

The Influence of Satellite-Derived Environmental and Oceanographic Parameters on Marine Turtle Time at Surface in the Gulf of Mexico

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
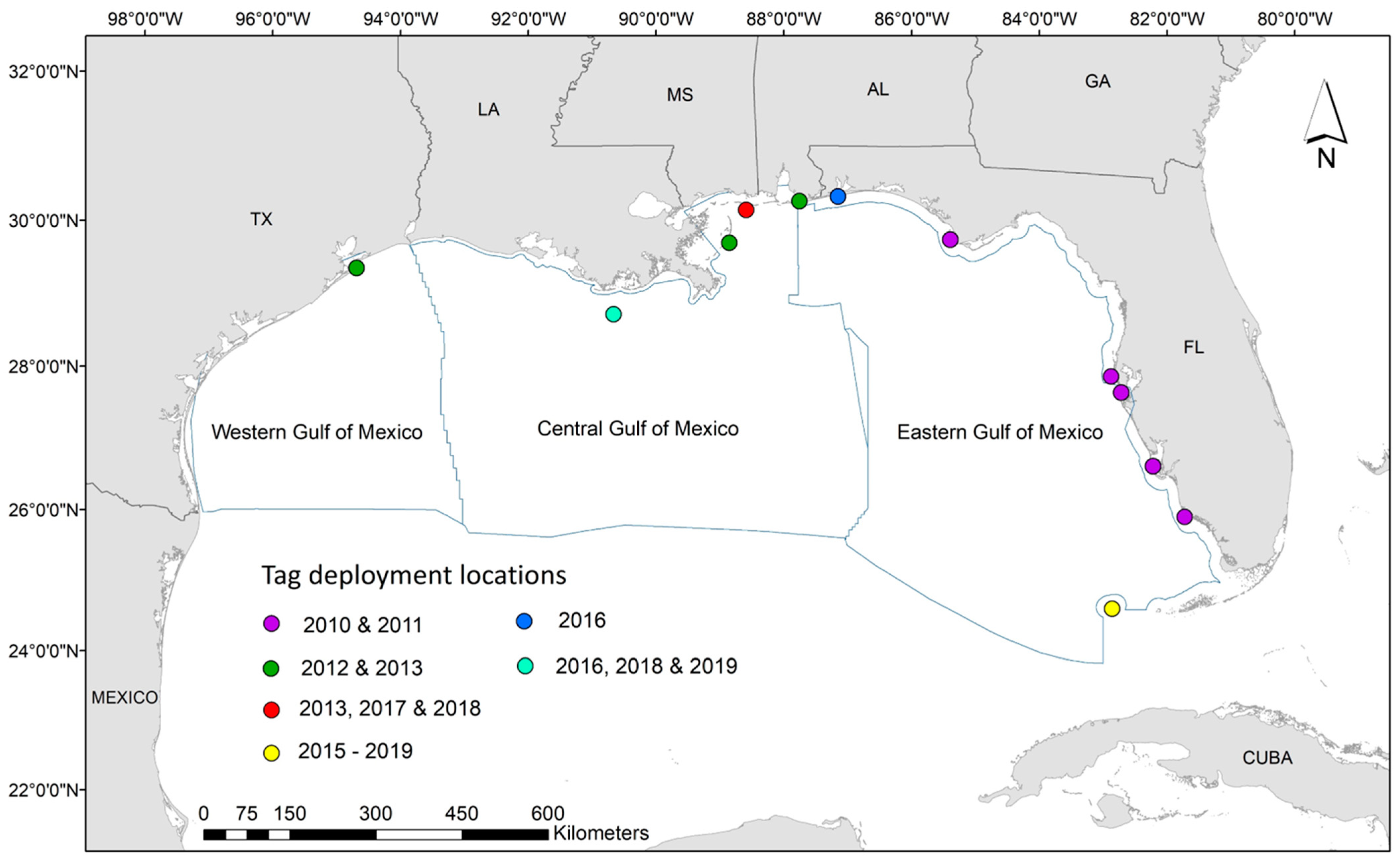
2.1. Data Collection
2.2. Linking Spatial and Dive Data
2.3. Environmental and Oceanographic Variables
2.4. Quantifying Environmental Influence on Time at Surface
3. Results
3.1. Data Collection
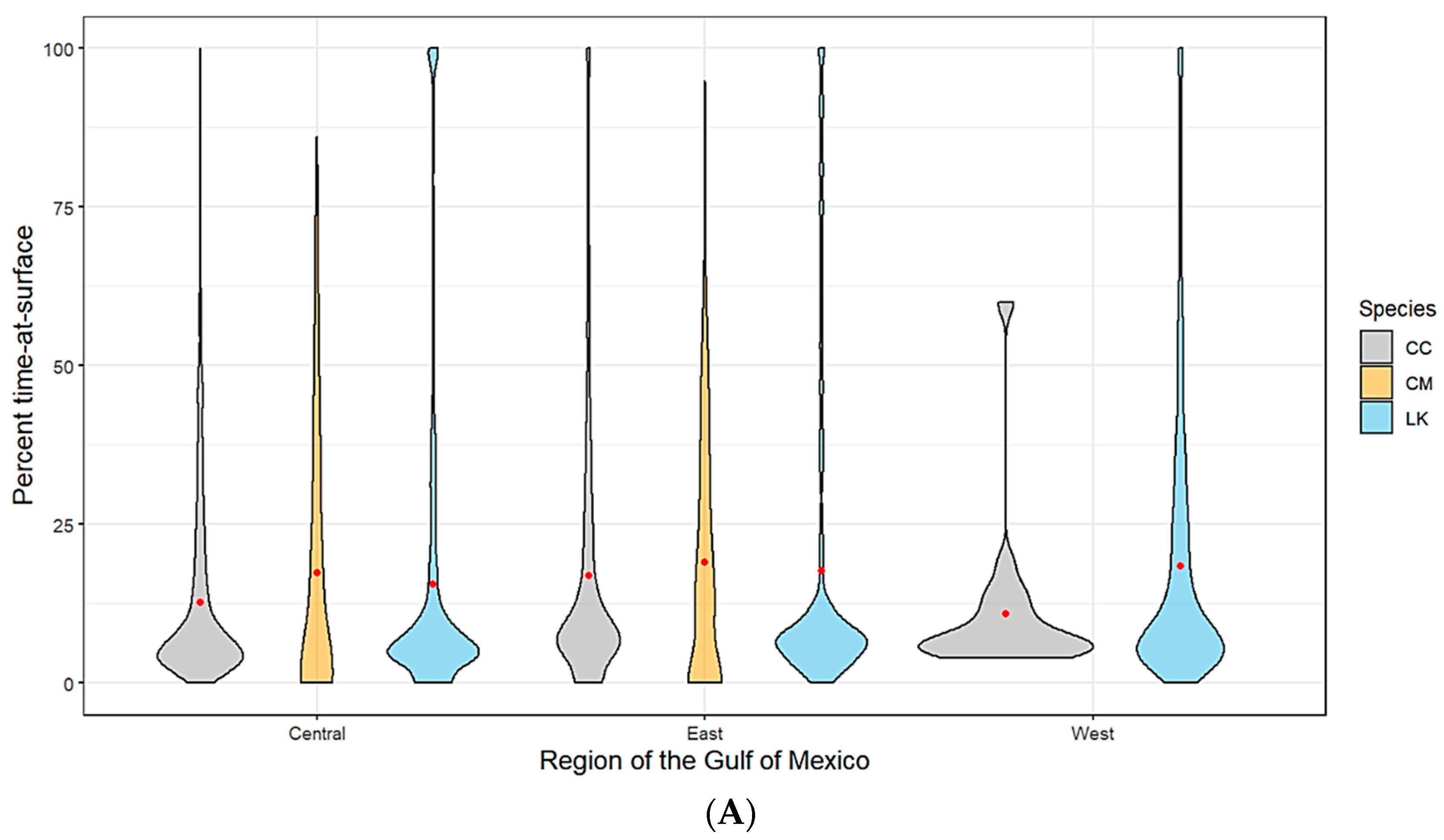
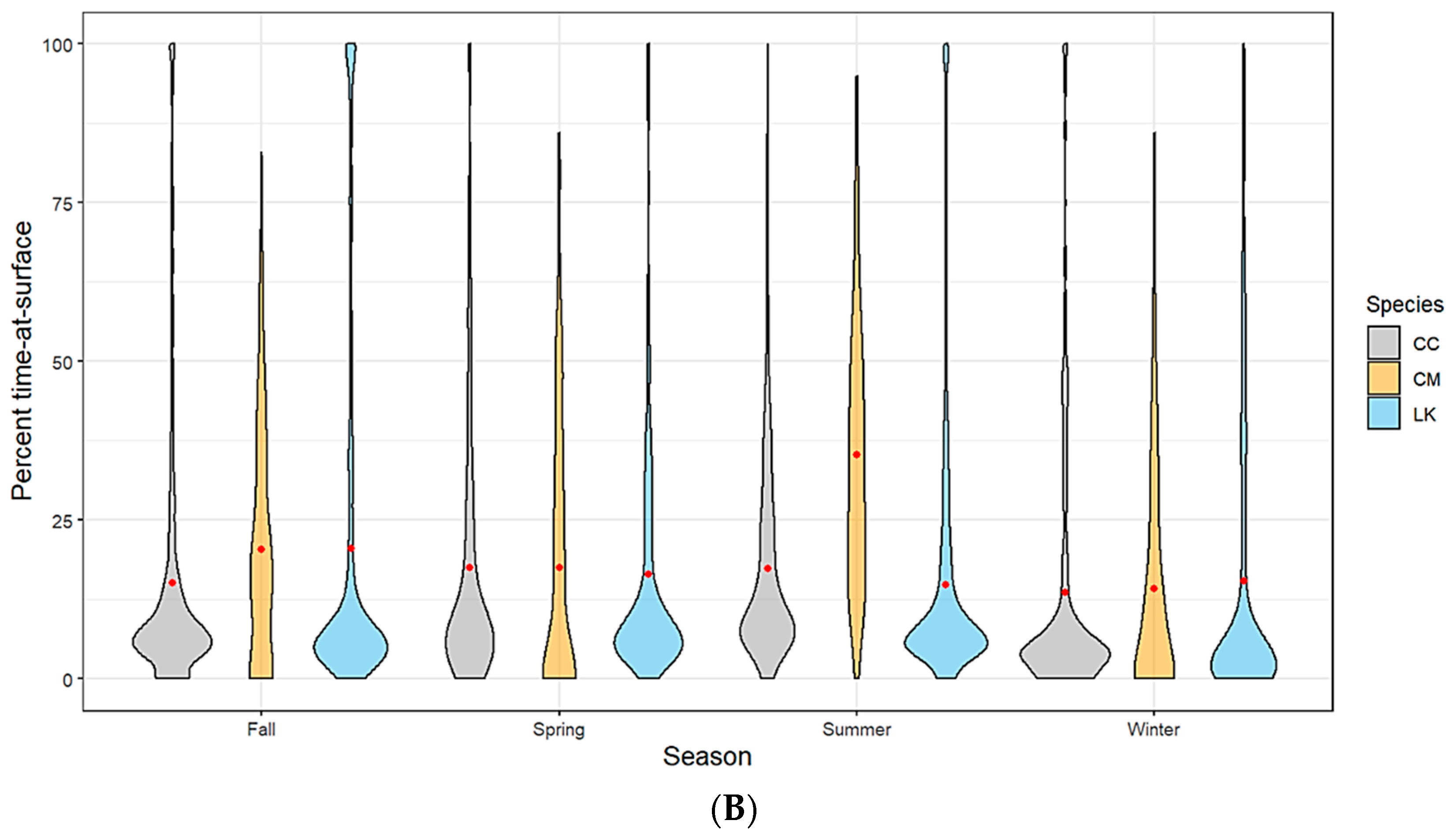
3.2. Time-at-Surface Summaries
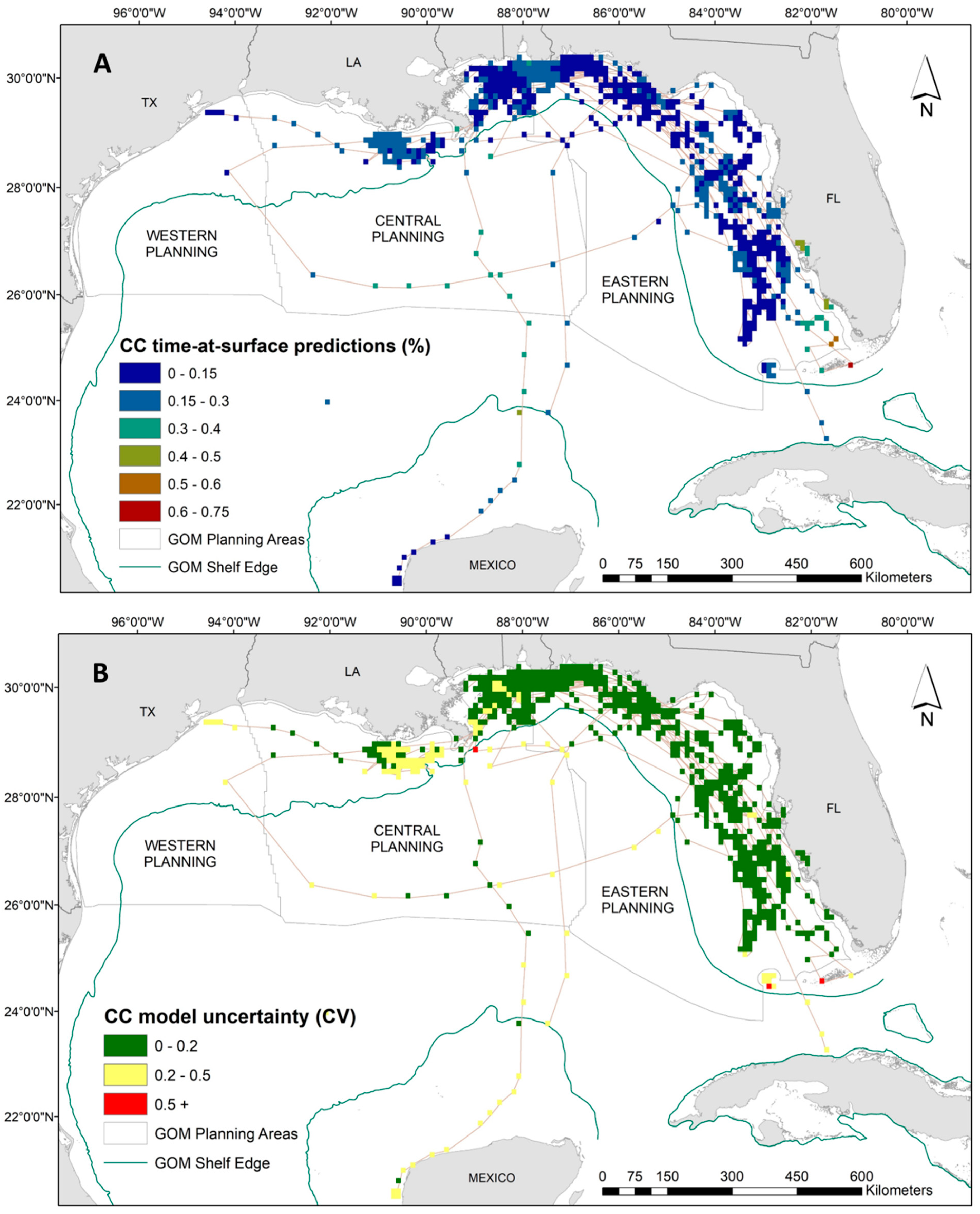
3.3. Modeling Time at Surface
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinsky, M.L.; Selden, R.L.; Kitchel, Z.J. Climate-Driven Shifts in Marine Species Ranges: Scaling from Organisms to Communities. Annu. Rev. Mar. Sci. 2020, 12, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.A.; Alves, J.A.; Gunnarsson, T.G. Mechanisms Driving Phenological and Range Change in Migratory Species. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180047. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, M.P.; Isaac, S.J. The Impacts of Climate Change on Marine Mammals: Early Signs of Significant Problems. Oryx 2007, 41, 19–26. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A.; et al. The Role of Biotic Interactions in Shaping Distributions and Realised Assemblages of Species: Implications for Species Distribution Modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Selig, E.R.; Turner, W.R.; Troëng, S.; Wallace, B.P.; Halpern, B.S.; Kaschner, K.; Lascelles, B.G.; Carpenter, K.E.; Mittermeier, R.A. Global Priorities for Marine Biodiversity Conservation. PLoS ONE 2014, 9, e82898. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.E.; Smith, B.J.; Burkholder, D.; Hart, K.M. Evaluating the Use of Marine Protected Areas by Endangered Species: A Habitat Selection Approach. Ecol. Solut. Evid. 2021, 2, e12035. [Google Scholar] [CrossRef]
- Pérez-Jorge, S.; Tobeña, M.; Prieto, R.; Vandeperre, F.; Calmettes, B.; Lehodey, P.; Silva, M.A. Environmental Drivers of Large-Scale Movements of Baleen Whales in the Mid-North Atlantic Ocean. Divers. Distrib. 2020, 26, 683–698. [Google Scholar] [CrossRef]
- Serratosa, J.; Hyrenbach, K.D.; Miranda-Urbina, D.; Portflitt-Toro, M.; Luna, N.; Luna-Jorquera, G. Environmental Drivers of Seabird At-Sea Distribution in the Eastern South Pacific Ocean: Assemblage Composition Across a Longitudinal Productivity Gradient. Front. Mar. Sci. 2020, 6, 838. [Google Scholar] [CrossRef]
