
Detecting the Spatial Variability of Seagrass Meadows and Their Consequences on Associated Macrofauna Benthic Activity Using Novel Drone Technology



Abstract
:1. Introduction
2. Materials and Methods
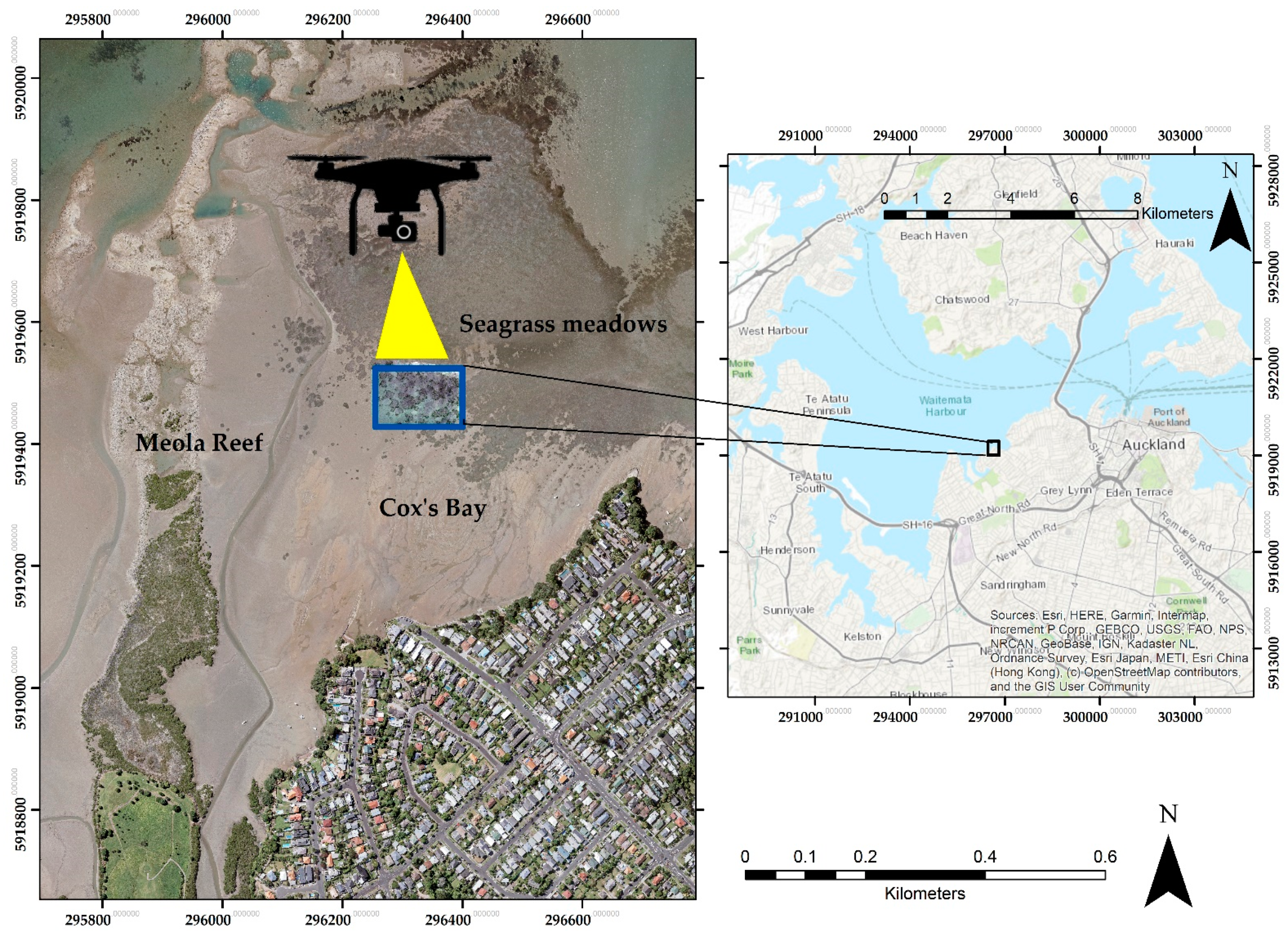
2.1. Study Site
2.2. Data Collection
2.3. On-Site Survey and Classification Schema
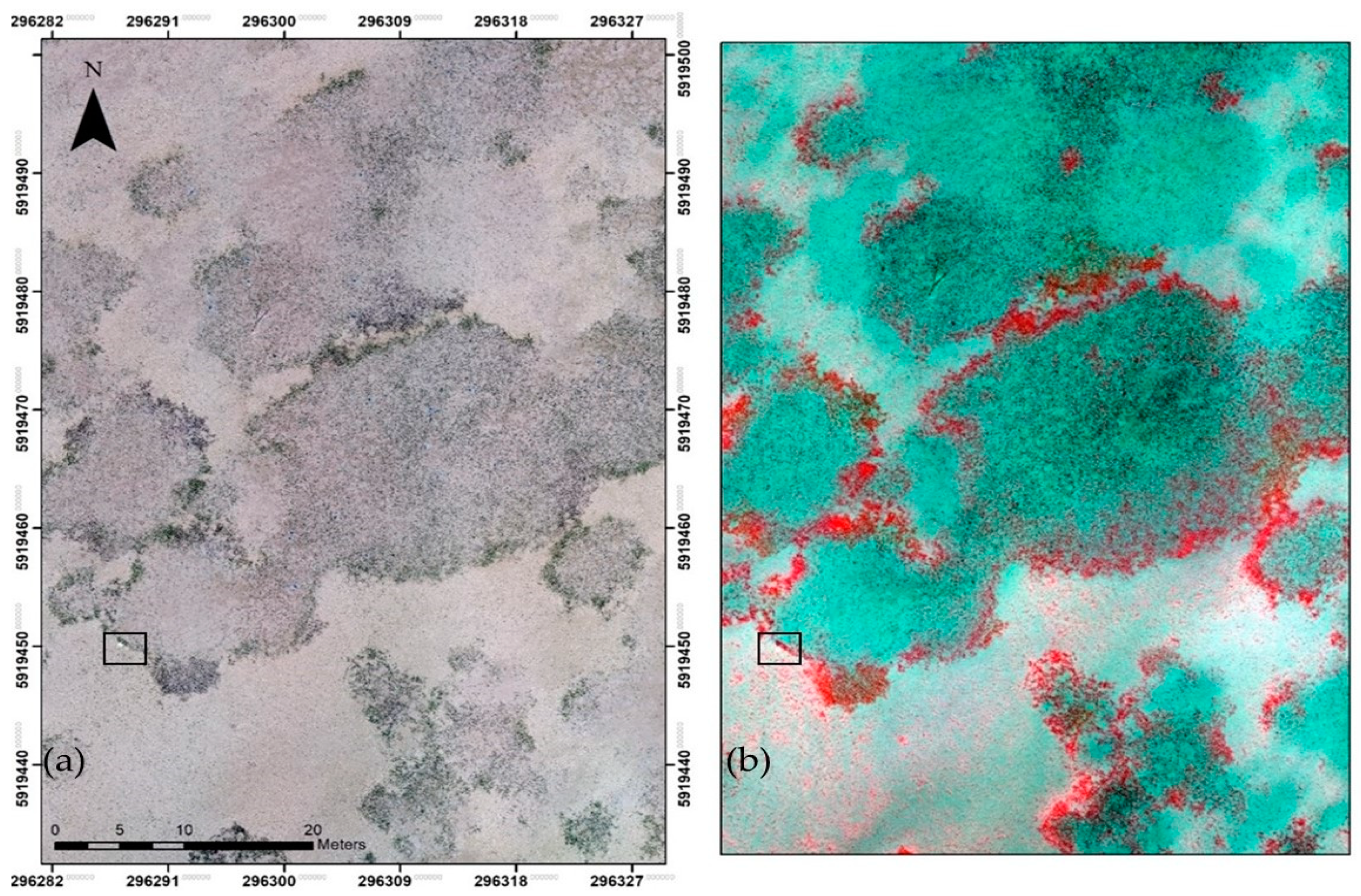
2.4. Identification of Macrofauna Benthic Activity from Their Feeding Burrows
2.5. Vegetation Indices for Supervised Classification
2.6. Support Vector Machine Classification
2.7. Accuracy Assessment
2.8. Habitat Maps: Post Classification Detecting Seasonal Change
3. Results
3.1. VIS Classification
VIS+NIR Classification
