
Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR


 , , ,
, , ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
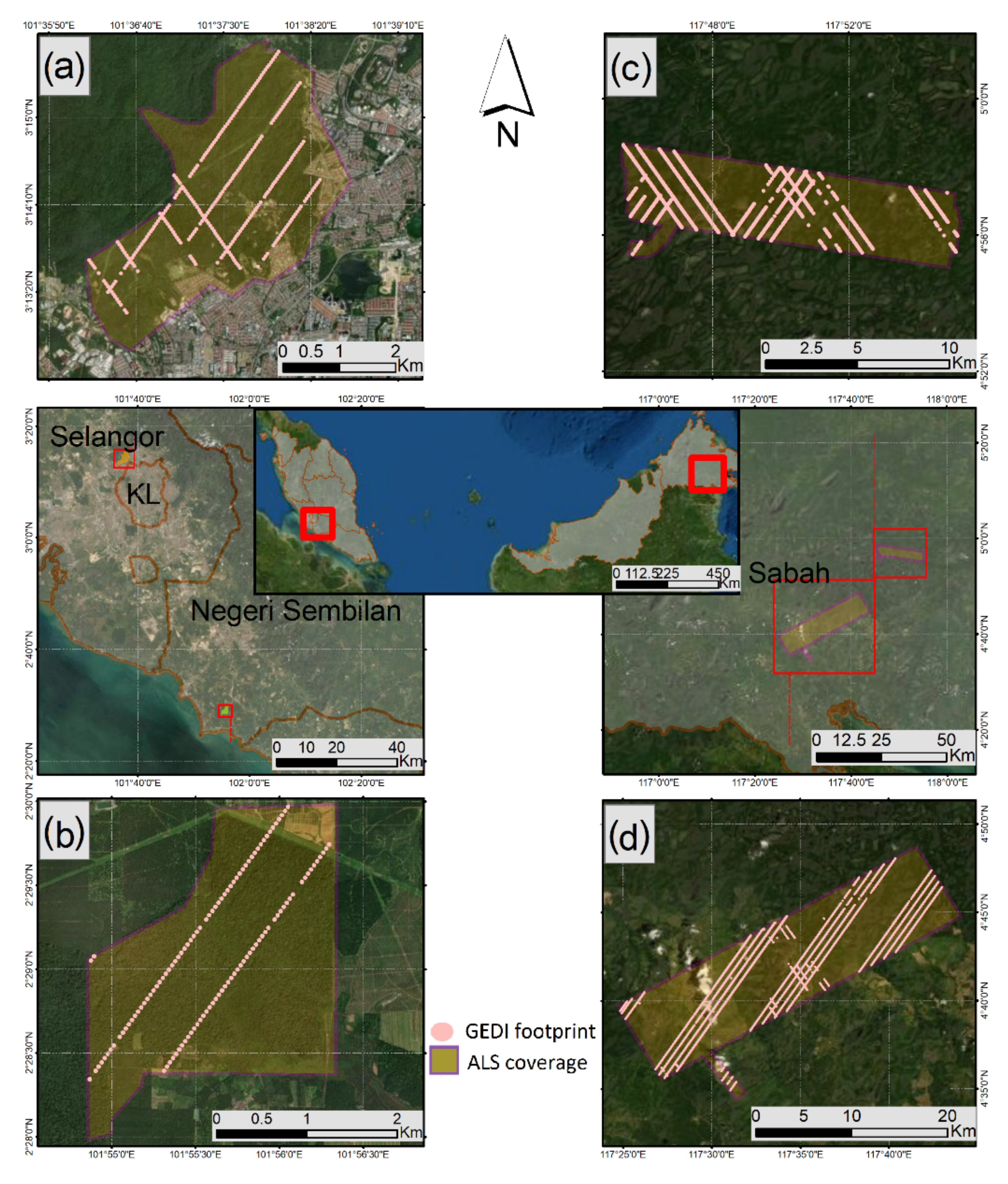
2.1. Study Sites
2.2. Remote Sensing Data
2.2.1. Airborne Laser Scanning Data and Data Pre-processing
2.2.2. GEDI Data
2.2.3. Climatic and Topographic Data
2.3. ALS and GEDI Canopy Height Comparison
2.4. Canopy Height Modeling and Variables Importance
2.4.1. Machine Learning Algorithms
2.4.2. Model Development and Feature Selection
2.4.3. Model Validation
2.5. Exploring the Bivariate and Multivariate Relationships
2.5.1. Bivariate Relationships
2.5.2. Multivariate Relationships
3. Results
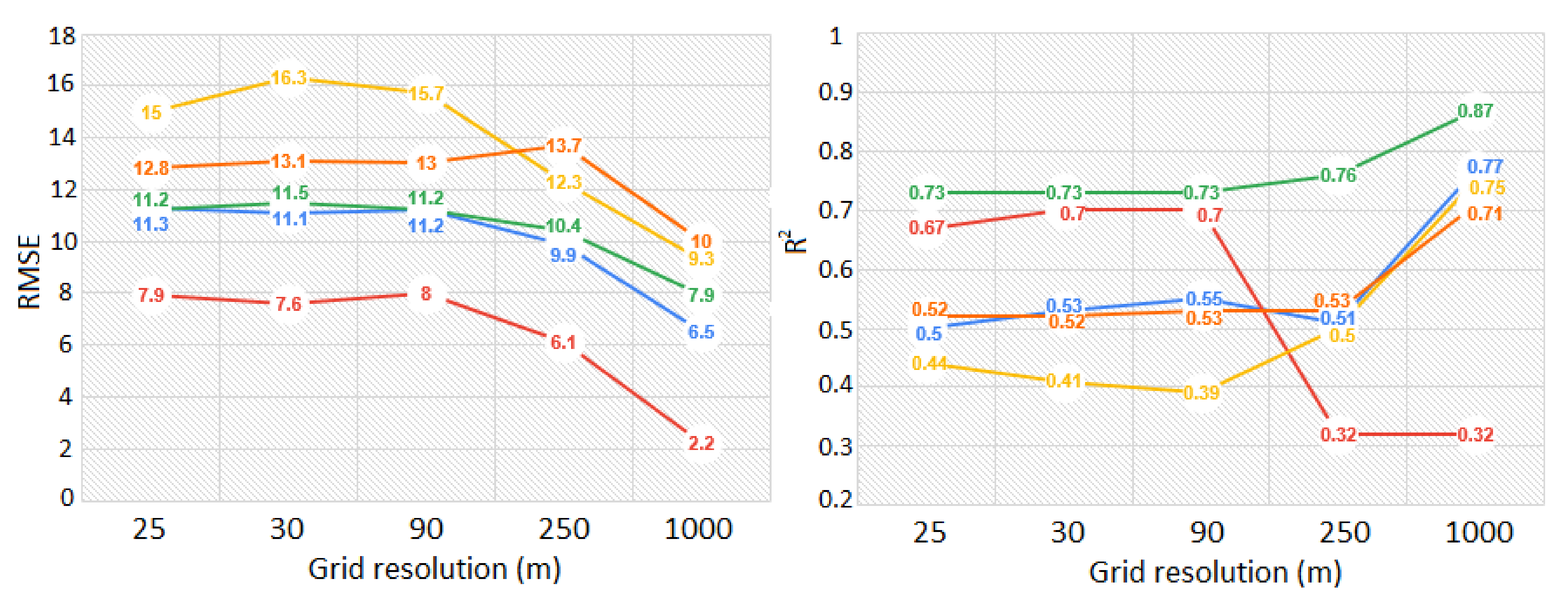
3.1. Comparison between ALS and GEDI-Derived Canopy Height
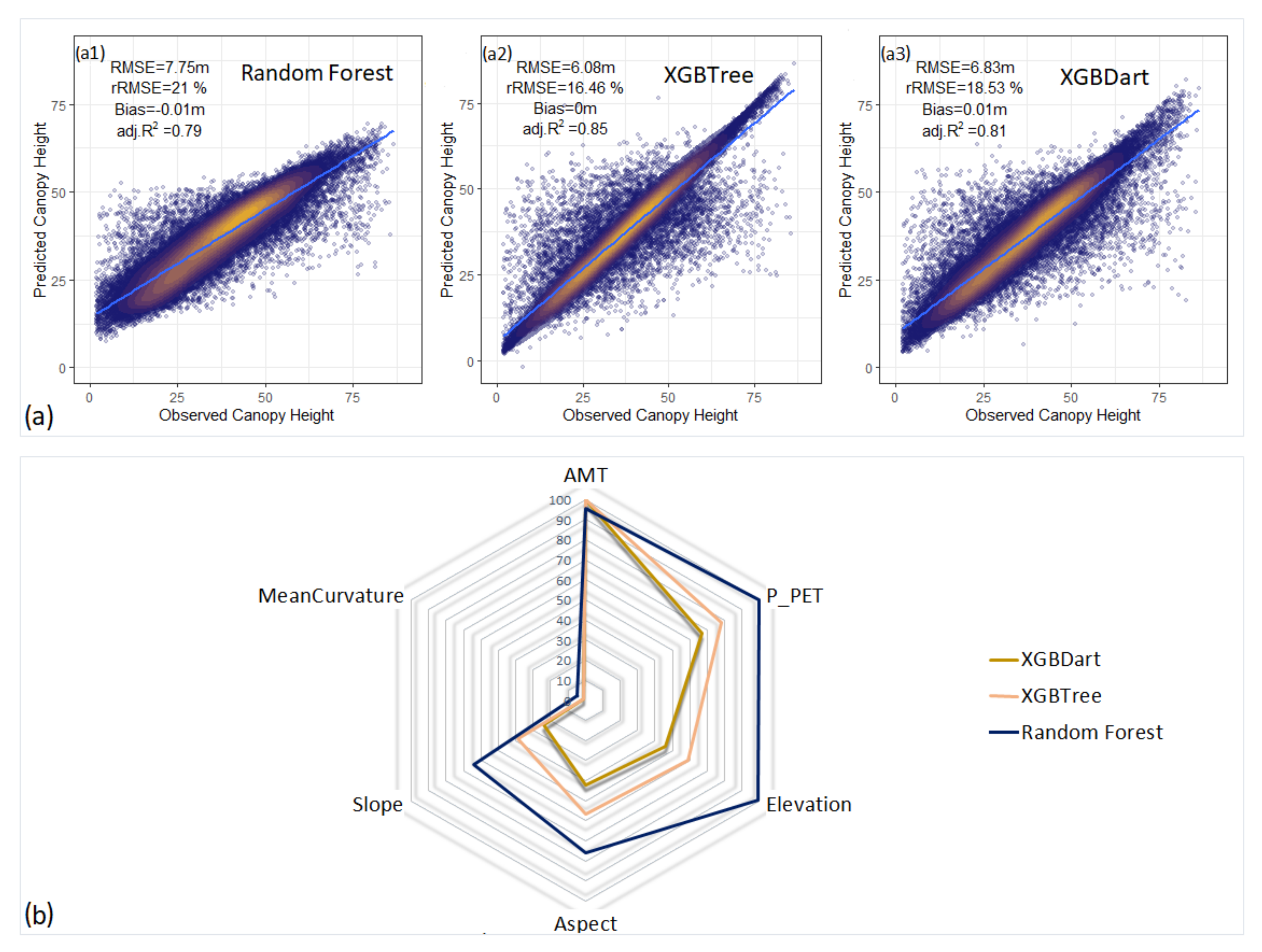
3.2. Machine Learning-Derived Canopy Height Models
3.3. Exploring the Bivariate and Multivariate Relationships
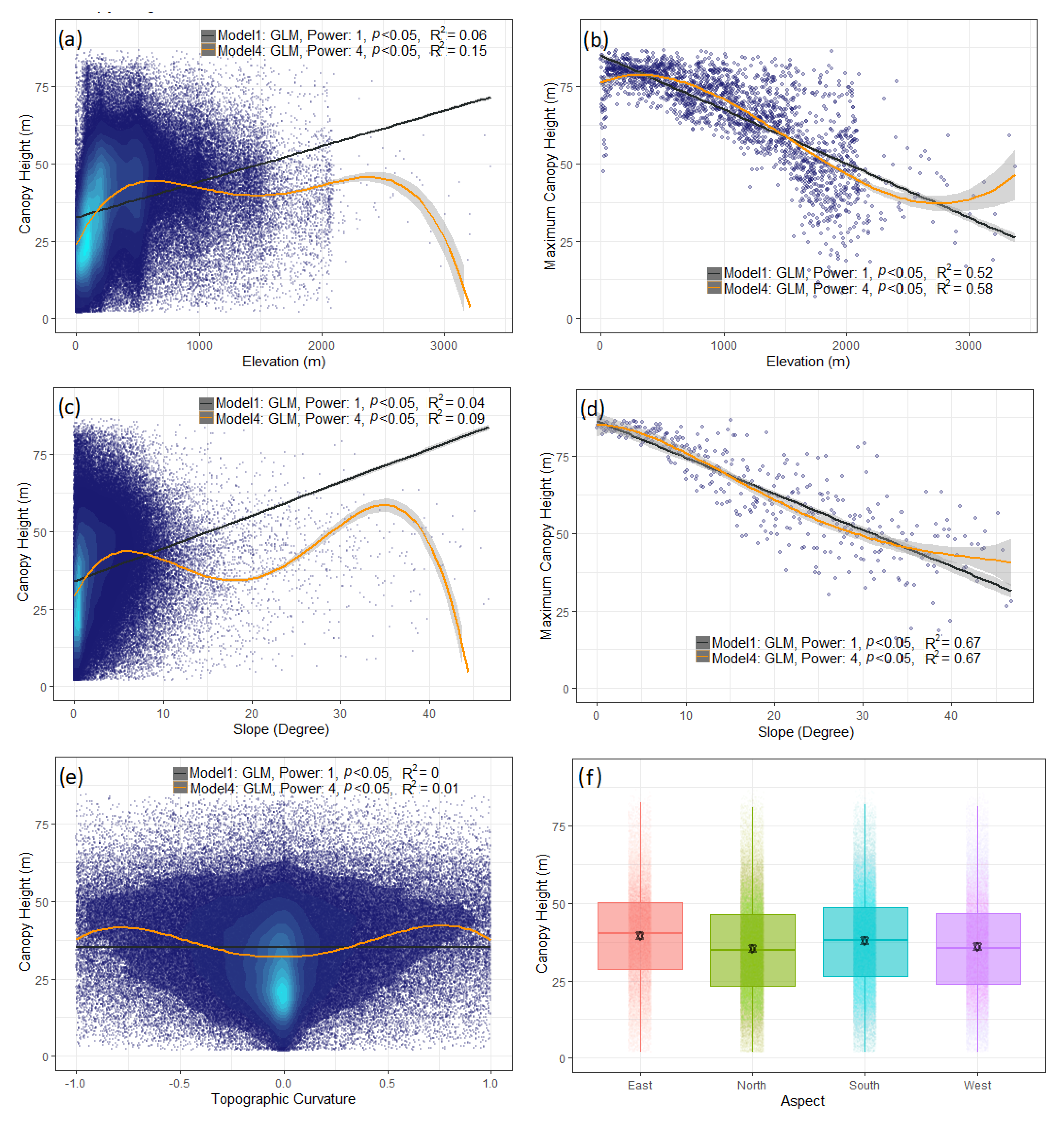
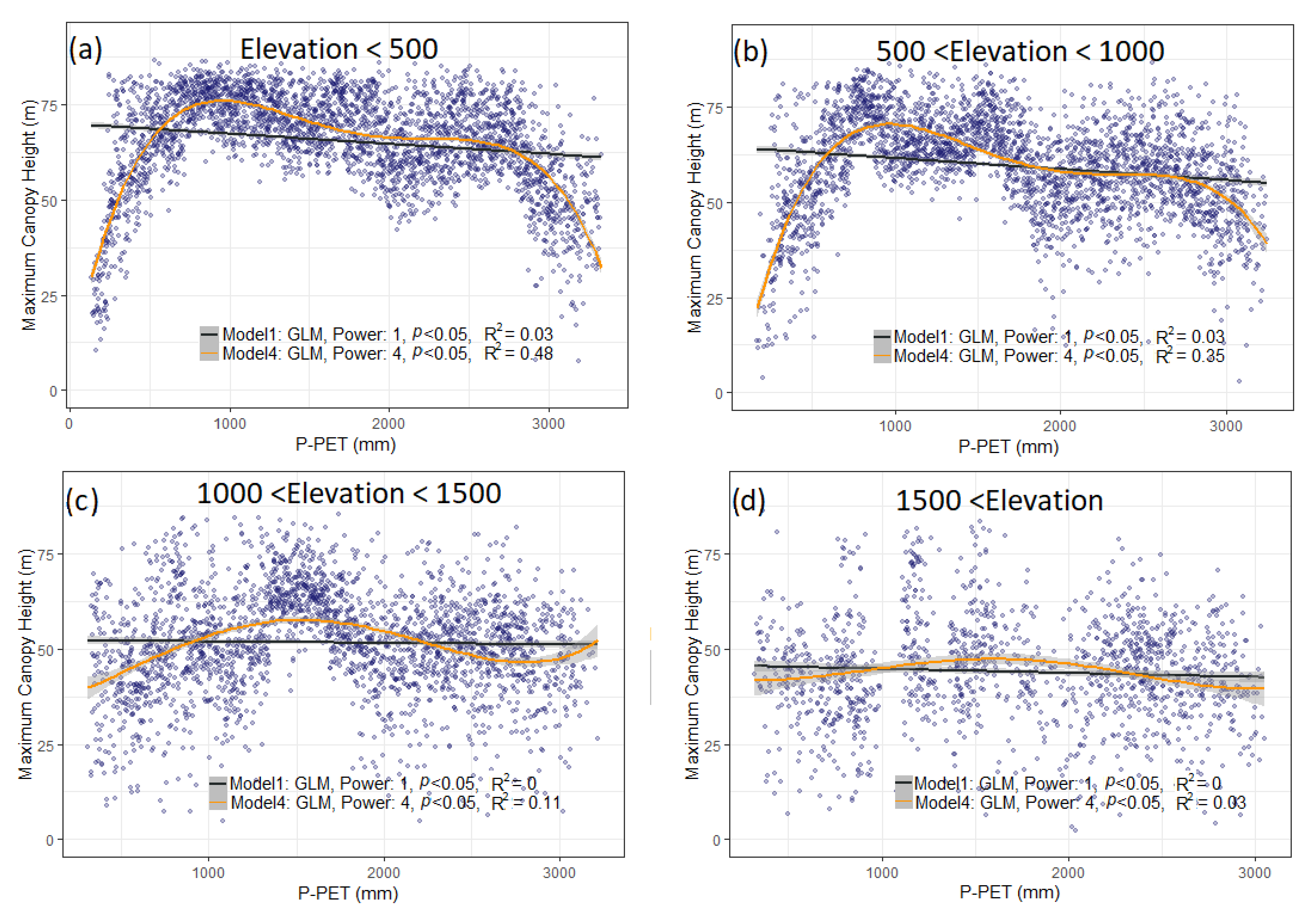
3.3.1. Bivariate Analysis for Canopy Height and Climatic Variables
3.3.2. Bivariate Analysis for Canopy Height and Topographical Variables
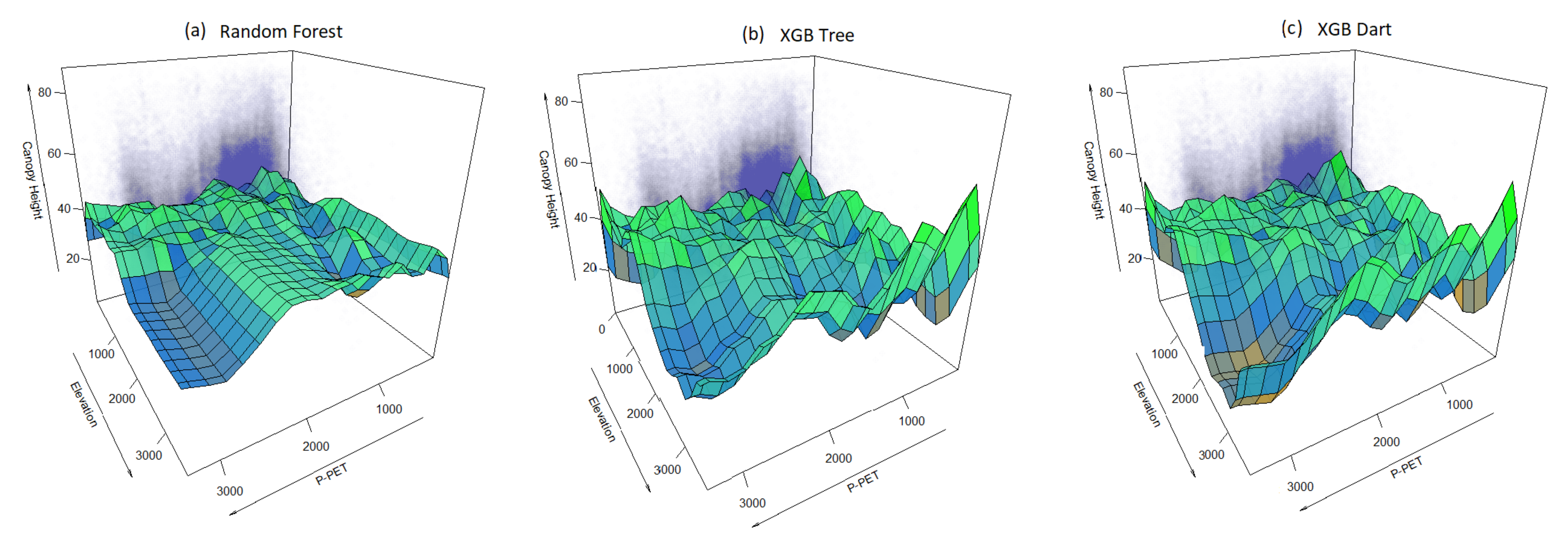
3.3.3. Multivariate Analysis for Canopy Height, Elevation and P-PET
4. Discussion
4.1. GEDI Validation and Resampling
4.2. Canopy Height Relationship with Climatic and Topographic Variables
4.2.1. Temperature
4.2.2. Water Availability
4.2.3. Topographic Variables
4.2.4. The Multivariate Relationship between P-PET-Elevation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, P.; Zhou, T.; Yi, C.; Fang, W.; Hendrey, G.; Zhao, X. Forest Drought Resistance Distinguished by Canopy Height. Environ. Res. Lett. 2018, 13, 075003. [Google