
Shifting of the Migration Route of White-Naped Crane (Antigone vipio) Due to Wetland Loss in China


 ,
,  and
and
Abstract
:1. Introduction
2. Data and Methods
2.1. White-Naped Crane Population Census
2.2. GPS Data Gathered in 1990s
2.3. GPS Data Collected in 2013–2014
2.4. Telemetric Data Processing
2.5. Habitat Utilization Modelling
2.6. Landcover Data
3. Results
3.1. White-Naped Crane Abundance at Different Sites along the Migratory Route
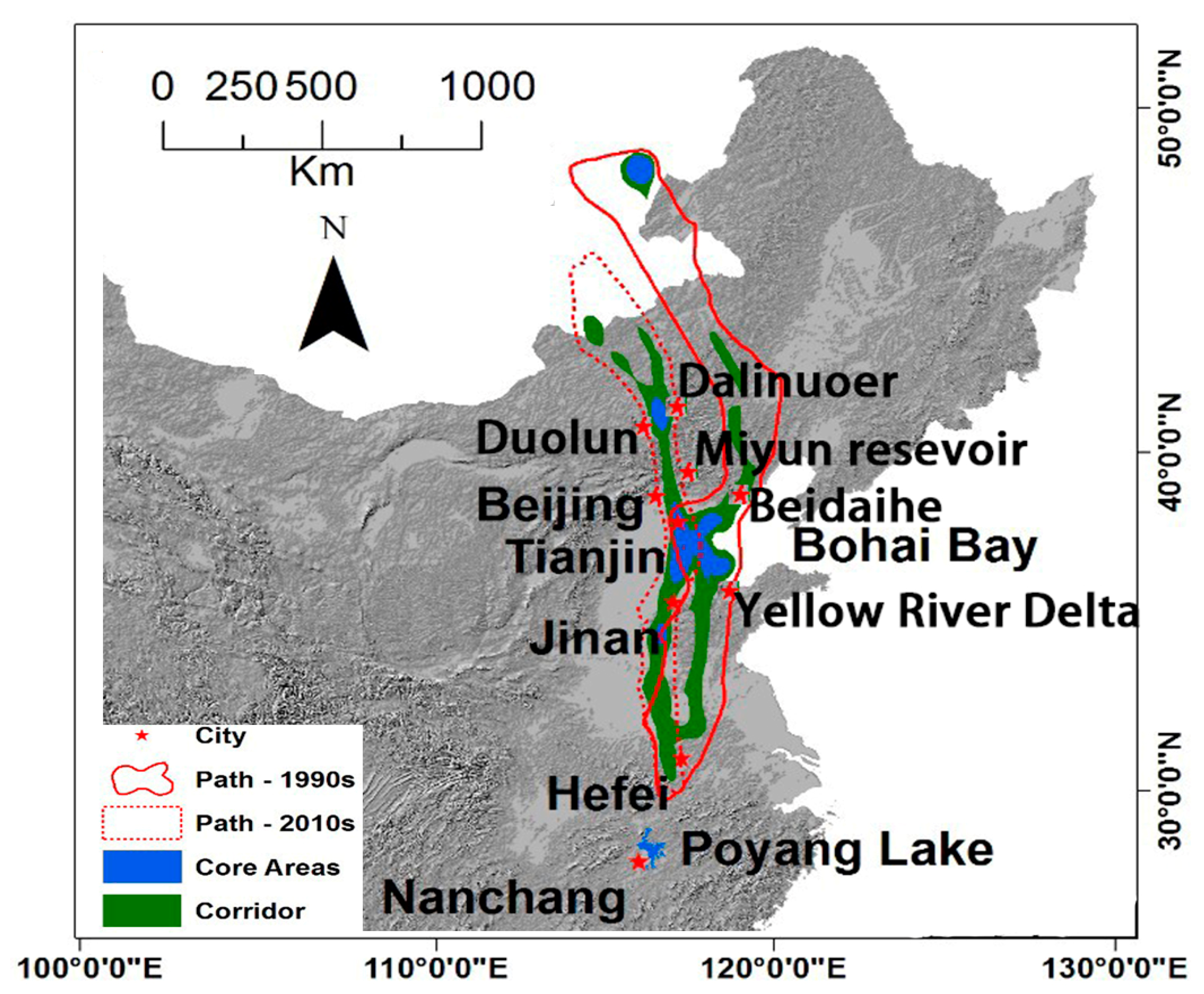
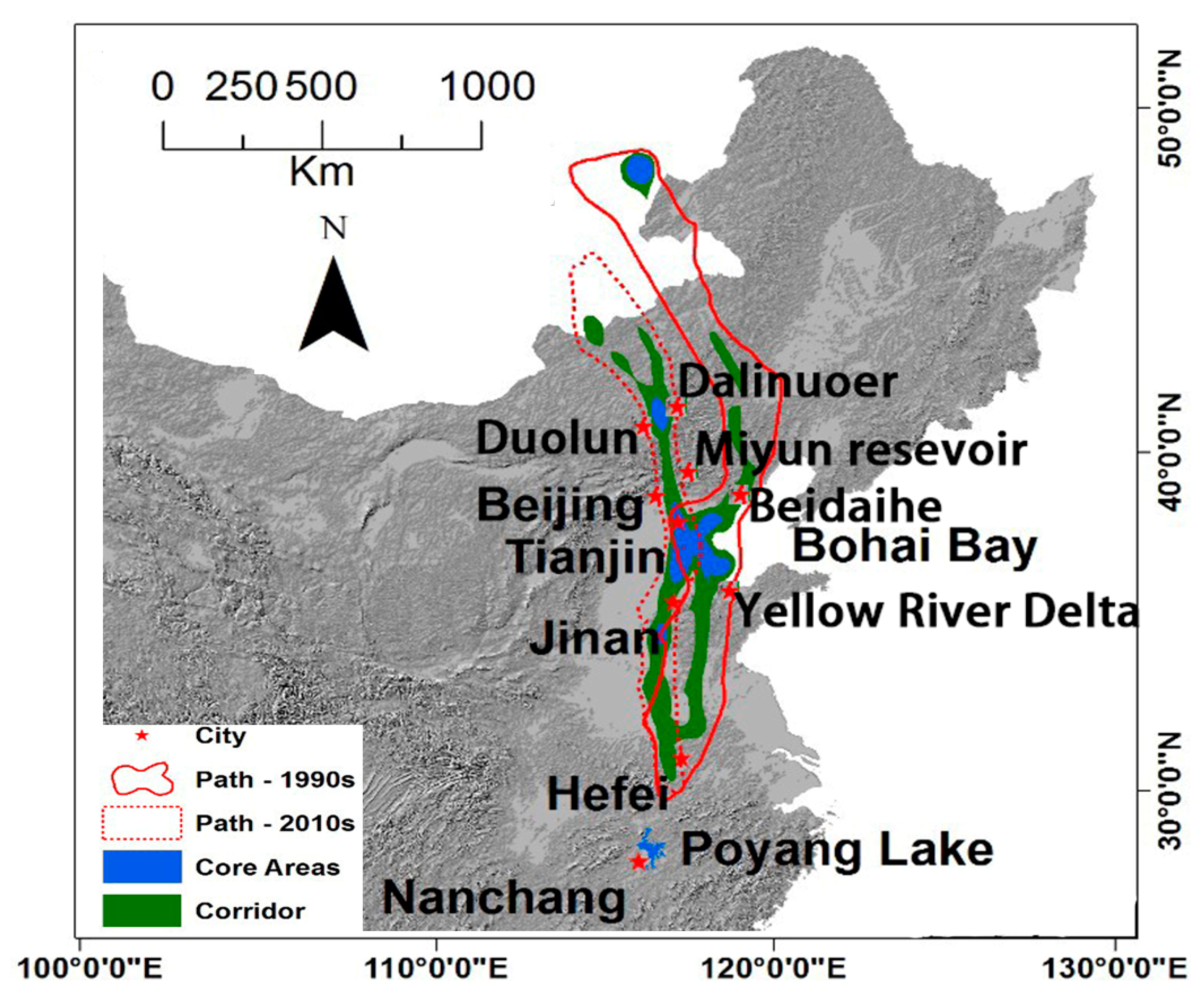
3.2. Migratory Paths at the Early 1990s and 2010s
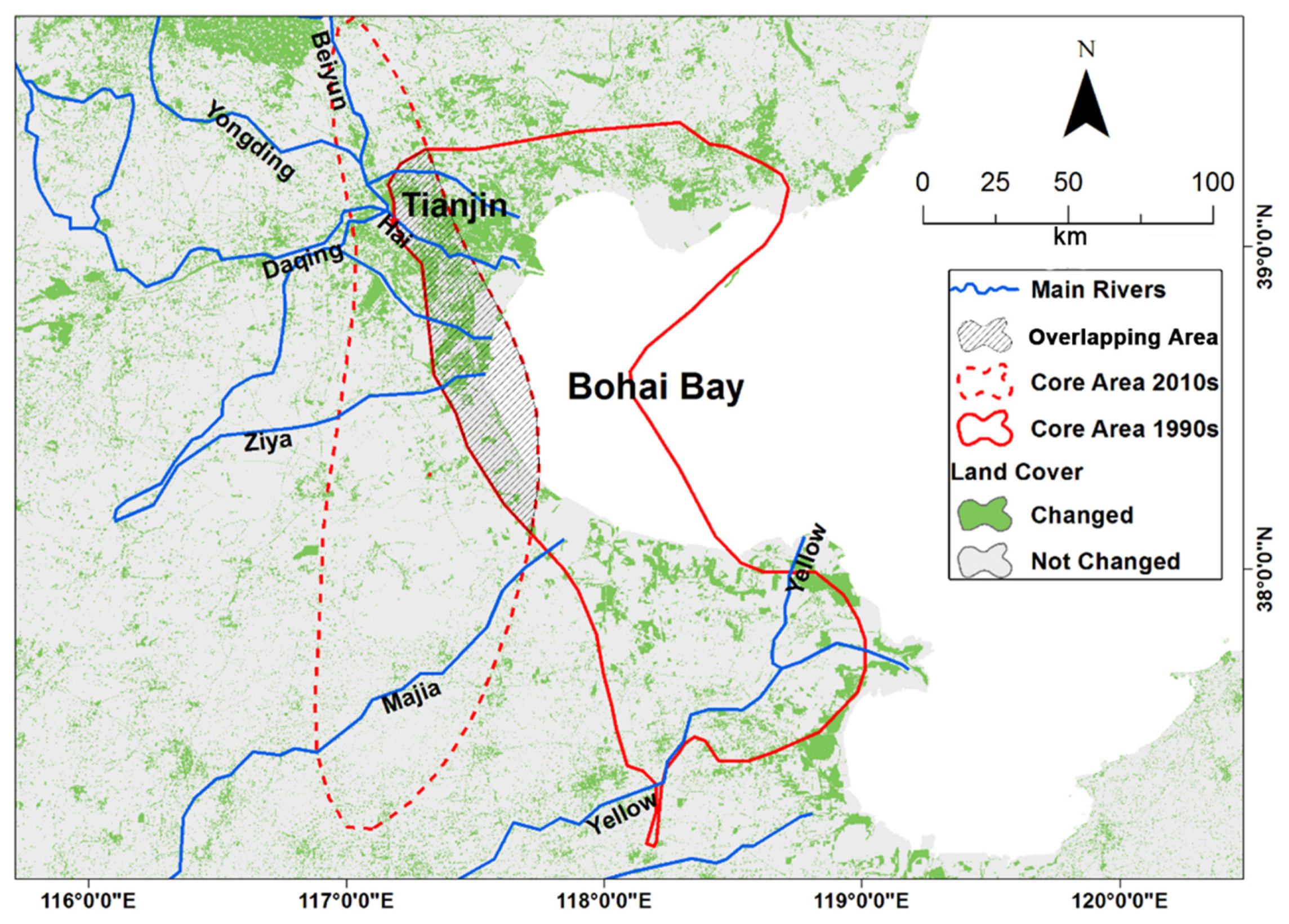
3.3. Landcover Changes within the Beidaihe Route
3.4. Landcover Changes within the Duolun Migratory Path
3.5. Landcover Change within the Bohai Bay Stopover Sites
3.6. Landcover Change within the Beidaihe Migratory Path North of Bohai Bay
4. Discussion
4.1. Overall Landcover Change Patterns and Their Impacts on White-Naped Crane Migration
4.2. Habitat Quality in Bohai Bay Region Could Be the Bottleneck of White-Naped Crane Conservation
4.3. The Possible Abandonment of the Beidaihe Route and Emergence of Duolun Route
4.4. Conservation Implications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kirby, J.S.; Stattersfield, A.J.; Butchart, S.H.; Evans, M.I.; Grimmett, R.F.; Jones, V.R.; O’Sullivan, J.; Tucker, G.M.; Newton, I. Key conservation issues for migratory land-and waterbird species on the world’s major flyways. Bird Conserv. Int. 2008, 18, S49–S73. [Google Scholar] [CrossRef] [Green Version]
- Studds, C.E.; Kendall, B.E.; Murray, N.J.; Wilson, H.B.; Rogers, D.I.; Clemens, R.S.; Gosbell, K.; Hassell, C.J.; Jessop, R.; Melville, D.S. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 2017, 8, 14895. [Google Scholar] [CrossRef] [PubMed]