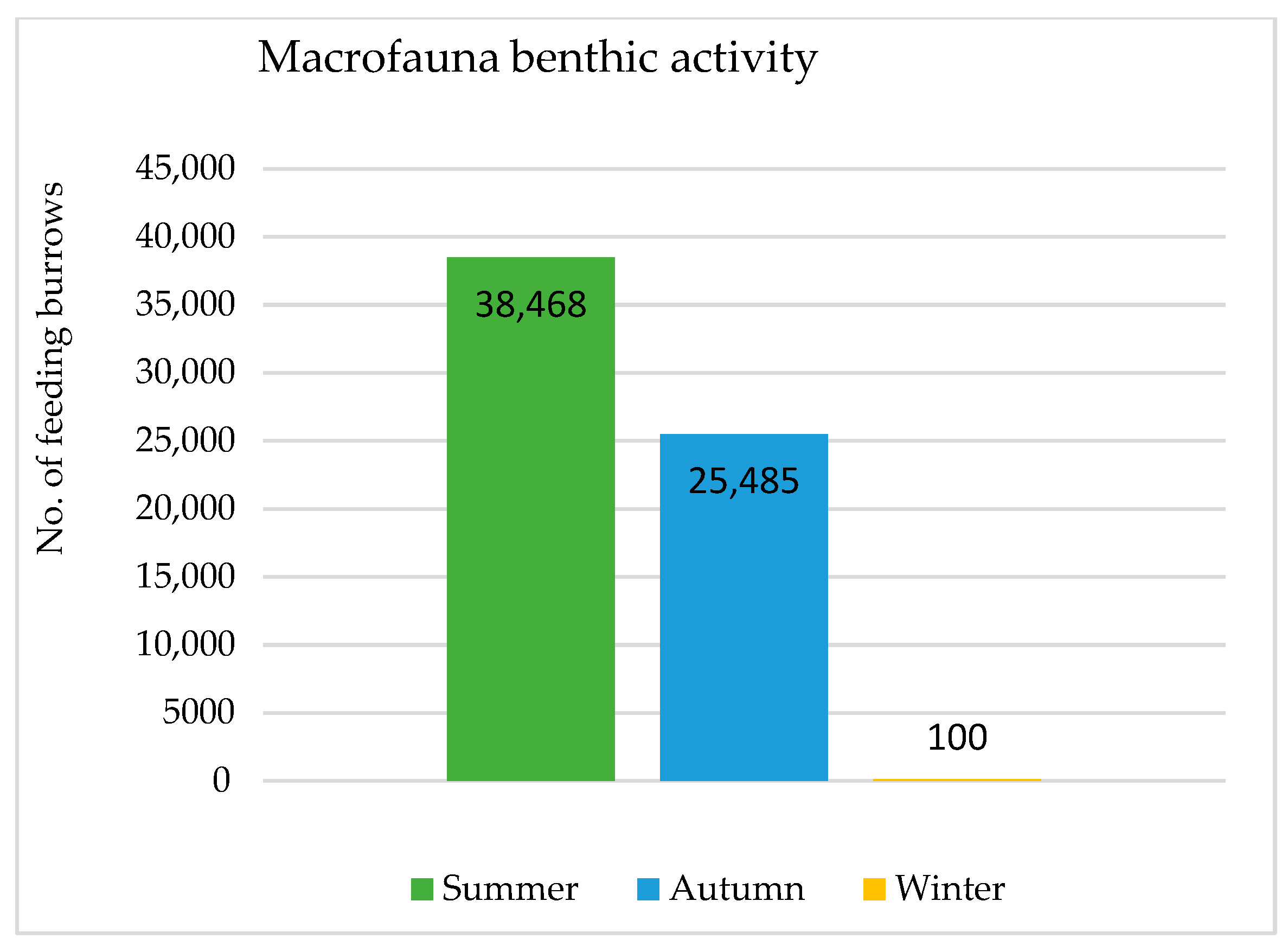
3.2. Identification of Macrofauna Benthic Activity from Their Feeding Burrows
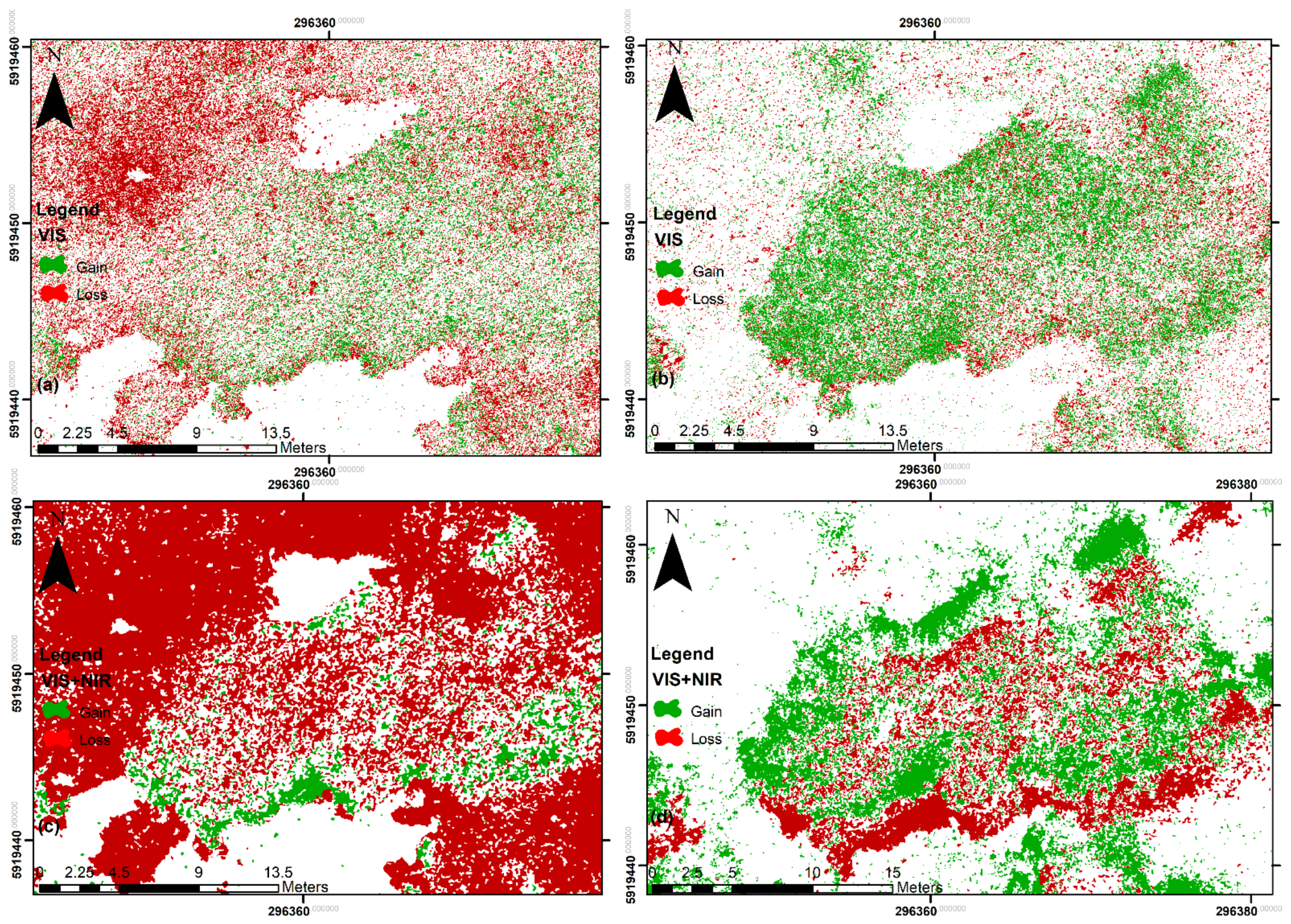
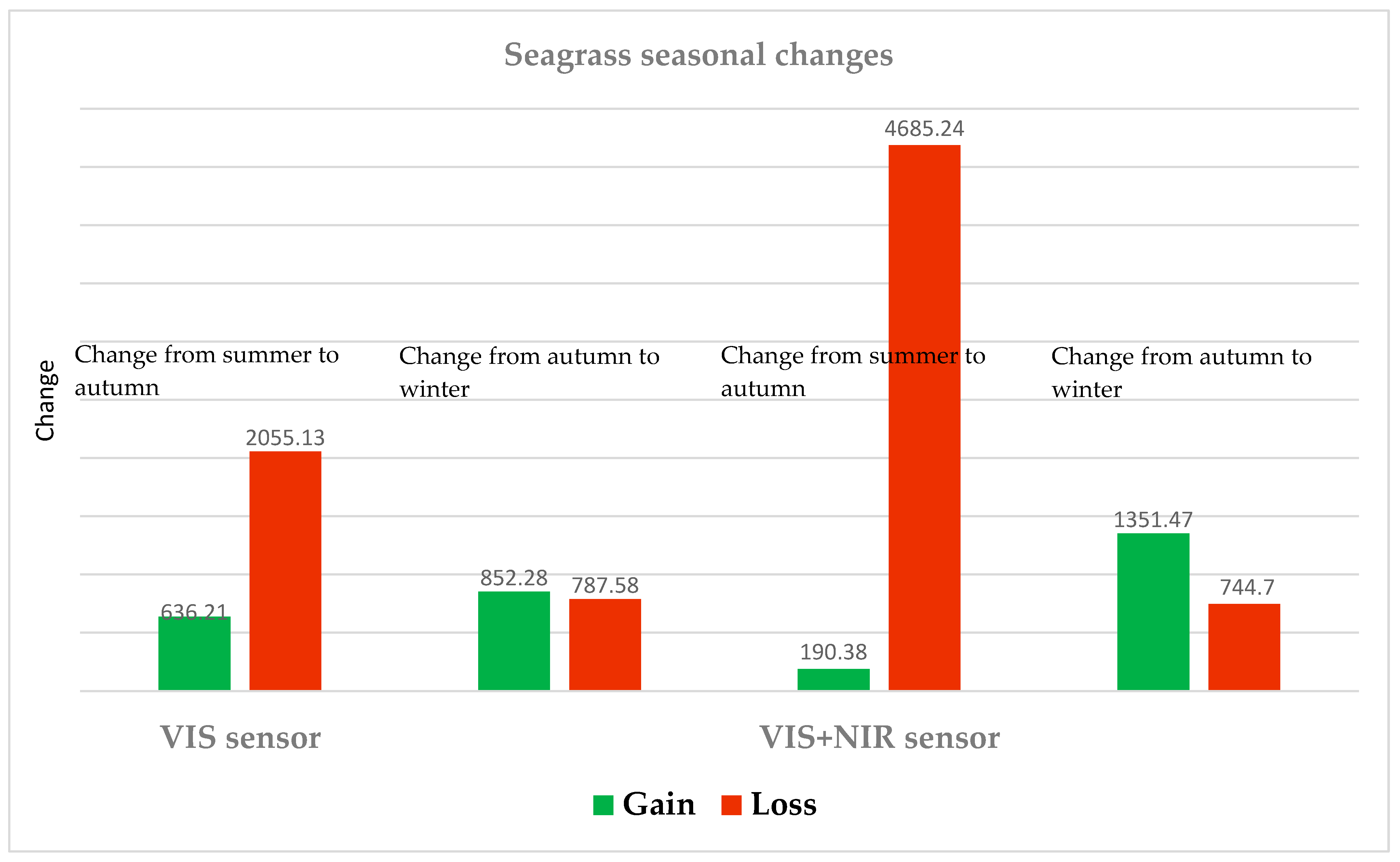
3.3. Habitat Maps: Post Classification Seasonal Change Detection
4. Discussion
4.1. The Performance of the Sensors and the SVM Classifier
VIS+NIR Classification Analysis
4.2. Identification of Macrofauna Benthic Activity Feeding Burrows
4.3. Habitat Maps: Post Classification Seasonal Change Detection
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anderson, T.J.; Morrison, M.; Macdiarmid, A.B.; Clark, M.R.; Archino, R.D.; Tracey, D.M.; Wadhwa, S. Review of New Zealand’s Key Biogenic Habitats; Report; The