Scholar] [CrossRef] [Green Version]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-Evaluation of Forest Biomass Carbon Stocks and Lessons from the World’s Most Carbon-Dense Forests. Proc. Natl. Acad. Sci. USA 2009, 106, 11635–11640. [Google Scholar] [CrossRef] [Green Version]
- Lefsky, M.A.; Harding, D.J.; Keller, M.; Cohen, W.B.; Carabajal, C.C.; Del Bom Espirito-Santo, F.; Hunter, M.O.; de Oliveira, R., Jr. Estimates of Forest Canopy Height and Aboveground Biomass Using ICESat: ICESAT ESTIMATES OF CANOPY HEIGHT. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Marselis, S.M.; Tang, H.; Armston, J.; Abernethy, K.; Alonso, A.; Barbier, N.; Bissiengou, P.; Jeffery, K.; Kenfack, D.; Labrière, N.; et al. Exploring the Relation between Remotely Sensed Vertical Canopy Structure and Tree Species Diversity in Gabon. Environ. Res. Lett. 2019, 14, 094013. [Google Scholar] [CrossRef]
- Xu, P.; Zhou, T.; Yi, C.; Luo, H.; Zhao, X.; Fang, W.; Gao, S.; Liu, X. Impacts of Water Stress on Forest Recovery and Its Interaction with Canopy Height. Int. J. Environ. Res. Public Health 2018, 15, 1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubayah, R.O.; Sheldon, S.L.; Clark, D.B.; Hofton, M.A.; Blair, J.B.; Hurtt, G.C.; Chazdon, R.L. Estimation of Tropical Forest Height and Biomass Dynamics Using Lidar Remote Sensing at La Selva, Costa Rica: FOREST DYNAMICS USING LIDAR. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; Mcnulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J. Climate Change and Forest Disturbances: Climate Change Can Affect Forests by Altering the Frequency, Intensity, Duration, and Timing of Fire, Drought, Introduced Species, Insect and Pathogen Outbreaks, Hurricanes, Windstorms, Ice