
Vegetation Growth Analysis of UNESCO World Heritage Hyrcanian Forests Using Multi-Sensor Optical Remote Sensing Data




Abstract
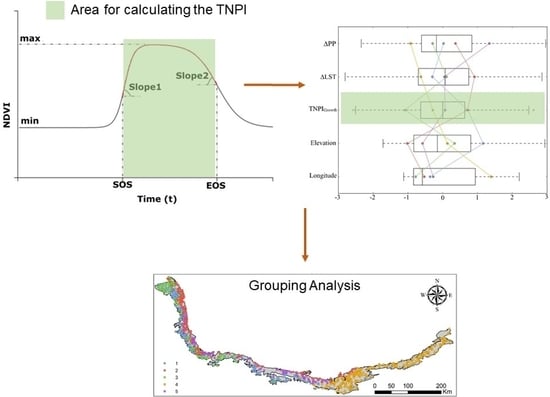
:
1. Introduction
2. Materials and Methods
2.1. Subsection
2.2. Data and Statistics
2.2.1. MODIS NDVI Data
2.2.2. Sentinel-2 Data
2.2.3. Elevation Data
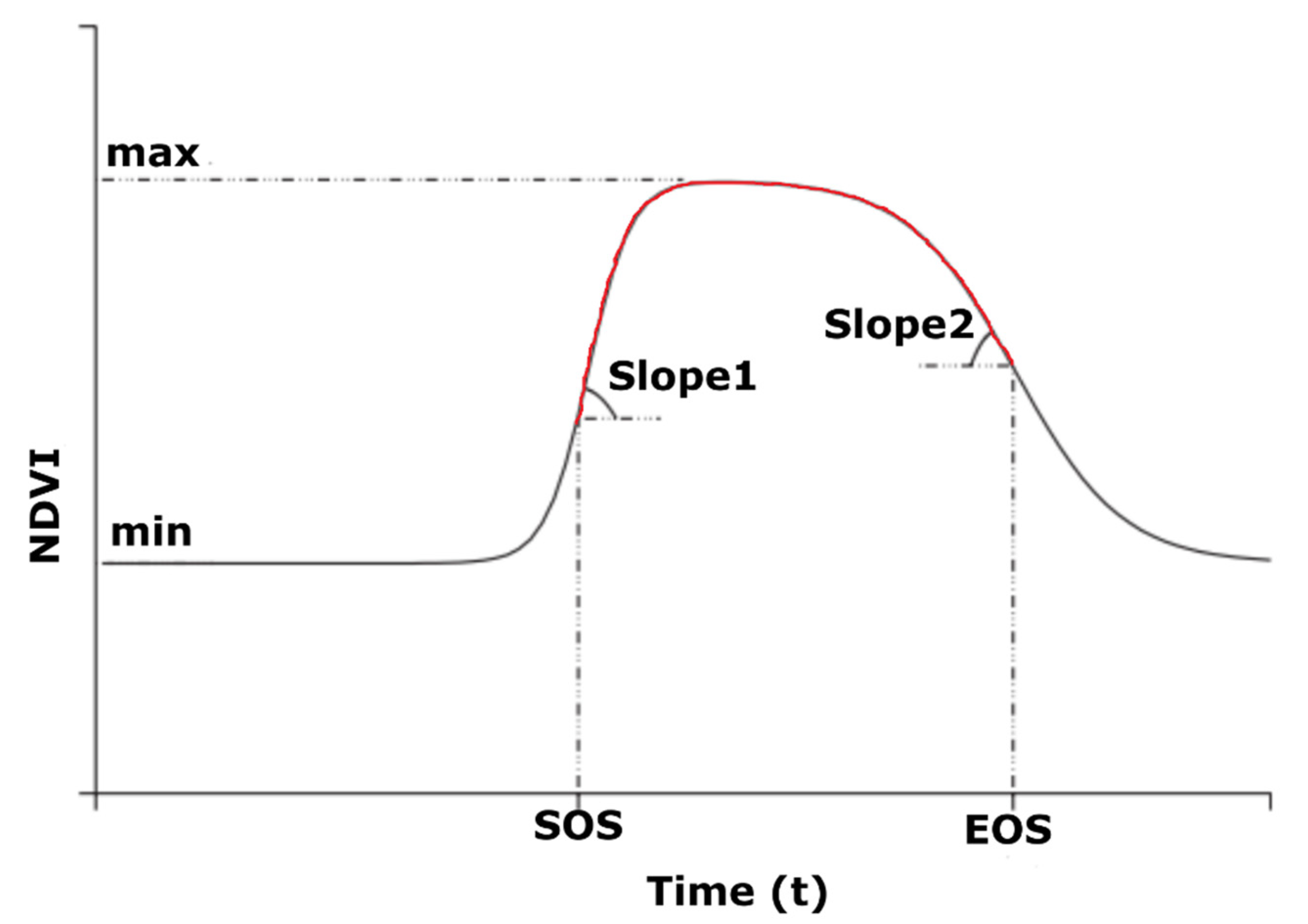
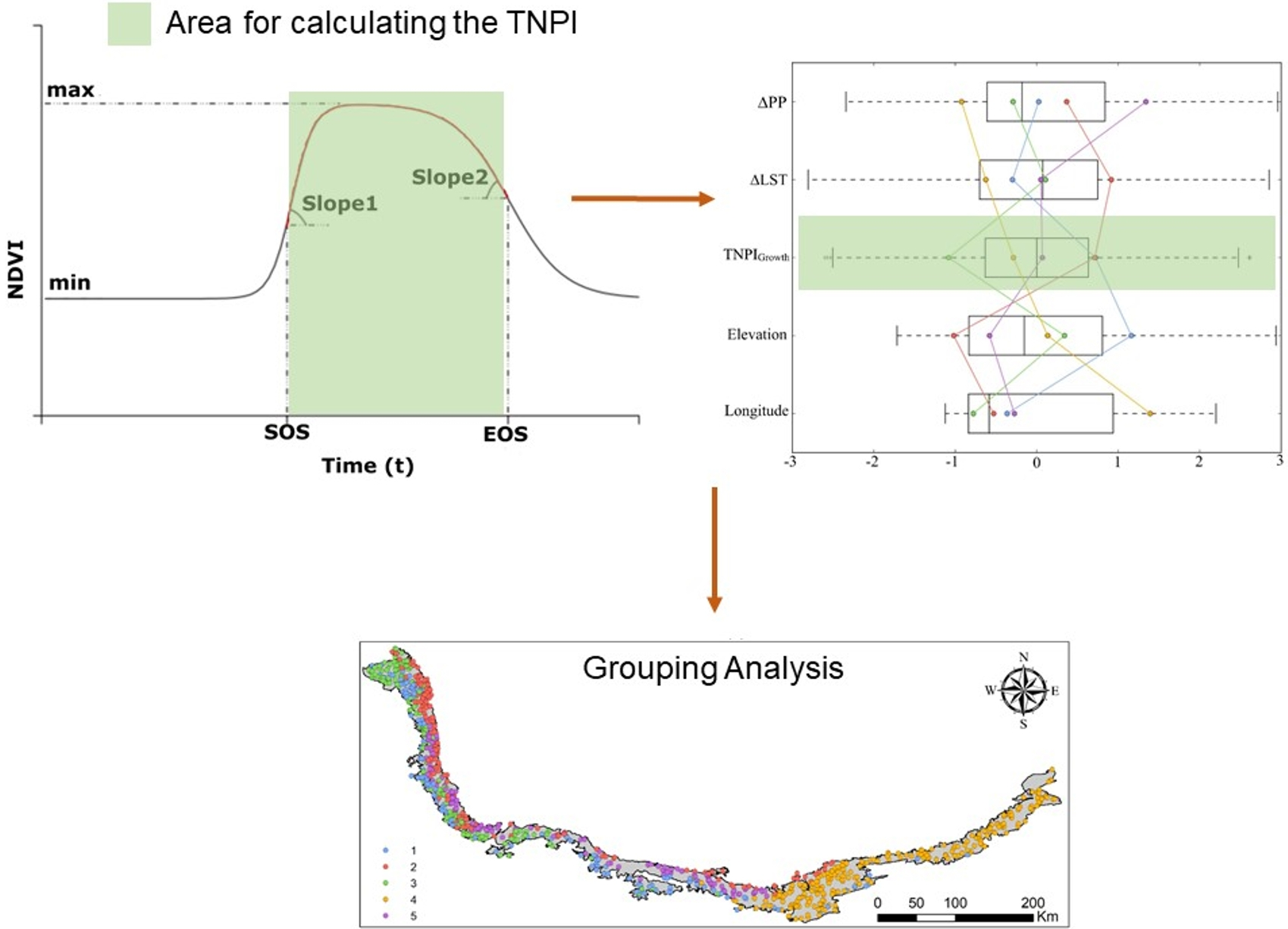
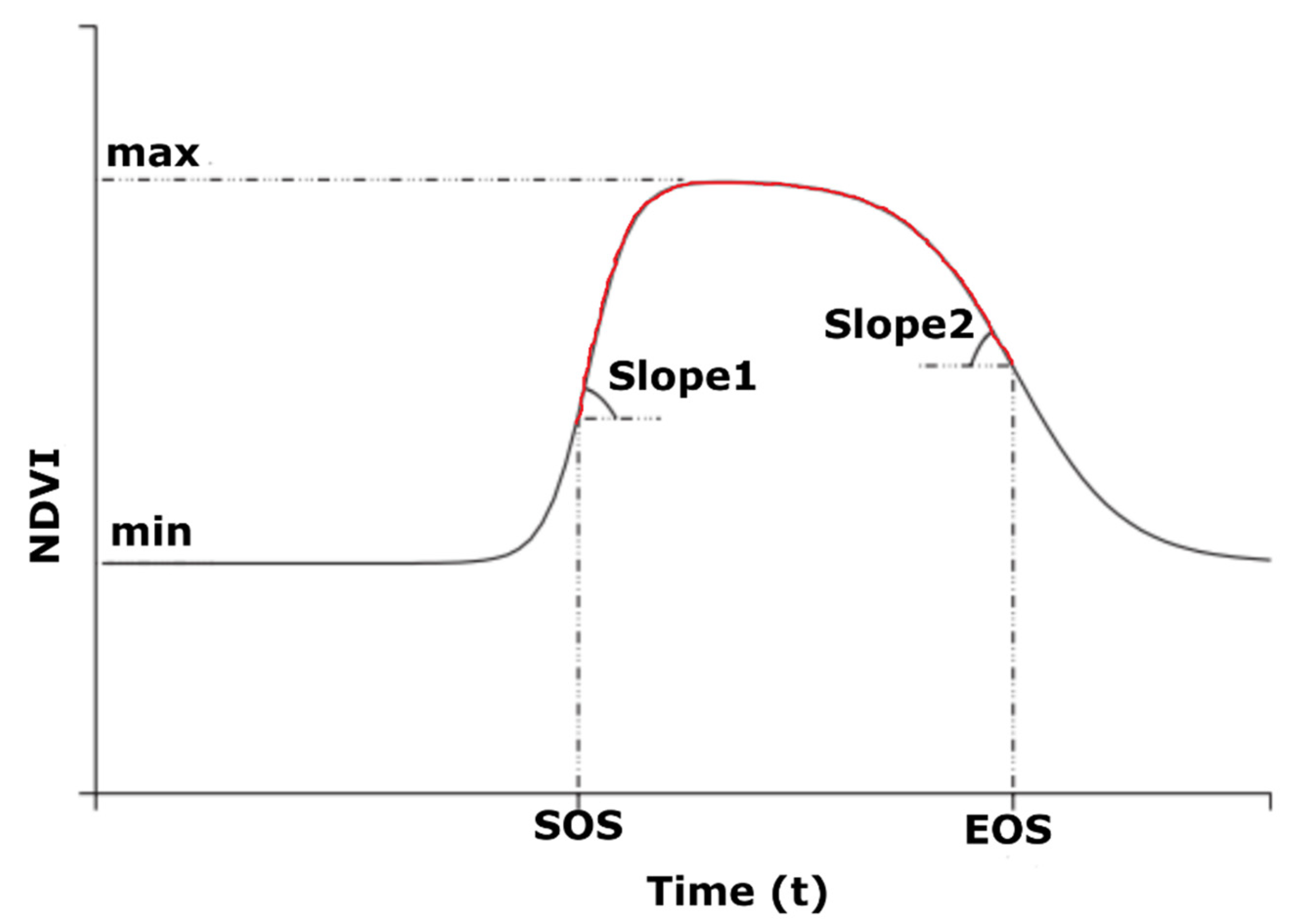
2.2.4. NDVI Curve Fitting
2.2.5. Computation of TNPI from Multi-Temporal NDVI
2.2.6. Precipitation Data
2.2.7. MODIS Land Surface Temperature Data
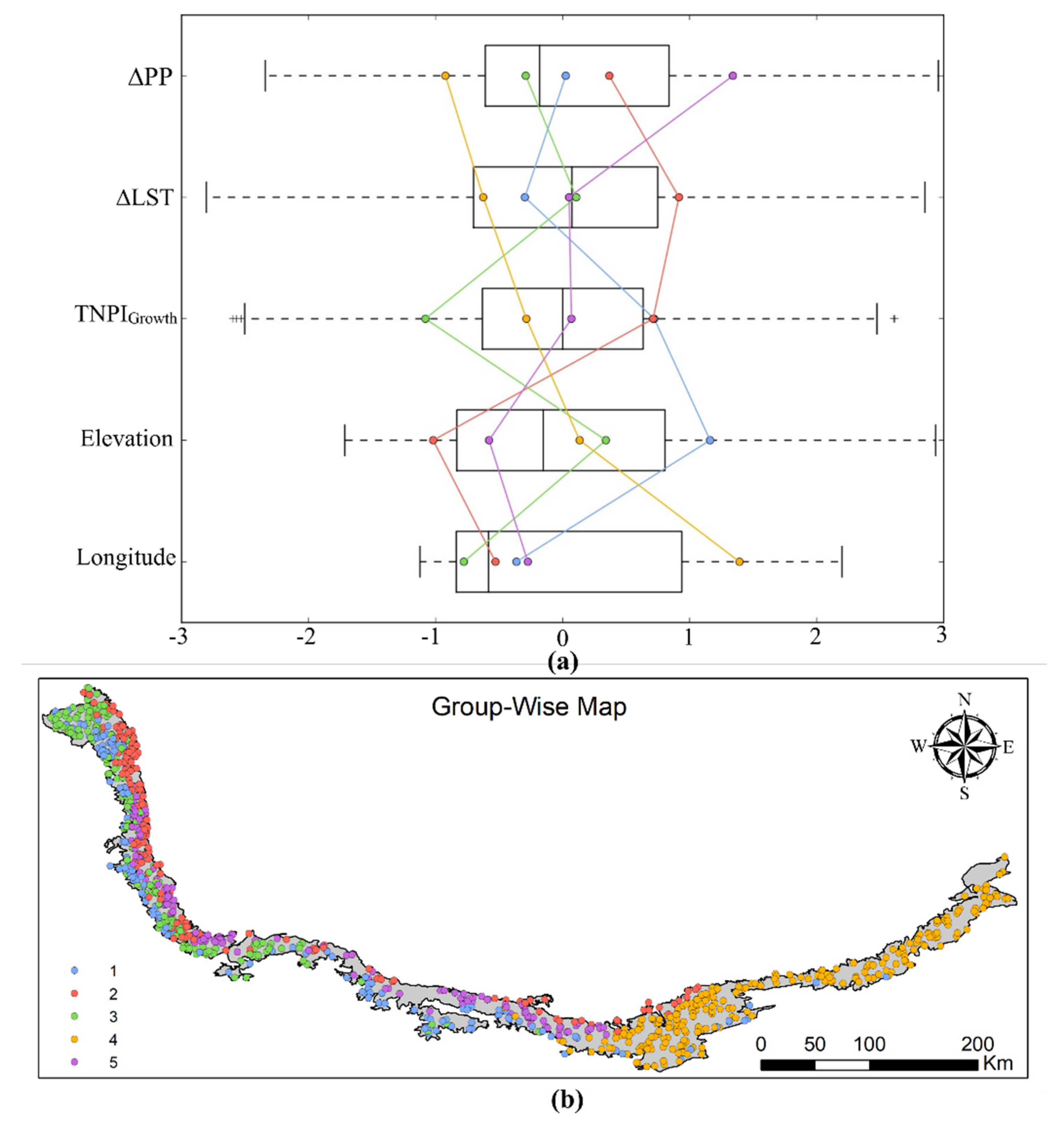
2.2.8. Grouping Analysis
2.2.9. Modelling Sentinel-2-Derived TNPI Growth with Environmental Factors
3. Results
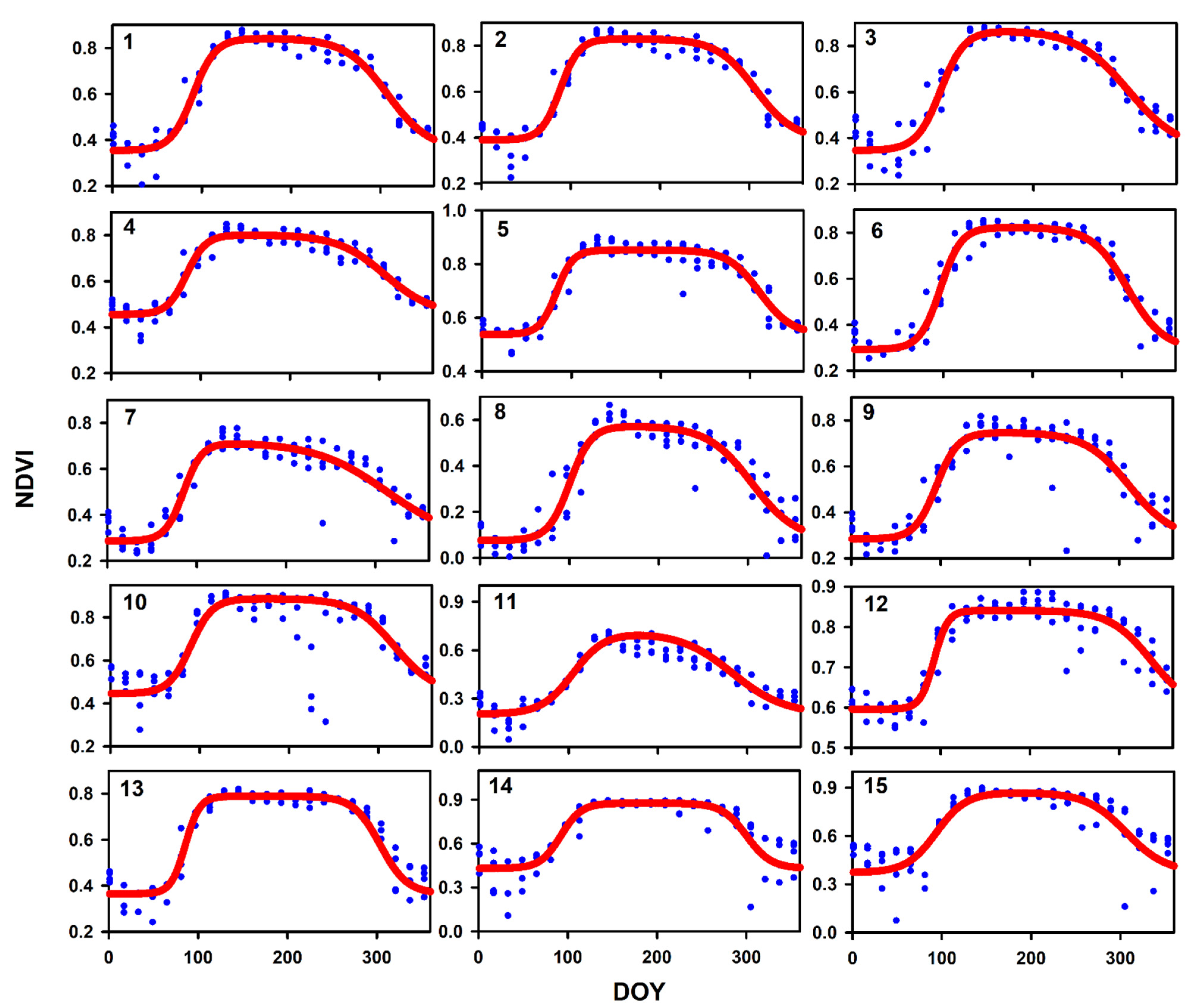
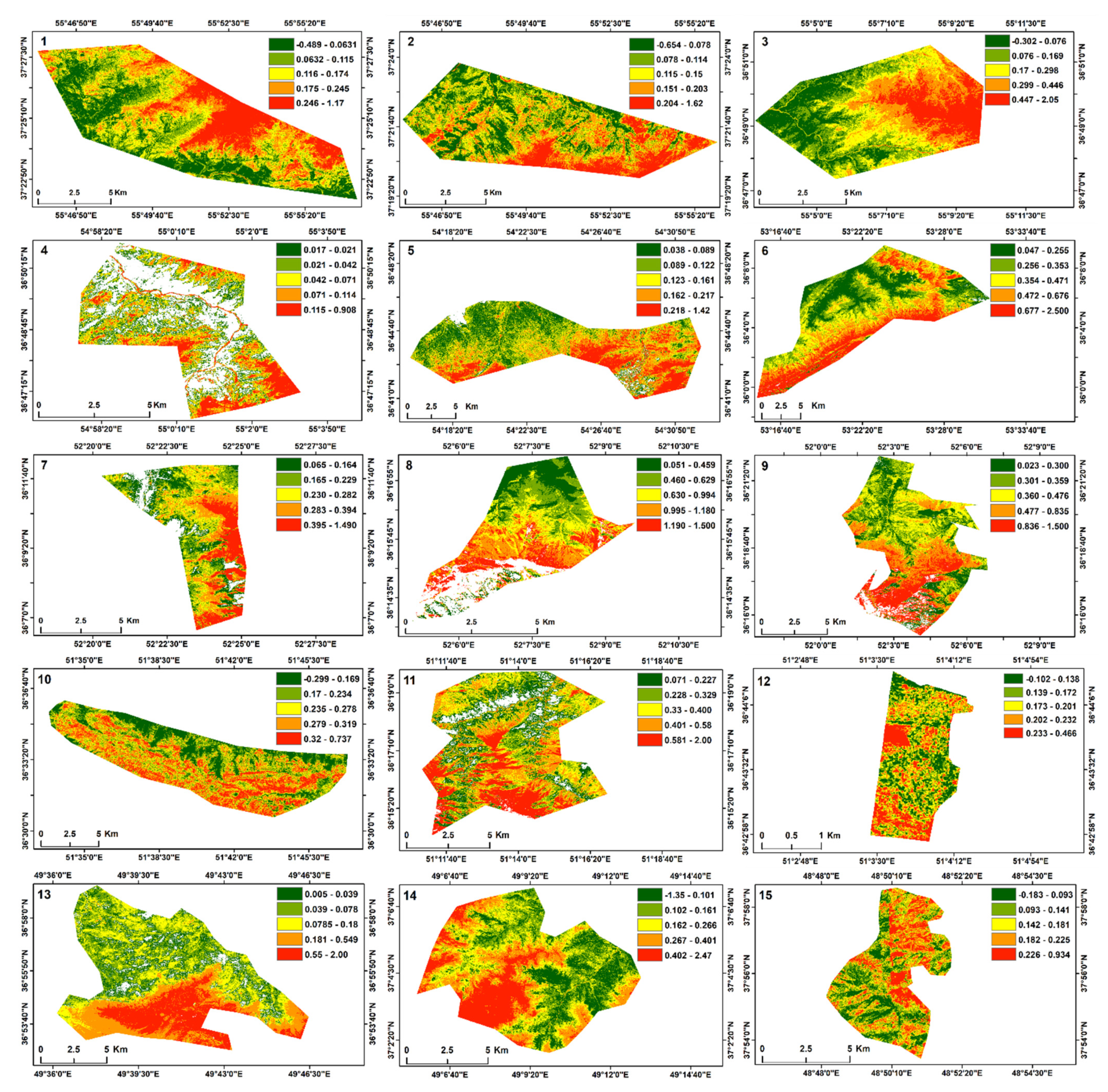
3.1. Phenology Patterns Derived from MODIS NDVI
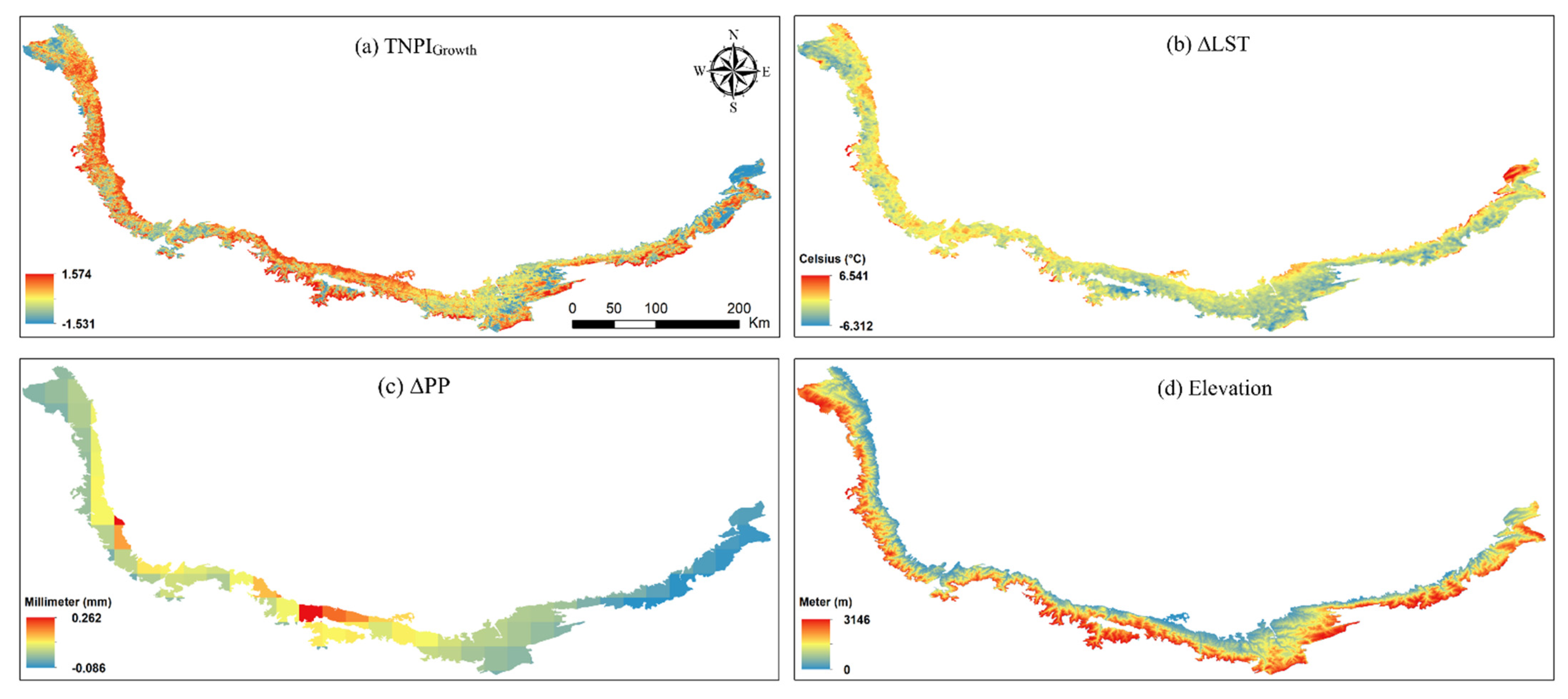
3.2. MODIS-Based Vegetation Growth Analysis and Effects of Environmental Factors
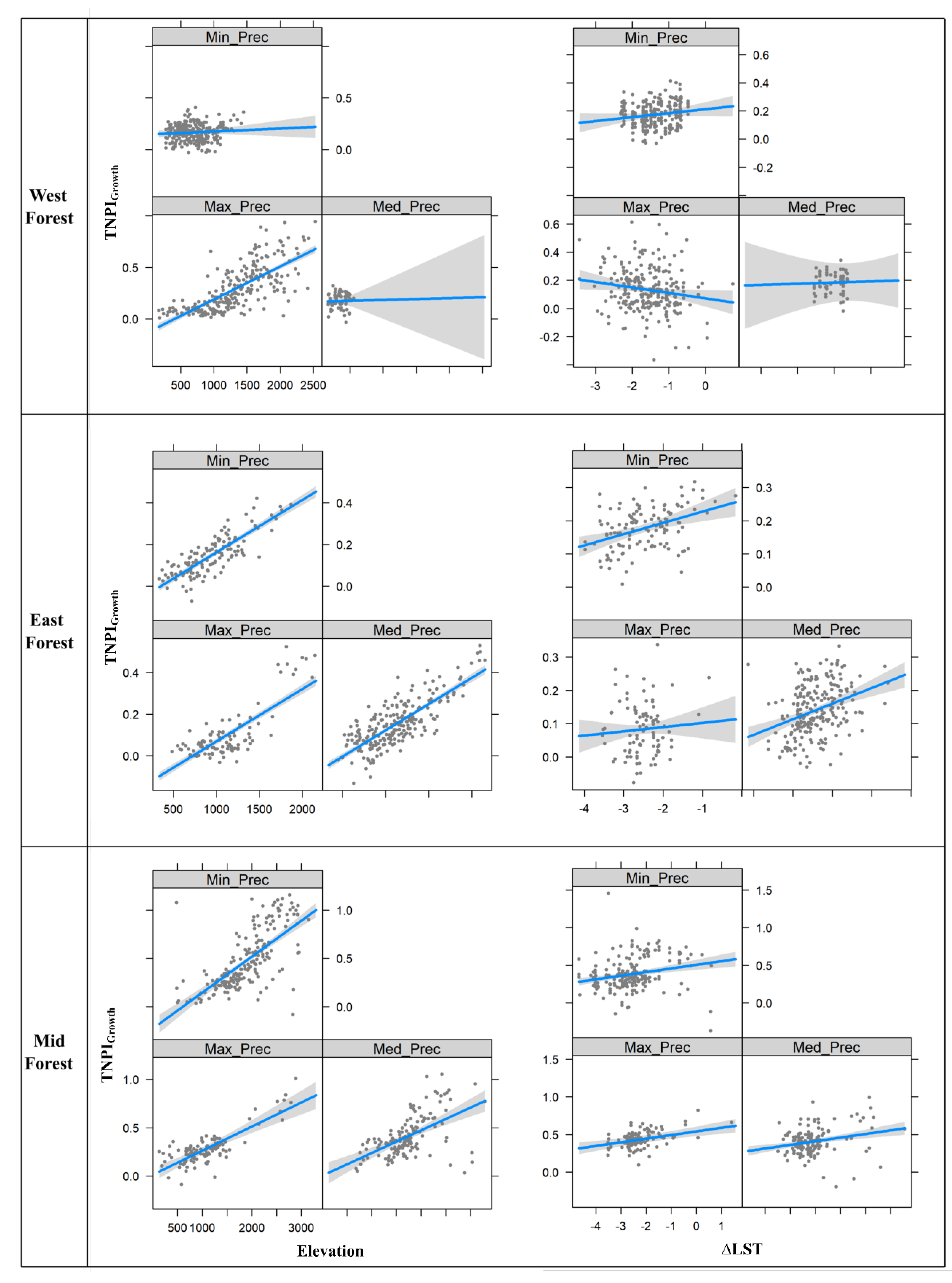
3.3. Multiple Regression Relationships between Sentinel-2-Derived TNPIGrowth and Environmental Variables
4. Discussion
4.1. The Observed Phenology Patterns
4.2. MODIS-Based Vegetation Growth Analysis
4.3. Multiple Regression Results on the Sentinel-2 Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Soudani, K.; le Maire, G.; Dufrêne, E.; François, C.; Delpierre, N.; Ulrich, E.; Cecchini, S. Evaluation of the onset of green-up in temperate deciduous broadleaf forests derived from Moderate Resolution Imaging Spectroradiometer (MODIS) data. Remote Sens. Environ. 