
Geospatial Approaches to Monitoring the Spread of Invasive Species of Solidago spp.


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Monitored Plant Species
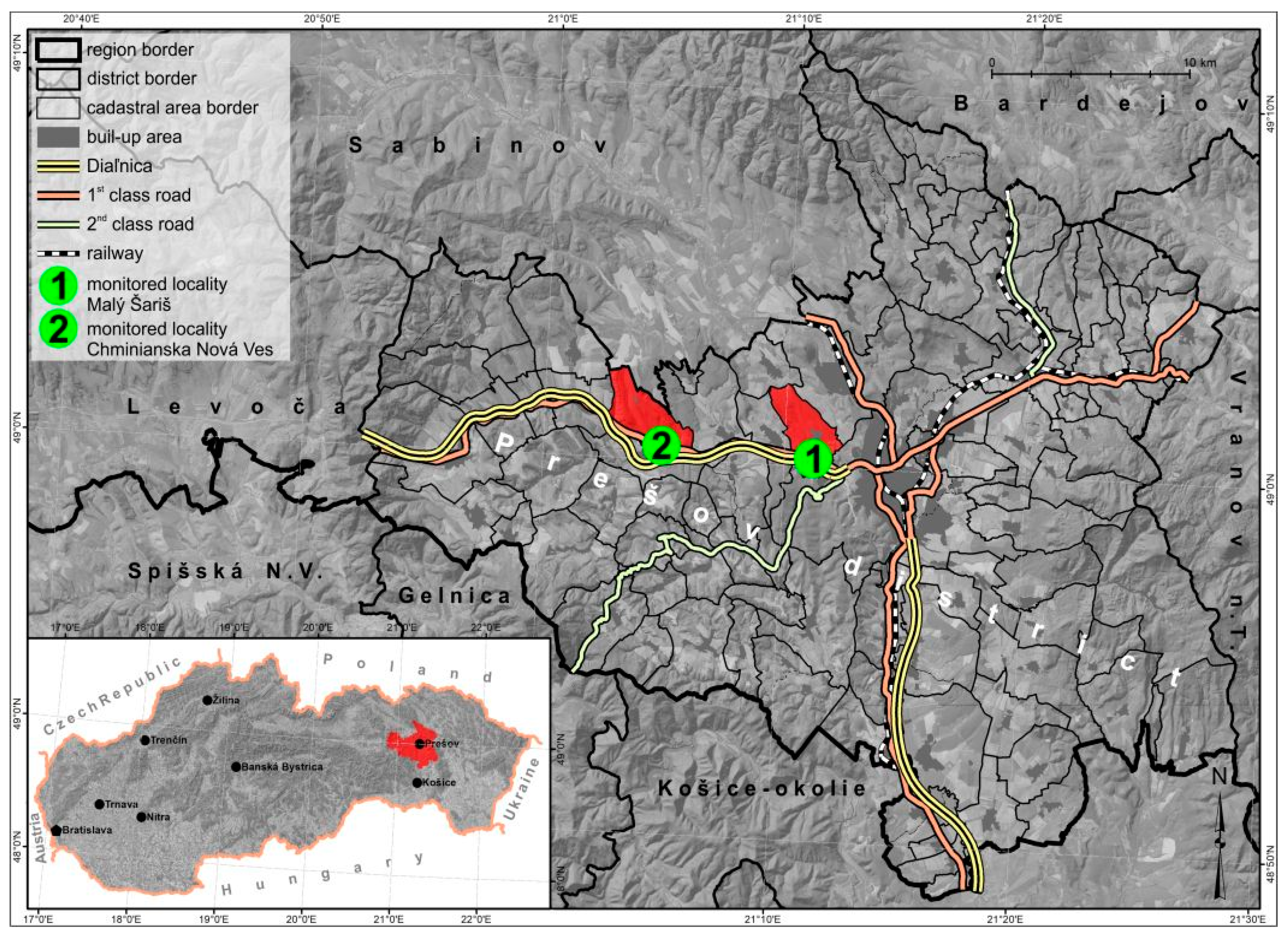
2.2. Monitored Localities
2.3. Locality of Malý Šariš
2.4. Locality Chminianska Nová Ves
2.5. Monitoring of the Solidago spp. Spreading by Using GNSS
2.6. Analysis of the Changes in Solidago spp. Spreading by Using Geospatial Analysis in GIS
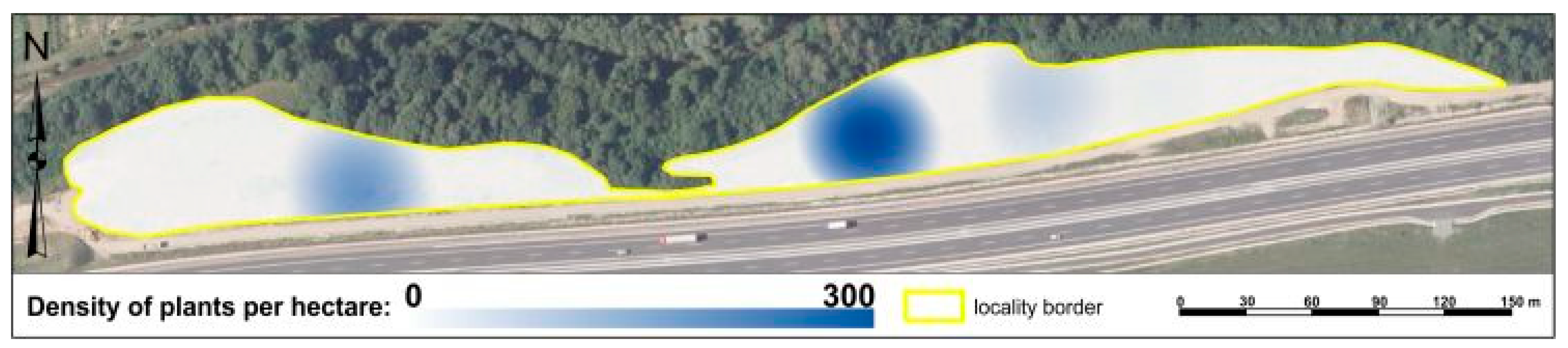
2.7. Kernel Analysis of Plant Density
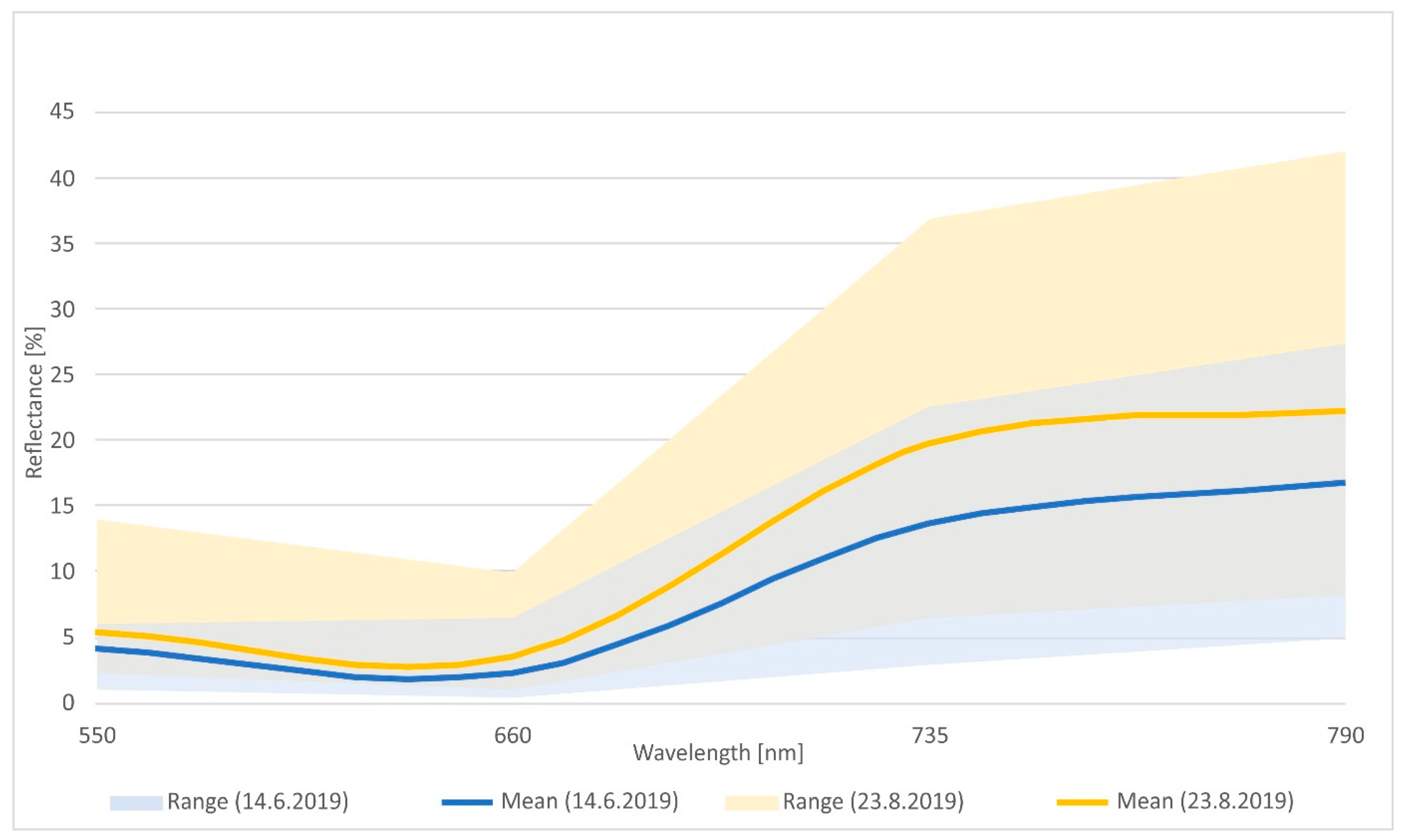
2.8. Evaluation of Spectral Reflectance of Selected Invasive Plants Using Multispectral Drone Imaging
3. Results
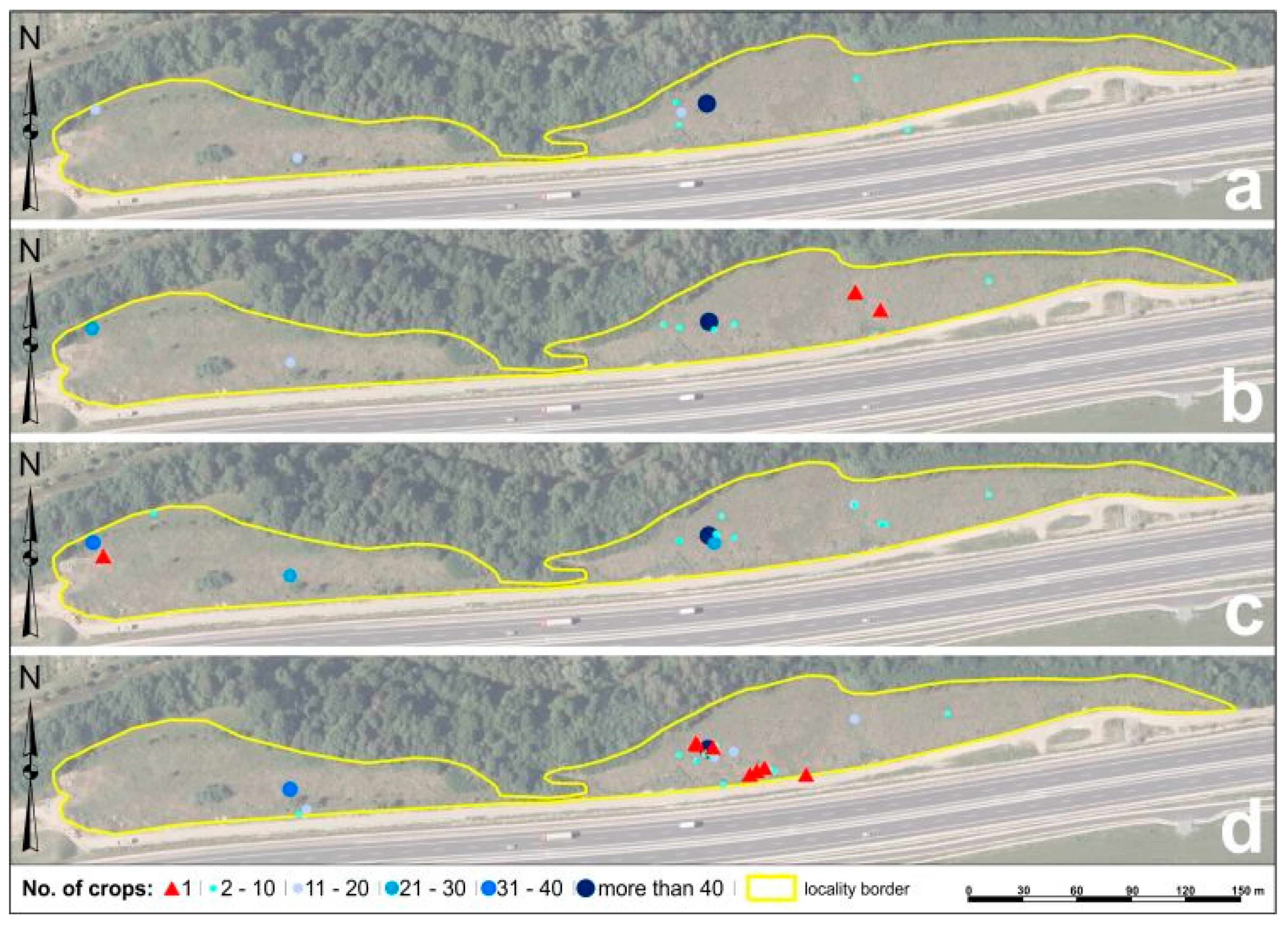
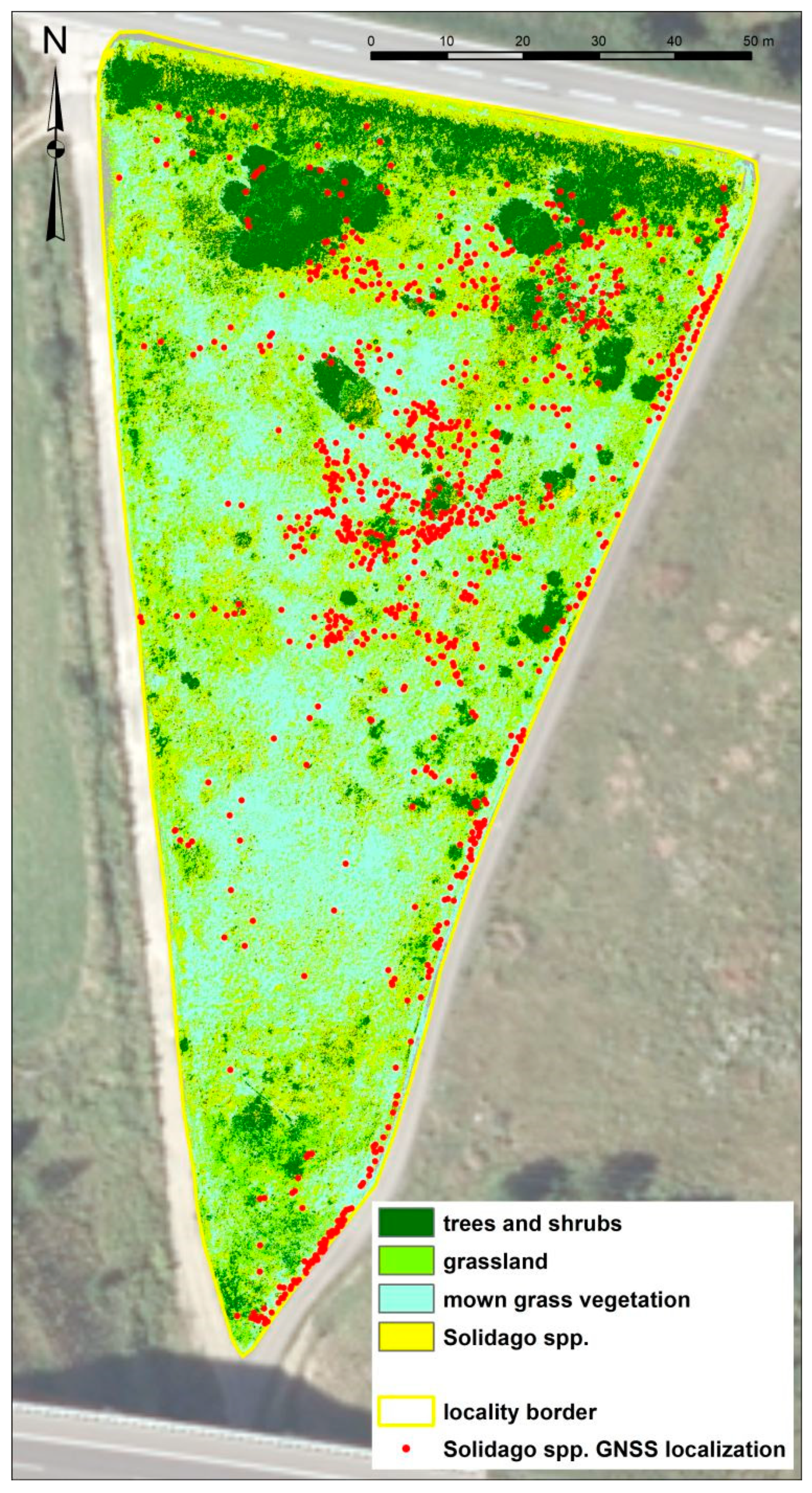
3.1. Mapping the Spread of Solidago spp. via GNSS at the Research Localities
3.1.1. Mapping the Spread of Solidago spp. via GNSS at the Locality of Malý Šariš
3.1.2. Mapping the Spread of Solidago spp. via GNSS at the Chminianska Nová Ves Locality
3.2. Analysis of Solidago spp. Changes in Propagation through Spatial Analyzes Using GIS in Research Localities
3.2.1. Analysis of Solidago spp. Changes in Propagation through Spatial Analyzes Using GIS in Malý Šariš
3.2.2. Analysis of Solidago spp. Changes in Propagation through Spatial Analyzes by GIS in Chminianska Nová Ves
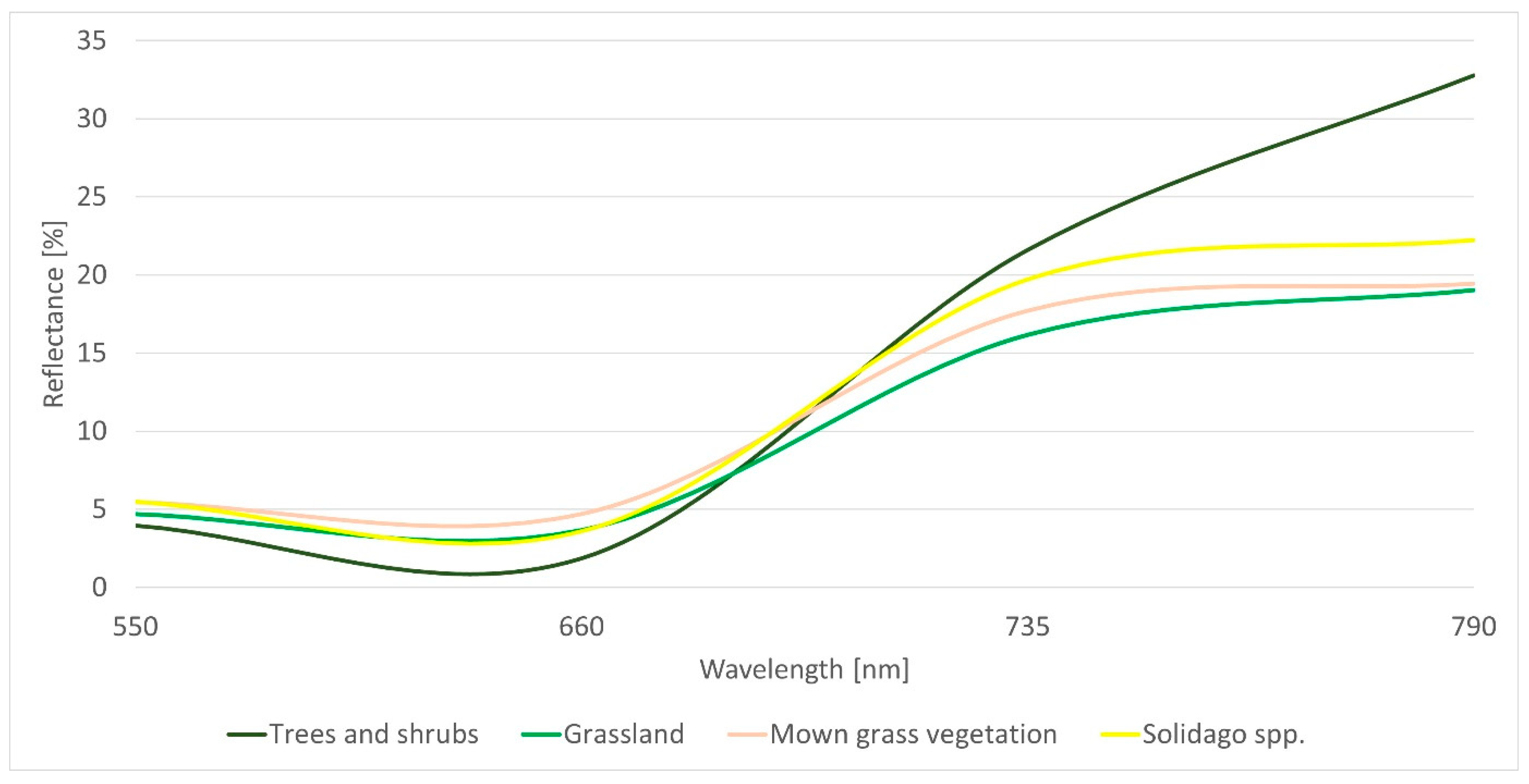
3.3. Spectral Analysis of Solidago spp. Stands in the Locality of Malý Šariš
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle and global change. Annu. Rev. Ecol. Evol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Levine, J.M.; Vila, M.; D’Antonio, C.M.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. B 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Vitousek, P.M.; D’Antonio, C.M.; Asner, G.P. Invasions and ecosystems: Vulnerabilities and the contribution of new technologies. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley Blackwell: Oxford, UK, 2011; pp. 277–288. [Google Scholar]