Storms, or Landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The Limits to Tree Height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global Patterns in Plant Height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Larjavaara, M.; Auvinen, M.; Kantola, A.; Mäkelä, A. Wind and Gravity in Shaping Picea Trunks. Trees 2021, 35, 1587–1599. [Google Scholar] [CrossRef]
- Wang, B.; Fang, S.; Wang, Y.; Guo, Q.; Hu, T.; Mi, X.; Lin, L.; Jin, G.; Coomes, D.A.; Yuan, Z.; et al. The Shift from Energy to Water Limitation in Local Canopy Height from Temperate to Tropical Forests in China. Forests 2022, 13, 639. [Google Scholar] [CrossRef]
- Tao, S.; Guo, Q.; Li, C.; Wang, Z.; Fang, J. Global Patterns and Determinants of Forest Canopy Height. Ecology 2016, 97, 3265–3270. [Google Scholar] [CrossRef]
- Zhang, J.; Nielsen, S.E.; Mao, L.; Chen, S.; Svenning, J.-C. Regional and Historical Factors Supplement Current Climate in Shaping Global Forest Canopy Height. J. Ecol. 2016, 104, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Fricker, G.A.; Synes, N.W.; Serra-Diaz, J.M.; North, M.P.; Davis, F.W.; Franklin, J. More than Climate? Predictors of Tree Canopy Height Vary with Scale in Complex Terrain, Sierra Nevada, CA (USA). For. Ecol. Manag. 2019, 434, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Farhadur Rahman, M.; Onoda, Y.; Kitajima, K. Forest Canopy Height Variation in Relation to Topography and Forest Types in Central Japan with LiDAR. For. Ecol. Manag. 2022, 503, 119792. [Google Scholar] [CrossRef]
- Dubayah, R.; Rich, P.M. Topographic Solar Radiation Models for GIS. Int. J. Geogr. Inf. Syst. 1995, 9, 405–419. [Google Scholar] [CrossRef]