National Institute of Water and Atmospheric Research (NIWA): Auckland, New Zealand, 2019; p. 190. Available online: https://environment.govt.nz/assets/Publications/Files/NZ-biogenic-habitat-review.pdf (accessed on 15 June 2021).
- Matheson, F.; Wadhwa, S. Seagrass in Porirua Harbour: Preliminary Assessment of Restoration Potential; NIWA Report for Greater Wellington Regional Council; Unpublished Report; Greater Wellington Regional Council: Masterton, New Zealand, 2012; 35p.
- Calleja, F.; Galván, C.; Silio-Calzada, A.; Juanes, J.A.; Ondiviela, B. Long-term analysis of Zostera noltei: A retrospective approach for understanding seagrasses’ dynamics. Mar. Environ. Res. 2017, 130, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Losada, I.; Hendriks, I.E.; Mazarrasa, I.; Marbà, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.; Schwarz, A.M. Management and conservation of seagrass in New Zealand: An introduction. Sci. Conserv. 2006, 264, 1–90. [Google Scholar]
- Attrill, M.J.; Strong, J.A.; Rowden, A.A. Are macroinvertebrate communities influenced by seagrass structural complexity? Ecography 2000, 23, 114–121. [Google Scholar] [CrossRef]
- Grech, A.; Chartrand, K.; Erftemeijer, P.; Fonseca, M.; McKenzie, L.; Rasheed, M.; Taylor, H.; Coles, R. A comparison of threats, vulnerabilities and management approaches in global seagrass bioregions. Environ. Res. Lett. 2012, 7, 024006. [Google Scholar] [CrossRef]
- Lin, H.; Sun, T.; Zhou, Y.; Gu, R.; Zhang, X.; Yang, W. Which genes in a typical intertidal Seagrass (Zostera japonica) indicate pollution? Front. Plant Sci. 2018, 9, 1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orth, R.J.; Carruthers, T.J.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A Global Crisis for Seagrass Ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.A.; Lowe, M.L.; Grant, C.M.; Smith, P.J.; Carbines, G.; Reed, J.; Bury, S.J.; Brown, J. Seagrass Meadows as Species and Productivity Hot Burrows New Zealand Aquatic Environment and Species Report No 137, Volume 6480, Issue 137. 2014. Available online: http://fs.fish.govt.nz (accessed on 15 June 2021).
