
Leveraging Machine Learning and Geo-Tagged Citizen Science Data to Disentangle the Factors of Avian Mortality Events at the Species Level


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
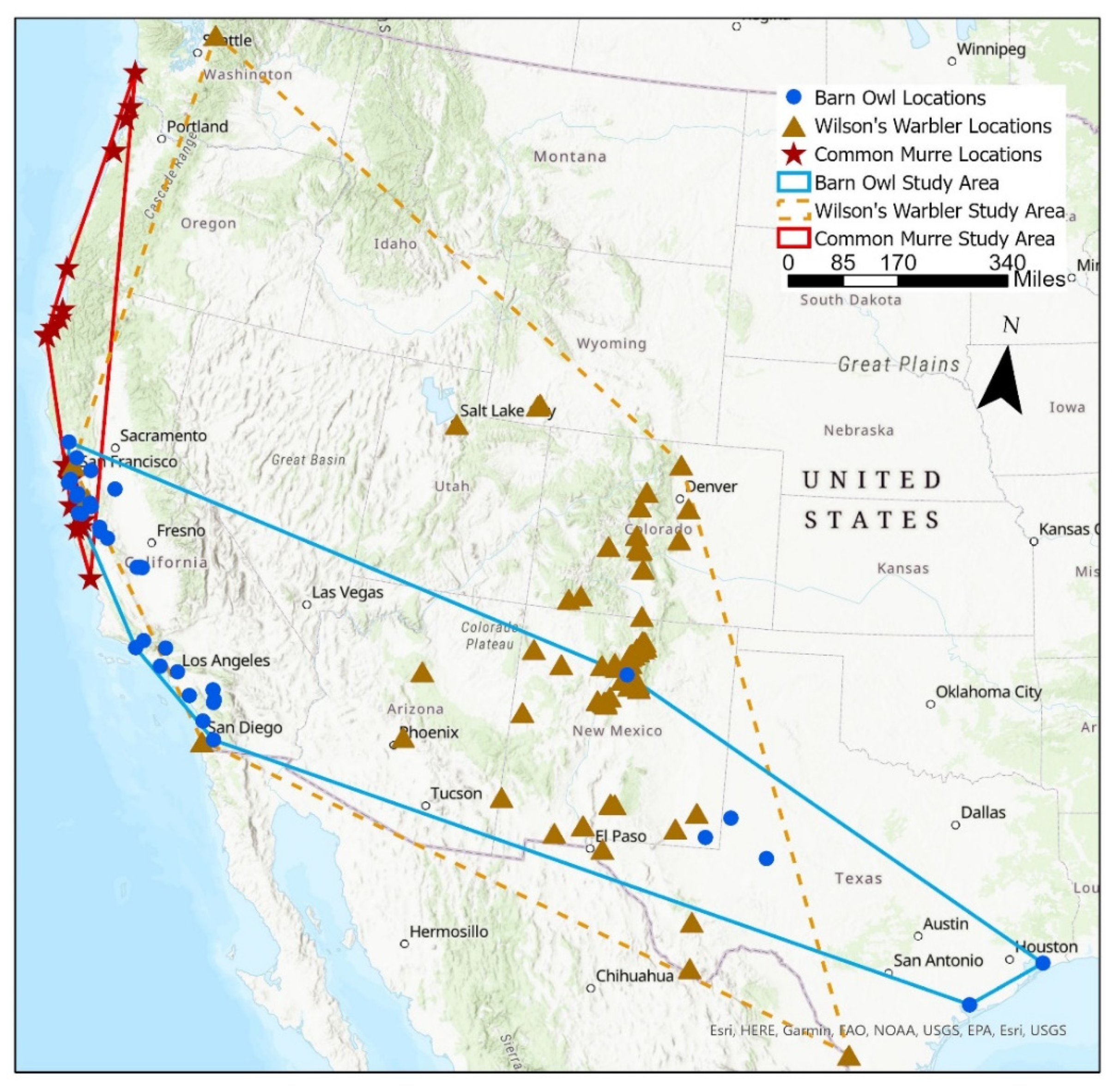
2.1. Citizen Science Avian Mortality Data and Study Area
2.2. Earth Observation Data
2.3. Ensemble Random Forest Model
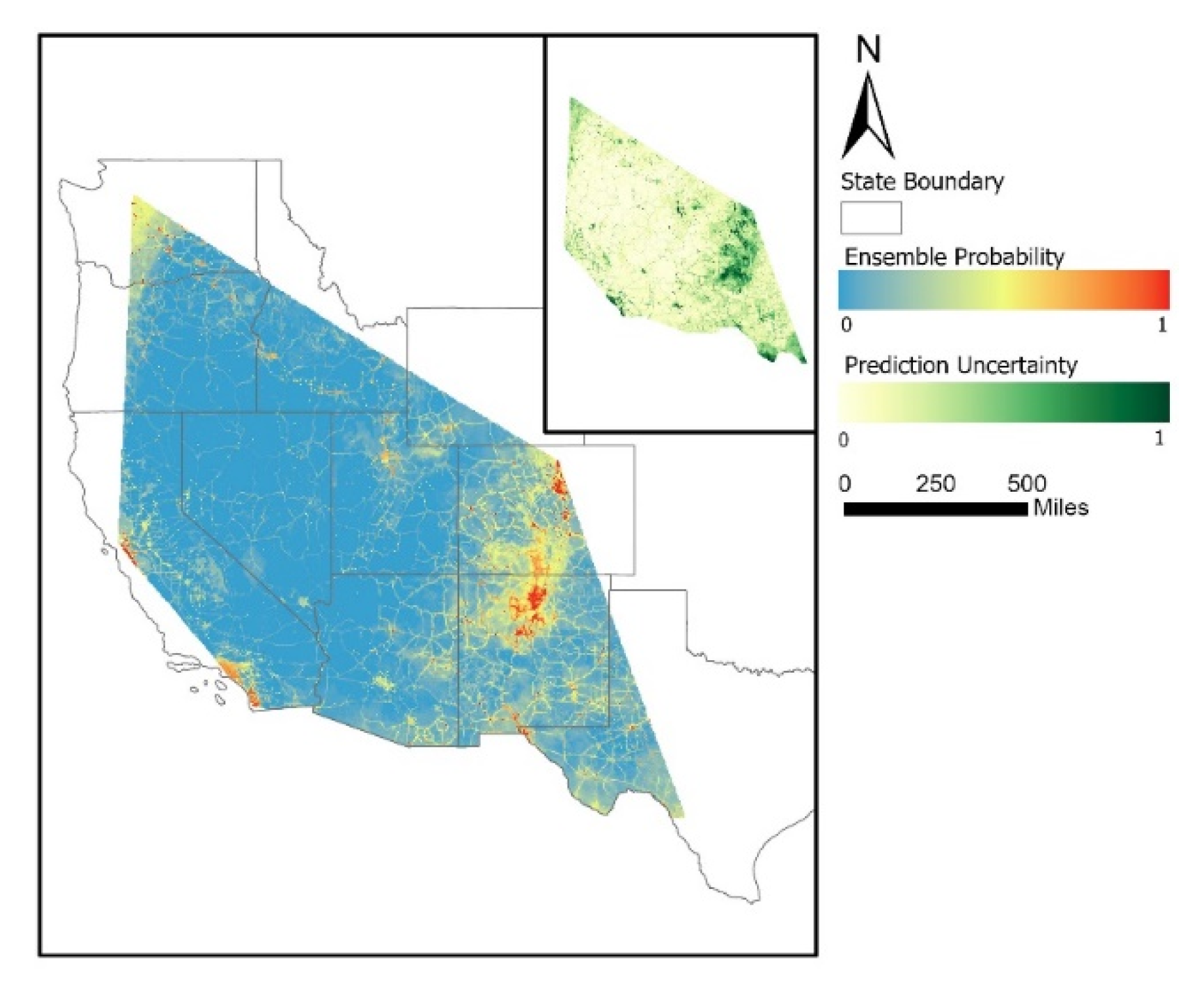
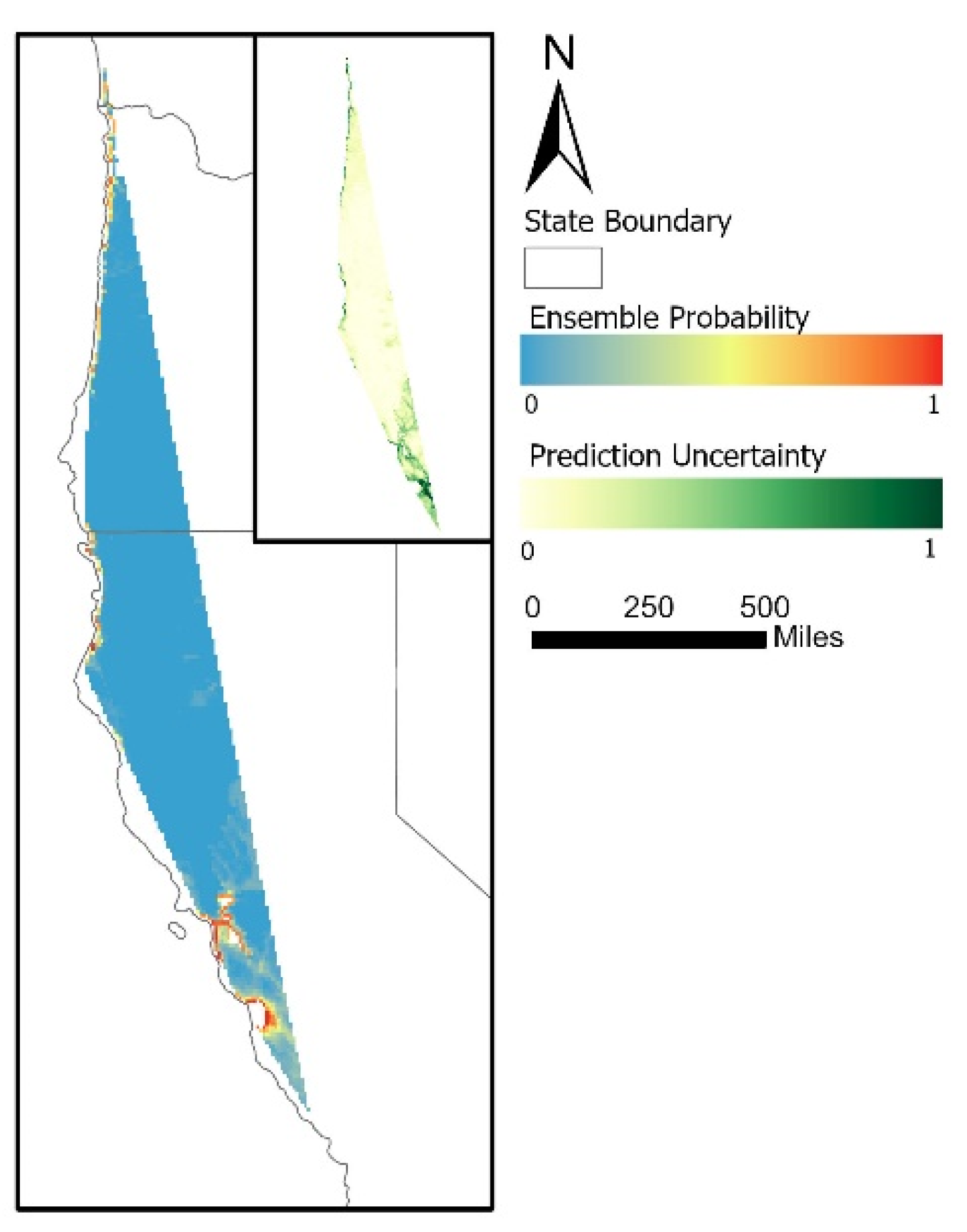
3. Results
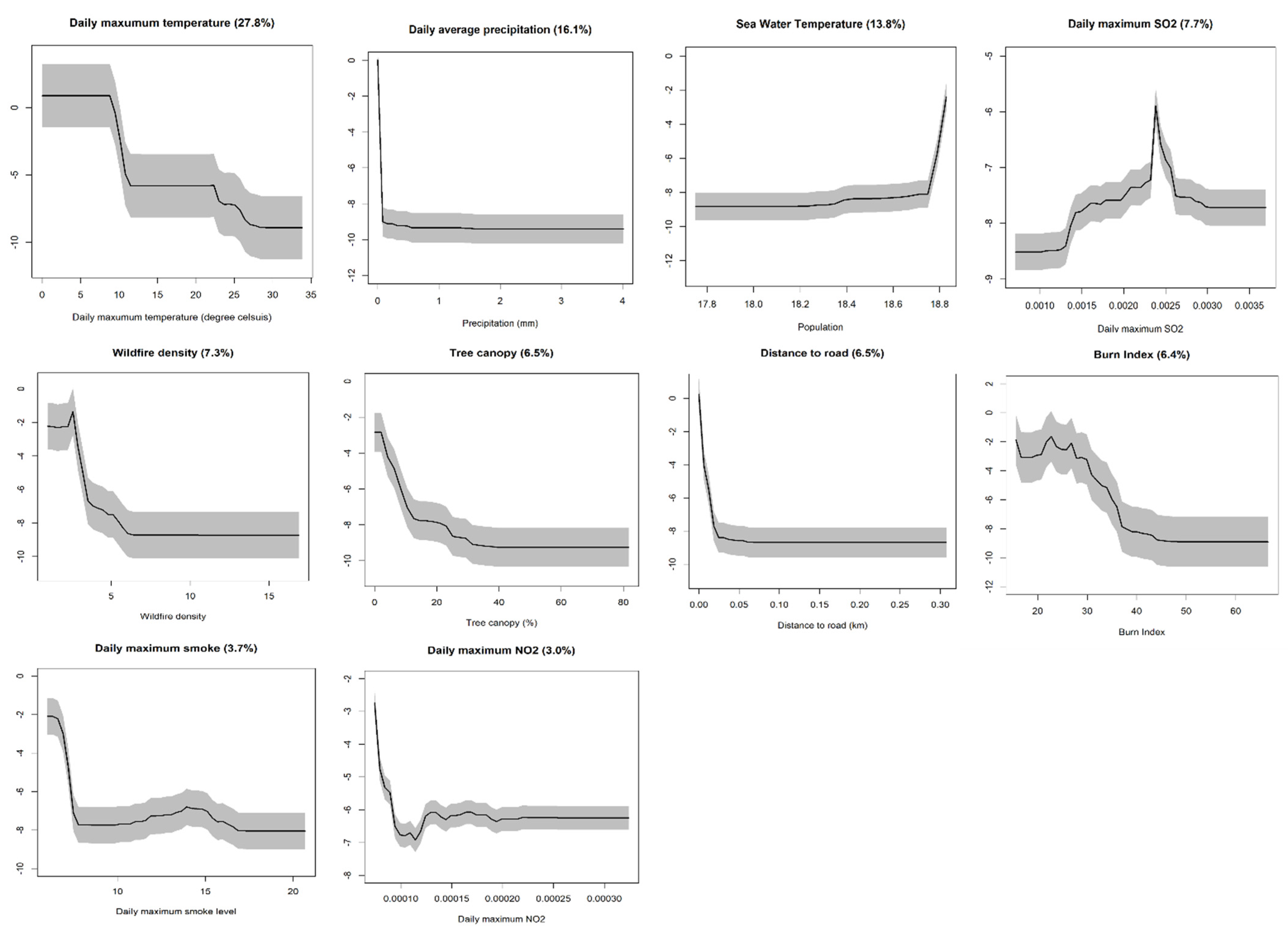
3.1. Wilson’s Warbler’s Mortality
3.2. Barn Owl’s Mortality
3.3. Common Murre’s Mortality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological Responses to Recent Climate Change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.