- Wauchope, H.S.; Shaw, J.D.; Varpe, Ø.; Lappo, E.G.; Boertmann, D.; Lanctot, R.B.; Fuller, R.A. Rapid climate-driven loss of breeding habitat for Arctic migratory birds. Glob. Chang. Biol. 2017, 23, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Si, Y.; Takekawa, J.; Liu, Q.; Prins, H.H.; Yin, S.; Prosser, D.J.; Gong, P.; de Boer, W.F. A network approach to prioritize conservation efforts for migratory birds. Conserv. Biol. 2019, 34, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Si, Y.; Wang, Y.; Zhang, Y.; Prins, H.H.; Cao, L.; de Boer, W.F. Loss of functional connectivity in migration networks induces population decline in migratory birds. Ecol. Appl. 2019, 29, e1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runge, C.A.; Watson, J.E.; Butchart, S.H.; Hanson, J.O.; Possingham, H.P.; Fuller, R.A. Protected areas and global conservation of migratory birds. Science 2015, 350, 1255–1258. [Google Scholar] [CrossRef] [Green Version]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Baillie, J.E.; Hilton-Taylor, C.; Stuart, S.N. The 2004 IUCN Red List of Threatened Species. A Global Species Assessment; International Union for Conservation of Nature (IUCN): Gland, Switzerland; Cambridge, UK, 2004. [Google Scholar]
- Runge, C.A.; Martin, T.G.; Possingham, H.P.; Willis, S.G.; Fuller, R.A. Conserving mobile species. Front. Ecol. Environ. 2014, 12, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Trierweiler, C.; Klaassen, R.H.; Drent, R.H.; Exo, K.; Komdeur, J.; Bairlein, F.; Koks, B.J. Migratory connectivity and population-specific migration routes in a long-distance migratory bird. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132897. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Jia, Y.; Zuo, A.; Zeng, Q.; Shi, L.; Zhou, Y.; Zhang, H.; Lu, C.; Lei, G.; Wen, L. Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway. Int. J. Environ. Res. Public Health 2019, 16, 1147. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.D.; Hilmarsson, J.O.; Einarsson, O.; Walsh, A.J.; Boyd, H.; Kristiansen, J.N. Staging site fidelity of Greenland White-fronted Geese Anser albifrons flavirostris in Iceland. Bird Study 2002, 49, 42–49. [Google Scholar] [CrossRef]
- Gong, D.; Shi, P.; Wang, J. Daily precipitation changes in the semi-arid region over northern China. J. Arid Environ. 2004, 59, 771–784. [Google Scholar] [CrossRef]
