
GPR-Based Automatic Identification of Root Zones of Influence Using HDBSCAN

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
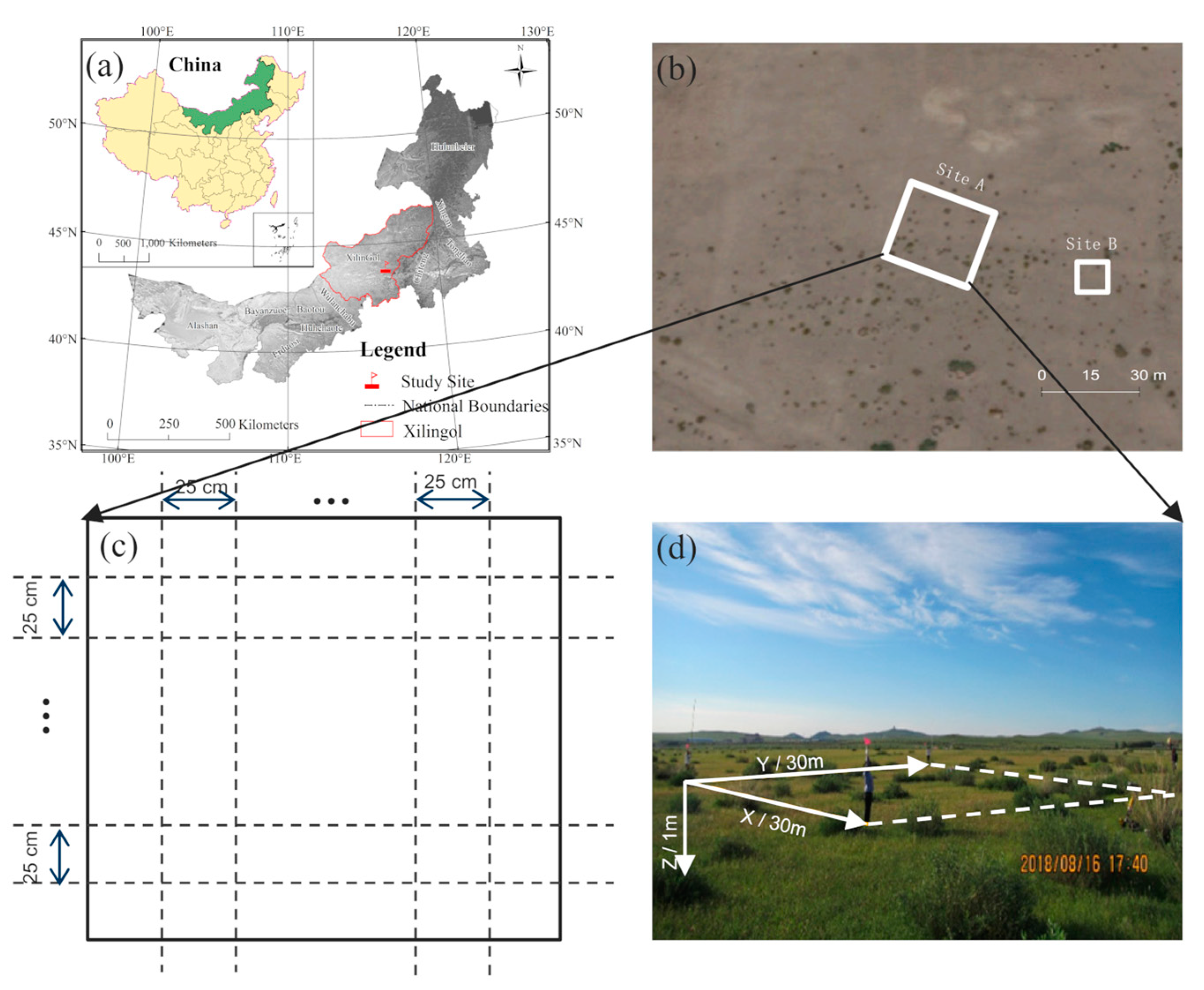
2.1. Study Site
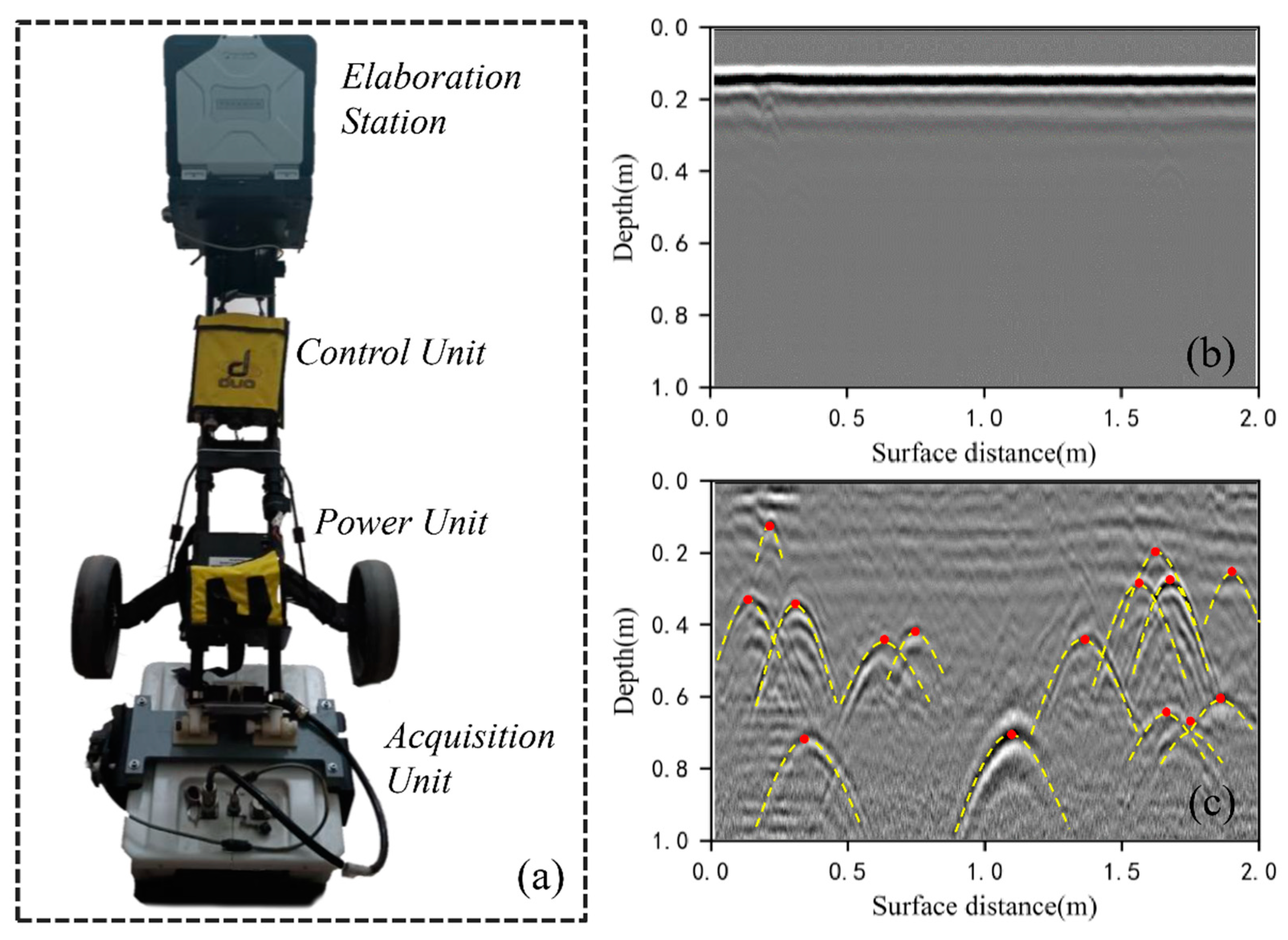
2.2. Field Data Collection
2.3. GPR Data Processing
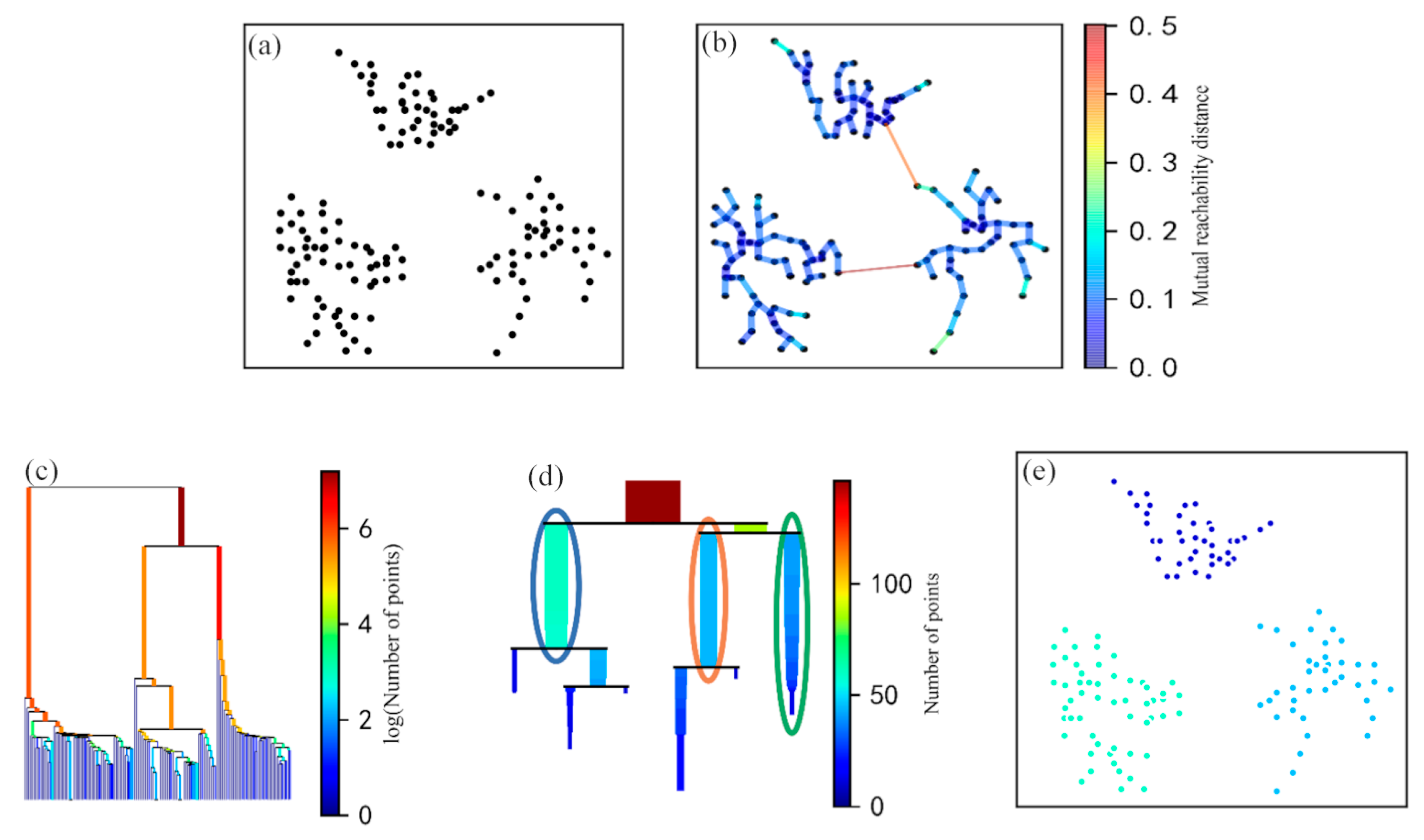
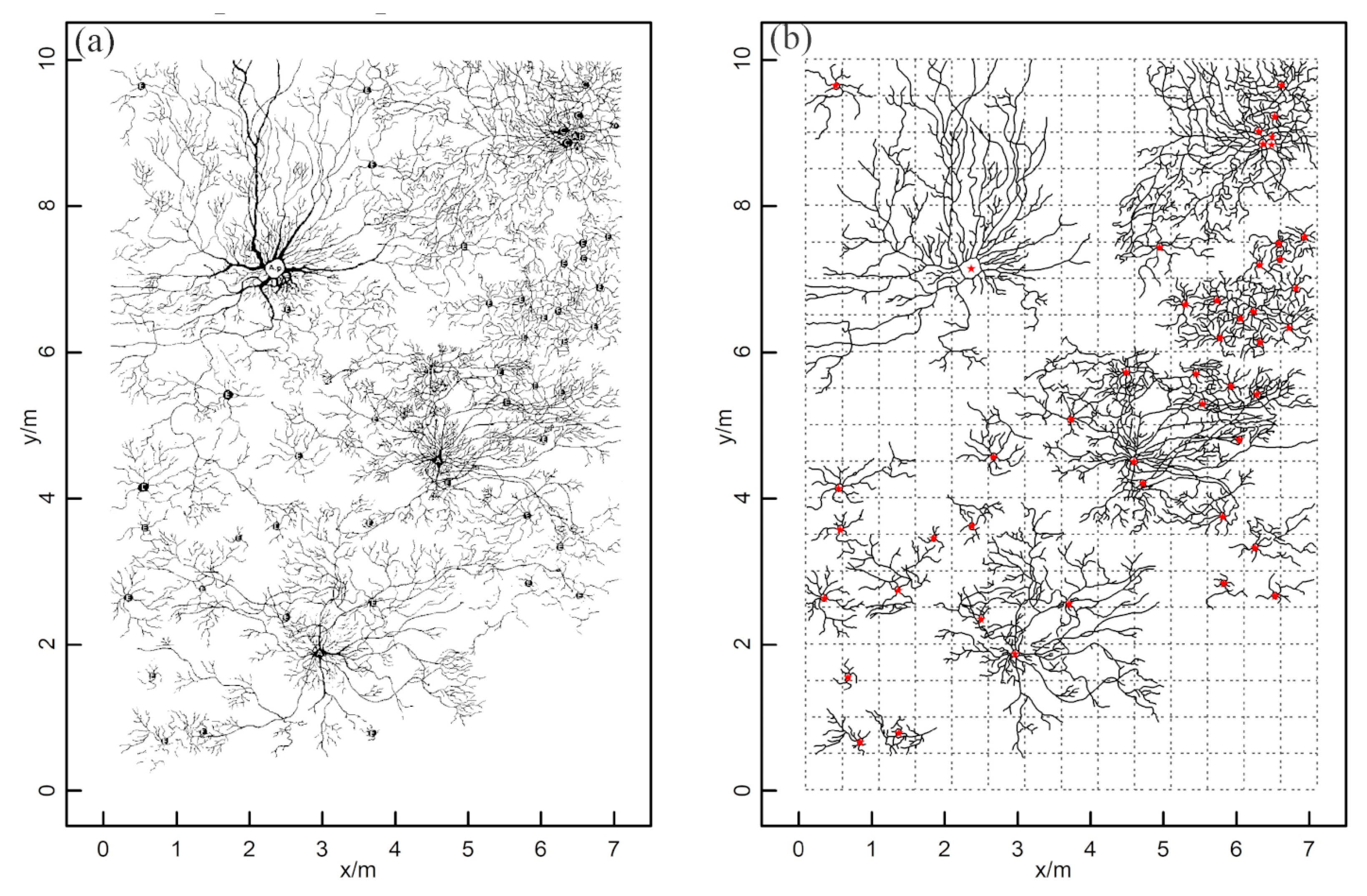
2.4. Identification of the ZOI from GPR-Based Root Measurements
2.5. Validation Method
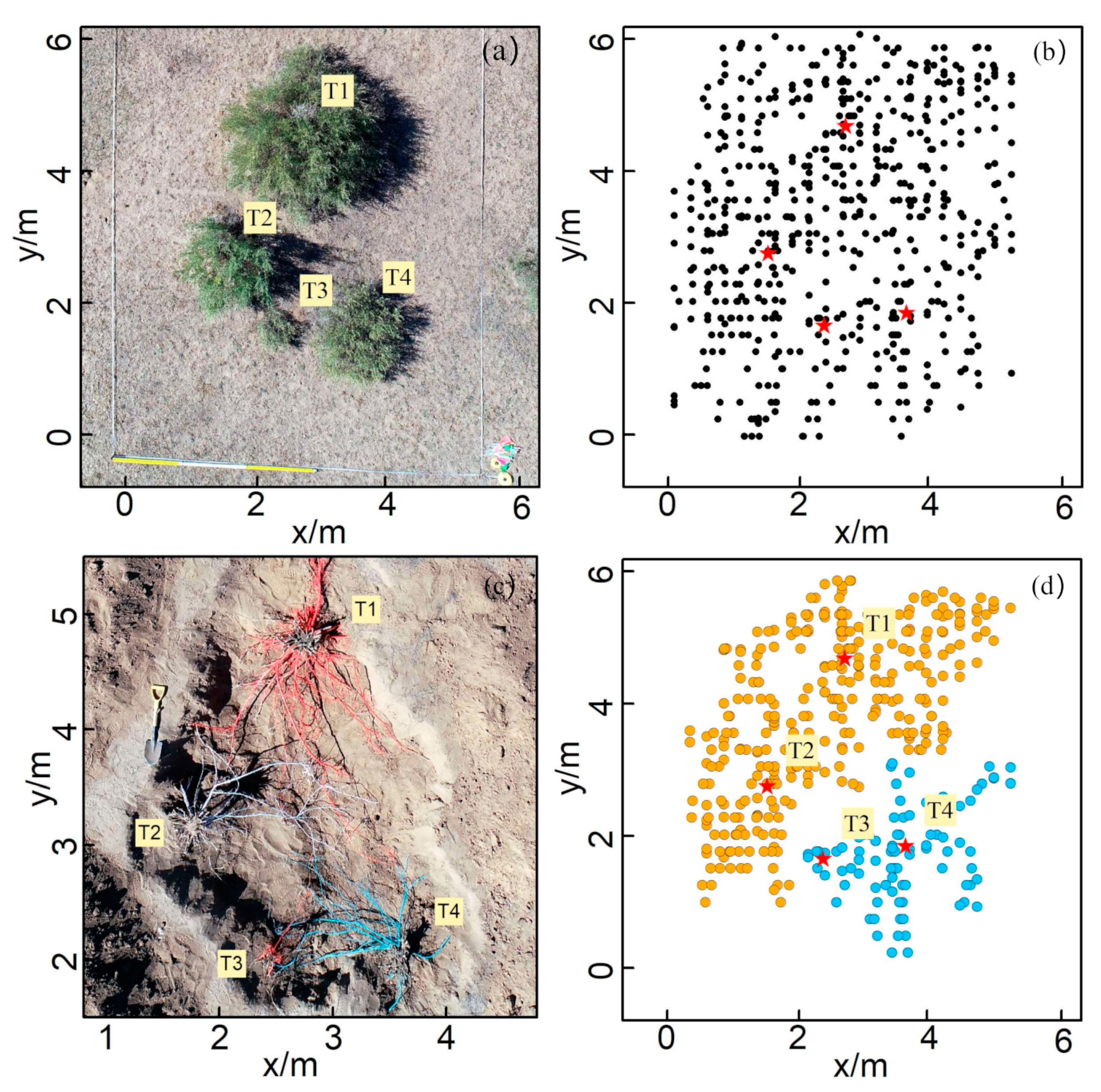
2.5.1. Validation with Field Data
2.5.2. Validation with Simulated Data
2.6. Analysis of the Characteristics of ZOI
3. Results
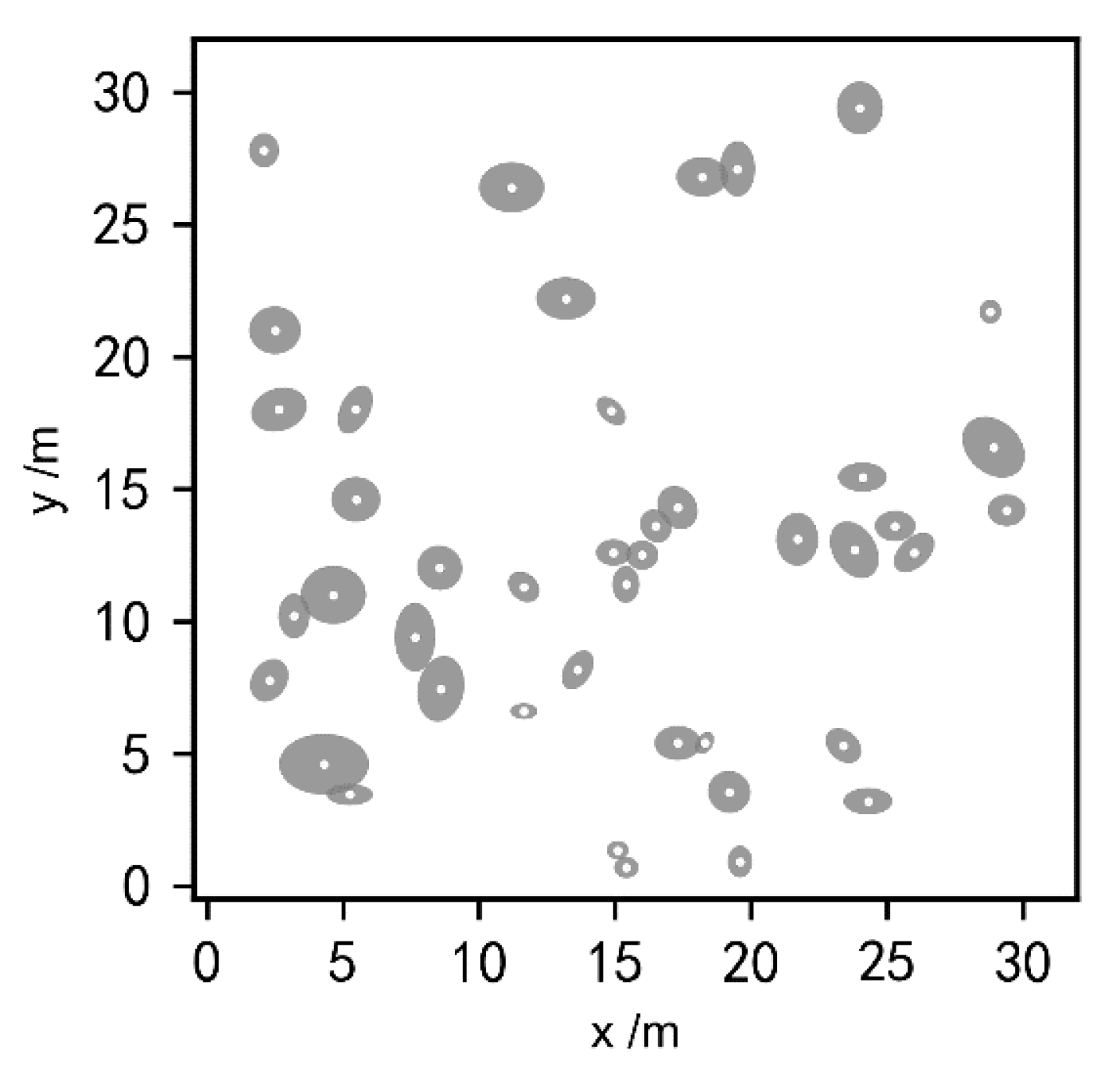
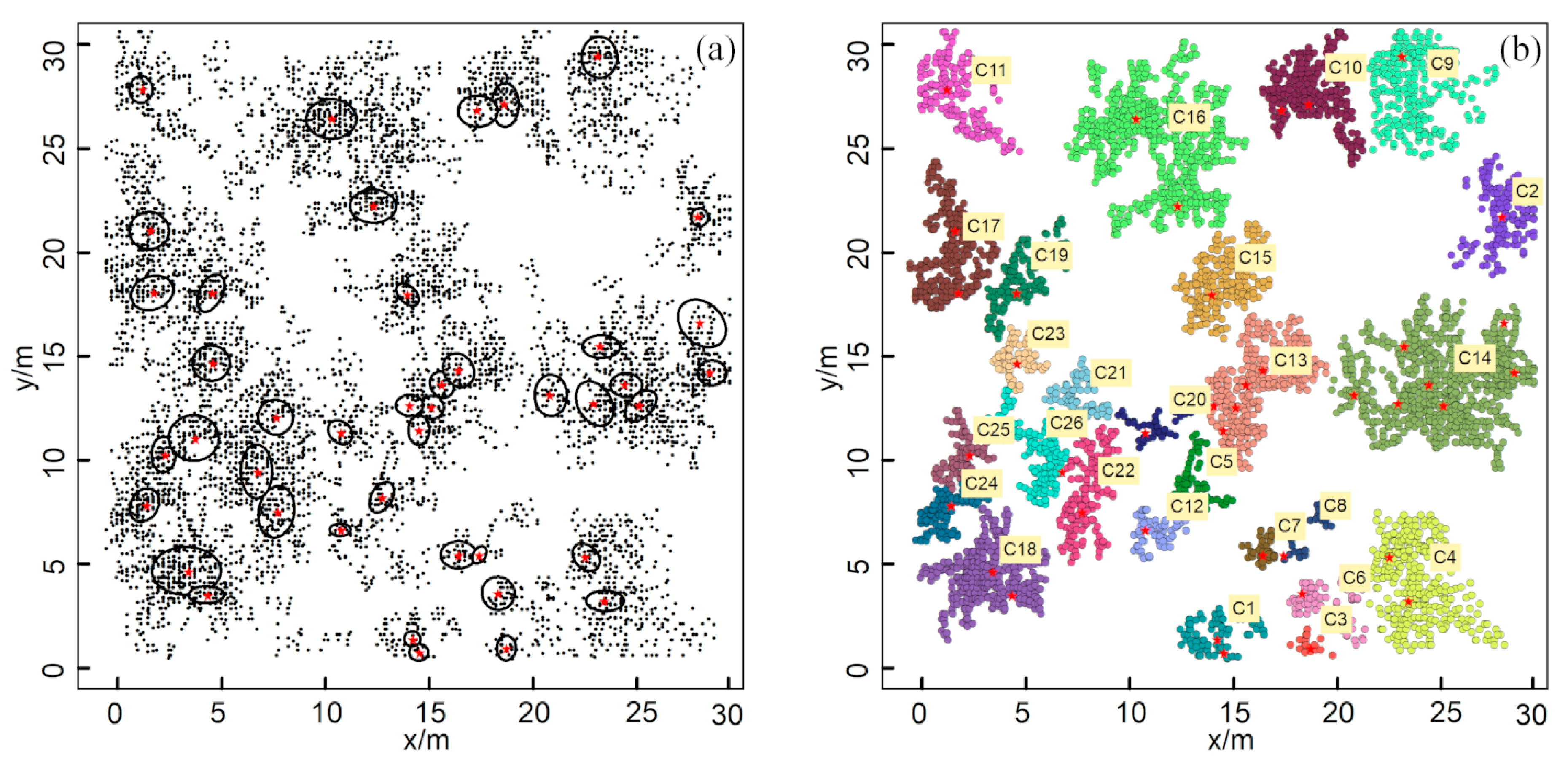
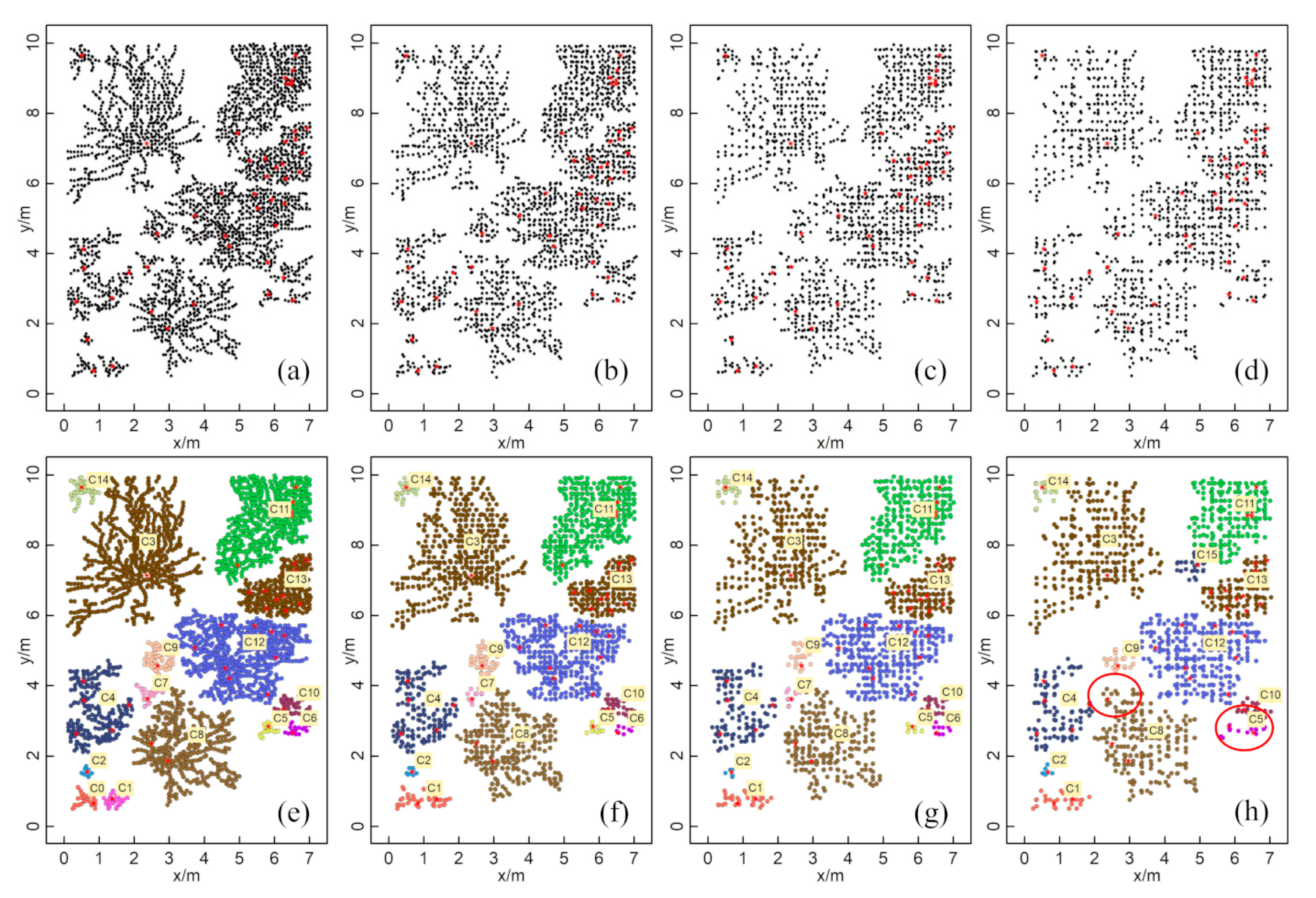
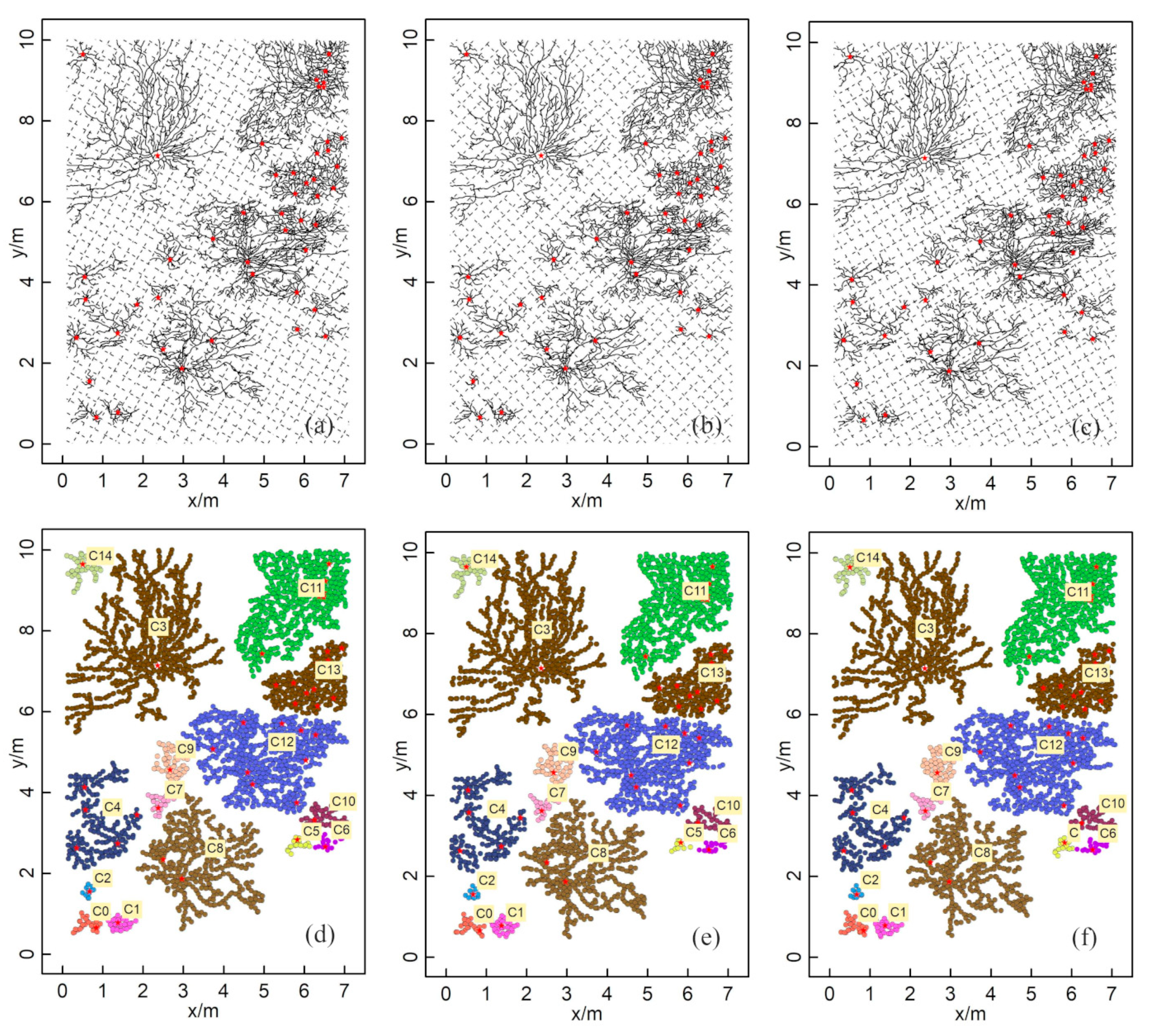
3.1. The Extraction of ZOIs Based on Field Data
3.2. The Validation of ZOIs Based on the Simulated Data
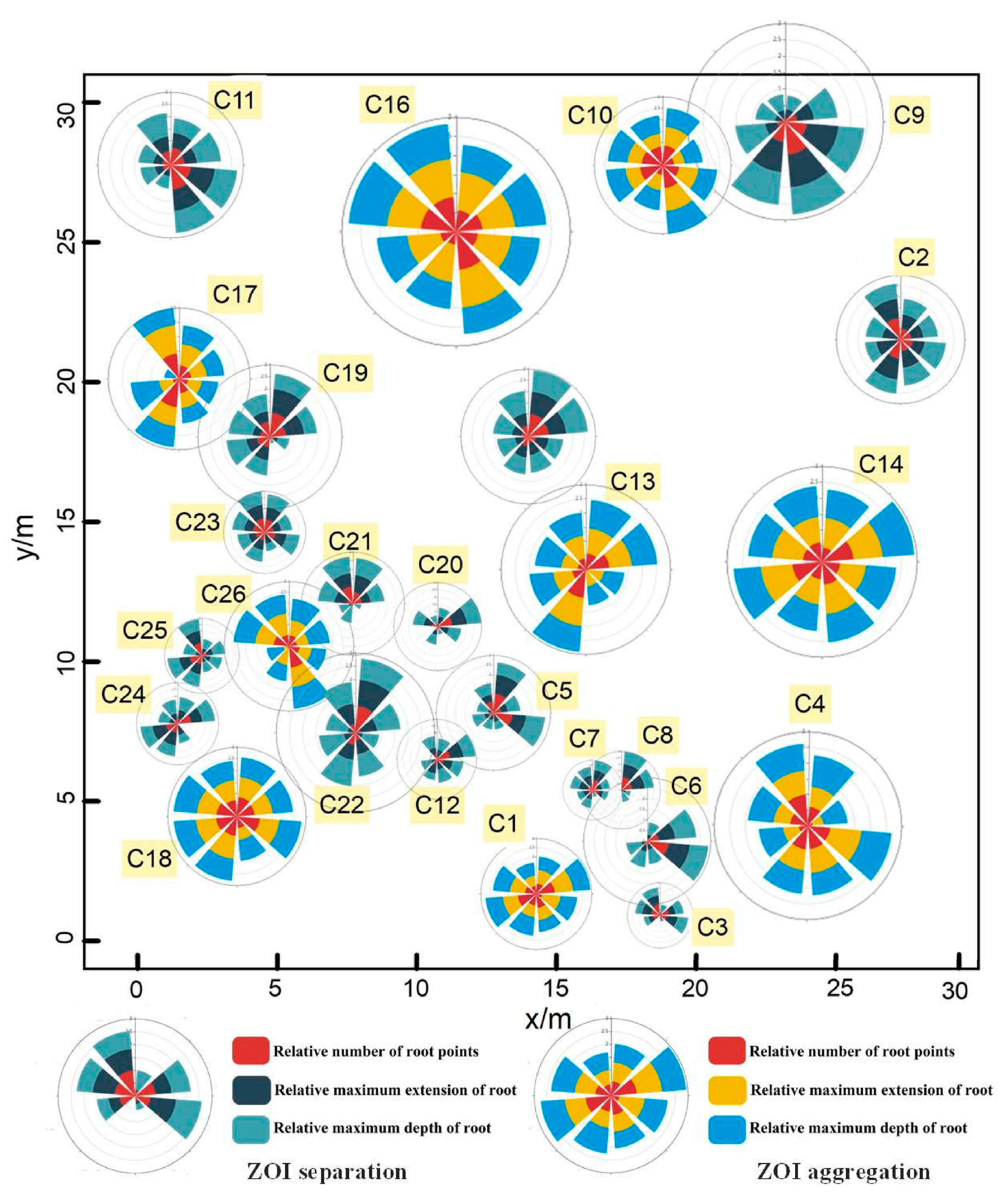
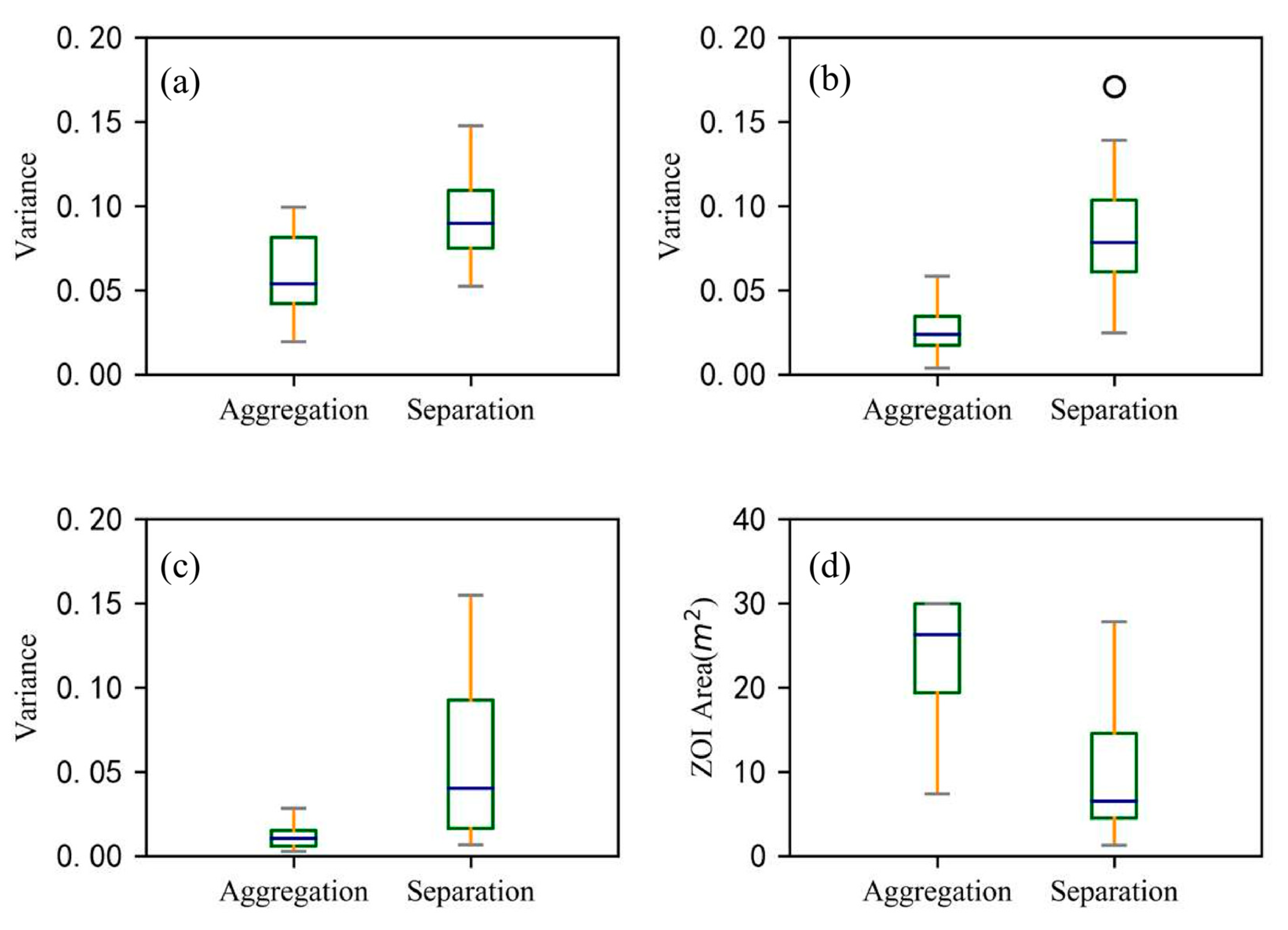
3.3. Characteristics of ZOI
4. Discussion
4.1. Advantages and Applicability of the Method
4.2. Characteristics of ZOIs on the C. microphylla Population Scale
4.3. Limitations of this Method
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antenna Frequency (MHz) | Soil Type | Soil Texture | Soil Drainage Condition | Plant Species | Maximum Detection Depth (m) | Minimum Detectable Root Diameter (cm) | Reference | |
|---|---|---|---|---|---|---|---|---|
| Sand (%) | Clay and Silt (%) | |||||||
| 250 | - | - | - | Poor | Colophospermum mopane | 4.00 | - | Schoor and Colvin [61] |
| 400 | Gergeville soil | 65 | 35 | Well | Pinus taeda | 1.00 | 3.7 | Butnor et al. [19] |
| 400 | Lynchburg soil | 70 | 30 | Poor | Pinus taeda | 1.30 | - | Butnor et al. [19] |
| 450 | Loamy deluvium soil | 30–60 | 40–70 | Well | Quercus petraea | 2.20 | 3.0–4.0 | Hruška et al. [20] |
| 450 | Loess-Clay soil | <50 * | >50 * | Poor | Acer campestre | 2.00 | 2.0–3.0 | Čermák et al. [62] |
| 450 | Loess-Clay soil | <50 * | >50* | Well | Pinus nigra | 2.50 | 2 | Stokes et al. [63] |
| 500 | River sand | 100 * | 0 * | Well | Eucalyptus sp. | - | 1 | Barton and Montagub [35] |