-R. Community and Ecosystem Responses to Recent Climate Change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.; Flint, L.; Syphard, A.D.; Moritz, M.A.; Buckley, L.B.; McCullough, I.M. Fine-Grain Modeling of Species’ Response to Climate Change: Holdouts, Stepping-Stones, and Microrefugia. Trends Ecol. Evol. 2014, 29, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.G.; Lee-Yaw, J.A.; Sunday, J.M.; Hargreaves, A.L. Expanding, Shifting and Shrinking: The Impact of Global Warming on Species’ Elevational Distributions. Glob. Ecol. Biogeogr. 2018, 27, 1268–1276. [Google Scholar] [CrossRef]
- Caula, S.; Marty, P.; Martin, J.-L. Seasonal Variation in Species Composition of an Urban Bird Community in Mediterranean France. Landsc. Urban Plan. 2008, 87, 1–9. [Google Scholar] [CrossRef]
- Jones, K.R.; Watson, J.E.; Possingham, H.P.; Klein, C.J. Incorporating Climate Change into Spatial Conservation Prioritisation: A Review. Biol. Conserv. 2016, 194, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Seabrook, L.; McAlpine, C.; Baxter, G.; Rhodes, J.; Bradley, A.; Lunney, D. Drought-Driven Change in Wildlife Distribution and Numbers: A Case Study of Koalas in South West Queensland. Wildl. Res. 2011, 38, 509–524. [Google Scholar] [CrossRef]
- Lam, S.S.; Waugh, C.; Peng, W.; Sonne, C. Wildfire Puts Koalas at Risk of Extinction. Science 2020, 367, 750. [Google Scholar] [CrossRef]
- Boyle, W.A.; Norris, D.R.; Guglielmo, C.G. Storms Drive Altitudinal Migration in a Tropical Bird. Proc. R. Soc. B Biol. Sci. 2010, 277, 2511–2519. [Google Scholar] [CrossRef] [Green Version]
- Waide, R.B. Summary of the Response of Animal Populations to Hurricanes in the Caribbean. Biotropica 1991, 23, 508–512. [Google Scholar] [CrossRef]
- Johnson, K. The Southwest Is Facing an ‘Unprecedented’ Migratory Bird Die-Off. Audubon Mag. 2020. Available online: https://www.audubon.org/news/the-southwest-facing-unprecedented-migratory-bird-die (accessed on 21 September 2021).
