
Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
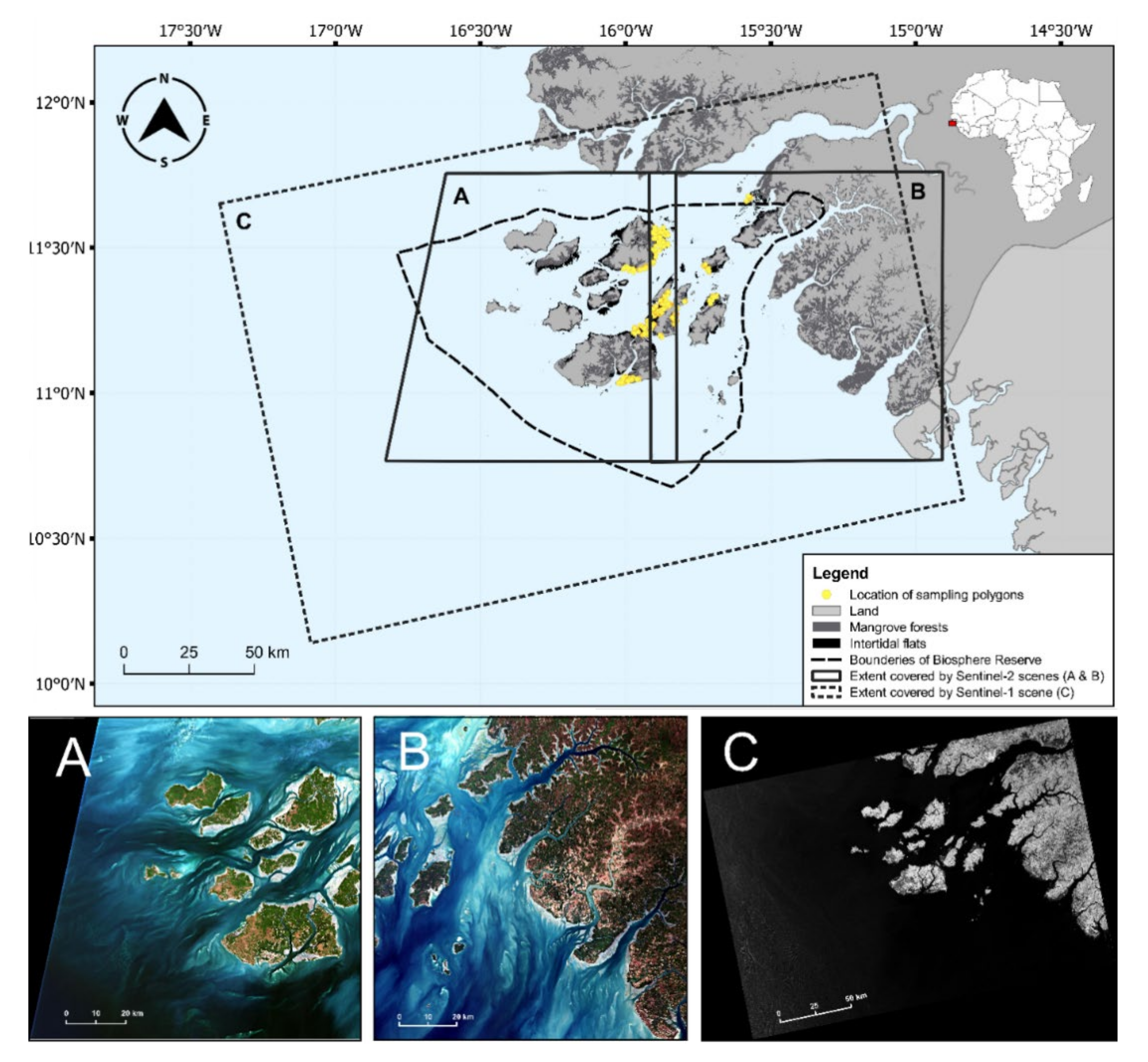
2.1. Study Area
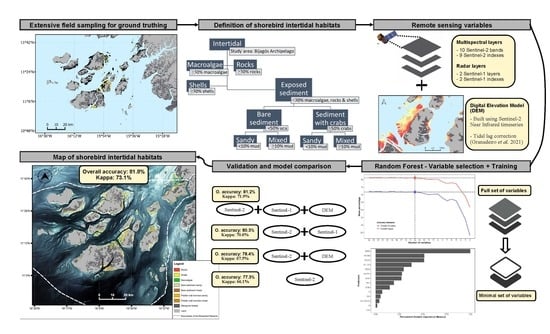
2.2. Overall Workflow
2.3. Field Reference Data
2.3.1. Field Sampling
2.3.2. Grain Size Analysis
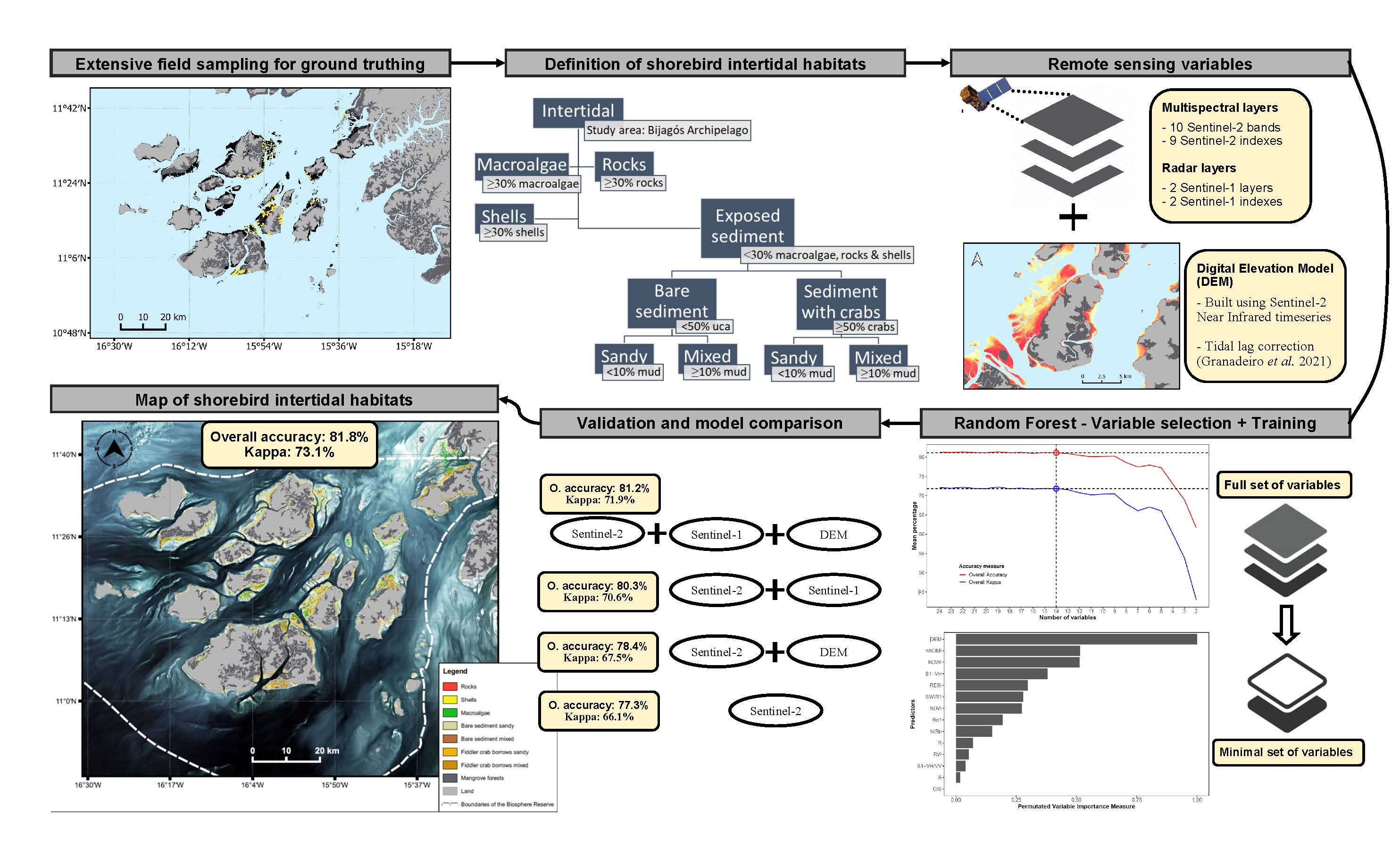
2.4. Definition of Shorebird Intertidal Habitats
2.5. Satellite Imagery Selection and Processing
2.5.1. Selection of Satellite Images
2.5.2. Sentinel-1 and Sentinel-2 Image Pre-Processing Methods
2.5.3. Intercalibration of Sentinel-2 Scenes
2.5.4. Extraction of the Intertidal Area Water and Land Mask
2.5.5. Digital Elevation Model and Final Study Area Extent
2.6. Predictors for Supervised Classification
2.7. Supervised Classification with Random Forest Algorithm
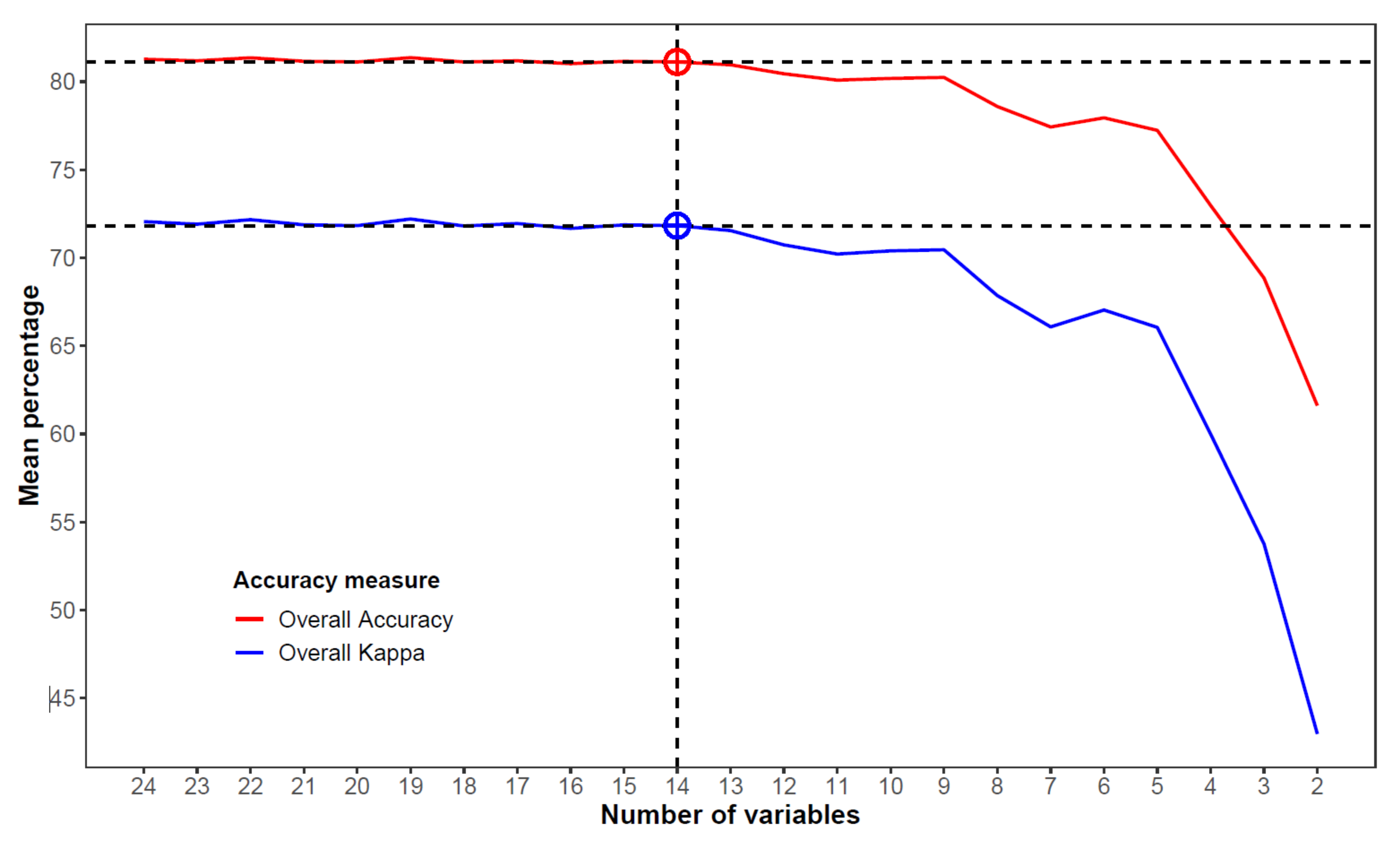
2.8. Variable Selection
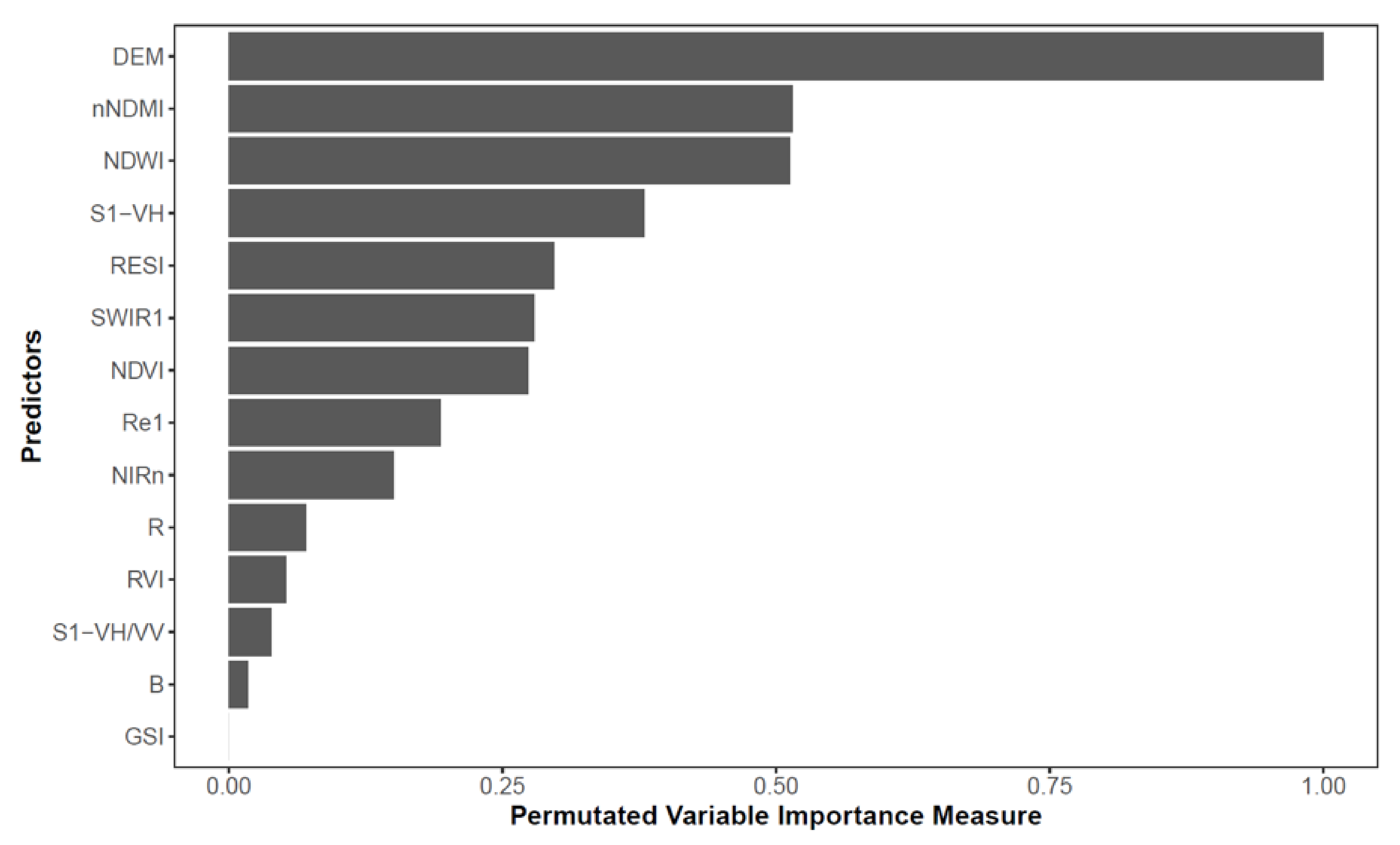
2.9. Variable Importance Metrics
2.10. Map of Intertidal Habitat Class Predictions
2.11. Post-Classification Filtering
2.12. Validation and Assessment of Model Approach
3. Results
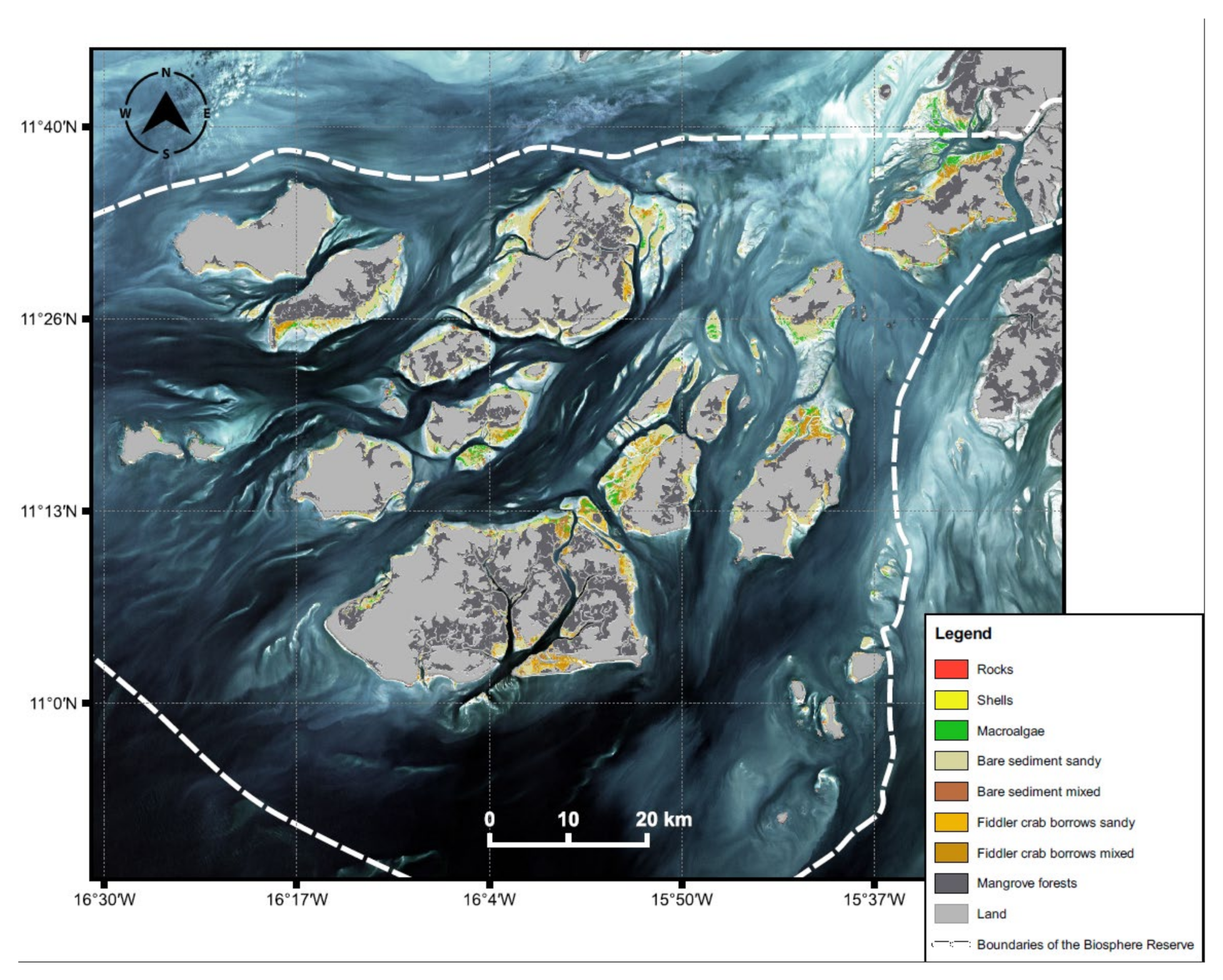
3.1. Extent of Intertidal Area of the Bijagós Archipelago
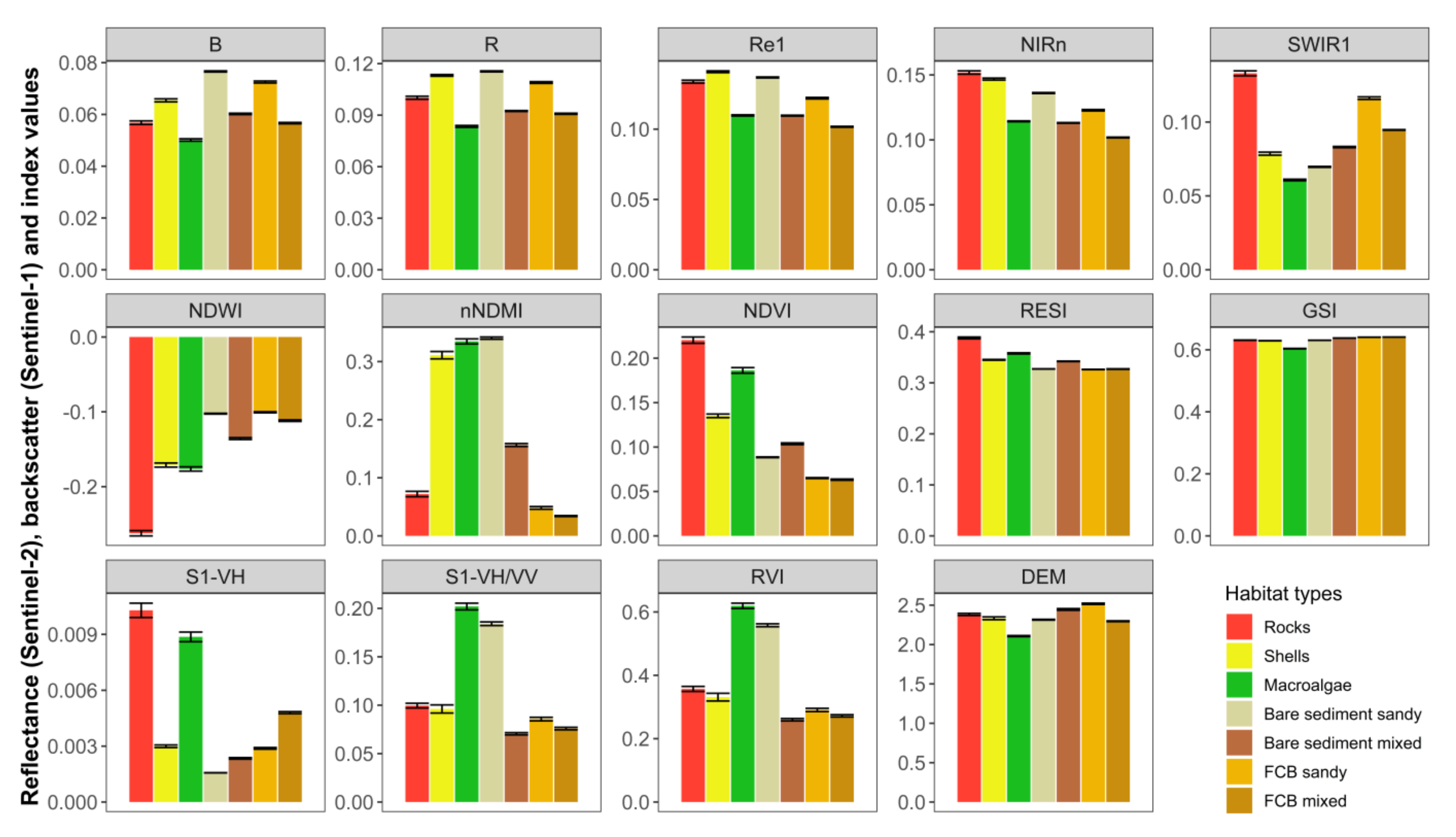
3.2. Field Reference Data and Definition of Shorebird Intertidal Habitats
3.3. Variable Selection and Final Random Forest Model
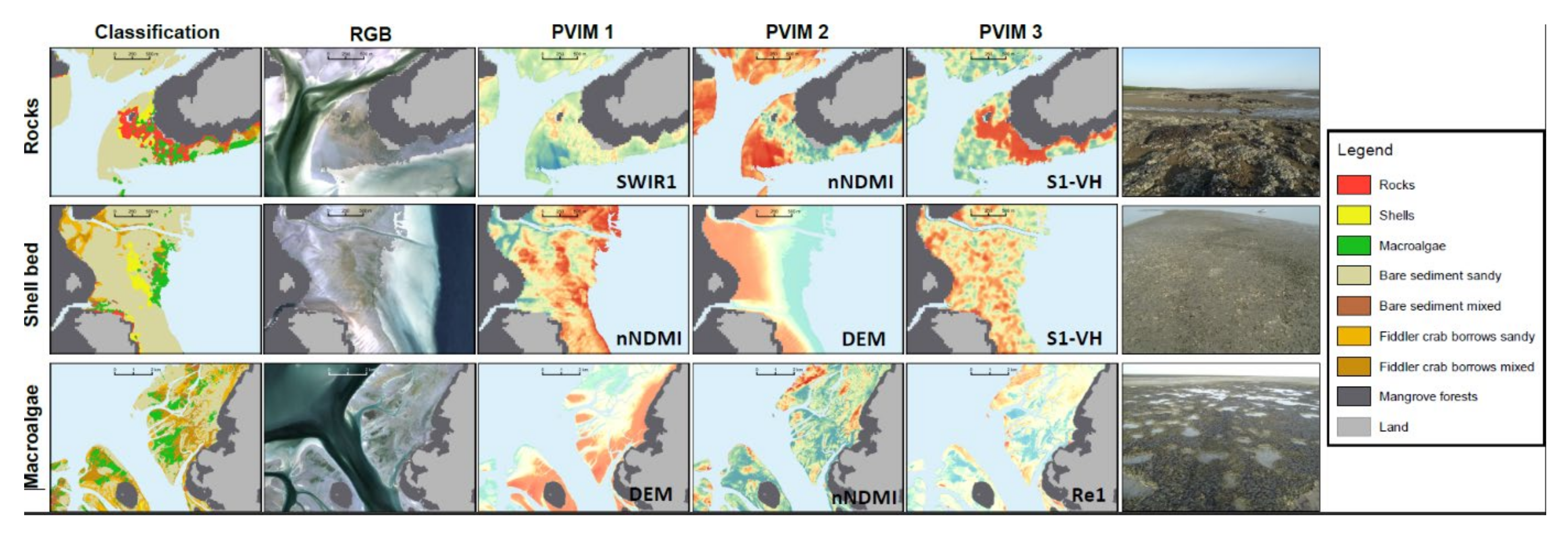
3.4. Variable Importance
3.5. Post-Classification Filtering
3.6. Analysis of the Final Intertidal Habitat Prediction Map
3.7. Comparison between Final Model and Models without Sentinel-1 and DEM
4. Discussion
4.1. Classification Performance
4.2. Multi-Sensor Approach and Variable Importance
4.3. Extent and Distribution of Intertidal Habitats
4.4. Implications for Shorebirds and Benthic Invertebrates
4.5. Applications to Monitoring and Management in the Bijagós Archipelago
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, N.J.; Phinn, S.R.; DeWitt, M.; Ferrari, R.; Johnston, R.; Lyons, M.B.; Clinton, N.; Thau, D.; Fuller, R.A. The global distribution and trajectory of tidal flats. Nature 2019, 565, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Beninger, P.G. Mudflat Ecology; Beninger, P.G., Ed.; Aquatic, Ec.; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-99192-4. [Google Scholar]
- van der Zee, E.M.; Angelini, C.; Govers, L.L.; Christianen, M.J.A.; Altieri, A.H.; van der Reijden, K.J.; Silliman, B.R.; van de Koppel, J.; van der Geest, M.; van Gils, J.A.; et al. How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152326. [Google Scholar] [CrossRef] [PubMed]