| 800 | River sand | 100 * | 0 * | Well | Eucalyptus sp. | 1.55 | <1.0 | Barton and Montagu [35] |
| 900 | Loamy sand | 92 | 7 | - | Prunus persica | 1.14 | 2.5 | Cox et al. [37] |
| 900 | Loamy sand | 85 | 15 | - | Prunus persica | - | 1.2 | Cox et al. [37] |
| 900 | Sand | 100 * | 0 * | Poor | Cryptomeria japonica | - | 1.1 | Dannoura et al. [64] |
| 900 | Sand | 100 * | 0 * | Well | Cryptomeria japonica | 0.80 | 1.9 | Hirano et al. [58] |
| 1000 | River sand | 100 * | 0 * | Well | Eucalyptus sp. | 1.55 | <1.0 | Barton and Montagu [35] |
| 1500 | Sand | 100 * | 0 * | Well | - | - | 0.25 | Wielopolski et al. [65] |
| 1500 | Lakeland soil | 90 | 10 | Well | Populus deltoides | 0.45 | 0.6 | Butnor et al. [19] |
| 1500 | Wakulla soil | 85–92 | 8–15 | Well | Pinus taeda | 0.50 | 0.5 | Butnor et al. [19] |
| 1500 | Gergeville soil | 65 | 35 | Well | Pinus taeda | 0.60 | - | Butnor et al. [19] |
| 1500 | Troup and Lucy soil | >70 * | <30 * | Well | Pinus taeda | 0.50 | 0.5 | Butnor et al. [36] |
| 1500 | Sandy Pomello soil | >90 * | <10 * | Well | Quercus sp | 0.60 | 0.5 | Stover et al. [66] |
| 2000 | Sand | 95 | 5 | Well | Ulmus pumila | 0.80 | 0.5 | Cui et al. [59] |
References
- Deans, J. Dynamics of coarse root production in a young plantation of Picea sitchensis. Forestry 1981, 54, 139–155. [Google Scholar] [CrossRef]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Wang, P.; Mou, P.; Li, Y.-B. Review of root nutrient foraging plasticity and root competition of plants. Chin. J. Plant Ecol. 2012, 36, 1184. [Google Scholar] [CrossRef]
- Schenk, H.J. Root competition: Beyond resource depletion. J. Ecol. 2006, 94, 725–739. [Google Scholar] [CrossRef]
- Casper, B.B.; Schenk, H.J.; Jackson, R.B. Defining a plant’s belowground zone of influence. Ecology 2003, 84, 2313–2321. [Google Scholar] [CrossRef]
- Mou, P.; Mitchell, R.J.; Jones, R.H. Ecological field theory model: A mechanistic approach to simulate plant–plant interactions in southeastern forest ecosystems. Can. J. For. Res. 1993, 23, 2180–2193. [Google Scholar] [CrossRef]
- Biondini, M.E.; Grygiel, C.E. Landscape distribution of organisms and the scaling of soil resources. Am. Nat. 1994, 143, 1026–1054. [Google Scholar] [CrossRef]