- Fourqurean, J.W.; Duarte, C.M.; Kennedy, H.; Marbà, N.; Holmer, M.; Mateo, M.A.; Apostolaki, E.T.; Kendrick, G.A.; Krause-Jensen, D.; McGlathery, K.J.; et al. Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 2012, 5, 505–509. [Google Scholar] [CrossRef]
- Matheson, F.E.; Reed, J.; Dos Santos, V.M.; Mackay, G.; Cummings, V. Seagrass rehabilitation: Successful transplants and evaluation of methods at different spatial scales. N. Z. J. Mar. Freshw. Res. 2017, 51, 96–109. [Google Scholar] [CrossRef]
- Chand, S.; Bollard, B. Low altitude spatial assessment and monitoring of intertidal seagrass meadows beyond the visible spectrum using a remotely piloted aircraft system. Estuar. Coast. Shelf Sci. 2021, 255, 107299. [Google Scholar] [CrossRef]
- Inglis, G.J. The seagrasses of New Zealand. In World Atlas of Seagrasses; Green, E.P., Short, F.T., Eds.; University of California Press: Berkeley, CA, USA, 2003; pp. 148–157. [Google Scholar]
- Kettles, H.; Bell, R. Estuarine ecosystems. In Freshwater Conservation under a Changing Climate, Proceedings of the Workshop Hosted by the Department of Conservation, Wellington, New Zealand, 10–11 December 2013; Robertson, H., Bowie, S., White, R., Death, R., Collins, D., Eds.; Department of Conservation: Christchurch, New Zealand, 2016; pp. 24–30. [Google Scholar]
- de Lange, P.J.; Rolfe, J.R.; Barkla, J.W.; Courtney, S.P.; Champion, P.D.; Perrie, L.R.; Beadel, S.M.; Ford, K.A.; Breitwieser, I.; Schönberger, I.; et al. Conservation status of New Zealand indigenous vascular plants, 2017. New Zealand Threat Classification Series 2017, 22. 82p. Available online: https://www.doc.govt.nz/documents/science-and-technical/nztcs22entire.pdf (accessed on 15 June 2021).
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, N.-T.; Manley-Harris, M.; Pham, T.-D.; Hawes, I. Detecting Multi-Decadal Changes in Seagrass Cover in Tauranga Harbour, New Zealand, Using Landsat Imagery and Boosting Ensemble Classification Techniques. ISPRS Int. J. Geo-Inf. 2021, 10, 371. [Google Scholar] [CrossRef]
- Hobley, B.; Arosio, R.; French, G.; Bremner, J.; Dolphin, T.; Mackiewicz, M. Semi-Supervised Segmentation for Coastal Monitoring Seagrass Using RPA Imagery. Remote Sens. 2021, 13, 1741. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; McKenzie, L.J.; Collier, C.J.; Cullen-Unsworth, L.C.; Duarte, C.M.; Eklöf, J.S.; Jarvis, J.C.; Jones, B.L.; Nordlund, L.M. Global challenges for seagrass conservation. AMBIO 2018, 48, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Johnston, D.W. Unoccupied Aircraft Systems in Marine Science and Conservation. Annu. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.; Gaston, K.J. Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Front. Ecol. Environ. 2013, 11, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Bujang, J.; Zakaria, M.; Hashim, M. Application of Landsat images to seagrass areal cover change analysis for Lawas, Terengganu and Kelantan of Malaysia. Cont. Shelf Res. 2015, 110, 124–148. [Google Scholar] [CrossRef]
- Duffy, J.; Pratt, L.; Anderson, K.; Land, P.; Shutler, J.D. Spatial assessment of intertidal seagrass meadows using optical imaging systems and a lightweight drone. Estuar. Coast. Shelf Sci. 2018, 200, 169–180. [Google Scholar] [CrossRef]
- Joyce, K.E.; Duce, S.; Leahy, S.M.; Leon, J.; Maier, S.W. Principles and practice of acquiring drone-based image data in marine environments. Mar. Freshw. Res. 2018, 70, 952–963. [Google Scholar] [CrossRef]
- Chayhard, S.; Manthachitra, V.; Naulchawee, K.; Buranapratheprat, A. Multitemporal mapping of seagrass distribution by using integrated remote sensing data in Kung Kraben Bay (KKB), Chanthaburi Province, Thailand. Int. J. Agric. Technol. 2018, 14, 161–170. [Google Scholar]
- Konar, B.; Iken, K. The use of unmanned aerial vehicle imagery in intertidal monitoring. Deep. Sea Res. Part II 2018, 147, 79–86. [Google Scholar] [CrossRef]
- Nahirnick, N.K.; Hunter, P.; Costa, M.; Schroeder, S.; Sharma, T. Benefits and Challenges of UAS Imagery for Eelgrass (Zostera marina) Mapping in Small Estuaries of the Canadian West Coast. J. Coast. Res. 2019, 35, 673–683. [Google Scholar] [CrossRef]
- Riniatsih, I.; Ambariyanto, A.; Yudiati, E.; Redjeki, S.; Hartati, R. Monitoring the seagrass ecosystem using the unmanned aerial vehicle (UAV) in coastal water of Jepara. IOP Conf. Ser. Earth Environ. Sci. 2021, 674, 012075. [Google Scholar] [CrossRef]