- Hochscheid, S. Why We Mind Sea Turtles’ Underwater Business: A Review on the Study of Diving Behavior. J. Exp. Mar. Biol. Ecol. 2014, 450, 118–136. [Google Scholar] [CrossRef]
- Roncon, G.; Bestley, S.; McMahon, C.R.; Wienecke, B.; Hindell, M.A. View from below: Inferring Behavior and Physiology of Southern Ocean Marine Predators from Dive Telemetry. Front. Mar. Sci. 2018, 5, 464. [Google Scholar] [CrossRef] [Green Version]
- Bauer, R.K.; Fromentin, J.-M.; Demarcq, H.; Bonhommeau, S. Habitat Use, Vertical and Horizontal Behaviour of Atlantic Bluefin Tuna (Thunnus thynnus) in the Northwestern Mediterranean Sea in Relation to Oceanographic Conditions. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 141, 248–261. [Google Scholar] [CrossRef]
- Iverson, A.R.; Fujisaki, I.; Lamont, M.M.; Hart, K.M. Loggerhead Sea Turtle (Caretta caretta) Diving Changes with Productivity, Behavioral Mode, and Sea Surface Temperature. PLoS ONE 2019, 14, e0220372. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Suganuma, H.; Kondo, S.; Sato, K. Long Dive Capacity of Olive Ridley Turtles (Lepidochelys olivacea) at High Water Temperature during the Post-Nesting Foraging Period in the Arafura Sea. J. Exp. Mar. Biol. Ecol. 2022, 546, 151649. [Google Scholar] [CrossRef]
- Freitas, C.; Caldeira, R.; Reis, J.; Dellinger, T. Foraging Behavior of Juvenile Loggerhead Sea Turtles in the Open Ocean: From Lévy Exploration to Area-Restricted Search. Mar. Ecol. Prog. Ser. 2018, 595, 203–215. [Google Scholar] [CrossRef]
- Owen, K.; Jenner, K.C.S.; Jenner, M.N.M.; McCauley, R.D.; Andrews, R.D. Water Temperature Correlates with Baleen Whale Foraging Behaviour at Multiple Scales in the Antarctic. Mar. Freshw. Res. 2019, 70, 19–32. [Google Scholar] [CrossRef]
- Madrak, S.; Lewison, R.; Eguchi, T.; Klimley, A.; Seminoff, J. Effects of Ambient Temperature on Dive Behavior of East Pacific Green Turtles before and after a Power Plant Closure. Mar. Ecol. Prog. Ser. 2022, 683, 157–168. [Google Scholar] [CrossRef]
- McIntyre, T.; Ansorge, I.J.; Bornemann, H.; Plötz, J.; Tosh, C.A.; Bester, M.N. Elephant Seal Dive Behaviour Is Influenced by Ocean Temperature: Implications for Climate Change Impacts on an Ocean Predator. Mar. Ecol. Prog. Ser. 2011, 441, 257–272. [Google Scholar] [CrossRef]