- Bella, I.E. A new competition model for individual trees. For. Sci. 1971, 17, 364–372. [Google Scholar]
- Berger, U.; Hildenbrandt, H. A new approach to spatially explicit modelling of forest dynamics: Spacing, ageing and neighbourhood competition of mangrove trees. Ecol. Modell. 2000, 132, 287–302. [Google Scholar] [CrossRef]
- Brisson, J.; Reynolds, J.F. The effect of neighbors on root distribution in a creosotebush (Larrea tridentata) population. Ecology 1994, 75, 1693–1702. [Google Scholar] [CrossRef]
- Brisson, J.; Reynolds, J.F. Effects of Compensatory Growth On Populationprocesses: A Simulation Study. Ecology 1997, 78, 2378–2384. [Google Scholar] [CrossRef]
- O’Brien, E.E.; Brown, J.S.; Moll, J.D. Roots in space: A spatially explicit model for below-ground competition in plants. Proc. R. Soc. B 2007, 274, 929–935. [Google Scholar] [CrossRef]
- Martin, M.; Snaydon, R.; Drennan, D. Lithium as a non-radioactive tracer for roots of intercropped species. Plant Soil 1982, 64, 203–208. [Google Scholar] [CrossRef]
- Schiffers, K.; Tielbörger, K.; Tietjen, B.; Jeltsch, F. Root plasticity buffers competition among plants: Theory meets experimental data. Ecology 2011, 92, 610–620. [Google Scholar] [CrossRef]
- Kummerow, J.; Krause, D.; Jow, W. Root systems of chaparral shrubs. Oecologia 1977, 29, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Zenone, T.; Morelli, G.; Teobaldelli, M.; Fischanger, F.; Matteucci, M.; Sordini, M.; Armani, A.; Ferrè, C.; Chiti, T.; Seufert, G. Preliminary use of ground-penetrating radar and electrical resistivity tomography to study tree roots in pine forests and poplar plantations. Funct. Plant Biol. 2008, 35, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Daniels, D.J. Surface-Penetrating Radar. Electron. Commun. Eng. J. 1996, 8, 165–182. [Google Scholar] [CrossRef]
- Butnor, J.R.; Doolittle, J.; Kress, L.; Cohen, S.; Johnsen, K.H. Use of ground-penetrating radar to study tree roots in the southeastern United States. Tree Physiol. 2001, 21, 1269–1278. [Google Scholar] [CrossRef]