- van Gils, J.A.; van der Geest, M.; Jansen, E.J.; Govers, L.L.; de Fouw, J.; Piersma, T. Trophic cascade induced by molluscivore predator alters pore-water biogeochemistry via competitive release of prey. Ecology 2012, 93, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Booty, J.M.; Underwood, G.J.C.; Parris, A.; Davies, R.G.; Tolhurst, T.J. Shorebirds Affect Ecosystem Functioning on an Intertidal Mudflat. Front. Mar. Sci. 2020, 7, 685. [Google Scholar] [CrossRef]
- van de Kam, J.; Ens, B.; Piersma, T.; Zwarts, L. Shorebirds: An Illustrated Behavioural Ecology; KNNV Publishers: Utrecht, The Netherlands, 2004. [Google Scholar]
- Lourenço, P.M.; Catry, T.; Granadeiro, J.P. Diet and feeding ecology of the wintering shorebird assemblage in the Bijagós archipelago, Guinea-Bissau. J. Sea Res. 2017, 128, 52–60. [Google Scholar] [CrossRef]
- Stroud, D.A.; Baker, A.; Blanco, D.E.; Davidson, N.C.; Delany, S.; Ganter, B.; Gill, R.; González, P.; Haanstra, L.; Morrison, R.I.G.; et al. The conservation and population status of the world’s waders at the turn of the millennium In Waterbirds around the World; Boere, G.C., Galbraith, C.A., Stroud, D.A., Eds.; The Stationery Office: London, UK, 2006; pp. 643–648. [Google Scholar]
- Sutherland, W.J.; Alves, J.A.; Amano, T.; Chang, C.H.; Davidson, N.C.; Max Finlayson, C.; Gill, J.A.; Gill, R.E., Jr.; González, P.M.; Gunnarsson, T.G.; et al. A horizon scanning assessment of current and potential future threats to migratory shorebirds. Ibis 2012, 154, 663–679. [Google Scholar] [CrossRef]
- East Atlantic Flyway Assessment 2017: The Status of Coastal Waterbird Populations and Their Sites; Van Roomen, M., Nagy, S., Citegetse, G., Schekkerman, H., Eds.; Wadden Sea Flyway Initiative; Wetlands International; Wadden Sea Flyway Initiative: Wilhelmshaven, Germany; Wetlands International: Wageningen, The Netherlands; BirdLife International: Cambridge, UK, 2018. [Google Scholar]
- Oudman, T.; Schekkerman, H.; Kidee, A.; Van Roomen, M.; Camara, M.; Smit, C.; Horn, J.T.; Piersma, T.; El-Hacen, E.H.M. Changes in the waterbird community of the Parc National du Banc d’Arguin, Mauritania, 1980–2017. Bird Conserv. Int. 2020, 30, 618–633. [Google Scholar] [CrossRef]
- van Gils, J.A.; Lisovski, S.; Lok, T.; Meissner, W.; Ozarowska, A.; de Fouw, J.; Rakhimberdiev, E.; Soloviev, M.Y.; Piersma, T.; Klaassen, M. Body shrinkage due to Arctic warming reduces red knot fitness in tropical wintering range. Science 2016, 352, 819–821. [Google Scholar] [CrossRef]