- Arrowsmith, L.; Sequeira, A.; Pattiaratchi, C.; Meekan, M. Water Temperature Is a Key Driver of Horizontal and Vertical Movements of an Ocean Giant, the Whale Shark Rhincodon typus. Mar. Ecol. Prog. Ser. 2021, 679, 101–114. [Google Scholar] [CrossRef]
- Loredo, S.A.; Orben, R.A.; Suryan, R.M.; Lyons, D.E.; Adams, J.; Stephensen, S.W. Spatial and Temporal Diving Behavior of Non-Breeding Common Murres during Two Summers of Contrasting Ocean Conditions. J. Exp. Mar. Biol. Ecol. 2019, 517, 13–24. [Google Scholar] [CrossRef]
- Brodie, S.J.; Thorson, J.T.; Carroll, G.; Hazen, E.L.; Bograd, S.; Haltuch, M.A.; Holsman, K.K.; Kotwicki, S.; Samhouri, J.F.; Willis-Norton, E.; et al. Trade-Offs in Covariate Selection for Species Distribution Models: A Methodological Comparison. Ecography 2020, 43, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Castro, S.; Regos, A.; Gonçalves, J.F.; Alcaraz-Segura, D.; Honrado, J. Remotely Sensed Variables of Ecosystem Functioning Support Robust Predictions of Abundance Patterns for Rare Species. Remote Sens. 2019, 11, 2086. [Google Scholar] [CrossRef]
- Chassignet, E.P.; Hurlburt, H.E.; Smedstad, O.M.; Halliwell, G.R.; Hogan, P.J.; Wallcraft, A.J.; Baraille, R.; Bleck, R. The HYCOM (HYbrid Coordinate Ocean Model) Data Assimilative System. J. Mar. Syst. 2007, 65, 60–83. [Google Scholar] [CrossRef]
- Shepard, A.N.; Valentine, J.F.; D’Elia, C.F.; Yoskowitz, D.W.; Dismukes, D.E. Economic Impact of Gulf of Mexico Ecosystem Goods and Services and Integration into Restoration Decision-Making. Gulf Mex. Sci. 2013, 31, 10–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, C. Ocean Temperature and Color Frontal Zones in the Gulf of Mexico: Where, When, and Why. J. Geophys. Res. Ocean. 2021, 126, e2021JC017544. [Google Scholar] [CrossRef]
- Valverde, R.A.; Holzwart, K.R. Sea Turtles of the Gulf of Mexico BT—Habitats and Biota of the Gulf of Mexico: Before the Deepwater Horizon Oil Spill. In Volume 2: Fish Resources, Fisheries, Sea Turtles, Avian Resources, Marine Mammals, Diseases and Mortalities; Ward, C.H., Ed.; Springer: New York, NY, USA, 2017; pp. 1189–1351. ISBN 978-1-4939-3456-0. [Google Scholar]
- Rabotyagov, S.S.; Kling, C.L.; Gassman, P.W.; Rabalais, N.N.; Turner, R.E. The Economics of Dead Zones: Causes, Impacts, Policy Challenges, and a Model of the Gulf of Mexico Hypoxic Zone. Rev. Environ. Econ. Policy 2014, 8, 58–79. [Google Scholar] [CrossRef]