- Geroy, I.J.; Gribb, M.M.; Marshall, H.P.; Chandler, D.G.; Benner, S.G.; McNamara, J.P. Aspect Influences on Soil Water Retention and Storage: ASPECT AND SOIL WATER RETENTION. Hydrol. Processes 2011, 25, 3836–3842. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Davies, S.J.; Chuyong, G.B.; Kenfack, D.; et al. Soil Resources and Topography Shape Local Tree Community Structure in Tropical Forests. Proc. Biol. Sci. 2013, 280, 20122532. [Google Scholar] [CrossRef]
- Klein, T.; Randin, C.; Körner, C. Water Availability Predicts Forest Canopy Height at the Global Scale. Ecol. Lett. 2015, 18, 1311–1320. [Google Scholar] [CrossRef]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping Forest Canopy Height Globally with Spaceborne Lidar. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Cheng, X.; Chen, Z. Performance Evaluation of GEDI and ICESat-2 Laser Altimeter Data for Terrain and Canopy Height Retrievals. Remote Sens. Environ. 2021, 264, 112571. [Google Scholar] [CrossRef]
- Dubayah, R.; Blair, J.B.; Goetz, S.; Fatoyinbo, L.; Hansen, M.; Healey, S.; Hofton, M.; Hurtt, G.; Kellner, J.; Luthcke, S.; et al. The Global Ecosystem Dynamics Investigation: High-Resolution Laser Ranging of the Earth’s Forests and Topography. Sci. Remote Sens. 2020, 1, 100002. [Google Scholar] [CrossRef]
- Abdul Razak, M.A.; Mohamed, M.; Alona, C.L.; Omar, H.; Misman, M.A. Tree Species Richness, Diversity and Distribution at Sungai Menyala Forest Reserve, Negeri Sembilan. IOP Conf. Ser. Earth Environ. Sci. 2019, 269, 012003. [Google Scholar] [CrossRef]
- Wyatt-Smith, J. Ecological Studies on Malayan Forests. Composition and Dynamic Studies in Lowland Evergreen Rain Forest in Two 5-Acre Plots in Bukit Lagong and Sungei Menyala Forest Reserves and in Two Half-Acre Plots in Sungei Menyala Forest; Research Pamphlet No. 101; Forest Research Institute, Forest Department: Kuala Lumpur, Malaysia, 1966. [Google Scholar]