2008, 112, 2643–2655. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Melaas, E.K.; Friedl, M.A.; Zhu, Z. Detecting interannual variation in deciduous broadleaf forest phenology using Landsat TM/ETM+ data. Remote Sens. Environ. 2013, 132, 176–185. [Google Scholar] [CrossRef]
- Lambert, J.; Drenou, C.; Denux, J.P.; Balent, G.; Cheret, V. Monitoring forest decline through remote sensing time series analysis. Giscience Remote Sens. 2013, 50, 437–457. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Glenn, E.P.; Huete, A.R.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: What vegetation indices can and cannot tell us about the landscape. Sensors 2008, 8, 2136–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen Trong, H.; Kappas, M. Land Cover and Forest Type Classification by Values of Vegetation Indices and Forest Structure of Tropical Lowland Forests in Central Vietnam. Int. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Khare, S.; Ghosh, S.K.; Latifi, H.; Vijay, S.; Dahms, T. Seasonal-based analysis of vegetation response to environmental variables in the mountainous forests of western himalaya using landsat 8 data. Int. J. Remote Sens. 2017, 38, 4418–4442. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, G.M.; Parthiban, S.; Thummalu, N.; Christy, A. Ndvi: Vegetation change detection using remote sensing and gis–A case study of Vellore District. Procedia Comput. Sci. 2015, 57, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Freitas, S.R.; Mello, M.C.S.; Cruz, C.B.M. Relationships between forest structure and vegetation indices in Atlantic rainforest. For. Ecol. Manag. 2005, 218, 353–362. [Google Scholar] [CrossRef]