- Wilson, M.; Tucker, A.D.; Beedholm, K.; Mann, D.A. Changes of Loggerhead Turtle (Caretta caretta) Dive Behavior Associated with Tropical Storm Passage during the Inter-Nesting Period. J. Exp. Biol. 2017, 220, 3432–3441. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.; Bailey, D.; Maung-douglass, E.; Partyka, M.; Sempier, S.; Skelton, T. Advancements in Understanding Ocean Circulation and Tracking the Movements of Oil; GOMSG-G-21-001; National Oceanic and Athmospheric Administration: Silver Spring, MA, USA, 2021. [Google Scholar]
- Maxwell, S.M.; Hazen, E.L.; Lewison, R.L.; Dunn, D.C.; Bailey, H.; Bograd, S.J.; Briscoe, D.K.; Fossette, S.; Hobday, A.J.; Bennett, M.; et al. Dynamic Ocean Management: Defining and Conceptualizing Real-Time Management of the Ocean. Mar. Policy 2015, 58, 42–50. [Google Scholar] [CrossRef]
- Hart, K.M.; Lamont, M.M.; Iverson, A.R.; Smith, B.J. The Importance of the Northeastern Gulf of Mexico to Foraging Loggerhead Sea Turtles. Front. Mar. Sci. 2020, 7, 330. [Google Scholar] [CrossRef]
- Crowe, L.M.; Hatch, J.M.; Patel, S.H.; Smolowitz, R.J.; Haas, H.L. Riders on the Storm: Loggerhead Sea Turtles Detect and Respond to a Major Hurricane in the Northwest Atlantic Ocean. Mov. Ecol. 2020, 8, 32. [Google Scholar] [CrossRef]
- Wildermann, N.E.; Sasso, C.R.; Stokes, L.W.; Snodgrass, D.; Fuentes, M.M.P.B. Habitat Use and Behavior of Multiple Species of Marine Turtles at a Foraging Area in the Northeastern Gulf of Mexico. Front. Mar. Sci. 2019, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Hatch, J.M.; Haas, H.L.; Sasso, C.R.; Patel, S.H.; Smolowitz, R.J. Estimating the Complex Patterns of Survey Availability for Loggerhead Turtles. J. Wildl. Manag. 2022, 86, e22208. [Google Scholar] [CrossRef]
- Polovina, J.J.; Balazs, G.H.; Howell, E.A.; Parker, D.M.; Seki, M.P.; Dutton, P.H. Forage and Migration Habitat of Loggerhead (Caretta caretta) and Olive Ridley (Lepidochelys olivacea) Sea Turtles in the Central North Pacific Ocean. Fish. Oceanogr. 2004, 13, 36–51. [Google Scholar] [CrossRef]
- Shillinger, G.L.; Swithenbank, A.M.; Bailey, H.; Bograd, S.J.; Castelton, M.R.; Wallace, B.P.; Spotila, J.R.; Paladino, F.V.; Piedra, R.; Block, B.A. Vertical and Horizontal Habitat Preferences of Post-Nesting Leatherback Turtles in the South Pacific Ocean. Mar. Ecol. Prog. Ser. 2011, 422, 275–289. [Google Scholar] [CrossRef]
- Scales, K.L.; Miller, P.I.; Varo-Cruz, N.; Hodgson, D.J.; Hawkes, L.A.; Godley, B.J. Oceanic Loggerhead Turtles Caretta caretta Associate with Thermal Fronts: Evidence from the Canary Current Large Marine Ecosystem. Mar. Ecol. Prog. Ser. 2015, 519, 195–207. [Google Scholar] [CrossRef]