- Ventura, D.; Bonifazi, A.; Gravina, M.F.; Belluscio, A.; Ardizzone, G. Mapping and Classification of Ecologically Sensitive Marine Habitats Using Unmanned Aerial Vehicle (UAV) Imagery and Object-Based Image Analysis (OBIA). Remote Sens. 2018, 10, 1331. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Hawthorne, T.L.; Searson, H.; Duffy, E. High-Resolution UAV Mapping for Investigating Eelgrass Beds along the West Coast of North America. In Proceedings of the IGARSS 2020—2020 IEEE International Geoscience and Remote Sensing Symposium, Waikoloa, HI, USA, 26 September–2 October 2020; pp. 6317–6320. [Google Scholar] [CrossRef]
- Martin, R.; Ellis, J.; Brabyn, L.; Campbell, M. Seasonal change-mapping of estuarine intertidal seagrass (Zostera muelleri) using multispectral imagery flown by remotely piloted aircraft (RPA) at Wharekawa Harbour, New Zealand. Estuar. Coast. Shelf Sci. 2020, 246, 107046. [Google Scholar] [CrossRef]
- Rodil, I.F.; Lohrer, A.M.; Attard, K.M.; Hewitt, J.E.; Thrush, S.F.; Norkko, A. Macrofauna communities across a seascape of seagrass meadows: Environmental drivers, species patterns and conservation implications. Biodivers. Conserv. 2021, 30, 3023–3043. [Google Scholar] [CrossRef]
- Lundquist, C.J.; Jones, T.C.; Parkes, S.M.; Bulmer, R.H. Seasonal change in benthic community structure and sediment characteristics after natural recolonization of the seagrass Zostera muelleri. Sci. Rep. 2018, 8, 13250. [Google Scholar] [CrossRef] [PubMed]
- Micheli, F.; Bishop, M.J.; Peterson, C.H.; Rivera, J. Alteration of seagrass species composition and function over two decades. Ecol. Monogr. 2008, 78, 225–244. [Google Scholar] [CrossRef]
- Foley, M.M.; Shears, N.T. Te Tokaroa Meola Reef Intertidal Reef Ecological Monitoring Programme: 2001 to 2017. Auckland Council Technical Report, New Zealand, TR2019/004. 2019. Available online: https://knowledgeauckland.org.nz/media/1061/tr2019-004-tokaroa-meola-reef-intertidal-monitoring-2001-to-2017.pdf (accessed on 15 June 2021).
- Turner, S.J. Growth and productivity of intertidal Zostera capricorni in New Zealand estuaries. N. Z. J. Mar. Freshw. Res. 2007, 41, 77–90. [Google Scholar] [CrossRef]
- Saunders, M.I.; Atkinson, S.; Klein, C.J.; Weber, T.; Possingham, H.P. Increased sediment loads cause non-linear decreases in seagrass suitable habitat extent. PLoS ONE 2017, 12, e0187284. [Google Scholar] [CrossRef] [Green Version]
- LINZ Land Information New Zealand. Available online: https://www.linz.govt.nz/ (accessed on 1 November 2020).
- Pix4D. Available online: www.Pix4D.com (accessed on 30 April 2020).
- Cunliffe, A.M.; Brazier, R.E.; Anderson, K. Ultra-fine grain landscape-scale quantification of dryland vegetation structure with drone-acquired structure-from-motion photogrammetry. Remote Sens. Environ. 2016, 183, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, D. APPENDIX 1: Pohnpeian Clans. In Upon a Stone Altar: A History of the Island of Pohnpei to 1890; University of Hawai’i Press: Honolulu, HI, USA, 1988; p. 211. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indices: A Review of Developments and Applications. J. Sens. 2017, 2017, 1353691. [Google Scholar] [CrossRef] [Green Version]
- LeBourgeois, V.; Bégué, A.; Labbé, S.; Mallavan, B.; Prévot, L.; Roux, B. Can commercial digital cameras be used as multispectral sensors? A crop monitoring test. Sensors 2008, 8, 7300–7322. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Merzlyak, M.N. Remote sensing of chlorophyll concentration in higher plant leaves. Adv. Space Res. 1998, 22, 689–692. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel algorithms for remote estimation of vegetation fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Lobell, D.; Asner, G. Hyperion Studies of Crop Stress in Mexico. In Proceedings of the 12th Annual JPL Airborne Earth Science Workshop, Pasadena, CA, USA, 24–28 February 2003. [Google Scholar]
- Environment System Research Institute (ESRI). Accuracy Assessment for Image Classification. 2019. Available online: https://desktop.arcgis.com/en/arcmap/latest/manage-data/raster-and-images/accuracy-assessment-for-image-classification.htm (accessed on 15 June 2021).