- Insurance Information Institute Facts + Statistics: Wildfires 2020. Available online: https://www.iii.org/fact-statistic/facts-statistics-wildfires (accessed on 21 September 2021).
- New Mexico Department of Game & Fish Starvation. Unexpected Weather to Blame in Mass Migratory Songbird Mortality. 2020. Available online: https://www.wildlife.state.nm.us/starvation-unexpected-weather-to-blame-in-mass-migratory-songbird-mortality/ (accessed on 21 September 2021).
- Yang, D.; Yang, A.; Yang, J.; Xu, R.; Qiu, H. Unprecedented Migratory Bird Die-off: A Citizen-based Analysis on the Spatiotemporal Patterns of Mass Mortality Events in the Western United States. GeoHealth 2021, 5, e2021GH000395. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sánchez, A.; Renton, K.; Landgrave-Ramírez, R.; Mora-Aguilar, E.F.; Rojas-Soto, O. Ecological Niche Variation in the Wilson’s Warbler Cardellina Pusilla Complex. J. Avian Biol. 2015, 46, 516–527. [Google Scholar] [CrossRef]
- Curson, J.; Quinn, D.; Beadle, D. Warblers of the Americas: An Identification Guide; Houghton Mifflin Harcourt: Boston, MA, USA, 1994; ISBN 0-395-70998-9. [Google Scholar]
- Dunn, J.L.; Garrett, K. A Field Guide to Warblers of North America; Houghton Mifflin Harcourt: Boston, MA, USA, 1997; Volume 49, ISBN 0-395-78321-6. [Google Scholar]
- Huang, A.C.; Elliott, J.E.; Cheng, K.M.; Ritland, K.; Ritland, C.E.; Thomsen, S.K.; Hindmarch, S.; Martin, K. Barn Owls (Tyto Alba) in Western North America: Phylogeographic Structure, Connectivity, and Genetic Diversity. Conserv. Genet. 2016, 17, 357–367. [Google Scholar] [CrossRef]
- Briggs, K.T.; Tyler, W.B.; Lewis, D.B.; Carlson, D.R. Bird Communities at Sea off California: 1975 to 1983; Cooper Ornithological Society Inc.: Lawrence, KS, USA, 1987; ISBN 0-935868-36-4. [Google Scholar]
- Roletto, J.; Mortenson, J.; Harrald, I.; Hall, J.; Grella, L. Beached Bird Surveys and Chronic Oil Pollution in Central California. Mar. Ornithol. 2003, 31, 21–28. [Google Scholar]
- Gibble, C.; Duerr, R.; Bodenstein, B.; Lindquist, K.; Lindsey, J.; Beck, J.; Henkel, L.; Roletto, J.; Harvey, J.; Kudela, R. Investigation of a Largescale Common Murre (Uria Aalge) Mortality Event in California, USA, in 2015. J. Wildl. Dis. 2018, 54, 569–574. [Google Scholar] [CrossRef]
- Ballatore, A.; Mooney, P. Conceptualising the Geographic World: The Dimensions of Negotiation in Crowdsourced Cartography. Int. J. Geogr. Inf. Sci. 2015, 29, 2310–2327. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Franklin, S.E.; Guo, X.; Cattet, M. Remote Sensing of Ecology, Biodiversity and Conservation: A Review from the Perspective of Remote Sensing Specialists. Sensors 2010, 10, 9647–9667. [Google Scholar] [CrossRef]
- Pettorelli, N.; Safi, K.; Turner, W. Satellite Remote Sensing, Biodiversity Research and Conservation of the Future. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130190. [Google Scholar] [CrossRef]
- Guanter, L.; Bacour, C.; Schneider, A.; Aben, I.; van Kempen, T.A.; Maignan, F.; Retscher, C.; Köhler, P.; Frankenberg, C.; Joiner, J. The TROPOSIF Global Sun-Induced Fluorescence Dataset from the Sentinel-5P TROPOMI Mission. Earth Syst. Sci. Data 2021, 13, 5423–5440. [Google Scholar] [CrossRef]
- Buchhorn, M.; Lesiv, M.; Tsendbazar, N.-E.; Herold, M.; Bertels, L.; Smets, B. Copernicus Global Land Cover Layers—Collection 2. Remote Sens. 2020, 12, 1044. [Google Scholar] [CrossRef] [Green Version]
- Vîrghileanu, M.; Săvulescu, I.; Mihai, B.-A.; Nistor, C.; Dobre, R. Nitrogen Dioxide (NO2) Pollution Monitoring with Sentinel-5P Satellite Imagery over Europe during the Coronavirus Pandemic Outbreak. Remote Sens. 2020, 12, 3575. [Google Scholar] [CrossRef]
