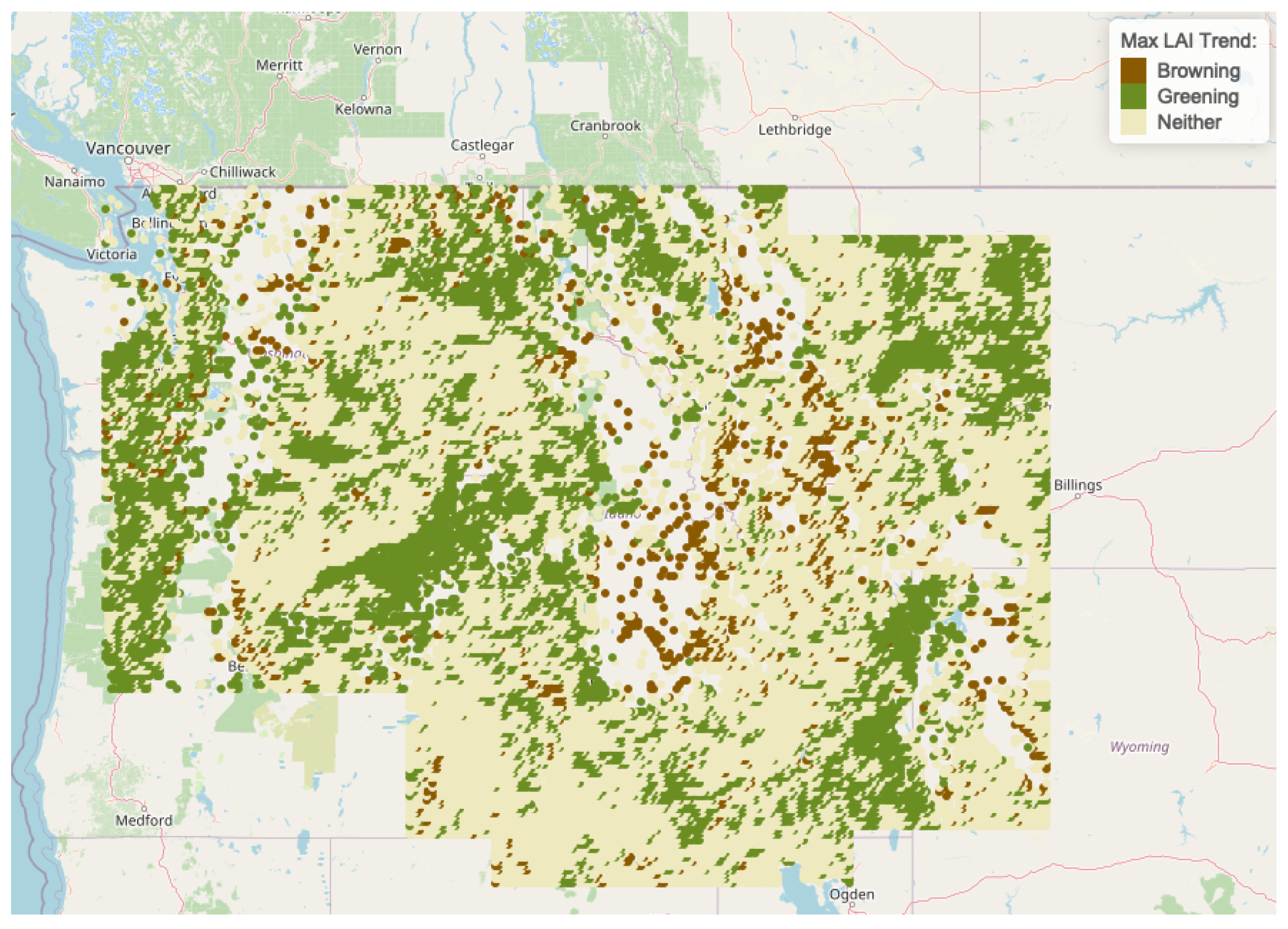
| Regions | Freq | Browning | Greening | Neither |
|---|---|---|---|---|
| Cluster 1 | 12,548 | 900 | 4087 | 7561 |
| Cluster 2 | 6737 | 104 | 2646 | 3987 |
| Cluster 3 | 5593 | 82 | 790 | 4721 |
| Cluster 4 | 742 | 5 | 517 | 220 |
| Cluster 5 | 1571 | 207 | 547 | 817 |
| Regions | Earlier | Later | Neither |
|---|---|---|---|
| Cluster 1 | 3885 | 157 | 8506 |
| Cluster 2 | 1398 | 119 | 5220 |
| Cluster 3 | 1049 | 60 | 4484 |
| Cluster 4 | 82 | 102 | 558 |
| Cluster 5 | 349 | 26 | 1196 |
| Regions | Freq | Max Diss | Avg Diss | Diameter | Separation |
|---|---|---|---|---|---|
| Cluster 1 | 12,548 | 0.6775 | 0.0744 | 1.0174 | 0.0025 |
| Cluster 2 | 6737 | 0.6746 | 0.0730 | 1.0496 | 0.0025 |
| Cluster 3 | 5593 | 0.8158 | 0.0956 | 1.1354 | 0.0033 |
| Cluster 4 | 742 | 0.9487 | 0.3087 | 1.2693 | 0.1110 |
| Cluster 5 | 1571 | 0.8639 | 0.2350 | 1.1874 | 0.0035 |
| Regions | Freq | Prop Agriculture | Prop Scrub | Prop Evergreen | Med Elev (m) |
|---|---|---|---|---|---|
| Cluster 1 | 12,548 | 0.130 | 0.293 | 0.238 | 1451.1 |
| Cluster 2 | 6737 | 0.161 | 0.455 | 0.155 | 1150.3 |
| Cluster 3 | 5593 | 0.174 | 0.667 | 0.010 | 942.0 |
| Cluster 4 | 742 | 0.009 | 0.241 | 0.481 | 337.8 |
| Cluster 5 | 1571 | 0.022 | 0.209 | 0.617 | 1776.8 |
| Regions | Attribute | Proportion of Variation | Component |
|---|---|---|---|
| Cluster 1 | LAI | 0.757 | 1st |
| Cluster 2 | LAI | 0.639 | 1st |
| Cluster 3 | LAI | 0.554 | 1st |
| Cluster 4 | LAI | 0.647 | 1st |
| Cluster 5 | LAI | 0.604 | 1st |
| Cluster 1 | Max Temp | 0.475 | 1st |