- Clausen, K.K.; Madsen, J. Philopatry in a changing world: Response of pink-footed geese Anser brachyrhynchus to the loss of a key autumn staging area due to restoration of Filsø Lake, Denmark. J. Ornithol. 2016, 157, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsson, G.A.; Lindström, Å. Spring migration of Sanderlings Calidris alba through SW Iceland: Wherefrom and whereto? Ardea-Wageningen 1992, 80, 315. [Google Scholar]
- Catry, P.; Encarnação, V.; Araújo, A.; Fearon, P.; Fearon, A.; Armelin, M.; Delaloye, P. Are long-distance migrant passerines faithful to their stopover sites? J. Avian Biol. 2004, 35, 170–181. [Google Scholar] [CrossRef]
- Bairlein, F. Migratory birds under threat. Science 2016, 354, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Martins, I.S.; Kastner, T.; Plutzar, C.; Theurl, M.C.; Eisenmenger, N.; Huijbregts, M.A.; Wood, R.; Stadler, K.; Bruckner, M. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nat. Ecol. Evol. 2019, 3, 628. [Google Scholar] [CrossRef] [PubMed]
- Clausen, K.K.; Clausen, P.; Fælled, C.C.; Mouritsen, K.N. Energetic consequences of a major change in habitat use: Endangered Brent Geese Branta bernicla hrota losing their main food resource. IBIS 2012, 154, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Alerstam, T. Bird Migration; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Lei, J.; Jia, Y.; Wang, Y.; Lei, G.; Lu, C.; Saintilan, N.; Wen, L. Behavioural plasticity and trophic niche shift: How wintering geese respond to habitat alteration. Freshw. Biol. 2019, 64, 1183–1195. [Google Scholar] [CrossRef]
- Tedeschi, A.; Sorrenti, M.; Bottazzo, M.; Spagnesi, M.; Telletxea, I.; Ibàñez, R.; Tormen, N.; De Pascalis, F.; Guidolin, L.; Rubolini, D. Interindividual variation and consistency of migratory behavior in the Eurasian woodcock. Curr. Zool. 2019, 66, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Na, X.; Kong, B.; Wang, Z.; Jiang, H.; Yu, H.; Zhao, Z.; Li, X.; Liu, C.; Dale, P. Identifying wetland change in China’s Sanjiang Plain using remote sensing. Wetlands 2009, 29, 302. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Si, Y.; Ji, L.; Gong, P. Dynamic response of East Asian Greater White-fronted Geese to changes of environment during migration: Use of multi-temporal species distribution model. Ecol. Model. 2017, 360, 70–79. [Google Scholar] [CrossRef]
- Kanai, Y.; Ueta, M.; Germogenov, N.; Nagendran, M.; Mita, N.; Higuchi, H. Migration routes and important resting areas of Siberian cranes (Grus leucogeranus) between northeastern Siberia and China as revealed by satellite tracking. Biol. Conserv. 2002, 106, 339–346. [Google Scholar] [CrossRef]
- Higuchi, H.; Ozaki, K.; Fujita, G.; Minton, J.; Ueta, M.; Soma, M.; Mita, N. Satellite Tracking of White-naped Crane Migration and the Importance of the Korean Demilitarized Zone. Conserv. Biol. 1996, 10, 806–812. [Google Scholar] [CrossRef]
- Harris, J.; Liying, S.; Higuchi, H.; Ueta, M.; Zhengwang, Z.; Yanyun, Z.; Xijun, N. Migratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G. monacha, as determined by satellite tracking. Forktail 2000, 16, 93–100. [Google Scholar]
- IUCN. Redlist: White-Naped Crane; International Union for Conservation of Nature (IUCN): Gland, Switzerland; Cambridge, UK, 2018; Volume 18. [Google Scholar]
- Wetland International. Waterbird Population Estimates, 5th ed.; Wetlands International: Wageningen, The Netherlands, 2013; Volume 2013. [Google Scholar]
- Meine, C.D.; Archibald, G.W. The Cranes: Status Survey and Conservation Action Plan; International Union for Conservation of Nature (IUCN): Gland, Switzerland; Cambridge, UK, 1996; p. 294. [Google Scholar]
- Harris, J.; Mirande, C. A global overview of cranes: Status, threats and conservation priorities. Chin. Birds 2013, 4, 189–209. [Google Scholar] [CrossRef] [Green Version]
- Fujita, G.; Hong Liang, G.; Ueta, M.; Goroshko, O.; Krever, V.; Ozaki, K.; Mita, N.; Higuchi, H. Comparing areas of suitable habitats along travelled and possible shortest routes in migration of White-naped Cranes Grus vipio in East Asia. IBIS 2004, 146, 461–474. [Google Scholar] [CrossRef]