- Bello, O.M.; Aina, Y.A. Satellite Remote Sensing as a Tool in Disaster Management and Sustainable Development: Towards a Synergistic Approach. Procedia-Soc. Behav. Sci. 2014, 120, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Kampichler, C.; Wieland, R.; Calmé, S.; Weissenberger, H.; Arriaga-Weiss, S. Classification in Conservation Biology: A Comparison of Five Machine-Learning Methods. Ecol. Inform. 2010, 5, 441–450. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Kracalik, I.T.; Kenu, E.; Ayamdooh, E.N.; Allegye-Cudjoe, E.; Polkuu, P.N.; Frimpong, J.A.; Nyarko, K.M.; Bower, W.A.; Traxler, R.; Blackburn, J.K. Modeling the Environmental Suitability of Anthrax in Ghana and Estimating Populations at Risk: Implications for Vaccination and Control. PLoS Negl. Trop. Dis. 2017, 11, e0005885. [Google Scholar] [CrossRef]
- Burgman, M.A.; Fox, J.C. Bias in Species Range Estimates from Minimum Convex Polygons: Implications for Conservation and Options for Improved Planning; Cambridge University Press: Cambridge, UK, 2003; Volume 6, pp. 19–28. [Google Scholar]
- Yang, A.; Yang, J.; Di Yang, R.X.; He, Y.; Aragon, A.; Qiu, H. Human Mobility to Parks Under the COVID-19 Pandemic and Wildfire Seasons in the Western and Central United States. GeoHealth 2021, 5, e2021GH000494. [Google Scholar] [CrossRef]
- Geffen, V.; Boersma, K.F.; Eskes, H.; Sneep, M.; Linden, T.; Zara, M.; Pepijn Veefkind, J. S5p TROPOMI NO2 Slant Column Retrieval: Method, Stability, Uncertainties and Comparisons with OMI. Atmos. Meas. Tech. 2020, 13, 1315–1335. [Google Scholar] [CrossRef] [Green Version]
- Harpconvert. Available online: http://stcorp.github.io/harp/doc/html/harpconvert.html (accessed on 13 February 2021).
- Verhoelst, T.; Compernolle, S.; Pinardi, G.; Lambert, J.-C.; Eskes, H.J.; Eichmann, K.-U.; Fjæraa, A.M.; Granville, J.; Niemeijer, S.; Cede, A. Ground-Based Validation of the Copernicus Sentinel-5p TROPOMI NO 2 Measurements with the NDACC ZSL-DOAS, MAX-DOAS and Pandonia Global Networks. Atmos. Meas. Tech. 2021, 14, 481–510. [Google Scholar] [CrossRef]
- Levelt, P.F.; Stein Zweers, D.C.; Aben, I.; Bauwens, M.; Borsdorff, T.; De Smedt, I.; Eskes, H.J.; Lerot, C.; Loyola, D.G.; Romahn, F. Air Quality Impacts of COVID-19 Lockdown Measures Detected from Space Using High Spatial Resolution Observations of Multiple Trace Gases from Sentinel-5P/TROPOMI. Atmos. Chem. Phys. Discuss. 2021, 1–53. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Buren, A.D.; Koen-Alonso, M.; Montevecchi, W.A. Linking Predator Diet and Prey Availability: Common Murres and Capelin in the Northwest Atlantic. Mar. Ecol. Prog. Ser. 2012, 445, 25–35. [Google Scholar] [CrossRef]
- McFarlane Tranquilla, L.A.; Montevecchi, W.A.; Fifield, D.A.; Hedd, A.; Gaston, A.J.; Robertson, G.J.; Phillips, R.A. Individual Winter Movement Strategies in Two Species of Murre (Uria Spp.) in the Northwest Atlantic. PLoS ONE 2014, 9, e90583. [Google Scholar] [CrossRef] [PubMed]
- McFarlane Tranquilla, L.; Montevecchi, W.; Hedd, A.; Regular, P.; Robertson, G.; Fifield, D.; Devillers, R. Ecological Segregation among Thick-Billed Murres (Uria Lomvia) and Common Murres (Uria Aalge) in the Northwest Atlantic Persists through the Nonbreeding Season. Can. J. Zool. 2015, 93, 447–460. [Google Scholar] [CrossRef]
- Cummings, J.A.; Smedstad, O.M. Variational Data Assimilation for the Global Ocean. In Data Assimilation for Atmospheric, Oceanic and Hydrologic Applications; Springer: Berlin/Heidelberg, Germany, 2013; Volume II, pp. 303–343. [Google Scholar]