- Foody, G.M. Status of land cover classification accuracy assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- Nielsen, A.A.; Conradsen, K.; Simpson, J.J. Multivariate Alteration Detection (MAD) and MAF Postprocessing in Multispectral, Bitemporal Image Data: New Approaches to Seasonal change Detection Studies. Remote Sens. Environ. 1998, 64, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lamb, J.B.; van de Water, J.A.J.M.; Bourne, D.G.; Altier, C.; Hein, M.Y.; Fiorenza, E.A.; Abu, N.; Jompa, J.; Harvell, C.D. Seagrass ecosystems reduce exposure to bacterial pathogens of humans, fishes, and invertebrates. Science 2017, 355, 731–733. [Google Scholar] [CrossRef]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; de Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountrakis, G.; Im, J.; Ogole, C. Support vector machines in remote sensing: A review. ISPRS J. Photogramm. Remote Sens. 2011, 66, 247–259. [Google Scholar] [CrossRef]
- Traganos, D.; Reinartz, P. Interannual seasonal change detection of mediterranean seagrasses using RapidEye image time series. Front. Plant Sci. 2018, 9, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.R.; Williams, S.L.; Duarte, C.M.; Heck, K.L.; Waycott, M. Associations of concern: Declining seagrasses and threatened dependent species. Front. Ecol. Environ. 2009, 7, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Alderson, S.L. The Analysis of Ecological Experiments in Which Overdispersion Is Present. Unpublished. Master’s Thesis, University of Auckland, Auckland, New Zealand, 1997. [Google Scholar]
- Bell, S.; Hicks, G. Marine landscapes and faunal recruitment: A field test with seagrasses and copepods. Mar. Ecol. Prog. Ser. 1991, 73, 61–68. [Google Scholar] [CrossRef]
- Berkenbusch, K.; Rowden, A.; Probert, P.K. Temporal and spatial variation in macrofauna community composition imposed by ghost shrimp Callianassa filholi bioturbation. Mar. Ecol. Prog. Ser. 2000, 192, 249–257. [Google Scholar] [CrossRef]
- Van Houte-Howes, K.S.S.; Turner, S.J.; Pilditch, C.A. Spatial differences in macroinvertebrate communities on intertidal sand flats: A comparison between seagrass habitats and unvegetated sediment in three estuaries on the Coromandel Peninsula, New Zealand. Estuaries 2004, 27, 945–957. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | VIS Orthomosaic | VIS+NIR Orthomosaic | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seasonal Time-Series | Summer | Autumn | Winter | Summer | Autumn | Winter | ||||||
| Density Classes | U-acc | P-acc | U-acc | P-acc | U-acc | P-acc | U-acc | P-acc | U-acc | P-acc | U-acc | P-acc |
| Absent | 0.99 | 0.99 | 0.98 | 0.99 | 0.97 | 0.99 | 1 | 0.99 | 1 | 0.99 | 1 | 0.99 |
| Low | 0.76 | 0.88 | 0.78 | 0.90 | 0.85 | 0.82 | 0.81 | 0.75 | 0.80 | 0.86 | 0.90 | 0.97 |
| Medium | 0.82 | 0.66 | 0.63 | 0.69 | 0.72 | 0.41 | 0.66 | 0.88 | 0.86 | 0.79 | 0.89 | 0.85 |
| High | 0.99 | 0.99 | 0.97 | 0.93 | 0.95 | 0.97 | 0.99 | 0.97 | 0.98 | 0.97 | 0.96 | 0.94 |
| Overall accuracy | 90% | 91% | 98% | 92% | 94% | 98% | ||||||
| Kappa | 0.92 | 0.87 | 0.90 | 0.95 | 0.88 | 0.95 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chand, S.; Bollard, B. Detecting the Spatial Variability of Seagrass Meadows and Their Consequences on Associated Macrofauna Benthic Activity Using Novel Drone Technology. Remote Sens. 2022, 14, 160. https://doi.org/10.3390/rs14010160
Chand S, Bollard B. Detecting the Spatial Variability of Seagrass Meadows and Their Consequences on Associated Macrofauna Benthic Activity Using Novel Drone Technology. Remote Sensing. 2022; 14(1):160. https://doi.org/10.3390/rs14010160
Chicago/Turabian StyleChand, Subhash, and Barbara Bollard. 2022. "Detecting the Spatial Variability of Seagrass Meadows and Their Consequences on Associated Macrofauna Benthic Activity Using Novel Drone Technology" Remote Sensing 14, no. 1: 160. https://doi.org/10.3390/rs14010160
APA StyleChand, S., & Bollard, B. (2022). Detecting the Spatial Variability of Seagrass Meadows and Their Consequences on Associated Macrofauna Benthic Activity Using Novel Drone Technology. Remote Sensing, 14(1), 160. https://doi.org/10.3390/rs14010160