- Higuchi, H.; Pierre, J.P.; Krever, V.; Andronov, V.; Fujita, G.; Ozaki, K.; Goroshko, O.; Ueta, M.; Smirensky, S.; Mita, N. Using a Remote Technology in Conservation: Satellite Tracking White-Naped Cranes in Russia and Asia. Conserv. Biol. 2004, 18, 136–147. [Google Scholar] [CrossRef]
- Costa, D.P.; Robinson, P.W.; Arnould, J.P.; Harrison, A.; Simmons, S.E.; Hassrick, J.L.; Hoskins, A.J.; Kirkman, S.P.; Oosthuizen, H.; Villegas-Amtmann, S. Accuracy of ARGOS locations of pinnipeds at-sea estimated using Fastloc GPS. PLoS ONE 2010, 5, e8677. [Google Scholar] [CrossRef] [Green Version]
- Calenge, C. Home Range Estimation in R: The AdehabitatHR Package; Office National de la Classe et de la Faune Sauvage: Auffargis, France, 2011. [Google Scholar]
- Graves, T.A.; Waller, J.S. Understanding the causes of missed global positioning system telemetry fixes. J. Wildl. Manag. 2006, 70, 844–851. [Google Scholar] [CrossRef]
- Kuhn, C.E.; Johnson, D.S.; Ream, R.R.; Gelatt, T.S. Advances in the tracking of marine species: Using GPS locations to evaluate satellite track data and a continuous-time movement model. Mar. Ecol. Prog. Ser. 2009, 393, 97–109. [Google Scholar] [CrossRef]
- Tomkiewicz, S.M.; Fuller, M.R.; Kie, J.G.; Bates, K.K. Global positioning system and associated technologies in animal behaviour and ecological research. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2163–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhamou, S. Dynamic approach to space and habitat use based on biased random bridges. PLoS ONE 2011, 6, e14592. [Google Scholar] [CrossRef] [Green Version]
- Horne, J.S.; Garton, E.O.; Krone, S.M.; Lewis, J.S. Analyzing animal movements using Brownian bridges. Ecology 2007, 88, 2354–2363. [Google Scholar] [CrossRef]
- Benhamou, S.; Cornélis, D. Incorporating movement behavior and barriers to improve kernel home range space use estimates. J. Wildl. Manag. 2010, 74, 1353–1360. [Google Scholar] [CrossRef]