- Yang, A.; Gomez, J.P.; Blackburn, J.K. Exploring Environmental Coverages of Species: A New Variable Contribution Estimation Methodology for Rulesets from the Genetic Algorithm for Rule-Set Prediction. PeerJ 2020, 8, e8968. [Google Scholar] [CrossRef] [PubMed]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-absences for Species Distribution Models: How, Where and How Many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Yang, A.; Proffitt, K.M.; Asher, V.; Ryan, S.J.; Blackburn, J.K. Sex-Specific Elk Resource Selection during the Anthrax Risk Period. J. Wildl. Manag. 2021, 85, 145–155. [Google Scholar] [CrossRef]
- Kulkarni, V.Y.; Sinha, P.K. Pruning of Random Forest Classifiers: A Survey and Future Directions. In Proceedings of the 2012 International Conference on Data Science & Engineering (ICDSE), Cochin, India, 18–20 July 2012; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2012; pp. 64–68. [Google Scholar]
- Ghosh, A.; Sharma, R.; Joshi, P. Random Forest Classification of Urban Landscape Using Landsat Archive and Ancillary Data: Combining Seasonal Maps with Decision Level Fusion. Appl. Geogr. 2014, 48, 31–41. [Google Scholar] [CrossRef]
- Probst, P.; Wright, M.N.; Boulesteix, A. Hyperparameters and Tuning Strategies for Random Forest. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2019, 9, e1301. [Google Scholar] [CrossRef] [Green Version]
- Deribe, K.; Cano, J.; Newport, M.J.; Golding, N.; Pullan, R.L.; Sime, H.; Gebretsadik, A.; Assefa, A.; Kebede, A.; Hailu, A. Mapping and Modelling the Geographical Distribution and Environmental Limits of Podoconiosis in Ethiopia. PLoS Negl. Trop. Dis. 2015, 9, e0003946. [Google Scholar] [CrossRef]
- Louppe, G.; Wehenkel, L.; Sutera, A.; Geurts, P. Understanding Variable Importances in Forests of Randomized Trees. In Proceedings of the Advances in Neural Information Processing Systems, Stateline, NV, USA, 5–10 December 2013. [Google Scholar]
- Marti, C.D.; Wagner, P.W. Winter Mortality in Common Barn-Owls and Its Effect on Population Density and Reproduction. Condor 1985, 87, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Handrich, Y.; Nicolas, L.; Maho, Y.L. Winter Starvation in Captive Common Barn-Owls: Physiological States and Reversible Limits. Auk 1993, 110, 458–469. [Google Scholar] [CrossRef]
- Sanderfoot, O.V.; Holloway, T. Air Pollution Impacts on Avian Species via Inhalation Exposure and Associated Outcomes. Environ. Res. Lett. 2017, 12, 083002. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.E.; Brain, J.D.; Wang, N. The Avian Respiratory System: A Unique Model for Studies of Respiratory Toxicosis and for Monitoring Air Quality. Environ. Health Perspect. 1997, 105, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.; Taylor, C.E.; Martin, J.M. Are Pro-Ecological Values Enough? Determining the Drivers and Extent of Participation in Citizen Science Programs. Hum. Dimens. Wildl. 2019, 24, 501–514. [Google Scholar] [CrossRef]
- Yang, D.; Yang, A.; Qiu, H.; Zhou, Y.; Herrero, H.; Fu, C.-S.; Yu, Q.; Tang, J. A Citizen-Contributed GIS Approach for Evaluating the Impacts of Land Use on Hurricane-Harvey-Induced Flooding in Houston Area. Land 2019, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Ballatore, A.; Jokar Arsanjani, J. Placing Wikimapia: An Exploratory Analysis. Int. J. Geogr. Inf. Sci. 2019, 33, 1633–1650. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Covariates | Descriptions | Sources |
|---|---|---|---|
| Wildfire effects | Distance to wildfire (km) | The distance to the closest wildfires | National Interagency Fire Center |
| Wildfire density (km2) | Density of wildfires | ||