- Nunes, M.; Ewers, R.; Turner, E.; Coomes, D. Mapping Aboveground Carbon in Oil Palm Plantations Using LiDAR: A Comparison of Tree-Centric versus Area-Based Approaches. Remote Sens. 2017, 9, 816. [Google Scholar] [CrossRef] [Green Version]
- Swinfield, T.; Both, S.; Riutta, T.; Bongalov, B.; Elias, D.; Majalap-Lee, N.; Ostle, N.; Svátek, M.; Kvasnica, J.; Milodowski, D.; et al. Imaging Spectroscopy Reveals the Effects of Topography and Logging on the Leaf Chemistry of Tropical Forest Canopy Trees. Glob. Chang. Biol. 2020, 26, 989–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swinfield, T.; Milodowski, D.; Jucker, T.; Michele, D.; Coomes, D. LiDAR Canopy Structure 2014, 2020 [Data set], Zenodo. Available online: https://doi.org/10.5281/zenodo.4020697 (accessed on 1 March 2022).
- Orme, D. Safe Web Safeproject.net. Available online: https://www.safeproject.net (accessed on 1 March 2022).
- CEDA Archive Web Browser. Available online: https://data.ceda.ac.uk/ (accessed on 1 March 2022).
- Roussel, J.-R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Meador, A.S.; Bourdon, J.-F.; de Boissieu, F.; Achim, A. LidR: An R Package for Analysis of Airborne Laser Scanning (ALS) Data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- Duncanson, L.; Kellner, J.R.; Armston, J.; Dubayah, R.; Minor, D.M.; Hancock, S.; Healey, S.P.; Patterson, P.L.; Saarela, S.; Marselis, S.; et al. Aboveground biomass density models for NASA’s Global Ecosystem Dynamics Investigation (GEDI) lidar mission. Remote Sens. Environ. 2022, 270, 112845. [Google Scholar] [CrossRef]