- Rakhimberdiev, E.; Duijns, S.; Karagicheva, J.; Camphuysen, C.J.; Dekinga, A.; Dekker, R.; Gavrilov, A.; ten Horn, J.; Jukema, J.; Saveliev, A.; et al. Fuelling conditions at staging sites can mitigate Arctic warming effects in a migratory bird. Nat. Commun. 2018, 9, 4263. [Google Scholar] [CrossRef]
- Salvig, J.C.; Asbirk, S.; Kjeldsen, J.P.; Rasmussen, P.A.F. Wintering waders in the Bijagos Archipelago, Guinee-Bissau 1992–1993. Ardea 1994, 82, 137–142. [Google Scholar]
- Zwarts, L. Numbers and distribution of coastal waders in guinea-bissau. Ardea 1988, 76, 42–55. [Google Scholar]
- Henriques, M.; Belo, J.R.; Sá, J.; Monteiro, H.; Alves, J.A.; Piersma, T.; Dodman, T.; van Roomen, M. The Bijagós Archipelago: A key area for waterbirds of the East Atlantic Flyway. In East Atlantic Flyway Assessment 2020: The Status of Coastal Waterbird Populations and Their Sites; van Roomen, M., Citegetse, G., Crowe, O., Dodman, T., Hagemeijer, W., Meise, K., Schekkerman, H., Eds.; Wadden Sea Flyway Initiative: Wilhelmshaven, Germany; Wetlands International: Wageningen, The Netherlands; BirdLife International: Cambridge, UK, 2022; pp. 81–93. [Google Scholar]
- Piersma, T. What is Habitat Quality? Dissecting a Research Portfolio on Shorebirds. In Birds and Habitat: Relationships in Changing Landscapes; Fuller, R., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 383–407. [Google Scholar]
- Lourenço, P.M.; Granadeiro, J.P.; Catry, T. Low macroinvertebrate biomass suggests limited food availability for shorebird communities in intertidal areas of the Bijagós archipelago (Guinea-Bissau). Hydrobiologia 2018, 816, 197–212. [Google Scholar] [CrossRef]
- Compton, T.J.; Holthuijsen, S.; Koolhaas, A.; Dekinga, A.; ten Horn, J.; Smith, J.; Galama, Y.; Brugge, M.; van der Wal, D.; van der Meer, J.; et al. Distinctly variable mudscapes: Distribution gradients of intertidal macrofauna across the Dutch Wadden Sea. J. Sea Res. 2013, 82, 103–116. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Meireles, S.; Pereira, T.; Gama, A.; Quintino, V. Spatial Patterns of Benthic Macroinvertebrates in Intertidal Areas of a Southern European Estuary: The Tagus, Portugal. Hydrobiologia 2006, 555, 99–113. [Google Scholar] [CrossRef]
- Passarelli, C.; Olivier, F.; Paterson, D.M.; Meziane, T.; Hubas, C. Organisms as cooperative ecosystem engineers in intertidal flats. J. Sea Res. 2014, 92, 92–101. [Google Scholar] [CrossRef]