| Maximum smoke | The average of the daily maximum smoke level | NOAA Hazard Mapping System Fire and Smoke Product | |
| Carbon monoxide (CO) (mol/m2) | Average of daily CO concentration | Sentinel-5P TROPOMI near-real-time (NRTI) Level-3 | |
| Sulfur dioxide (SO2) (mol/m2) | Average of daily SO2 concentration | ||
| Nitrogen dioxide (NO2) (mol/m2) | Average of daily NO2 concentration | ||
| Winter storm | Snow cover | Percentage of daily maximum snow cover | MOD10A1 V6 Snow Cover Daily Global 500 m |
| Climatic conditions that might reflect both | Maximum Temperature (Celsius) | Average of daily maximum temperature | gridMET dataset via “ClimateR” package |
| Precipitation (mm) | Average of daily precipitation | ||
| Humidity (kg/kg) | Average of daily humidity | ||
| Burn Index | Average of burn index | ||
| Wind (m/s) | Average of daily wind speed | ||
| Evapotranspiration (mm) | Average of daily evapotranspiration | ||
| Habitat | Forest, shrub, developed, agriculture, grass, and water | Binary indicator for forest, shrub, developed, agriculture, grass, and water land | 2019 National Land Cover Dataset a |
| Ocean temperature (Celsius) | Average of the daily maximum ocean temperature within a buffer of 100 km | Hybrid Coordinate Ocean Model | |
| Tree canopy (%) | Percentage of tree canopy coverage | Multi-Resolution Land Characteristics (MRLC) Consortium | |
| Anthropogenic effects on observation | County population | Population of the county where the observation was reported | Census Bureau |
| Distance to roads (km) | Euclidean distance to primary and secondary roads | TIGER/Line Census Data |
| Species | Model Structure | External Training AUC | External Testing AUC |
|---|---|---|---|
| Wilson’s Warbler | Maximum temperature + Precipitation + Burn index + Distance to roads + Maximum smoke + CO + SO2 + Forest + Wind + Developed + County population + Snow cover + Shrub + Evapotranspiration + Humidity + Wildfire Density | 0.96 (0.96, 0.97) | 0.97 (0.96, 0.98) |
| Barn Owl | Tree canopy + Precipitation + Distance to roads + Maximum smoke + SO2 + NO2 + Forest + Wind + Developed + County population + Snow cover | 0.95 (0.94, 0.95) | 0.93 (0.90, 0.95) |
| Common Murre | Maximum temperature + Tree canopy + Precipitation + Burn index + Ocean temperature + Distance to roads + Wildfire density + Maximum smoke + SO2 + NO2 | 0.98 (0.97, 0.99) | 0.98 (0.97, 0.99) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, A.; Rodriguez, M.; Yang, D.; Yang, J.; Cheng, W.; Cai, C.; Qiu, H. Leveraging Machine Learning and Geo-Tagged Citizen Science Data to Disentangle the Factors of Avian Mortality Events at the Species Level. Remote Sens. 2022, 14, 2369. https://doi.org/10.3390/rs14102369
Yang A, Rodriguez M, Yang D, Yang J, Cheng W, Cai C, Qiu H. Leveraging Machine Learning and Geo-Tagged Citizen Science Data to Disentangle the Factors of Avian Mortality Events at the Species Level. Remote Sensing. 2022; 14(10):2369. https://doi.org/10.3390/rs14102369
Chicago/Turabian StyleYang, Anni, Matthew Rodriguez, Di Yang, Jue Yang, Wenwen Cheng, Changjie Cai, and Han Qiu. 2022. "Leveraging Machine Learning and Geo-Tagged Citizen Science Data to Disentangle the Factors of Avian Mortality Events at the Species Level" Remote Sensing 14, no. 10: 2369. https://doi.org/10.3390/rs14102369
APA StyleYang, A., Rodriguez, M., Yang, D., Yang, J., Cheng, W., Cai, C., & Qiu, H. (2022). Leveraging Machine Learning and Geo-Tagged Citizen Science Data to Disentangle the Factors of Avian Mortality Events at the Species Level. Remote Sensing, 14(10), 2369. https://doi.org/10.3390/rs14102369