- Rouse, W.; Haas, H.; Deering, W. Monitoring Vegetation Systems in the Great Plains With Erts. Proc. Third ERTS Symp. 1974, 351, 309–317. [Google Scholar]
- Ding, Y.; Zhao, K.; Zheng, X.; Jiang, T. Temporal dynamics of spatial heterogeneity over cropland quantified by time-series NDVI, near infrared and red reflectance of Landsat 8 OLI imagery. Int. J. Appl. Earth Obs. Geoinf. 2014, 30, 139–145. [Google Scholar] [CrossRef]
- Pettorelli, N. The Normalized Difference Vegetation Index, 1st ed.; Oxford University Press: New York, NY, USA, 2013; ISBN 9780199693160. [Google Scholar]
- Nordberg, M.L.; Evertson, J. Vegetation index differencing and linear regression for change detection in a Swedish mountain range using Landsat TM (R) and ETM+((R)) imagery. L. Degrad. Dev. 2005, 16, 139–149. [Google Scholar] [CrossRef]
- Lillesand, T.; Kiefer, R.W.; Chipman, J. Remote Sensing and Image Interpretation, 7th ed.; John Wiley & Sons: New York, NY, USA, 2015; ISBN 978-1-118-34328-9. [Google Scholar]
- Khare, S.; Rossi, S. Phenology analysis of moist decedous forest using time series Landsat-8 remote sensing data. In Proceedings of the 2019 IEEE International Workshop on Metrology for Agriculture and Forestry, MetroAgriFor, Portici, Italy, 24–26 October 2019; pp. 127–131. [Google Scholar]
- Khare, S.; Latifi, H.; Ghosh, K. Phenology analysis of forest vegetation to environmental variables during pre- And post-monsoon seasons in western Himalayan region of India. In Proceedings of the International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences—ISPRS Archives, 2016. XXIII ISPRS Congress, Prague, Czech Republic, 12–19 July 2016. [Google Scholar]