- Umanzor, S.; Ladah, L.; Calderon-Aguilera, L.; Zertuche-González, J. Intertidal macroalgae influence macroinvertebrate distribution across stress scenarios. Mar. Ecol. Prog. Ser. 2017, 584, 67–77. [Google Scholar] [CrossRef]
- Chen, T.Y.; Hwang, G.W.; Mayfield, A.B.; Chen, C.P.; Lin, H.J. The relationship between intertidal soil composition and fiddler crab burrow depth. Ecol. Eng. 2017, 100, 256–260. [Google Scholar] [CrossRef]
- El-Hacen, E.H.M.; Bouma, T.J.; Oomen, P.; Piersma, T.; Olff, H. Large-scale ecosystem engineering by flamingos and fiddler crabs on West-African intertidal flats promote joint food availability. Oikos 2019, 128, 753–764. [Google Scholar] [CrossRef]
- Paulino, J.; Granadeiro, J.P.; Henriques, M.; Belo, J.; Catry, T. Composition and abundance of shorebird and macroinvertebrate communities differ according to densities of burrowing fiddler crabs in tropical intertidal flats. Hydrobiologia 2021, 848, 3905–3919. [Google Scholar] [CrossRef]
- Mathot, K.J.; Piersma, T.; Elner, R.W. Shorebirds as Integrators and Indicators of Mudflat Ecology. In Mudflat Ecology; Springer: Cham, Switzerland, 2018; pp. 309–338. [Google Scholar] [CrossRef]
- Zhang, C.; Gong, Z.; Qiu, H.; Zhang, Y.; Zhou, D. Mapping typical salt-marsh species in the Yellow River Delta wetland supported by temporal-spatial-spectral multidimensional features. Sci. Total Environ. 2021, 783, 147061. [Google Scholar] [CrossRef]
- Zoffoli, M.L.; Gernez, P.; Rosa, P.; Le Bris, A.; Brando, V.E.; Barillé, A.L.; Harin, N.; Peters, S.; Poser, K.; Spaias, L.; et al. Sentinel-2 remote sensing of Zostera noltei-dominated intertidal seagrass meadows. Remote Sens. Environ. 2020, 251, 112020. [Google Scholar] [CrossRef]
- Pottier, A.; Catry, T.; Trégarot, E.; Maréchal, J.-P.; Fayad, V.; David, G.; Sidi Cheikh, M.; Failler, P. Mapping coastal marine ecosystems of the National Park of Banc d’Arguin (PNBA) in Mauritania using Sentinel-2 imagery. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102419. [Google Scholar] [CrossRef]
- Gade, M.; Melchionna, S.; Stelzer, K.; Kohlus, J. Multi-frequency SAR data help improving the monitoring of intertidal flats on the German North Sea coast. Estuar. Coast. Shelf Sci. 2014, 140, 32–42. [Google Scholar] [CrossRef]
- Choi, J.K.; Ryu, J.H.; Lee, Y.K.; Yoo, H.R.; Woo, H.J.; Kim, C.H. Quantitative estimation of intertidal sediment characteristics using remote sensing and GIS. Estuar. Coast. Shelf Sci. 2010, 88, 125–134. [Google Scholar] [CrossRef]