- Hruska, J.; Čermák, J.; Šustek, S. Mapping tree root systems with ground-penetrating radar. Tree Physiol. 1999, 19, 125–130. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, L.; Cui, X.; Chen, J.; Cao, X.; Lin, H. Ground-penetrating radar-based automatic reconstruction of three-dimensional coarse root system architecture. Plant Soil 2014, 383, 155–172. [Google Scholar] [CrossRef]
- Cui, X.; Liu, X.; Cao, X.; Fan, B.; Zhang, Z.; Chen, J.; Chen, X.; Lin, H.; Guo, L. Pairing dual-frequency GPR in summer and winter enhances the detection and mapping of coarse roots in the semi-arid shrubland in China. Eur. J. Soil Biol. 2020, 71, 236–251. [Google Scholar] [CrossRef]
- Rahman, M.F.; Liu, W.; Suhaim, S.B.; Thirumuruganathan, S.; Zhang, N.; Das, G. Hdbscan: Density based clustering over location based services. arXiv 2016, arXiv:1602.03730. [Google Scholar]
- Campello, R.J.; Moulavi, D.; Zimek, A.; Sander, J. A framework for semi-supervised and unsupervised optimal extraction of clusters from hierarchies. Data Min. Knowl. Discov. 2013, 27, 344–371. [Google Scholar] [CrossRef]
- Veerhoek, L. Clustering Satellite Data to Define Eutrophication Monitoring Zones Based on Chlorophyll-a Concentration. Bachelor’s Thesis, Delft University of Technology, Delft, The Netherlands, 2020. [Google Scholar]
- Rosalina, E.; Salim, F.D.; Sellis, T. Automated density-based clustering of spatial urban data for interactive data exploration. In Proceedings of the 2017 IEEE Conference on Computer Communications Workshops (INFOCOM WKSHPS), Atlanta, GA, USA, 1–4 May 2017; pp. 295–300. [Google Scholar]
- Lin, C.-H.; Hsu, K.-C.; Johnson, K.R.; Luby, M.; Fann, Y.C. Applying density-based outlier identifications using multiple datasets for validation of stroke clinical outcomes. Int. J. Med. Inform. 2019, 132, 103988. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Niu, H.; Li, H.; Zhao, M.; Han, X.; Dong, X. Relationship between soil water content and vertical distribution of root system under different ground water gradients in Maowusu Sandy Land. J. Arid Land Resour. Environ. 2008, 22, 157–163. [Google Scholar]