- Sabeti, H. Forests, Trees, and Shrubs of Iran; Iran University Science Technology Press: Tehran, Iran, 1994. [Google Scholar]
- Sagheb-Talebi, K.; Pourhashemi, M.; Sajedi, T. Forests of Iran: A Treasure from the Past, a Hope for the Future; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 9400773706. [Google Scholar]
- Zohary, M. Geobotanical Foundations of the Middle East; Swets & Zeitlinger: Stuttgart, Amsterdam, 1973; ISBN 9026501579. [Google Scholar]
- Ramezani, E.; Marvie Mohadjer, M.R.; Knapp, H.-D.; Ahmadi, H.; Joosten, H. The late-Holocene vegetation history of the Central Caspian (Hyrcanian) forests of northern Iran. Holocene 2008, 18, 307–321. [Google Scholar] [CrossRef]
- Leroy, S.A.G.; Kakroodi, A.A.; Kroonenberg, S.; Lahijani, H.K.; Alimohammadian, H.; Nigarov, A. Holocene vegetation history and sea level changes in the SE corner of the Caspian Sea: Relevance to SW Asia climate. Quat. Sci. Rev. 2013, 70, 28–47. [Google Scholar] [CrossRef] [Green Version]
- IUCN. World Heritage Nomination-IUCN Technical Evaluation for Hyrcanian Forests (Islamic Rerublic of Iran); Eastern Azarbaijan Province of Islamic Republic of Iran: 2019. Available online: https://whc.unesco.org/en/list/1584/ (accessed on 12 July 2021).
- Didan, K. MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid V006; NASA: 2015. Available online: http://dx.doi.org/10.5067/MODIS/MOD13Q1.006 (accessed on 12 July 2021).