- Wang, W.; Yang, X.; Liu, G.; Zhou, H.; Ma, W.; Yu, Y.; Li, Z. Random forest classification of sediments on exposed intertidal flats using ALOS-2 quad-polarimetric SAR data. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, 41, 1191–1194. [Google Scholar] [CrossRef]
- Choi, J.K.; Oh, H.J.; Koo, B.J.; Lee, S.; Ryu, J.H. Macrobenthos habitat mapping in a tidal flat using remotely sensed data and a GIS-based probabilistic model. Mar. Pollut. Bull. 2011, 62, 564–572. [Google Scholar] [CrossRef]
- Müller, G.; Stelzer, K.; Smollich, S.; Gade, M.; Adolph, W.; Melchionna, S.; Kemme, L.; Geißler, J.; Millat, G.; Reimers, H.C.; et al. Remotely sensing the German Wadden Sea—A new approach to address national and international environmental legislation. Environ. Monit. Assess. 2016, 188, 595. [Google Scholar] [CrossRef]
- Wade, S.; Hickey, R. Mapping migratory wading bird feeding habitats using satellite imagery and field data, Eighty-Mile Beach, Western Australia. J. Coast. Res. 2008, 24, 759–770. [Google Scholar] [CrossRef][Green Version]
- Okoye, O.K.; Li, H.; Gong, Z. Retraction of invasive Spartina alterniflora and its effect on the habitat loss of endangered migratory bird species and their decline in YNNR using remote sensing technology. Ecol. Evol. 2020, 10, 13810–13824. [Google Scholar] [CrossRef]
- Jackson, M.V.; Fuller, R.A.; Gan, X.; Li, J.; Mao, D.; Melville, D.S.; Murray, N.J.; Wang, Z.; Choi, C.Y. Dual threat of tidal flat loss and invasive Spartina alterniflora endanger important shorebird habitat in coastal mainland China. J. Environ. Manage. 2021, 278, 111549. [Google Scholar] [CrossRef]
- Danufsky, T.; Colwell, M.A. Winter shorebird communities and tidal flat characteristics at Humboldt Bay, California. Condor 2003, 105, 117–129. [Google Scholar] [CrossRef]
- Morrison, R.I.G. The Use of Remote Sensing to Evaluate Shorebird Habitats and Populations on Prince Charles Island, Foxe Basin, Canada. Arctic 1997, 50, 55–75. [Google Scholar] [CrossRef]
- Zhang, T.; Tian, B.; Bo, S.; Yuan, L.; Zhao, Y.; Yuan, X. Mapping the conservation priority of migratory shorebird habitat on a dynamic deltaic coast. Estuar. Coast. Shelf Sci. 2018, 212, 219–232. [Google Scholar] [CrossRef]
- Schulte to Bühne, H.; Pettorelli, N. Better together: Integrating and fusing multispectral and radar satellite imagery to inform biodiversity monitoring, ecological research and conservation science. Methods Ecol. Evol. 2018, 9, 849–865. [Google Scholar] [CrossRef]
- ESA. The European Space Agency. Available online: https://sentinels.copernicus.eu/ (accessed on 12 December 2021).