- He, K.S.; Rocchini, D.; Neteler, M.; Nagendra, H. Benefits of hyperspectral remote sensing for tracking plant invasions. Divers. Distrib. 2011, 17, 381–392. [Google Scholar] [CrossRef]
- Pettorelli, N.; Laurance, W.F.; O’Brien, T.G.; Wegmann, M.; Nagendra, H.; Turner, W. Satellite remote sensing for applied ecologists: Opportunities and challenges. J. Appl. Ecol. 2014, 51, 839–848. [Google Scholar] [CrossRef]
- Peterson, A.T.; Navarro-Sigüenza, A.G.; Benítez-Díaz, H. The need for continued scientific collecting; a geographic analysis of Mexican bird specimens. Ibis 1998, 140, 288–294. [Google Scholar] [CrossRef]
- Schott, J.R. Remote Sensing: The Image Chain Approach; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Asrar, G. Introduction. In Theory and Applications of Optical Remote Sensing; Asrar, G., Ed.; John Wiley & Sons: New York, NY, USA, 1989; p. 1. [Google Scholar]
- Wan, H.; Tang, Y.; Jing, L.; Li, H.; Qiu, F.; Wu, W. Tree Species Classification of Forest Stands Using Multisource Remote Sensing Data. Remote Sens. 2021, 13, 144. [Google Scholar] [CrossRef]
- DeFries, R.S.; Townshend, J.R.G. NDVI-derived land cover classifications at a global scale. Int. J. Remote Sens. 1994, 15, 3567–3586. [Google Scholar] [CrossRef]
- DeFries, R.S.; Hansen, M.C.; Townshend, J.R.G.; Sohlberg, R.S. Global landcover classifications at 8 km spatial resolution: The use of training data derived from Landsat imagery in decision tree classifiers. Int. J. Remote Sens. 1998, 19, 3141–3168. [Google Scholar] [CrossRef]
- Mountrakis, G.; Im, J.; Ogole, C. Support vector machines in remote sensing: A review. ISPRS J. Photogram. Remote Sens. 2011, 66, 247–259. [Google Scholar] [CrossRef]
- Stathaki, T. Image Fusion: Algorithms and Applications; Elsevier Ltd.: London, UK, 2008; 493p. [Google Scholar]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Pettorelli, N. The Normalized Differential Vegetation Index; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Lukasová, V.; Bucha, T.; Škvareninová, J.; Škvarenina, J. Validation and application of European beech phenological metrics derived from MODIS data along an altitudinal gradient. Forests 2019, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.A.; Mustard, J.F. Characterizing the landscape dynamics of an invasive plant and risk of invasion using remote sensing. Ecol. Appl. 2006, 16, 1132–1147. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y. Evaluating High Resolution GeoEye-1 Satellite Imagery for Mapping Wildlife in Open Savannahs; International Institute for Geoinformation Science and Earth Observation: Enschede, The Netherlands, 2012; 52p. [Google Scholar]
- Clark, D.B.; Read, J.M.; Clark, M.L.; Murillo Cruz, A.; Fallas Dotti, M.; Clark, D.A. Application of 1-m and 4-m resolution satellite data to studies of tree demography, stand structure and land-use classification in tropical rain forest landscapes. Ecol. Appl. 2004, 14, 61–74. [Google Scholar] [CrossRef]
- Nagendra, H.; Lucas, R.; Honrado, J.P.; Jongman, R.H.G.; Tarantino, C.; Adamo, M.; Mairota, P. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Turner, W.; Rondinini, C.; Pettorelli, N.; Mora, B.; Leidner, A.K.; Szantoi, Z.; Buchanan, G.; Dech, S.; Dwyer, J.; Herold, M.; et al. Free and open-access satellite data are key to biodiversity conservation. Biol. Conserv. 2015, 182, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Andrew, M.E.; Ustin, S.L. Habitat suitability modelling of an invasive plant with advanced remote sensing data. Divers. Distrib. 2009, 15, 627–640. [Google Scholar] [CrossRef]
- Everitt, J.H.; Deloach, C.J. Remote sensing of Chinese tamarisk (Tamarix chinensis) and associated vegetation. Weed Sci. 1990, 38, 273–278. [Google Scholar] [CrossRef]
- Frazier, B.E.; Moore, B.C. Some tests of film types for remote sensing of purple loosestrife, Lythrum salicaria, at low-densities. Wetlands 1993, 13, 145–152. [Google Scholar] [CrossRef]
- Cook, G.D.; Setterfield, S.A.; Maddison, J.P. Shrub invasion of a tropical wetland: Implications for weed management. Ecol. Appl. 1996, 6, 531–537. [Google Scholar] [CrossRef]
- Brown, J.R.; Carter, J. Spatial and temporal patterns of exotic shrub invasion in an Australian tropical grassland. Landsc. Ecol. 1998, 13, 93–102. [Google Scholar] [CrossRef]