- Wang, J.; Guo, Y.; Yao, Y.; Tang, J.; Zhang, M. Distribution characteristics of root system and carbon stock of Caragana microphylla Lam. in Aohan sandificational area. J. Northwest A F Univ. 2017, 45, 103–110. [Google Scholar]
- Alamusa, J.D.; Tiefan, P. Soil moisture infiltration dynamics in plantation of Caragana microphylla in Heerqin sandy land. Chin. J. Ecol. 2004, 1, 56. [Google Scholar]
- Guo, L.; Chen, J.; Cui, X.; Fan, B.; Lin, H. Application of ground penetrating radar for coarse root detection and quantification: A review. Plant Soil 2013, 362, 1–23. [Google Scholar] [CrossRef]
- Murray, C.; Keiswetter, D. Application of magnetic and multi-frequency EM techniques for landfill investigations: Case histories. In Proceedings of the 11th EEGS Symposium on the Application of Geophysics to Engineering and Environmental Problems, Chicago, IL, USA, 22–26 March 1998; p. cp-203-00047. [Google Scholar]
- Sato, T.; Takada, K.; Wakayama, T.; Kimura, I.; Abe, T.; Shinbo, T. Automatic data processing procedure for ground probing radar. IEICE Trans. Commun. 1994, 77, 831–837. [Google Scholar]
- Barton, C.V.; Montagu, K.D. Detection of tree roots and determination of root diameters by ground penetrating radar under optimal conditions. Tree Physiol. 2004, 24, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Butnor, J.R.; Doolittle, J.; Johnsen, K.H.; Samuelson, L.; Stokes, T.; Kress, L. Utility of ground-penetrating radar as a root biomass survey tool in forest systems. Soil Sci. Soc. Am. J. 2003, 67, 1607–1615. [Google Scholar] [CrossRef]
- Cox, K.; Scherm, H.; Serman, N. Ground-penetrating radar to detect and quantify residual root fragments following peach orchard clearing. HortTechnology 2005, 15, 600–607. [Google Scholar] [CrossRef]
- Li, W.; Cui, X.; Guo, L.; Chen, J.; Chen, X.; Cao, X. Tree root automatic recognition in ground penetrating radar profiles based on randomized Hough transform. Remote Sens. 2016, 8, 430. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Astels, S. hdbscan: Hierarchical density based clustering. J. Open Res. Softw. 2017, 2, 205. [Google Scholar] [CrossRef]