- Calenge, C. Analysis of Animal Movements in R: The AdehabitatLT Package; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Monsarrat, S.; Benhamou, S.; Sarrazin, F.; Bessa-Gomes, C.; Bouten, W.; Duriez, O. How predictability of feeding patches affects home range and foraging habitat selection in avian social scavengers? PLoS ONE 2013, 8, e53077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoener, T.W. Sizes of feeding territories among birds. Ecology 1968, 49, 123–141. [Google Scholar] [CrossRef]
- Rödder, D.; Engler, J.O. Quantitative metrics of overlaps in Grinnellian niches: Advances and possible drawbacks. Glob. Ecol. Biogeogr. 2011, 20, 915–927. [Google Scholar] [CrossRef]
- Jenks, G.F. The data model concept in statistical mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Samuel, M.D.; Pierce, D.J.; Garton, E.O. Identifying areas of concentrated use within the home range. J. Anim. Ecol. 1985, 54, 711–719. [Google Scholar] [CrossRef]
- van Wijk, R.E.; Bauer, S.; Schaub, M. Repeatability of individual migration routes, wintering sites, and timing in a long-distance migrant bird. Ecol. Evol. 2016, 6, 8679–8685. [Google Scholar] [CrossRef] [PubMed]
- Senner, N.R.; Verhoeven, M.A.; Abad-Gómez, J.M.; Alves, J.A.; Hooijmeijer, J.C.; Howison, R.A.; Kentie, R.; Loonstra, A.H.; Masero, J.A.; Rocha, A. High Migratory Survival and Highly Variable Migratory Behavior in Black-Tailed Godwits. Front. Ecol. Evol. 2019, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.; Boertmann, D. Animal behavioral adaptation to changing landscapes: Spring-staging geese habituate to wind farms. Landsc. Ecol. 2008, 23, 1007–1011. [Google Scholar] [CrossRef]
- Rogers, D.I.; Yang, H.; Hassell, C.J.; Boyle, A.N.; Rogers, K.G.; Chen, B.; Zhang, Z.; Piersma, T. Red Knots (Calidris canutus piersmai and C. c. rogersi) depend on a small threatened staging area in Bohai Bay, China. Emu Austral. Ornithol. 2010, 110, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, H.; Tamura, M.; Higuchi, H. Migration routes and important stopover sites of endangered oriental white storks (Ciconia boyciana), as revealed by satellite tracking. Mem. Natl. Inst. Polar Res. 2004, 58, 162–178. [Google Scholar]