- Maheu-Giroux, M.; De Blois, S. Mapping the invasive species Phragmites australis in linear wetland corridors. Aquat. Bot. 2005, 83, 310–320. [Google Scholar] [CrossRef]
- Müllerová, J.; Pyšek, P.; Jarošík, V.; Pergl, J. Aerial photographs as a tool for assessing the regional dynamics of the invasive plant species Heracleum mantegazzianum. J. Appl. Ecol. 2005, 42, 1042–1053. [Google Scholar] [CrossRef]
- Ge, S.K.; Carruthers, R.; Gong, P.; Herrera, A. Texture analysis for mapping Tamarix parviflora using aerial photographs along the Cache Creek, California. Environ. Monit. Assess. 2006, 114, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Turner, W. Satellites: Make data freely accessible. Nature 2013, 498, 37. [Google Scholar] [CrossRef]
- Strand, H.; Höft, R.; Strittholt, J.; Miles, L.; Horning, N.; Fosnight, E.; Turner, W. Sourcebook on Remote Sensing and Biodiversity Indicators; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2007; Volume 32, 203p. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org/ (accessed on 10 June 2021).
- QGIS. A Free and Open Source Geographic Information System. Available online: www.qgis.org (accessed on 13 July 2021).
- GRASS Development Team. Geographic Resources Analysis Support System. Open Source Geospatial Foundation, USA. Available online: http://grass.osgeo.org (accessed on 13 July 2021).
- Blackburn, J.K. Evaluating the Spatial Ecology of Anthrax in North America: Examining Epidemiological Components across Multiple Geographic Scales Using a Gis-Based Approach. Ph.D. Thesis, Louisiana State University, Baton Rouge, LA, USA, 2006. [Google Scholar]
- Gruľová, D.; Koco, Š.; Muľová, A. Medziročná dynamika invázneho rastlinného druhu Solidago canadensis na vybraných lokalitách Prešovského okresu (Year-on-year dynamics of invasive plant species Solidago canadensis in selected localities of the Prešov district). Acta Univ. Prešoviensis 2017, 9, 57. [Google Scholar]