- Witherington, B.; Hirama, S.; Hardy, R. Young Sea Turtles of the Pelagic Sargassum-Dominated Drift Community: Habitat Use, Population Density, and Threats. Mar. Ecol. Prog. Ser. 2012, 463, 1–22. [Google Scholar] [CrossRef]
- Hart, K.M.; Guzy, J.C.; Smith, B.J. Drivers of Realized Satellite Tracking Duration in Marine Turtles. Mov. Ecol. 2021, 9, 1–14. [Google Scholar] [CrossRef]
- Breed, G.A.; Don Bowen, W.; Leonard, M.L. Behavioral Signature of Intraspecific Competition and Density Dependence in Colony-breeding Marine Predators. Ecol. Evol. 2013, 3, 3838–3854. [Google Scholar] [CrossRef]
- Jonsen, I. Joint Estimation over Multiple Individuals Improves Behavioural State Inference from Animal Movement Data. Sci. Rep. 2016, 6, 20625. [Google Scholar] [CrossRef]
- Hart, K.M.; Lamont, M.M.; Fujisaki, I.; Tucker, A.D.; Carthy, R.R. Common Coastal Foraging Areas for Loggerheads in the Gulf of Mexico: Opportunities for Marine Conservation. Biol. Conserv. 2012, 145, 185–194. [Google Scholar] [CrossRef]
- Jonsen, I.D.; Flemming, J.M.; Myers, R.A. Robust State–Space Modeling of Animal Movement Data. Ecology 2005, 86, 2874–2880. [Google Scholar] [CrossRef]
- Jonsen, I.D.; Basson, M.; Bestley, S.; Bravington, M.V.; Patterson, T.A.; Pedersen, M.W.; Thomson, R.; Thygesen, U.H.; Wotherspoon, S.J. State-Space Models for Bio-Loggers: A Methodological Road Map. Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 88, 34–46. [Google Scholar] [CrossRef]
- Belkin, I.M.; O’Reilly, J.E. An Algorithm for Oceanic Front Detection in Chlorophyll and SST Satellite Imagery. J. Mar. Syst. 2009, 78, 319–326. [Google Scholar] [CrossRef]
- Hu, C. An Empirical Approach to Derive MODIS Ocean Color Patterns under Severe Sun Glint. Geophys. Res. Lett. 2011, 38, L01603. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017; ISBN 9781315370279. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Hart, K.M.; Roberts, K.E.; Lamont, M.; Garrison, L.P.; Sasso, C.R. Surface Time for Sea Turtles in the Gulf of Mexico, 2010–2019; U.S. Geological Survey Data Release, 2022. [Google Scholar] [CrossRef]
- Fuentes, M.; Bell, I.; Hagihara, R.; Hamann, M.; Hazel, J.; Huth, A.; Seminoff, J.A.; Sobtzick, S.; Marsh, H. Improving In-Water Estimates of Marine Turtle Abundance by Adjusting Aerial Survey Counts for Perception and Availability Biases. J. Exp. Mar. Biol. Ecol. 2015, 471, 77–83. [Google Scholar] [CrossRef]