| Cluster 1 | Max Temp | 0.214 | 2nd |
| Cluster 2 | Max Temp | 0.481 | 1st |
| Cluster 2 | Max Temp | 0.210 | 2nd |
| Cluster 3 | Max Temp | 0.482 | 1st |
| Cluster 3 | Max Temp | 0.198 | 2nd |
| Cluster 4 | Max Temp | 0.446 | 1st |
| Cluster 4 | Max Temp | 0.233 | 2nd |
| Cluster 5 | Max Temp | 0.462 | 1st |
| Cluster 5 | Max Temp | 0.225 | 2nd |
| Cluster 1 | Precip | 0.903 | 1st |
| Cluster 2 | Precip | 0.914 | 1st |
| Cluster 3 | Precip | 0.917 | 1st |
| Cluster 4 | Precip | 0.917 | 1st |
| Cluster 5 | Precip | 0.912 | 1st |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whetten, A.B.; Demler, H.J. Detection of Multidecadal Changes in Vegetation Dynamics and Association with Intra-Annual Climate Variability in the Columbia River Basin. Remote Sens. 2022, 14, 569. https://doi.org/10.3390/rs14030569
Whetten AB, Demler HJ. Detection of Multidecadal Changes in Vegetation Dynamics and Association with Intra-Annual Climate Variability in the Columbia River Basin. Remote Sensing. 2022; 14(3):569. https://doi.org/10.3390/rs14030569
Chicago/Turabian StyleWhetten, Andrew B., and Hannah J. Demler. 2022. "Detection of Multidecadal Changes in Vegetation Dynamics and Association with Intra-Annual Climate Variability in the Columbia River Basin" Remote Sensing 14, no. 3: 569. https://doi.org/10.3390/rs14030569
APA StyleWhetten, A. B., & Demler, H. J. (2022). Detection of Multidecadal Changes in Vegetation Dynamics and Association with Intra-Annual Climate Variability in the Columbia River Basin. Remote Sensing, 14(3), 569. https://doi.org/10.3390/rs14030569