- Dubayah, R.; Hofton, M.; Blair, J.; Armston, J.; Tang, H.; Luthcke, S. GEDI L2A Elevation and Height Metrics Data Global Footprint Level V002; NASA EOSDIS Land Processes DAAC: Greenbelt, MD, USA, 2021. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 March 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas: NEW CLIMATE SURFACES FOR GLOBAL LAND AREAS. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Trabucco, A.; Zomer, R. Global Aridity Index and Potential Evapotranspiration (ET0); Climate Database; Figshare: Iasi, Romania, 2022; Volume 3. [Google Scholar] [CrossRef]
- Safanelli, J.; Poppiel, R.; Ruiz, L.; Bonfatti, B.; Mello, F.; Rizzo, R.; Demattê, J. Terrain Analysis in Google Earth Engine: A Method Adapted for High-Performance Global-Scale Analysis. ISPRS Int. J. Geoinf. 2020, 9, 400. [Google Scholar] [CrossRef]
- Zhu, J.; Shi, Y.; Fang, L.; Liu, X.; Ji, C. Patterns and Determinants of Wood Physical and Mechanical Properties across Major Tree Species in China. Sci. China Life Sci. 2015, 58, 602–612. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.-L.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional Variable Importance for Random Forests. BMC Bioinformatics 2008, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Strobl, C.; Zeileis, A. Danger: High Power! Exploring the Statistical Properties of a Test for Random Forest Variable Importance; Universitätsbibliothek der Ludwig-Maximilians-Universität München: Munich, Germany, 2008. [Google Scholar] [CrossRef]
- Kuhn, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel., B. The Caret Package. Vienna, Austria, 2012. Available online: https://cran.r-project.org/package=caret (accessed on 1 March 2022).