- Jung, R.; Adolph, W.; Ehlers, M.; Farke, H. A multi-sensor approach for detecting the different land covers of tidal flats in the German Wadden Sea—A case study at Norderney. Remote Sens. Environ. 2015, 170, 188–202. [Google Scholar] [CrossRef]
- Gade, M.; Wang, W.; Kemme, L. On the imaging of exposed intertidal flats by single- and dual-co-polarization Synthetic Aperture Radar. Remote Sens. Environ. 2018, 205, 315–328. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăgu, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Khatami, R.; Mountrakis, G.; Stehman, S.V. A meta-analysis of remote sensing research on supervised pixel-based land-cover image classification processes: General guidelines for practitioners and future research. Remote Sens. Environ. 2016, 177, 89–100. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, X. Improving land cover classification in an urbanized coastal area by random forests: The role of variable selection. Remote Sens. Environ. 2020, 251, 112105. [Google Scholar] [CrossRef]
- Campredon, P.; Catry, P. Bijagos Archipelago (Guinea-Bissau). In The Wetland Book. II: Distribution, Description, and Conservation; Finlayson, C.M., Milton, G.R., Prentice, R.C., Davidson, N.C., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 1333–1340. ISBN 9789400740006. [Google Scholar]
- Cuq, F.; Campredon, P.; Giraudet, J.; Giraudet, E.; Gourmelon, F.; Pennober, G.; Simão da Silva, A. Un Système D’information Géographique Pour L’aide à la Gestion Intégrée de L’archipel des Bijagos (Guinée-Bissau): Notice de la Carte, Constitution et Exploitation du SIG; Laboratoire Géosystèmes: Brest, France, 2001; ISBN 2-9516590-0-8. [Google Scholar]
- Pennober, G. Analyse Spatiale de L’environnement Côtier de L’archipel des Bijagos (Guinée Bissau). Ph.D. Thesis, Université de Bretagne Occidentale, Brest, France, 1999. [Google Scholar]
- Temudo, M.P.; Cabral, A.I. The Social Dynamics of Mangrove Forests in Guinea-Bissau, West Africa. Hum. Ecol. 2017, 45, 307–320. [Google Scholar] [CrossRef]
- Pennober, G. Dynamique Littorale d’un Delta Estuarien: Les Bijagos (Guinée-Bissau); Institut de Géographie et D’aménagement Régional: Nantes, France, 2003. [Google Scholar]
- Granadeiro, J.P.; Andrade, J.; Palmeirim, J.M. Modelling the distribution of shorebirds in estuarine areas using generalised additive models. J. Sea Res. 2004, 52, 227–240. [Google Scholar] [CrossRef]
- Ponsero, A.; Sturbois, A.; Desroy, N.; Le Mao, P.; Jones, A.; Fournier, J. How do macrobenthic resources concentrate foraging waders in large megatidal sandflats? Estuar. Coast. Shelf Sci. 2016, 178, 120–128. [Google Scholar] [CrossRef]
- Finn, P.G.; Catterall, C.P.; Driscoll, P.V. Prey versus substrate as determinants of habitat choice in a feeding shorebird. Estuar. Coast. Shelf Sci. 2008, 80, 381–390. [Google Scholar] [CrossRef]
- Lourenço, P.M.; Catry, P.; Lecoq, M.; Ramírez, I.; Granadeiro, J.P. Role of disturbance, geology and other environmental factors in determining abundance and diversity in coastal avian communities during winter. Mar. Ecol. Prog. Ser. 2013, 479, 223–234. [Google Scholar] [CrossRef]
- Green, L.; Blumstein, D.T.; Fong, P. Macroalgal Mats in a Eutrophic Estuary Obscure Visual Foraging Cues and Increase Variability in Prey Availability for Some Shorebirds. Estuaries and Coasts 2015, 38, 917–926. [Google Scholar] [CrossRef]
- Besterman, A.F.; Karpanty, S.M.; Pace, M.L. Impact of exotic macroalga on shorebirds varies with foraging specialization and spatial scale. PLoS ONE 2020, 15, e0231337. [Google Scholar] [CrossRef]
- Everett, R.A. Macroalgae in marine soft-sediment communities: Effects on benthic faunal assemblages. J. Exp. Mar. Bio. Ecol. 1994, 175, 253–274. [Google Scholar] [CrossRef]
- Somerfield, P.J.; McClelland, I.L.; McNeill, C.L.; Bolam, S.G.; Widdicombe, S. Environmental and sediment conditions, infaunal benthic communities and biodiversity in the Celtic Sea. Cont. Shelf Res. 2019, 185, 23–30. [Google Scholar] [CrossRef]
- Philippe, A.S.; Pinaud, D.; Cayatte, M.L.; Goulevant, C.; Lachaussée, N.; Pineau, P.; Karpytchev, M.; Bocher, P. Influence of environmental gradients on the distribution of benthic resources available for shorebirds on intertidal mudflats of Yves Bay, France. Estuar. Coast. Shelf Sci. 2016, 174, 71–81. [Google Scholar] [CrossRef]
- Folk, R.L.; Andrews, P.B.; Lewis, D.W. Detrital sedimentary rock classification and nomenclature for use in new zealand. New Zeal. J. Geol. Geophys. 1970, 13, 937–968. [Google Scholar] [CrossRef]
- ESA. Sentinel-2 User Handbook; European Space Agency: Paris, France, 2015. [Google Scholar]
- Richter, R.; Schläpfer, D. Atmospheric/Topographic Correction for Satellite Imagery; ReSe Applications Schlapfer: Wil, Switzerland, 2012; ISBN 9788578110796. [Google Scholar]
- Filipponi, F. Sentinel-1 GRD Preprocessing Workflow. Proceedings 2019, 18, 11. [Google Scholar] [CrossRef]
- Medasani, S.; Reddy, G.U. Analysis and Evaluation of Speckle Filters by Using Polarimetric Synthetic Aperture Radar Data Through Local Statistics. In Proceedings of the 2018 Second International Conference on Electronics, Communication and Aerospace Technology (ICECA), Coimbatore, India, 29–31 March 2018; pp. 169–174. [Google Scholar] [CrossRef]
- Abramov, S.; Rubel, O.; Lukin, V.; Kozhemiakin, R.; Kussul, N.; Shelestov, A.; Lavreniuk, M. Speckle reducing for Sentinel-1 SAR data. In Proceedings of the International Geoscience and Remote Sensing Symposium (IGARSS), Fort Worth, TX, USA, 23–28 July 2017; Institute of Electrical and Electronics Engineers Inc.: Piscataway, NJ, USA, 2017; Volume 2017, pp. 2353–2356. [Google Scholar]
- Granadeiro, J.P.; Belo, J.; Henriques, M.; Catalão, J.; Catry, T. Using Sentinel-2 images to estimate topography, tidal-stage lags and exposure periods over large intertidal areas. Remote Sens. 2021, 13, 320. [Google Scholar] [CrossRef]
- Legendre, P. Lmodel2: Model II Regression; R Package: Vienna, Austria, 2018. [Google Scholar]
- Catalão, J. Uma nova técnica para determinar a batimetria intertidal com imagens multiespectrais do satélite Sentinel-2. In Actas das 5.as Jornadas de Engenharia Hidrog; Instituto Hidrográfico: Lisbon, Portugal, 2018; pp. 19–22. [Google Scholar]
- Bué, I.; Catalão, J.; Semedo, Á. Intertidal bathymetry extraction with multispectral images: A logistic regression approach. Remote Sens. 2020, 12, 1311. [Google Scholar] [CrossRef]
- McFeeters, S.K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017; ISBN 9781315370279. [Google Scholar]
- Commo, F.; Bot, B.M. Package nplr: N-Parameter Logistic Regression. 2016. Available online: https://mran.revolutionanalytics.com/snapshot/2015-07-13/web/packages/nplr/vignettes/nplr.pdf (accessed on 16 May 2022).