- Belkin, I.M. Review Remote Sensing of Ocean Fronts in Marine Ecology and Fisheries. Remote Sens. 2021, 13, 883. [Google Scholar] [CrossRef]
- Gaspar, P.; Georges, J.Y.; Fossette, S.; Lenoble, A.; Ferraroli, S.; Le Maho, Y. Marine Animal Behaviour: Neglecting Ocean Currents Can Lead Us up the Wrong Track. Proc. R. Soc. B Biol. Sci. 2006, 273, 2697–2702. [Google Scholar] [CrossRef]
- Sequeira, A.M.M.; Rodríguez, J.P.; Eguíluz, V.M.; Harcourt, R.; Hindell, M.; Sims, D.W.; Duarte, C.M.; Costa, D.P.; Fernández-Gracia, J.; Ferreira, L.C.; et al. Convergence of Marine Megafauna Movement Patterns in Coastal and Open Oceans. Proc. Natl. Acad. Sci. USA 2018, 115, 3072–3077. [Google Scholar] [CrossRef]
- Della Penna, A.; De Monte, S.; Kestenare, E.; Guinet, C.; D’Ovidio, F. Quasi-Planktonic Behavior of Foraging Top Marine Predators. Sci. Rep. 2015, 5, 18063. [Google Scholar] [CrossRef] [Green Version]
- Cox, S.L.; Embling, C.B.; Hosegood, P.J.; Votier, S.C.; Ingram, S.N. Oceanographic Drivers of Marine Mammal and Seabird Habitat-Use across Shelf-Seas: A Guide to Key Features and Recommendations for Future Research and Conservation Management. Estuar. Coast. Shelf Sci. 2018, 212, 294–310. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Zárate, P.; Coyne, M.; Foley, D.G.; Parker, D.; Lyon, B.N.; Dutton, P.H. Post-Nesting Migrations of Galápagos Green Turtles Chelonia Mydas in Relation to Oceanographic Conditions: Integrating Satellite Telemetry with Remotely Sensed Ocean Data. Endanger. Species Res. 2008, 4, 57–72. [Google Scholar] [CrossRef]
- Trudelle, L.; Cerchio, S.; Zerbini, A.N.; Geyer, Y.; Mayer, F.X.; Jung, J.L.; Hervé, M.R.; Pous, S.; Pous, S.; Rosenbaum, H.C.; et al. Influence of Environmental Parameters on Movements and Habitat Utilization of Humpback Whales (Megaptera novaeangliae) in the Madagascar Breeding Ground. R. Soc. Open Sci. 2016, 3, 160616. [Google Scholar] [CrossRef] [PubMed]
- Lambardi, P.; Lutjeharms, J.R.E.; Mencacci, R.; Hays, G.C.; Luschi, P. Influence of Ocean Currents on Long-Distance Movement of Leatherback Sea Turtles in the Southwest Indian Ocean. Mar. Ecol. Prog. Ser. 2008, 353, 289–301. [Google Scholar] [CrossRef]
- Papastamatiou, Y.P.; Iosilevskii, G.; Di Santo, V.; Huveneers, C.; Hattab, T.; Planes, S.; Ballesta, L.; Mourier, J. Sharks Surf the Slope: Current Updrafts Reduce Energy Expenditure for Aggregating Marine Predators. J. Anim. Ecol. 2021, 90, 2302–2314. [Google Scholar] [CrossRef]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will Remote Sensing Shape the next Generation of Species Distribution Models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef]