- Eldridge, J.; Belkin, M.; Wang, Y. Beyond hartigan consistency: Merge distortion metric for hierarchical clustering. In Proceedings of the Conference on Learning Theory, Paris, France, 3–6 July 2015; pp. 588–606. [Google Scholar]
- Faget, M.; Nagel, K.A.; Walter, A.; Herrera, J.M.; Jahnke, S.; Schurr, U.; Temperton, V.M. Root–root interactions: Extending our perspective to be more inclusive of the range of theories in ecology and agriculture using in-vivo analyses. Ann. Bot. 2013, 112, 253–266. [Google Scholar] [CrossRef]
- Gupta, P.; Rustgi, S. Molecular markers from the transcribed/expressed region of the genome in higher plants. Funct. Integr. Genomics. 2004, 4, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Mahall, B.E.; Callaway, R.M. Root communication among desert shrubs. Proc. Natl. Acad. Sci. USA 1991, 88, 874–876. [Google Scholar] [CrossRef]
- Schmid, C.; Bauer, S.; Müller, B.; Bartelheimer, M. Belowground neighbor perception in Arabidopsis thaliana studied by transcriptome analysis: Roots of Hieracium pilosella cause biotic stress. Front. Plant Sci. 2013, 4, 296. [Google Scholar] [CrossRef]
- Semchenko, M.; Zobel, K.; Heinemeyer, A.; Hutchings, M.J. Foraging for space and avoidance of physical obstructions by plant roots: A comparative study of grasses from contrasting habitats. New Phytol. 2008, 179, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.J.; Callaway, R.M.; Mahall, B. Spatial root segregation: Are plants territorial? In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1999; Volume 28, pp. 145–180. [Google Scholar]
- Zahavi, A.; Zahavi, A. The Handicap Principle: A Missing Piece of Darwin’s Puzzle; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Schenk, H.J.; Seabloom, E.W. Evolutionary ecology of plant signals and toxins: A conceptual framework. In Plant Communication from an Ecological Perspective; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–19. [Google Scholar]
- Bartelheimer, M.; Steinlein, T.; Beyschlag, W. Aggregative root placement: A feature during interspecific competition in inland sand-dune habitats. Plant Soil 2006, 280, 101–114. [Google Scholar] [CrossRef]