- Muľová, A. Využitie Geopriestorových Technológií Pri Mapovaní Priestorového Šírenia Inváznych Rastlín (The Use of Geospatial Technologies in Mapping the Spatial Spread of Invasive Plants). Bachelor’s Thesis, Fakulta Humanitných a Prírodných Vied Prešovskej Univerzity v Prešove, Prešov, Slovakia, 2018. [Google Scholar]
- Jakobs, G.; Weber, E.; Edwards, P.J. Introduced plants of the invasive Solidago gigantea (Asteraceae) are larger and grow denser than conspecifics in the native range. Divers. Distrib. 2004, 10, 11–19. [Google Scholar] [CrossRef]
- Holden, P.B.; Rebelo, A.J.; New, M.G. Mapping invasive alien trees in water towers: A combined approach using satellite data fusion, drone technology and expert engagement. Remote. Sens. Appl. Soc. Environ. 2021, 21, 100448. [Google Scholar] [CrossRef]
- Karlson, M.; Ostwald, M.; Reese, H.; Bazié, H.R.; Tankoano, B. Assessing the potential of multi-seasonal WorldView-2 imagery for mapping West African agroforestry tree species. Int. J. Appl. Earth Obs. Geoinf. 2016, 50, 80–88. [Google Scholar] [CrossRef]
- Michez, A.; Piegay, H.; Lisein, J.; Claessens, H.; Lejeune, P. Classification of riparian forest species and health condition using multi-temporal and hyperspatial imagery from unmanned aerial system. Environ. Monit. Assess. 2016, 188, 146. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.P.; Wagner, F.H.; Aragão, L.E.O.C.; Shimabukuro, Y.E.; de Souza Filho, C.R. Tree species classification in tropical forests using visible to shortwave infrared WorldView-3 images and texture analysis. ISPRS J. Photogramm. Remote Sens. 2019, 149, 119–131. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Škvarenina, J.; Tomlain, J.; Hrvol’, J.; Škvareninová, J. Occurence of dry and wet periods in altitudinal vegetation stages of West Carpathians in Slovakia: Time-Series Analysis 1951–2005. In Bioclimatology and Natural Hazards; Strelcová, K., Matyas, C., Kleidon, A., Lapin, M., Matejka, F., Blazenec, M., Škvarenina, J., Holecy, J., Eds.; Springer: Amsterdam, The Netherlands, 2009; pp. 97–106. [Google Scholar] [CrossRef]