- Fuentes, M.M.P.B.; Meletis, Z.A.; Wildermann, N.E.; Ware, M. Conservation Interventions to Reduce Vessel Strikes on Sea Turtles: A Case Study in Florida. Mar. Policy 2021, 128, 104471. [Google Scholar] [CrossRef]
- Wallace, B.P.; Kot, C.Y.; Dimatteo, A.D.; Lee, T.; Crowder, L.B.; Lewison, R.L. Impacts of Fisheries Bycatch on Marine Turtle Populations Worldwide: Toward Conservation and Research Priorities. Ecosphere 2013, 4, 1–49. [Google Scholar] [CrossRef]
- Jonsen, I.D.; McMahon, C.R.; Patterson, T.A.; Auger-Méthé, M.; Harcourt, R.; Hindell, M.A.; Bestley, S. Movement Responses to Environment: Fast Inference of Variation among Southern Elephant Seals with a Mixed Effects Model. Ecology 2019, 100, e02566. [Google Scholar] [CrossRef]
- Conners, M.G.; Michelot, T.; Heywood, E.I.; Orben, R.A.; Phillips, R.A.; Vyssotski, A.L.; Shaffer, S.A.; Thorne, L.H. Hidden Markov Models Identify Major Movement Modes in Accelerometer and Magnetometer Data from Four Albatross Species. Mov. Ecol. 2021, 9, 7. [Google Scholar] [CrossRef]
- Hawkes, L.A.; Broderick, A.C.; Godfrey, M.H.; Godley, B.J. Climate Change and Marine Turtles. Endanger. Species Res. 2009, 7, 137–154. [Google Scholar] [CrossRef]
- Roberts, J.J.; Best, B.D.; Mannocci, L.; Fujioka, E.; Halpin, P.N.; Palka, D.L.; Garrison, L.P.; Mullin, K.D.; Cole, T.V.N.; Khan, C.B.; et al. Habitat-Based Cetacean Density Models for the U.S. Atlantic and Gulf of Mexico. Sci. Rep. 2016, 6, 22615. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.A.; Forney, K.A.; Foley, D.G.; Smith, R.C.; Moore, T.J.; Barlow, J. Predicting Seasonal Density Patterns of California Cetaceans Based on Habitat Models. Endanger. Species Res. 2014, 23, 1–22. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Eguchi, T.; Carretta, J.; Allen, C.D.; Prosperi, D.; Rangel, R.; Gilpatrick, J.W.; Forney, K.; Peckham, S.H. Loggerhead Sea Turtle Abundance at a Foraging Hotspot in the Eastern Pacific Ocean: Implications for at-Sea Conservation. Endanger. Species Res. 2014, 24, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Rappucci, G.; Barry, K.; Foster, M.; Garrison, L.P.; Litz, J. GoMMAPPS Fall Aerial Abundance Survey during October–November 2018. In GOMMAPPS Summary Report; Bureau of Ocean Energy Management: Washington, DC, USA, 2018. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Source | Time Window | Spatial Resolution |