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Wilson, E.H.; Sader, S.A. Detection of forest harvest type using multiple dates of Landsat TM imagery. Remote Sens. Environ. 2002, 80, 385–396. [Google Scholar] [CrossRef]
- Xiao, X.; Boles, S.; Liu, J.; Zhuang, D.; Frolking, S.; Li, C.; Salas, W.; Moore, B. Mapping paddy rice agriculture in southern China using multi-temporal MODIS images. Remote Sens. Environ. 2005, 95, 480–492. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Qi, J.; Kerr, Y.; Chehbouni, A. External factor consideration in vegetation index development. In Proceedings of the International Symposium on Physical Measurements and Signatures in Remote Sensing, Val-d’isere, France, 17–22 January 1994. [Google Scholar]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Xiao, C.; Li, P.; Feng, Z.; Liu, Y.; Zhang, X. Sentinel-2 red-edge spectral indices (RESI) suitability for mapping rubber boom in Luang Namtha Province, northern Lao PDR. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102176. [Google Scholar] [CrossRef]
- Chen, W.; Liu, L.; Zhang, C.; Wang, J.; Wang, J.; Pan, Y. Monitoring the seasonal bare soil areas in Beijing using multi-temporal TM images. Int. Geosci. Remote Sens. Symp. 2004, 5, 3379–3382. [Google Scholar] [CrossRef]
- Xiao, J.; Shen, Y.; Tateishi, R.; Bayaer, W. Development of topsoil grain size index for monitoring desertification in arid land using remote sensing. Int. J. Remote Sens. 2006, 27, 2411–2422. [Google Scholar] [CrossRef]
- Tuszynski, J. caTools: Moving Window Statistics, GIF, Base64, ROC AUC, etc. R package version 1. 2021. [Google Scholar]
- Liaw, A.; Wiener, M. Package ‘randomForest’: Breiman and Cutler’s Random Forests for Classification and Regression. R package version 4. 2018. [Google Scholar]
- Gregorutti, B.; Michel, B.; Saint-Pierre, P. Correlation and variable importance in random forests. Stat. Comput. 2017, 27, 659–678. [Google Scholar] [CrossRef]
- Pelletier, C.; Valero, S.; Inglada, J.; Champion, N.; Dedieu, G. Assessing the robustness of Random Forests to map land cover with high resolution satellite image time series over large areas. Remote Sens. Environ. 2016, 187, 156–168. [Google Scholar] [CrossRef]
- Speiser, J.L.; Miller, M.E.; Tooze, J.; Ip, E. A comparison of random forest variable selection methods for classification prediction modeling. Expert Syst. Appl. 2019, 134, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Lunetta, K.L.; Hayward, L.B.; Segal, J.; van Eerdewegh, P. Screening large-scale association study data: Exploiting interactions using random forests. BMC Genet. 2004, 5, 32. [Google Scholar] [CrossRef]
- Diaz-Uriarte, R. GeneSrF and varSelRF: A web-based tool and R package for gene selection and classification using random forest. BMC Bioinform. 2007, 8, 328. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.M.; Tuleau-Malot, C. VSURF: An R package for variable selection using random forests. R J. 2015, 7, 19–33. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef]
- Lu, Q.; Huang, X.; Liu, T.; Zhang, L. A structural similarity-based label-smoothing algorithm for the post-processing of land-cover classification. Remote Sens. Lett. 2016, 7, 437–445. [Google Scholar] [CrossRef]
- Zhu, X. Land cover classification using moderate resolution satellite imagery and random forests with post-hoc smoothing. J. Spat. Sci. 2013, 58, 323–337. [Google Scholar] [CrossRef]
- Huang, X.; Lu, Q.; Zhang, L.; Plaza, A. New postprocessing methods for remote sensing image classification: A systematic study. IEEE Trans. Geosci. Remote Sens. 2014, 52, 7140–7159. [Google Scholar] [CrossRef]
- Schindler, K. An overview and comparison of smooth labeling methods for land-cover classification. IEEE Trans. Geosci. Remote Sens. 2012, 50, 4534–4545. [Google Scholar] [CrossRef]
- Kuhn, M. Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar]
- Velez, D.R.; White, B.C.; Motsinger, A.A.; Bush, W.S.; Ritchie, M.D.; Williams, S.M.; Moore, J.H. A balanced accuracy function for epistasis modeling in imbalanced datasets using multifactor dimensionality reduction. Genet. Epidemiol. 2007, 31, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, M.; Leutner, B.; Dech, S. Remote Sensing and GIS for Ecologists: Using Open Source Software; Wegmann, M., Leutner, B., Dech, S., Eds.; Pelagic Publishing: Exeter, UK, 2016; ISBN 978-1-78427-028-5. [Google Scholar]
- Stehman, S.V. Selecting and interpreting measures of thematic classification accuracy. Remote Sens. Environ. 1997, 62, 77–89. [Google Scholar] [CrossRef]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Kim, K.L.; Kim, B.J.; Lee, Y.K.; Ryu, J.H. Generation of a large-scale surface sediment classification map using unmanned aerial vehicle (UAV) data: A case study at the Hwang-do tidal flat, Korea. Remote Sens. 2019, 11, 229. [Google Scholar] [CrossRef]
- Adolph, W.; Farke, H.; Lehner, S.; Ehlers, M. Remote sensing intertidal flats with TerraSAR-X. A SAR perspective of the structural elements of a tidal basin for monitoring the Wadden Sea. Remote Sens. 2018, 10, 1085. [Google Scholar] [CrossRef]
- Kohlus, J.; Stelzer, K.; Müller, G.; Smollich, S. Mapping seagrass (Zostera) by remote sensing in the Schleswig-Holstein Wadden Sea. Estuar. Coast. Shelf Sci. 2020, 238, 106699. [Google Scholar] [CrossRef]
- Fitton, J.M.; Rennie, A.F.; Hansom, J.D.; Muir, F.M.E. Remotely sensed mapping of the intertidal zone: A Sentinel-2 and Google Earth Engine methodology. Remote Sens. Appl. Soc. Environ. 2021, 22, 100499. [Google Scholar] [CrossRef]
- Bousbih, S.; Zribi, M.; Pelletier, C.; Gorrab, A.; Lili-Chabaane, Z.; Baghdadi, N.; Aissa, N.B.; Mougenot, B. Soil texture estimation using radar and optical data from Sentinel-1 and Sentinel-2. Remote Sens. 2019, 11, 1520. [Google Scholar] [CrossRef]
- Ibrahim, E.; Adam, S.; De Wever, A.; Govaerts, A.; Vervoort, A.; Monbaliu, J. Investigating spatial resolutions of imagery for intertidal sediment characterization using geostatistics. Cont. Shelf Res. 2014, 85, 117–125. [Google Scholar] [CrossRef]
- Wang, W.; Yang, X.; Li, X.; Chen, K.; Liu, G.; Li, Z.; Gade, M. A Fully Polarimetric SAR Imagery Classification Scheme for Mud and Sand Flats in Intertidal Zones. IEEE Trans. Geosci. Remote Sens. 2017, 55, 1734–1742. [Google Scholar] [CrossRef]
- Deroin, J.P. Combining ALOS and ERS-2 SAR data for the characterization of tidal flats. Case study from the Baie des Veys, Normandy, France. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 183–194. [Google Scholar] [CrossRef]
- Wang, X.; Xiao, X.; Zou, Z.; Chen, B.; Ma, J.; Dong, J.; Doughty, R.B.; Zhong, Q.; Qin, Y.; Dai, S.; et al. Tracking annual changes of coastal tidal flats in China during 1986–2016 through analyses of Landsat images with Google Earth Engine. Remote Sens. Environ. 2020, 238, 110987. [Google Scholar] [CrossRef]
- Ryu, J.H.; Na, Y.H.; Won, J.S.; Doerffer, R. A critical grain size for Landsat ETM+ investigations into intertidal sediments: A case study of the Gomso tidal flats, Korea. Estuar. Coast. Shelf Sci. 2004, 60, 491–502. [Google Scholar] [CrossRef]
- Choi, J.K.; Eom, J.A.; Ryu, J.H. Spatial relationships between surface sedimentary facies distribution and topography using remotely sensed data: Example from the Ganghwa tidal flat, Korea. Mar. Geol. 2011, 280, 205–211. [Google Scholar] [CrossRef]
- Instituto Hidrográfico. Vol II Países Africanos de Língua Oficial Portuguesa e Macau. In Tabelas de Marés; Instituto Hidrográfico: Lisboa, Portugal, 2021. [Google Scholar]
- Hill, N.K.; Woodworth, B.K.; Phinn, S.R.; Murray, N.J.; Fuller, R.A. Global protected-area coverage and human pressure on tidal flats. Conserv. Biol. 2021, 35, 933–943. [Google Scholar] [CrossRef]