- Liaw, A.; Wiener, M. Classification and Regression by random Forest. R News 2002, 2, 18–22. [Google Scholar]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016. [Google Scholar] [CrossRef] [Green Version]
- Arjasakusuma, S.; Swahyu Kusuma, S.; Phinn, S. Evaluating Variable Selection and Machine Learning Algorithms for Estimating Forest Heights by Combining Lidar and Hyperspectral Data. ISPRS Int. J. Geoinf. 2020, 9, 507. [Google Scholar] [CrossRef]
- Kacic, P.; Hirner, A.; Da Ponte, E. Fusing Sentinel-1 and -2 to Model GEDI-Derived Vegetation Structure Characteristics in GEE for the Paraguayan Chaco. Remote Sens. 2021, 13, 5105. [Google Scholar] [CrossRef]
- Anchang, J.Y.; Prihodko, L.; Ji, W.; Kumar, S.S.; Ross, C.W.; Yu, Q.; Lind, B.; Sarr, M.A.; Diouf, A.A.; Hanan, N.P. Toward Operational Mapping of Woody Canopy Cover in Tropical Savannas Using Google Earth Engine. Front. Environ. Sci. 2020, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Molnar, C. Interpretable Machine Learning. Github.io. Available online: https://christophm.github.io/interpretable-ml-book/ (accessed on 31 May 2022).
- Ross, C.W.; Hanan, N.P.; Prihodko, L.; Anchang, J.; Ji, W.; Yu, Q. Woody-Biomass Projections and Drivers of Change in Sub-Saharan Africa. Nat. Clim. Chang. 2021, 11, 449–455. [Google Scholar] [CrossRef]
- Francini, S.; D’Amico, G.; Vangi, E.; Borghi, C.; Chirici, G. Integrating GEDI and Landsat: Spaceborne Lidar and Four Decades of Optical Imagery for the Analysis of Forest Disturbances and Biomass Changes in Italy. Sensors 2022, 22, 2015. [Google Scholar] [CrossRef]
- Adrah, E.; Mohd Jaafar, W.S.W.; Bajaj, S.; Omar, H.; Leite, R.V.; Silva, C.A.; Cardil, A.; Mohan, M. Analyzing Canopy Height Variations in Secondary Tropical Forests of Malaysia Using NASA GEDI. IOP Conf. Ser. Earth Environ. Sci. 2021, 880, 012031. [Google Scholar] [CrossRef]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Silva, C.E.; et al. Mapping Global Forest Canopy Height through Integration of GEDI and Landsat Data. Remote Sens. Environ. 2021, 253, 112165. [Google Scholar] [CrossRef]
- Dorado-Roda, I.; Pascual, A.; Godinho, S.; Silva, C.; Botequim, B.; Rodríguez-Gonzálvez, P.; González-Ferreiro, E.; Guerra-Hernández, J. Assessing the Accuracy of GEDI Data for Canopy Height and Aboveground Biomass Estimates in Mediterranean Forests. Remote Sens. 2021, 13, 2279. [Google Scholar] [CrossRef]
- Adam, M.; Urbazaev, M.; Dubois, C.; Schmullius, C. Accuracy Assessment of GEDI Terrain Elevation and Canopy Height Estimates in European Temperate Forests: Influence of Environmental and Acquisition Parameters. Remote Sens. 2020, 12, 3948. [Google Scholar] [CrossRef]
- Hilbert, C.; Schmullius, C. Influence of Surface Topography on ICESat/GLAS Forest Height Estimation and Waveform Shape. Remote Sens. 2012, 4, 2210–2235. [Google Scholar] [CrossRef] [Green Version]
- Lang, N.; Kalischek, N.; Armston, J.; Schindler, K.; Dubayah, R.; Wegner, J.D. Global Canopy Height Regression and Uncertainty Estimation from GEDI LIDAR Waveforms with Deep Ensembles. Remote Sens. Environ. 2022, 268, 112760. [Google Scholar] [CrossRef]
- Yu, Q.; Ji, W.; Prihodko, L.; Ross, C.W.; Anchang, J.Y.; Hanan, N.P. Study Becomes Insight: Ecological Learning from Machine Learning. Methods Ecol. Evol. 2021, 12, 2117–2128. [Google Scholar] [CrossRef]
- Clark, D.B.; Hurtado, J.; Saatchi, S.S. Tropical Rain Forest Structure, Tree Growth and Dynamics along a 2700-m Elevational Transect in Costa Rica. PLoS ONE 2015, 10, e0122905. [Google Scholar] [CrossRef]
- Malhi, Y.; Silman, M.; Salinas, N.; Bush, M.; Meir, P.; Saatchi, S. Introduction: Elevation Gradients in the Tropics: Laboratories for Ecosystem Ecology and Global Change Research. Glob. Change Biol. 2010, 16, 3171–3175. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Givnish, T.J. Tree Diversity in Relation to Tree Height: Alternative Perspectives. Ecol. Lett. 2017, 20, 395–397. [Google Scholar] [CrossRef]
- Blom, C.W.; Voesenek, L.A. Flooding: The Survival Strategies of Plants. Trends Ecol. Evol. 1996, 11, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Schuur, E.A.G. Productivity and Global Climate Revisited: The Sensitivity of Tropical Forest Growth to Precipitation. Ecology 2003, 84, 1165–1170. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud Cover Limits Net CO2 Uptake and Growth of a Rainforest Tree during Tropical Rainy Seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [Green Version]