- Lukasová, V.; Vido, J.; Škvareninová, J.; Bičárová, S.; Hlavatá, H.; Borsányi, P.; Škvarenina, J. Autumn phenological response of European Beech to summer drought and heat. Water 2020, 12, 2610. [Google Scholar] [CrossRef]
- Babálová, D.; Škvareninová, J.; Fazekaš, J.; Vyskot, I. The dynamics of the phenological development of four woody species in south-west and central Slovakia. Sustainability 2018, 10, 1497. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.H.; Schmid, B. Experimental demography of the old-field perennial Solidago altissima: The dynamic of the shoot population. J. Ecol. 1999, 87, 17–27. [Google Scholar] [CrossRef]
- Schulte to Bühne, H.; Pettorelli, N. Better Together: Integrating and Fusing Multispectral and Radar Satellite Imagery to Inform Biodiversity Monitoring, Ecological Research and Conservation Science. Methods Ecol. Evol. 2018, 9, 849–865. [Google Scholar] [CrossRef]
- Rapinel, S.; Hubert-Moy, L. One-Class Classification of Natural Vegetation Using Remote Sensing: A Review. Remote Sens. 2021, 13, 1892. [Google Scholar] [CrossRef]
- Kattenborn, T.; Lopatin, J.; Förster, M.; Braun, A.C.; Fassnacht, F.E. UAV Data as Alternative to Field Sampling to Map Woody Invasive Species Based on Combined Sentinel-1 and Sentinel-2 Data. Remote Sens. Environ. 2019, 227, 61–73. [Google Scholar] [CrossRef]
- Alexandridis, T.K.; Tamouridou, A.A.; Pantazi, X.E.; Lagopodi, A.L.; Kashefi, J.; Ovakoglou, G.; Polychronos, V.; Moshou, D. Novelty Detection Classifiers in Weed Mapping: Silybum marianum Detection on UAV Multispectral Images. Sensors 2017, 17, 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabat-Tomala, A.; Raczko, E.; Zagajewski, B. Comparison of Support Vector Machine and Random Forest Algorithms for Invasive and Expansive Species Classification using Airborne Hyperspectral Data. Remote Sens. 2020, 12, 516. [Google Scholar] [CrossRef] [Green Version]
- Davis, F.W.; Seo, C.; Zielinski, W.J. Regional variation in home-range-scale habitat models for fisher (Martes pennanti) in California. Ecol. Appl. 2007, 17, 2195–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, S78–S91. [Google Scholar] [CrossRef]