- Ritter, P. A vector-based slope and aspect generation algorithm. Photogramm. Eng. Remote Sens. 1987, 53, 1109–1111. [Google Scholar]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Khare, S.; Drolet, G.; Sylvain, J.D.; Paré, M.C.; Rossi, S. Assessment of spatio-temporal patterns of black spruce bud phenology across Quebec based on MODIS-NDVI time series and field observations. Remote Sens. 2019, 11, 2745. [Google Scholar] [CrossRef] [Green Version]
- Škerlak, B.; Sprenger, M.; Wernli, H. A global climatology of stratosphere-troposphere exchange using the ERA-Interim data set from 1979 to 2011. Atmos. Chem. Phys. 2014, 14, 913–937. [Google Scholar] [CrossRef] [Green Version]
- QGIS Development Team QGIS Geographic Information System. 2021. Available online: https://qgis.org/en/site/ (accessed on 15 July 2021).
- ESRI, R. ArcGIS desktop: Release 10. Environ. Syst. Res. Inst. CA 2011. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-desktop/overview (accessed on 20 July 2021).
- ESRI. How Grouping Analysis Works—ArcGIS Pro|ArcGIS Desktop. Available online: https://pro.arcgis.com/en/pro-app/tool-reference/spatial-statistics/how-grouping-analysis-works.htm (accessed on 20 July 2021).
- Latifi, H.; Heurich, M.; Hartig, F.; Müller, J.; Krzystek, P.; Jehl, H.; Dech, S. Estimating over- and understorey canopy density of temperate mixed stands by airborne LiDAR data. Forestry 2016, 89, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Barton, K. MuMIn: Multi--Model Inference (R Package Version 1.13. 4). R--project. org/package= MuMIn. 2015. Available online: http://CRAN (accessed on 22 July 2021).
- Hurvich, C.; Tsai, C. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef]
- Kiapasha, K.; Darvishsefat, A.A.; Julien, Y.; Sobrino, J.A.; Zargham, N.; Attarod, P.; Schaepman, M.E. Trends in Phenological Parameters and Relationship Between Land Surface Phenology and Climate Data in the Hyrcanian Forests of Iran. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 4961–4970. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the European continent from NDVI3g data and MODIS time series. Remote Sens. 2014, 6, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Detsch, F.; Otte, I.; Appelhans, T.; Hemp, A.; Nauss, T. Seasonal and long-term vegetation dynamics from 1-km GIMMS-based NDVI time series at Mt. Kilimanjaro, Tanzania. Remote Sens. Environ. 2016, 178, 70–83. [Google Scholar] [CrossRef]