|---|---|---|---|---|
| Sea surface temperature | °C | ERDDAP OISST-V2-AVHRR | Daily | 3.5 km (0.04°) |
| Sea surface temperature anomaly | °C | ERDDAP OISST-V2-AVHRR | Daily | 3.5 km (0.04°) |
| Sea surface salinity | PSU | HYCOM + NCODA | Daily | 3.5 km (0.04°) |
| Sea surface height | m | HYCOM + NCODA | Daily | 3.5 km (0.04°) |
| Bottom depth | m | SRTM30 | Static | ~1 km (30’) |
| Distance to shore | km | Derived from Harris et al. (2014) | Static | NA |
| Distance to shelf | km | Derived from Harris et al. (2014) | Static | NA |
| Current strength | m/s | HYCOM + NCODA | Daily | 3.5 km (0.04°) |
| Current direction | Degrees | HYCOM + NCODA | Daily | 3.5 km (0.04°) |
| Frontal gradient magnitude-color fronts | 1/km | USF Optical Oceanography Laboratory; BOA front detection algorithm (Belkin and O’Reilly, 2009) based on MODIS/Aqua 9-km CI | Weekly | ~9 km |
| Frontal gradient magnitude-thermal fronts | °C/km | USF Optical Oceanography Laboratory; BOA front detection algorithm (Belkin and O’Reilly, 2009) based on MODIS/Aqua 9-km SST | Weekly | ~9 km |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, K.E.; Garrison, L.P.; Ortega-Ortiz, J.; Hu, C.; Zhang, Y.; Sasso, C.R.; Lamont, M.; Hart, K.M. The Influence of Satellite-Derived Environmental and Oceanographic Parameters on Marine Turtle Time at Surface in the Gulf of Mexico. Remote Sens. 2022, 14, 4534. https://doi.org/10.3390/rs14184534
Roberts KE, Garrison LP, Ortega-Ortiz J, Hu C, Zhang Y, Sasso CR, Lamont M, Hart KM. The Influence of Satellite-Derived Environmental and Oceanographic Parameters on Marine Turtle Time at Surface in the Gulf of Mexico. Remote Sensing. 2022; 14(18):4534. https://doi.org/10.3390/rs14184534
Chicago/Turabian StyleRoberts, Kelsey E., Lance P. Garrison, Joel Ortega-Ortiz, Chuanmin Hu, Yingjun Zhang, Christopher R. Sasso, Margaret Lamont, and Kristen M. Hart. 2022. "The Influence of Satellite-Derived Environmental and Oceanographic Parameters on Marine Turtle Time at Surface in the Gulf of Mexico" Remote Sensing 14, no. 18: 4534. https://doi.org/10.3390/rs14184534
APA StyleRoberts, K. E., Garrison, L. P., Ortega-Ortiz, J., Hu, C., Zhang, Y., Sasso, C. R., Lamont, M., & Hart, K. M. (2022). The Influence of Satellite-Derived Environmental and Oceanographic Parameters on Marine Turtle Time at Surface in the Gulf of Mexico. Remote Sensing, 14(18), 4534. https://doi.org/10.3390/rs14184534