- Ustin, S.L.; Gitelson, A.A.; Jacquemoud, S.; Schaepman, M.; Asner, G.P.; Gamon, J.A.; Zarco-Tejada, P. Retrieval of foliar information about plant pigment systems from high resolution spectroscopy. Remote Sens. Environ. 2009, 113, S67–S77. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, M.R.; Numata, S.; Hosaka, T. Mapping an invasive goldenrod of Solidago altissima in urban landscape of Japan using multi-scale remote sensing and knowledge-based classification. Ecol. Indic. 2020, 111, 105975. [Google Scholar] [CrossRef]
- Ishii, J.; Washitani, I. Early detection of the invasive alien plant Solidago altissima in moist tall grassland using hyperspectral imagery. Int. J. Remote Sens. 2013, 34, 5926–5936. [Google Scholar] [CrossRef]
- Dorigo, W.; Lucieer, A.; Podobnikar, T.; Čarni, A. Mapping invasive Fallopia japonica by combined spectral, spatial, and temporal analysis of digital orthophotos. Int. J. Appl. Earth Obs. Geoinf. 2012, 19, 185–195. [Google Scholar] [CrossRef]
- Groeneveld, D.P.; Watson, R.P. Near-infrared discrimination of leafless saltcedar in wintertime Landsat TM. Int. J. Remote Sens. 2008, 29, 3577–3588. [Google Scholar] [CrossRef]
- Asner, G.P. Hyperspectral remote sensing of canopy chemistry, physiology and diversity in tropical rainforests. In Hyperspectral Remote Sensing of Tropical and Subtropical Forests; Kalacska, M., Sanchez-Azofeifa, G.A., Eds.; Taylor and Francis Group: Oxon, UK, 2008; pp. 261–288. [Google Scholar]
- Everitt, J.H.; Anderson, G.L.; Escobar, D.E.; Davis, M.R.; Spencer, N.R.; Andrascik, R.J. Use of remote sensing for detecting and mapping leafy spurge (Euphorbia esula). Weed Technol. 1995, 9, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Everitt, J.H.; Escobar, D.E.; Alaniz, M.A.; Davis, M.R.; Richerson, J.V. Using spatial information technologies to map Chinese tamarisk (Tamarix chinensis) infestations. Weed Sci. 1996, 44, 194–201. [Google Scholar] [CrossRef]
- US National Agriculture Imagery Program. Available online: http://www.fsa.usda.gov (accessed on 12 June 2021).