- Ameztegui, A.; Rodrigues, M.; Gelabert, P.J.; Lavaquiol, B.; Coll, L. Maximum Height of Mountain Forests Abruptly Decreases above an Elevation Breakpoint. GIsci Remote Sens. 2021, 58, 442–454. [Google Scholar] [CrossRef]
- Körner, C. The Use of “altitude” in Ecological Research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range Dynamics of Mountain Plants Decrease with Elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848–1853. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.; Spehn, E. A Humboldtian View of Mountains. Science 2019, 365, 1061. [Google Scholar] [CrossRef] [Green Version]
- Hofhansl, F.; Wanek, W.; Drage, S.; Huber, W.; Weissenhofer, A.; Richter, A. Topography Strongly Affects Atmospheric Deposition and Canopy Exchange Processes in Different Types of Wet Lowland Rainforest, Southwest Costa Rica. Biogeochemistry 2011, 106, 371–396. [Google Scholar] [CrossRef]
- Spracklen, D.V.; Righelato, R. Tropical Montane Forests Are a Larger than Expected Global Carbon Store. Biogeosciences 2014, 11, 2741–2754. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Kolyaie, S.; Morton, D.C.; Zimmerman, J.K.; Uriarte, M. Effects of topography on tropical forest structure depend on climate context. J. Ecol. 2020, 108, 145–159. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | Resolution | Reference Studies | Source of Used Dataset |
|---|---|---|---|
| Annual mean temperature (AMT) * Mean temperature of wettest quarter (MTWQ) Mean temperature of driest quarter (MTDQ) Annual mean precipitation (AP) Precipitation of the wettest month (PWM) | 1 × 1 km | [9,12,19,38] | World Climate |
| Precipitation minus potential evapotranspiration (P-PET) * | 1 × 1 km | [12,13,19] | CIGAR |
| Elevation * Slope * Aspect * Mean curvature * Gaussian curvature Vertical curvature Horizontal curvature Max/Min curvature | 1 × 1 km | [14,15] | SRTM |
| Forest polygons | Vector | - | FRIM |
| Grid Resolution | 25 m | 30, 90, 250, 1000 m |
|---|---|---|
| Considered GEDI metrics | rh50, rh75, rh90, rh95, rh100 | rh50, rh75, rh90, rh95, rh100 |
| GEDI Gridding | No aggregation; comparison at footprint level | Max aggregation (rh-Max) 90th percentile (rh-90) Mean (rh-Mean) |
| ALS Gridding | Max aggregation (HALS-Max) 95th percentile (HALS-95) 90th percentile (HALS-90) Mean (HALS-Mean) | |
| Comparison statistics | R2 Mean absolute error (MAE) Root mean square error (RMSE) relative RMSE (rRMSE) | |
| Study Sites | Peninsular Part | Borneo Part | |||
|---|---|---|---|---|---|
| FRIM | Negeri Sembilan | Danum | SAFE 1 | SAFE 2 | |
| Area (km2) | 12.9 | 6.6 | 75.5 | 196.3 | 200.3 |
| GEDI shots (n) | 405 | 73 | 1807 | 3620 | 5128 |
| Average GEDI shot per 1 km2 | 21.5 | 10.6 | 22.5 | 26.5 | 27.3 |
| rh90 GEDI average height (m) | 25.7 | 30 | 41.6 | 23.7 | 23 |
| ALS average height (m) | 21.5 | 24.5 | 33 | 14.6 | 18.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adrah, E.; Wan Mohd Jaafar, W.S.; Omar, H.; Bajaj, S.; Leite, R.V.; Mazlan, S.M.; Silva, C.A.; Chel Gee Ooi, M.; Mohd Said, M.N.; Abdul Maulud, K.N.; et al. Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR. Remote Sens. 2022, 14, 3172. https://doi.org/10.3390/rs14133172
Adrah E, Wan Mohd Jaafar WS, Omar H, Bajaj S, Leite RV, Mazlan SM, Silva CA, Chel Gee Ooi M, Mohd Said MN, Abdul Maulud KN, et al. Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR. Remote Sensing. 2022; 14(13):3172. https://doi.org/10.3390/rs14133172
Chicago/Turabian StyleAdrah, Esmaeel, Wan Shafrina Wan Mohd Jaafar, Hamdan Omar, Shaurya Bajaj, Rodrigo Vieira Leite, Siti Munirah Mazlan, Carlos Alberto Silva, Maggie Chel Gee Ooi, Mohd Nizam Mohd Said, Khairul Nizam Abdul Maulud, and et al. 2022. "Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR" Remote Sensing 14, no. 13: 3172. https://doi.org/10.3390/rs14133172
APA StyleAdrah, E., Wan Mohd Jaafar, W. S., Omar, H., Bajaj, S., Leite, R. V., Mazlan, S. M., Silva, C. A., Chel Gee Ooi, M., Mohd Said, M. N., Abdul Maulud, K. N., Cardil, A., & Mohan, M. (2022). Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR. Remote Sensing, 14(13), 3172. https://doi.org/10.3390/rs14133172