- Marshall, M.; Okuto, E.; Kang, Y.; Opiyo, E.; Ahmed, M. Global assessment of vegetation index and phenology lab (VIP) and global inventory modeling and mapping studies (GIMMS) version 3 products. Biogeosciences 2016, 13, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Galiano, V.F.; Dash, J.; Atkinson, P.M. Characterising the land surface phenology of Europe using decadal MERIS data. Remote Sens. 2015, 7, 9390–9409. [Google Scholar] [CrossRef] [Green Version]
- Moradi, H.; Naqinezhad, A.; Siadati, S.; Yousefi, Y.; Attar, F.; Etemad, V.; Reif, A. Elevational gradient and vegetation-environmental relationships in the central Hyrcanian forests of northern Iran. Nord. J. Bot. 2016, 34, 1–14. [Google Scholar] [CrossRef]
- Gholizadeh, H.; Naqinezhad, A.; Chytrý, M. Classification of the Hyrcanian forest vegetation, Northern Iran. Appl. Veg. Sci. 2020, 23, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Naqinezhad, A.; Zare-Maivan, H.; Gholizadeh, H. A floristic survey of the Hyrcanian forests in Northern Iran, using two lowland-mountain transects. J. For. Res. 2015, 26, 187–199. [Google Scholar] [CrossRef]
- Khalili, A. Precipitation patterns of central Elburz. Arch. für Meteorol. Geophys. Bioklimatol. B 1973, 21, 215–232. [Google Scholar] [CrossRef]
- Kahnamoie, M.H.M.; Bijker, W.; Sagheb–Talebi, K. The relation between annual diameter increment of Fagus orientalis and environmental factors (Hyrcanian forest). Improv. Silvic. Beech 2004, 76. Available online: https://www.iufro.org/download/file/5366/4507/11000-beech-proceedings-tehran-04_pdf/#page=79 (accessed on 13 July 2021).
- Noroozi, J.; Körner, C. A bioclimatic characterization of high elevation habitats in the Alborz mountains of Iran. Alp. Bot. 2018, 128, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, O.; Shirvani, Z.; Buchroithner, M.F. Spatiotemporal drought evaluation of Hyrcanian deciduous forests and semi-steppe rangelands using moderate resolution imaging spectroradiometer time series in Northeast Iran. L. Degrad. Dev. 2018, 29, 2525–2541. [Google Scholar] [CrossRef]