- Zwarts, L. The winter exploitation of Fiddler Crabs Uca tangeri by waders in Guinea-Bissau. Ardea 1985, 73, 3–12. [Google Scholar]
- East Atlantic Flyway Assessment 2020: The Status of Coastal Waterbird Populations and Their Sites; Van Roomen, M., Citegetse, G., Crowe, O., Dodman, T., Hagemeijer, W., Meise, K., Schekkerman, H., Eds.; Wadden Sea Flyway Initiative: Wilhelmshaven, Germany; Wetlands International: Wageningen, The Netherlands; BirdLife International: Cambridge, UK, 2022. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictors | Sensor (Reference) | Description |
|---|---|---|
| Band 2—Blue (B) | Sentinel-2 | Native resolution: 10 m; Central wavelength: 490 nm |
| Band 3—Green (G) | Sentinel-2 | Native resolution: 10 m; Central wavelength: 560 nm |
| Band 4—Red (R) | Sentinel-2 | Native resolution: 10 m; Central wavelength: 665 nm |
| Band 5—Vegetation Red Edge (Re1) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 705 nm |
| Band 6—Vegetation Red Edge (Re2) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 740 nm |
| Band 7—Vegetation Red Edge (Re3) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 783 nm |
| Band 8—Near Infrared (NIR) | Sentinel-2 | Native resolution: 10 m; Central wavelength: 842 nm |
| Band 8A—Near Infrared narrow (NIRn) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 865 nm |
| Band 11—Short-wave infrared (SWIR1) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 1610 nm |
| Band 12—Short-wave infrared (SWIR2) | Sentinel-2 | Native resolution: 20 m; Central wavelength: 2190 nm |
| NDWI—Normalised Vegetation Water Index | Sentinel-2 [73] | (G − NIR)/(G + NIR) |
| mNDWI—modified Normalised Difference Water Index | Sentinel-2 [76] | (G − SWIR1)/(G + SWIR1) |
| NDMI—Normalised Difference Moisture Index | Sentinel-2 [77] | (NIR − SWIR1)/(NIR + SWIR1) |
| nNDMI—narrow Normalised Difference Moisture Index | Sentinel-2 [78] | (NIRn − SWIR1)/(NIRn + SWIR1) |
| NDVI—Normalised Difference Vegetation Index | Sentinel-2 [79] | (NIR − R)/(NIR + R) |
| MSAVI Version 2—Modified Soil-Adjusted Vegetation Index | Sentinel-2 [80,81] | (2NIR + 1 − √(2NIR + 1)2 − 8(NIR − R))/2 |
| RESI—Red-edge Normalised Index | Sentinel-2 [82] | (Re3 + Re2 − Re1)/(Re3 + Re2 + Re1) |
| BSI—Bare Sediment Index | Sentinel-2 [83] | (SWIR1 + R) − (NIR + B))/(SWIR1 + R) + (NIR + B)) |
| GSI—Topsoil Grain Size Index | Sentinel-2 [84] | (R + B)/(R + B + G) |
| S1-VV polarisation—vertical transmit, vertical receive | Sentinel-1 | Native resolution: 5 m × 20 m, resampled to 10 m × 10 m |
| S1-VH cross polarisation—vertical transmit, vertical receive | Sentinel-1 | Native resolution: 5 m × 20 m, resampled to 10 m × 10 m |
| RVI—Radar Vegetation Index | Sentinel-1 | 4σVH0/(σVV0 + σVH0) |
| S1-VH/VV—Radar Cross-polarisation ratio | Sentinel-1 | σVH0/σVV0 |
| Digital Elevation Model (DEM) | Sentinel-2 [69] | Including heights ranging 1.65–4.69 m |
| Habitat Types | Defining Thresholds | Median Water Film Cover (%) | Mud Content (Mean % ± SD) | n Polygons | n Pixels |
|---|---|---|---|---|---|
| Rocks | ≥30% of rock cover. | 0 | N/A | 88 | 428 |
| Shell beds | ≥30% of shell cover. | 20 | N/A | 53 | 609 |
| Macroalgae | ≥30% of macroalgae cover and ≤50 FCB cover. | 30 | N/A | 116 | 3155 |
| Bare sediment sandy | ≤50% of high-density FCB cover and ≤30% of rocks, shells, and macroalgae cover and <10% of mud content. | 35 | 2 ± 2.3 | 408 | 12,777 |
| Bare sediment mixed | ≤50% of high-density FCB cover and ≤30% of rocks, shells, and macroalgae cover and ≥10% of mud content. | 30 | 15.3 ± 6.5 | 36 | 1219 |
| FCB sandy | ≥50% of high-density FCB cover and ≤30% of rocks, shells, and macroalgae cover and <10% of mud content. | 0 | 4.7 ± 2.9 | 179 | 4119 |
| FCB mixed | ≥50% of high-density FCB cover and ≤30% of rocks, shells, and macroalgae cover and ≥10% of mud content. | 5 | 19.9 ± 10.3 | 143 | 3534 |
| Predictions | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Rocks | Shell Beds | Macroalgae | Bare Sediment Sandy | Bare Sediment Mixed | FCB Sandy | FCB Mixed | Total | User’s Accuracy | |
| Rocks | 95 | 0 | 41 | 1 | 0 | 0 | 0 | 137 | 69.3 |
| Shell beds | 3 | 107 | 0 | 8 | 0 | 0 | 0 | 118 | 90.7 |
| Macroalgae | 7 | 3 | 685 | 23 | 3 | 3 | 90 | 814 | 84.2 |
| Bare sediment sandy | 14 | 56 | 184 | 3573 | 219 | 192 | 26 | 4264 | 83.8 |
| Bare sediment mixed | 0 | 0 | 18 | 2 | 132 | 32 | 4 | 188 | 70.2 |
| FCB sandy | 1 | 8 | 11 | 186 | 4 | 929 | 168 | 1307 | 71.1 |
| FCB mixed | 8 | 9 | 8 | 40 | 8 | 80 | 772 | 925 | 83.5 |
| Total | 128 | 183 | 947 | 3833 | 366 | 1236 | 1060 | 7753 | - |
| Producer’s Accuracy | 74.2 | 58.5 | 72.3 | 93.2 | 36.1 | 75.2 | 72.8 | - | |
| Overall Accuracy | 81.2 | ||||||||
| Kappa Coefficient | 71.9 | ||||||||
| Stats | Unfiltered | Filtered | |
|---|---|---|---|
| Overall Accuracy | 81.2 | 81.8 | |
| Kappa Coefficient | 71.9 | 73.1 | |
| Rocks | User’s Accuracy | 69.3 | 66.2 |
| Producer’s Accuracy | 74.2 | 76.6 | |
| Shell beds | User’s Accuracy | 90.7 | 90.1 |
| Producer’s Accuracy | 58.5 | 59.6 | |
| Macroalgae | User’s Accuracy | 84.2 | 88.3 |
| Producer’s Accuracy | 72.3 | 73.8 | |
| Bare sediment sandy | User’s Accuracy | 83.8 | 83.8 |
| Producer’s Accuracy | 93.2 | 93.6 | |
| Bare sediment mixed | User’s Accuracy | 70.2 | 72.4 |
| Producer’s Accuracy | 36.1 | 36.7 | |
| Fiddler crab burrow sandy | User’s Accuracy | 71.1 | 72.3 |
| Producer’s Accuracy | 75.2 | 75.4 | |
| Fiddler crab burrow mixed | User’s Accuracy | 83.5 | 84.0 |
| Producer’s Accuracy | 72.8 | 75.1 | |
| Habitat Type | Area (km2) | % of Total |
|---|---|---|
| Rocks | 15.7 | 3.7 |
| Shell beds | 6.7 | 1.6 |
| Macroalgae | 49.6 | 11.6 |
| Bare sediment sandy | 252.3 | 59.1 |
| Bare sediment mixed | 7.9 | 1.9 |
| FCB sandy | 38.2 | 8.9 |
| FCB mixed | 56.3 | 13.2 |
| Total | 426.8 | 100.0 |
| Stats | Final Model S2 + S1 + DEM 14 Predictors | S2 Model 14 Predictors | S2 + S1 Model 14 Predictors | S2 + DEM Model 16 Predictors | |
|---|---|---|---|---|---|
| Overall Accuracy | 81.2 | 77.3 | 80.3 | 78.4 | |
| Kappa | 71.9 | 66.1 | 70.6 | 67.5 | |
| Rocks | User’s Accuracy | 69.3 | 92.9 | 75.2 | 88.2 |
| Producer’s Accuracy | 74.2 | 61.7 | 68.8 | 64.1 | |
| Shell beds | User’s Accuracy | 90.7 | 59.9 | 91.7 | 72.4 |
| Producer’s Accuracy | 58.5 | 48.1 | 54.7 | 53.0 | |
| Macroalgae | User’s Accuracy | 84.2 | 75.7 | 82.7 | 79.3 |
| Producer’s Accuracy | 72.3 | 77.5 | 72.0 | 78.7 | |
| Bare sediment sandy | User’s Accuracy | 83.8 | 80.9 | 83.4 | 81.3 |
| Producer’s Accuracy | 93.2 | 90.9 | 93.0 | 92.1 | |
| Bare sediment mixed | User’s Accuracy | 70.2 | 58.0 | 60.3 | 62.2 |
| Producer’s Accuracy | 36.1 | 28.7 | 32.8 | 27.9 | |
| FCB sandy | User’s Accuracy | 71.1 | 67.1 | 70.5 | 67.3 |
| Producer’s Accuracy | 75.2 | 63.3 | 71.6 | 64.9 | |
| FCB mixed | User’s Accuracy | 83.5 | 80.4 | 80.8 | 81.0 |
| Producer’s Accuracy | 72.8 | 68.1 | 74.1 | 67.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriques, M.; Catry, T.; Belo, J.R.; Piersma, T.; Pontes, S.; Granadeiro, J.P. Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds. Remote Sens. 2022, 14, 3260. https://doi.org/10.3390/rs14143260
Henriques M, Catry T, Belo JR, Piersma T, Pontes S, Granadeiro JP. Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds. Remote Sensing. 2022; 14(14):3260. https://doi.org/10.3390/rs14143260
Chicago/Turabian StyleHenriques, Mohamed, Teresa Catry, João Ricardo Belo, Theunis Piersma, Samuel Pontes, and José Pedro Granadeiro. 2022. "Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds" Remote Sensing 14, no. 14: 3260. https://doi.org/10.3390/rs14143260
APA StyleHenriques, M., Catry, T., Belo, J. R., Piersma, T., Pontes, S., & Granadeiro, J. P. (2022). Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds. Remote Sensing, 14(14), 3260. https://doi.org/10.3390/rs14143260