- Semchenko, M.; John, E.A.; Hutchings, M.J. Effects of physical connection and genetic identity of neighbouring ramets on root-placement patterns in two clonal species. New Phytol. 2007, 176, 644–654. [Google Scholar] [CrossRef]
- Caffaro, M.M.; Vivanco, J.M.; Botto, J.; Rubio, G. Root architecture of Arabidopsis is affected by competition with neighbouring plants. Plant Growth Regul. 2013, 70, 141–147. [Google Scholar] [CrossRef]
- Falik, O.; Reides, P.; Gersani, M.; Novoplansky, A. Root navigation by self inhibition. Plant Cell Environ. 2005, 28, 562–569. [Google Scholar] [CrossRef]
- Schmid, C.; Bauer, S.; Bartelheimer, M. Should I stay or should I go? Roots segregate in response to competition intensity. Plant Soil 2015, 391, 283–291. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Cabal, C.; Martínez-García, R.; de Castro Aguilar, A.; Valladares, F.; Pacala, S.W. The exploitative segregation of plant roots. Science 2020, 370, 1197–1199. [Google Scholar] [CrossRef]
- Thomas, S.C.; Weiner, J. Including competitive asymmetry in measures of local interference in plant populations. Oecologia 1989, 80, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Ganatsas, P.; Spanos, I. Root system asymmetry of Mediterranean pines. In Eco-And Ground Bio-Engineering: The Use of Vegetation to Improve Slope Stability; Springer: Berlin/Heidelberg, Germany, 2007; pp. 127–134. [Google Scholar]
- Hirano, Y.; Dannoura, M.; Aono, K.; Igarashi, T.; Ishii, M.; Yamase, K.; Makita, N.; Kanazawa, Y. Limiting factors in the detection of tree roots using ground-penetrating radar. Plant Soil 2009, 319, 15–24. [Google Scholar] [CrossRef]
- Cui, X.; Chen, J.; Shen, J.; Cao, X.; Chen, X.; Zhu, X. Modeling tree root diameter and biomass by ground-penetrating radar. Sci. China Earth Sci. 2011, 54, 711–719. [Google Scholar] [CrossRef]
- Malzer, C.; Baum, M. HDBSCAN (): An Alternative Cluster Extraction Method for HDBSCAN. arXiv 2019, arXiv:1911.02282. [Google Scholar]
- Van Schoor, M.; Colvin, C. Tree root mapping with ground penetrating radar. In Proceedings of the 11th SAGA Biennial Technical Meeting and Exhibition, Swaziland, South Africa, 16–18 September 2009; p. cp-241-00077. [Google Scholar]