- Abdi, O.; Shirvani, Z.; Buchroithner, M.F. Forest drought-induced diversity of Hyrcanian individual-tree mortality affected by meteorological and hydrological droughts by analyzing moderate resolution imaging spectroradiometer products and spatial autoregressive models over northeast Iran. Agric. For. Meteorol. 2019, 275, 265–276. [Google Scholar] [CrossRef]
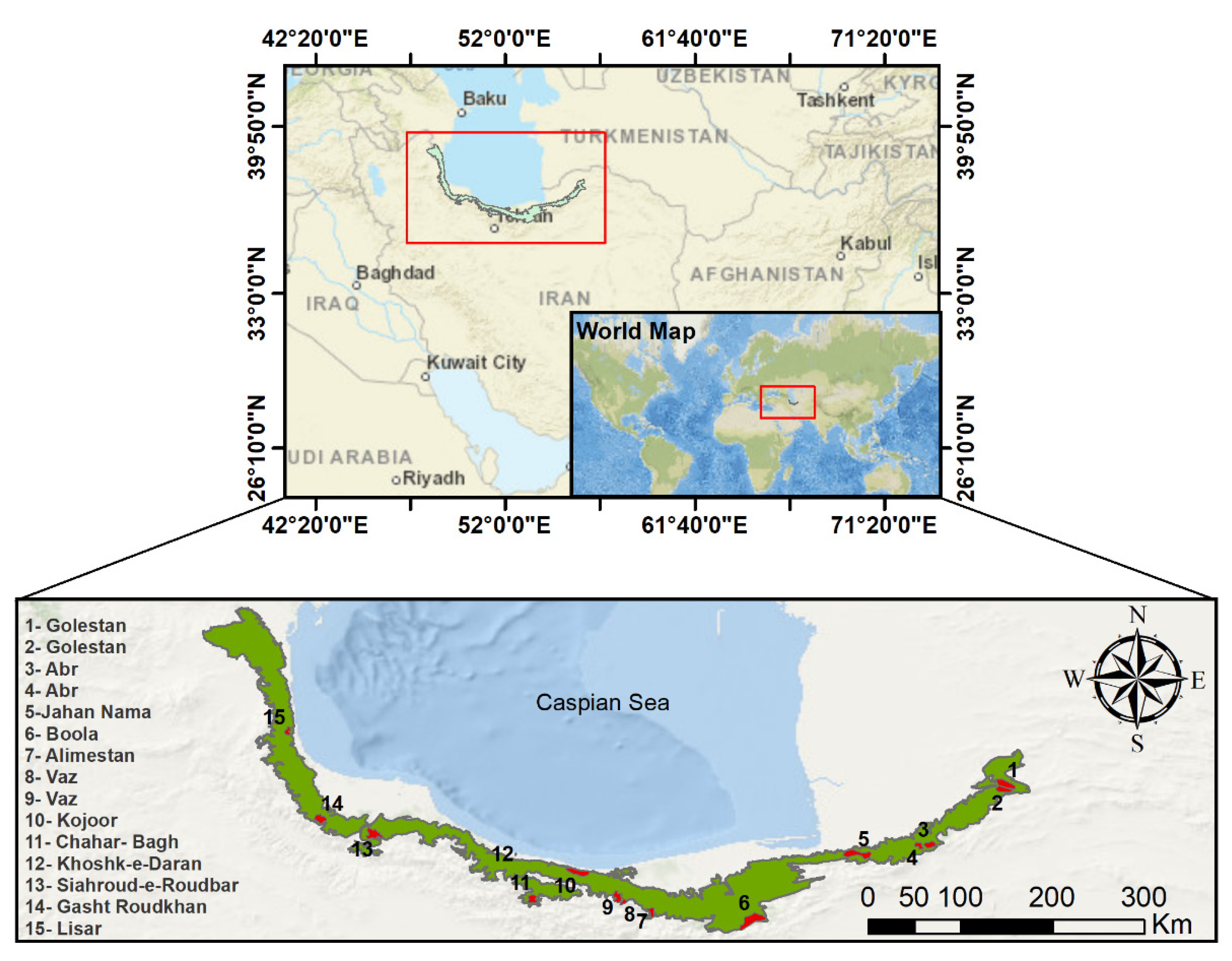
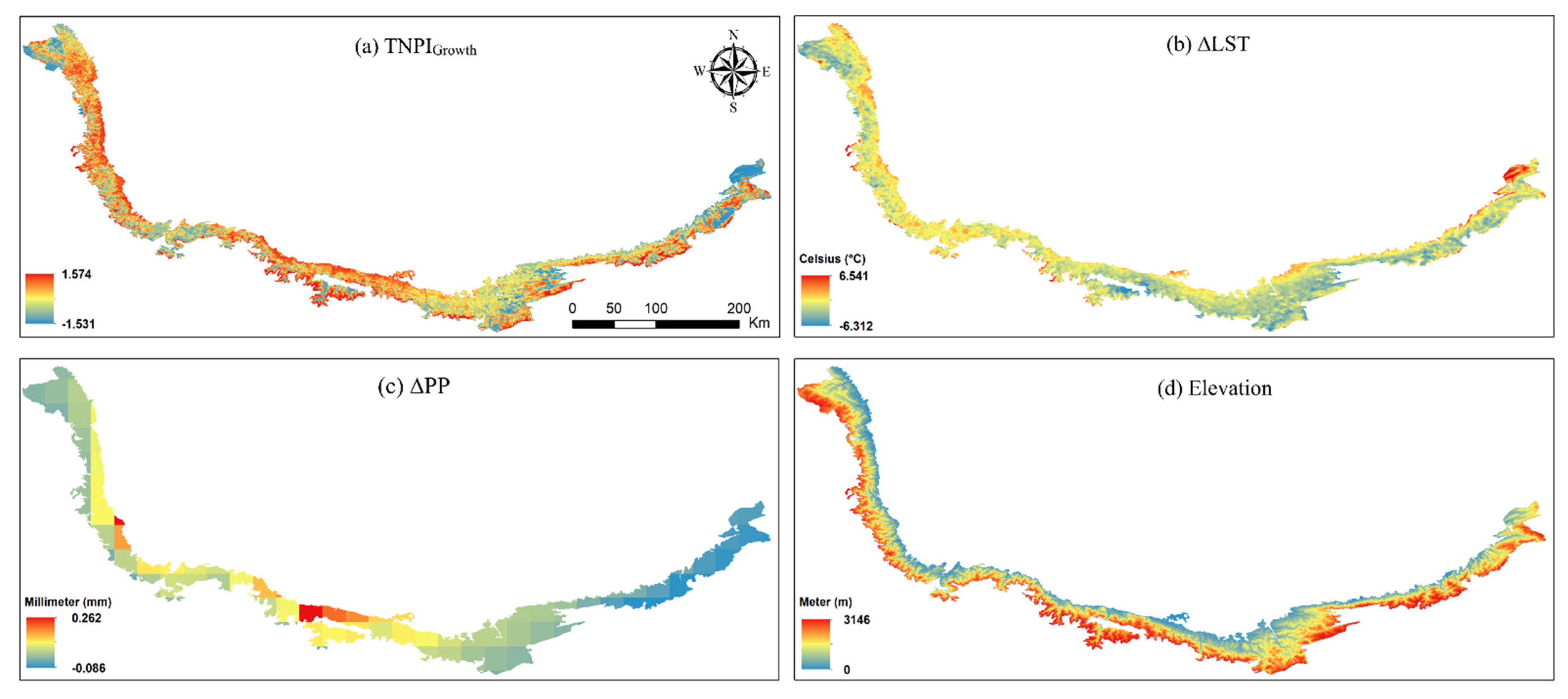
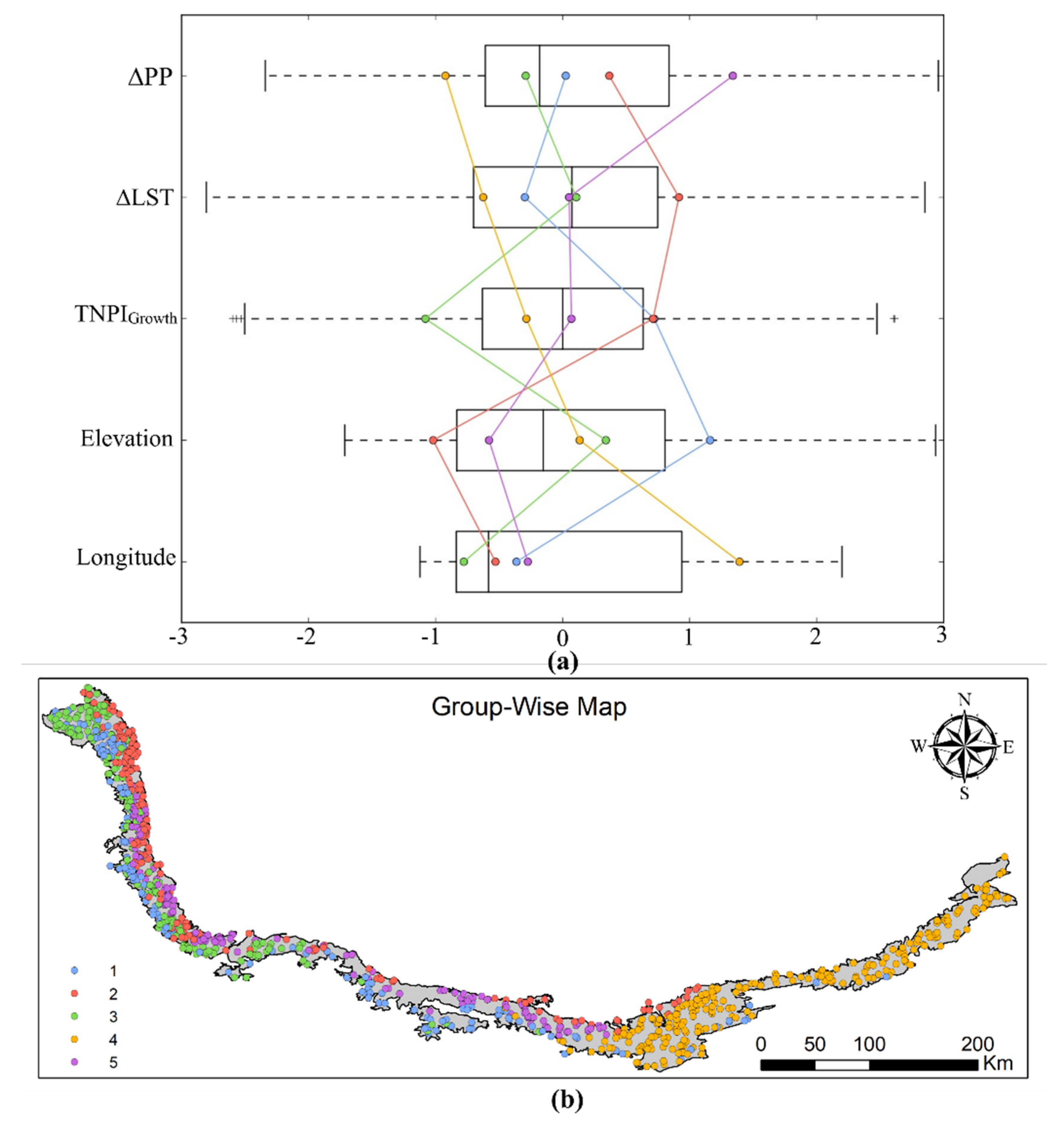
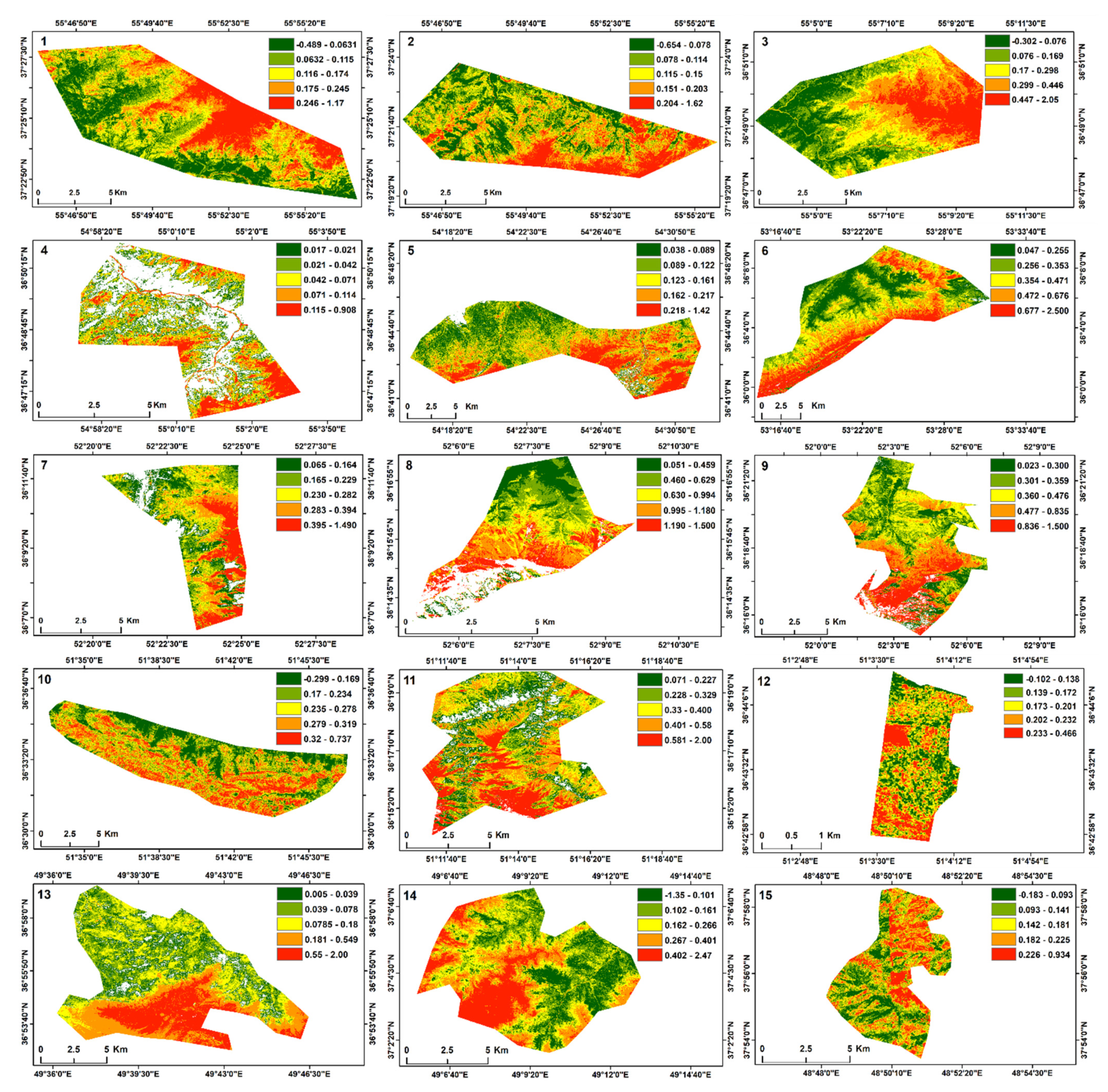
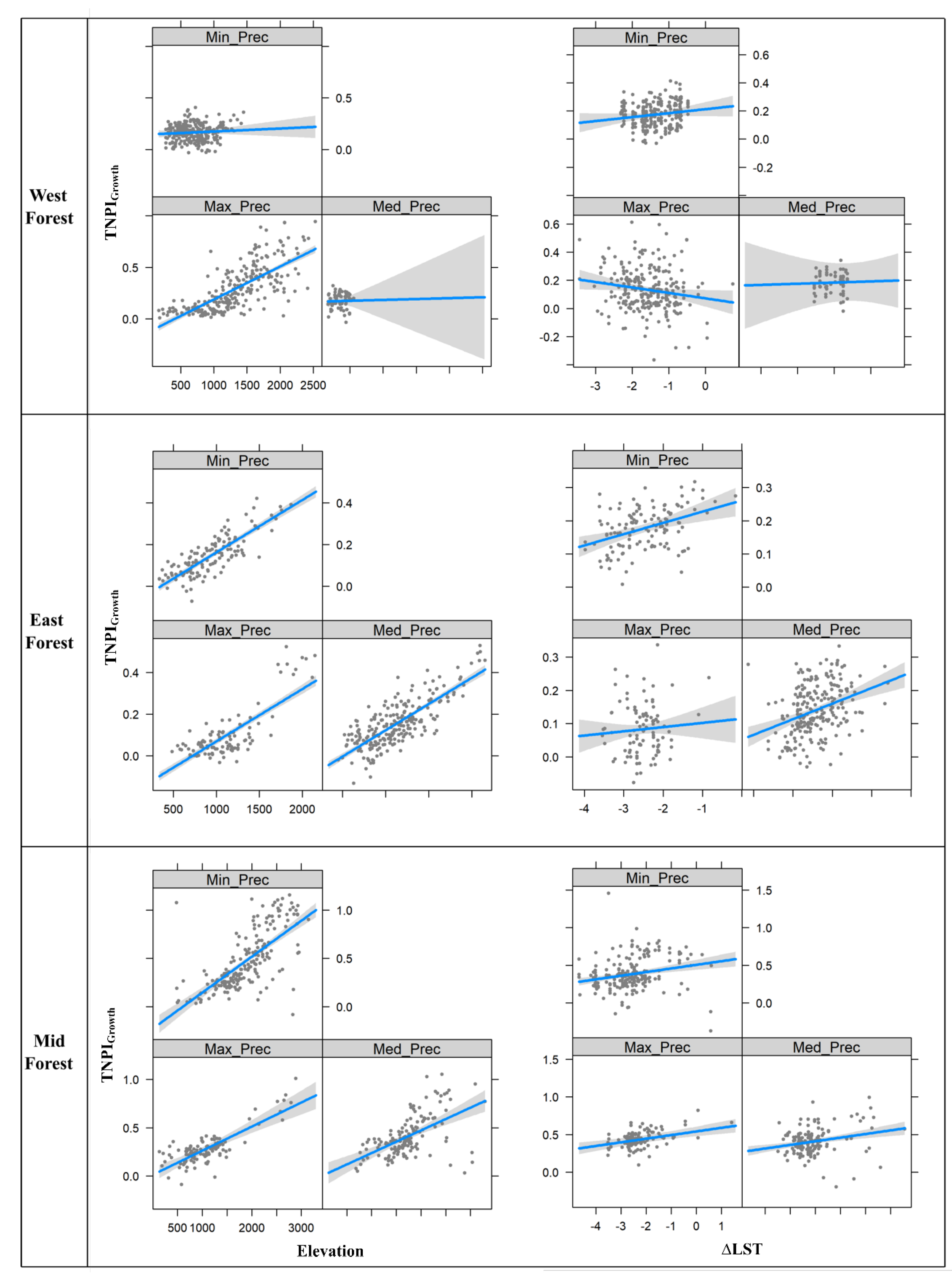

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
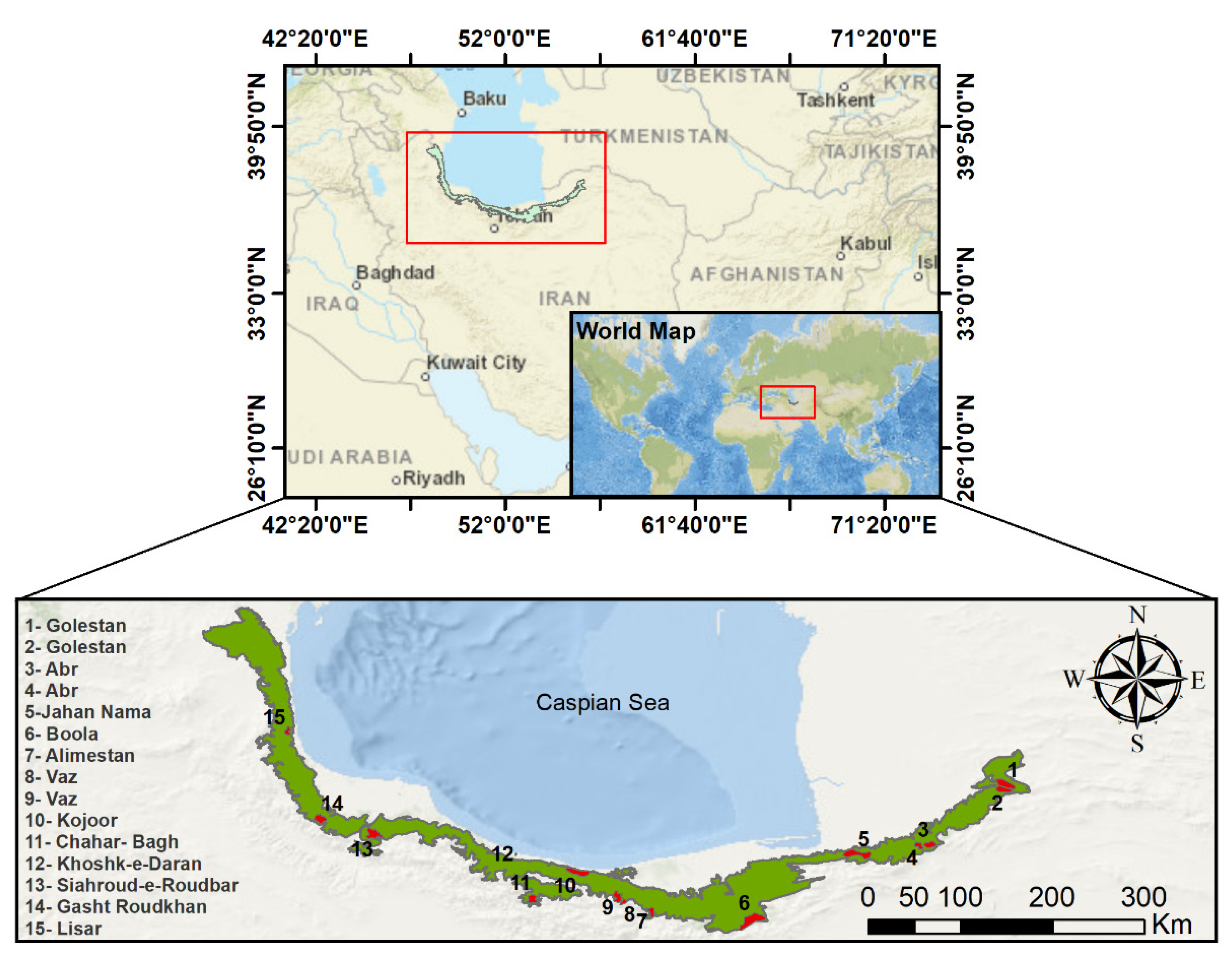
| Site | Coordinate | Annual Mean Temperature (°C) | Altitude (m) | Area (ha) |
|---|---|---|---|---|
| Golestan (North) | 55°43’27.4” E, 37°25’17.3” N | 12.40 | 1004 | 17,873.18 |
| Golestan (South) | 55°43’32.3” E, 37°20’26.4” N | 12.58 | 1034 | 10,658.08 |
| Abr (East) | 54°56’41.6” E, 36°48’45.3” N | 11.12 | 1728 | 6672.52 |
| Abr (West) | 55°6’3.4” E, 36°48’57.0” N | 12.04 | 1134 | 10,991.08 |
| Jahan Nama | 54°24’5.5” E, 36°39’55.0” N | −8.68 | 974 | 11,339.73 |
| Boola | 53°23’37.5” E, 36°5’55.8” N | 9.70 | 1641 | 17,516.47 |
| Alimestan | 52°24’14.2” E, 36°10’24.9” N | 10.78 | 1321 | 394.30 |
| Vaz (East) | 52°7’30.2” E, 36°16’44.8” N | 12.78 | 2713 | 2218.16 |
| Vaz (West) | 52°3’39.8” E, 36°18’26.9” N | 10.30 | 1690 | 4692.37 |
| Kojoor | 51°40’3.5” E, 36°32’45.7” N | 12.30 | 1086 | 14,891.80 |
| Chahar- Bagh | 51°13’1.7” E, 36°15’30.8” N | 9.52 | 1855 | 6886.44 |
| Khoshk-e-Daran | 51°3’50.3” E, 36°43’38.1” N | 13.64 | 8 | 214.47 |
| Siahroud-e-Roudbar | 49°40’19.3” E, 36°53’59.2” N | 12.08 | 988 | 11,197.40 |
| Gasht Roudkhan | 49°9’9.9” E, 37°3’56.0” N | 10.19 | 1280 | 10,541.13 |
| Lisar | 48°49’56.4” E, 37°56’8.0” N | 15.06 | 914 | 3397.61 |
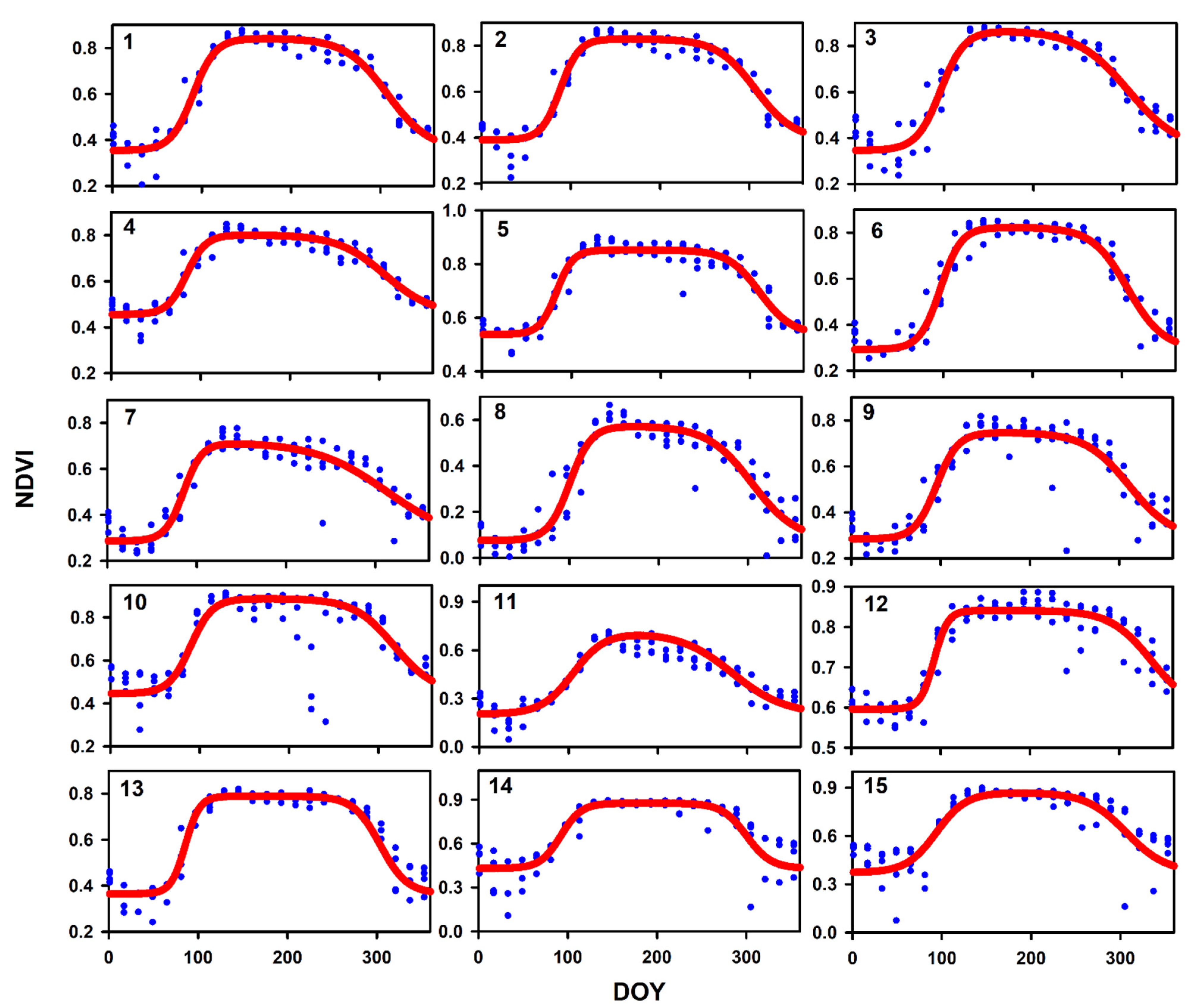
| Site | NDVImin | NDVImax | SOS | EOS | Slope 1 | Slope 2 | LOS | RMSE |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.354 | 0.842 | 90.38 | 308.08 | 0.080 | 0.043 | 217.70 | 0.0485 |
| 2 | 0.389 | 0.830 | 88.20 | 307.00 | 0.095 | 0.046 | 218.80 | 0.0451 |
| 3 | 0.346 | 0.868 | 96.09 | 306.04 | 0.074 | 0.034 | 209.94 | 0.0614 |
| 4 | 0.455 | 0.802 | 84.16 | 306.30 | 0.092 | 0.038 | 222.14 | 0.0374 |
| 5 | 0.538 | 0.852 | 83.17 | 310.70 | 0.107 | 0.056 | 227.52 | 0.0371 |