- Everitt, J.H.; Escobar, D.E.; Davis, M.R. Reflectance and image characteristics of selected noxious rangeland species. J. Range Manag. 2001, 54, A106–A120. [Google Scholar] [CrossRef]
- Huang, C.; Asner, G.P. Applications of Remote Sensing to Alien Invasive Plant Studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Date | Locality |
|---|---|---|
| 2016 | 27 and 28 September | Malý Šariš |
| 2016 | 25 September | Chmin. N. Ves |
| 2017 | 8 September | Malý Šariš |
| 2017 | 8 September | Chmin. N. Ves |
| 2018 | 27 August | Malý Šariš |
| 2018 | 27 August | Chmin. N. Ves |
| 2019 | 23 August | Malý Šariš |
| 2019 | 23 August | Chmin. N. Ves |
| locality | Malý Šariš | Chminianska Nová Ves | ||||||
|---|---|---|---|---|---|---|---|---|
| Year | 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 |
| Total number of shoots | 1258 | 1865 | 2523 | 3934 | 116 | 156 | 244 | 209 |
| Number of genets | 221 | 265 | 412 | 661 | 8 | 13 | 17 | 24 |
| Number of ramets | 37 | 161 | 180 | 324 | 0 | 3 | 0 | 7 |
| Minimum number of plants in genet | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Maximum number plants in genet | 32 | 48 | 50 | 42 | 61 | 85 | 100 | 41 |
| Percentage of increase/decrease in shoots | - | 48.30% | 35.30% | 55.90% | - | 34.50% | 56.40% | −14.30% |
| Chminianska Nová Ves | |
|---|---|
| Density | Acreage (m2) |
| (shoots numb./ha) | |
| <50 | 8144.4 |
| 50–100 | 1115.6 |
| 100–150 | 432.3 |
| 150–200 | 289.5 |
| >200 | 293.5 |
| Malý Šariš | |
| Density | Acreage (m2) |
| (shoots numb./ha) | |
| <1000 | 5327.3 |
| 1000–2000 | 1182.4 |
| 2000–3000 | 469.6 |
| 3000–4000 | 177.8 |
| 4000–5000 | 31.8 |
| 5000–6000 | 18.3 |
| >6000 | 13.3 |
| Reference | |||||
|---|---|---|---|---|---|
| Class | Trees and Shrubs | Grassland | Mown Grass Vegetation | Solidago spp. | |
| Classified | Trees and Shrubs | 0.066 | 0.066 | 0.033 | 0.017 |
| Grassland | 0.000 | 0.107 | 0.024 | 0.024 | |
| Mown Grass Vegetation | 0.036 | 0.125 | 0.054 | 0.018 | |
| Solidago spp. | 0.000 | 0.068 | 0.000 | 0.051 | |
| Total | 0.102 | 0.367 | 0.110 | 0.109 | |
| Standard Error | 0.037 | 0.054 | 0.039 | 0.038 | |
| Confidence Interval | 837.0 | 1224.0 | 897.0 | 862.0 | |
| 95% CI Area | 1641.0 | 2400.0 | 1758.0 | 1690.0 | |
| Producer’s Accuracy [%] | 64.9 | 15.8 | 48.5 | 46.7 | |
| User’s Accuracy [%] | 36.4 | 69.2 | 23.1 | 42.9 | |
| Kappa hat | 0.29 | 0.04 | 0.14 | 0.36 | |
| Overall Accuracy [%] | 27.8004 | ||||
| Kappa hat Classification | 0.1381 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koco, Š.; Dubravská, A.; Vilček, J.; Gruľová, D. Geospatial Approaches to Monitoring the Spread of Invasive Species of Solidago spp. Remote Sens. 2021, 13, 4787. https://doi.org/10.3390/rs13234787
Koco Š, Dubravská A, Vilček J, Gruľová D. Geospatial Approaches to Monitoring the Spread of Invasive Species of Solidago spp. Remote Sensing. 2021; 13(23):4787. https://doi.org/10.3390/rs13234787
Chicago/Turabian StyleKoco, Štefan, Anna Dubravská, Jozef Vilček, and Daniela Gruľová. 2021. "Geospatial Approaches to Monitoring the Spread of Invasive Species of Solidago spp." Remote Sensing 13, no. 23: 4787. https://doi.org/10.3390/rs13234787
APA StyleKoco, Š., Dubravská, A., Vilček, J., & Gruľová, D. (2021). Geospatial Approaches to Monitoring the Spread of Invasive Species of Solidago spp. Remote Sensing, 13(23), 4787. https://doi.org/10.3390/rs13234787