- Cermak, J.; Hruska, J.; Martinkova, M.; Prax, A. Urban tree root systems and their survival near houses analyzed using ground penetrating radar and sap flow techniques. Plant Soil 2000, 219, 103–116. [Google Scholar] [CrossRef]
- Stokes, A.; Fourcaud, T.; Hruska, J.; Cermak, J.; Nadyezdhina, N.; Nadyezhdin, V.; Praus, L. An evaluation of different methods to investigate root system architecture of urban trees in situ: I. Ground-penetrating radar. J. Arboric. 2002, 28, 2–10. [Google Scholar]
- Dannoura, M.; Hirano, Y.; Igarashi, T.; Ishii, M.; Aono, K.; Yamase, K.; Kanazawa, Y. Detection of Cryptomeria japonica roots with ground penetrating radar. Plant Biosyst. 2008, 142, 375–380. [Google Scholar] [CrossRef]
- Wielopolski, L.; Hendrey, G.; Daniels, J.J.; McGuigan, M. Imaging tree root systems in situ. In Proceedings of the Eighth International Conference on Ground Penetrating Radar, Gold Coast, Australia, 23–26 May 2000; pp. 642–646. [Google Scholar]
- Stover, D.B.; Day, F.P.; Butnor, J.R.; Drake, B.G. Effect of elevated CO2 on coarse-root biomass in Florida scrub detected by ground-penetrating radar. Ecology 2007, 88, 1328–1334. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Quan, Z.; Chen, X.; Zhang, Z.; Zhou, J.; Liu, X.; Chen, J.; Cao, X.; Guo, L. GPR-Based Automatic Identification of Root Zones of Influence Using HDBSCAN. Remote Sens. 2021, 13, 1227. https://doi.org/10.3390/rs13061227
Cui X, Quan Z, Chen X, Zhang Z, Zhou J, Liu X, Chen J, Cao X, Guo L. GPR-Based Automatic Identification of Root Zones of Influence Using HDBSCAN. Remote Sensing. 2021; 13(6):1227. https://doi.org/10.3390/rs13061227
Chicago/Turabian StyleCui, Xihong, Zhenxian Quan, Xuehong Chen, Zheng Zhang, Junxiong Zhou, Xinbo Liu, Jin Chen, Xin Cao, and Li Guo. 2021. "GPR-Based Automatic Identification of Root Zones of Influence Using HDBSCAN" Remote Sensing 13, no. 6: 1227. https://doi.org/10.3390/rs13061227
APA StyleCui, X., Quan, Z., Chen, X., Zhang, Z., Zhou, J., Liu, X., Chen, J., Cao, X., & Guo, L. (2021). GPR-Based Automatic Identification of Root Zones of Influence Using HDBSCAN. Remote Sensing, 13(6), 1227. https://doi.org/10.3390/rs13061227