| 6 | 0.292 | 0.823 | 96.89 | 306.47 | 0.086 | 0.050 | 209.58 | 0.0565 |
| 7 | 0.286 | 0.716 | 85.25 | 312.78 | 0.096 | 0.025 | 227.53 | 0.0650 |
| 8 | 0.075 | 0.574 | 100.29 | 304.45 | 0.083 | 0.040 | 204.16 | 0.0710 |
| 9 | 0.284 | 0.748 | 95.32 | 311.17 | 0.083 | 0.041 | 215.86 | 0.0811 |
| 10 | 0.445 | 0.887 | 89.74 | 318.04 | 0.081 | 0.044 | 228.30 | 0.1095 |
| 11 | 0.202 | 0.713 | 105.16 | 283.35 | 0.056 | 0.034 | 178.19 | 0.0551 |
| 12 | 0.596 | 0.840 | 92.69 | 335.30 | 0.131 | 0.044 | 242.61 | 0.0357 |
| 13 | 0.364 | 0.789 | 85.65 | 303.98 | 0.125 | 0.067 | 218.33 | 0.0658 |
| 14 | 0.430 | 0.876 | 92.21 | 300.16 | 0.087 | 0.073 | 207.95 | 0.0982 |
| 15 | 0.372 | 0.870 | 93.95 | 306.48 | 0.060 | 0.045 | 212.53 | 0.0942 |
| Variable | Mean | Std. Dev | Min | Max | R2 |
|---|---|---|---|---|---|
| Longitude | 50.7655 | 2.3889 | 48.0830 | 56.0200 | 0.7203 |
| ΔPP | 39.6805 | 53.9826 | −86.7000 | 199.3500 | 0.5625 |
| Elevation | 946.1155 | 554.6567 | −5.0000 | 2575.0000 | 0.5309 |
| TNPIGrowth | 0.0126 | 0.0416 | −0.0954 | 0.1215 | 0.4424 |
| ΔLST | −2.0399 | 0.9361 | −4.6656 | 0.6304 | 0.2932 |
| Selected Model | Total No. of Samples | R2 | Adj.R2 | RMSE | p Value |
|---|---|---|---|---|---|
| 500 | 0.5267 | 0.522 | 0.116 | <0.0001 | |
| 500 | 0.6078 | 0.602 | 0.069 | <0.0001 | |
| 500 | 0.5678 | 0.5618 | 0.181 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khare, S.; Latifi, H.; Khare, S. Vegetation Growth Analysis of UNESCO World Heritage Hyrcanian Forests Using Multi-Sensor Optical Remote Sensing Data. Remote Sens. 2021, 13, 3965. https://doi.org/10.3390/rs13193965
Khare S, Latifi H, Khare S. Vegetation Growth Analysis of UNESCO World Heritage Hyrcanian Forests Using Multi-Sensor Optical Remote Sensing Data. Remote Sensing. 2021; 13(19):3965. https://doi.org/10.3390/rs13193965
Chicago/Turabian StyleKhare, Suyash, Hooman Latifi, and Siddhartha Khare. 2021. "Vegetation Growth Analysis of UNESCO World Heritage Hyrcanian Forests Using Multi-Sensor Optical Remote Sensing Data" Remote Sensing 13, no. 19: 3965. https://doi.org/10.3390/rs13193965
APA StyleKhare, S., Latifi, H., & Khare, S. (2021). Vegetation Growth Analysis of UNESCO World Heritage Hyrcanian Forests Using Multi-Sensor Optical Remote Sensing Data. Remote Sensing, 13(19), 3965. https://doi.org/10.3390/rs13193965