- Murray, N.J.; Fuller, R.A. Protecting stopover habitat for migratory shorebirds in East Asia. J. Ornithol. 2015, 156, 217–225. [Google Scholar] [CrossRef]
- Iwamura, T.; Possingham, H.P.; Chadès, I.; Minton, C.; Murray, N.J.; Rogers, D.I.; Treml, E.A.; Fuller, R.A. Migratory connectivity magnifies the consequences of habitat loss from sea-level rise for shorebird populations. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130325. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.D.; Jabłoński, P.G.; Higuchi, H. Winter foraging of threatened cranes in the Demilitarized Zone of Korea: Behavioral evidence for the conservation importance of unplowed rice fields. Biol. Conserv. 2007, 138, 286–289. [Google Scholar] [CrossRef]
- Fujita, G.; Harris, J.; Bold, A.; Tseveenmyadag, N.; Chuluunbaatar, S. Habitat preference of Demoiselle and White-naped Cranes, Anthropoides virgo and Grus vipio, breeding in Mongolia. Future Cranes Wetl. 1994, 93–96. [Google Scholar]
- Kinzel, P.J.; Nelson, J.M.; Heckman, A.K. Response of sandhill crane (Grus canadensis) riverine roosting habitat to changes in stage and sandbar morphology. River Res. Appl. 2009, 25, 135–152. [Google Scholar] [CrossRef]
- Gullestad, N.; Owen, M.; Nugent, M.J. Numbers and distribution of Barnacle Geese Branta leucopsis on Norwegian staging islands and the importance of the staging area to the Svalbard population. Nor. Polarinst. Skr. 1984, 181, 57–65. [Google Scholar]
- Tulbure, M.G.; Broich, M.; Stehman, S.V.; Kommareddy, A. Surface water extent dynamics from three decades of seasonally continuous Landsat time series at subcontinental scale in a semi-arid region. Remote Sens. Environ. 2016, 178, 142–157. [Google Scholar] [CrossRef]
- Berthold, P. Control of Bird Migration; Springer: New York, NY, USA, 1996. [Google Scholar]
- Miller-Rushing, A.J.; Høye, T.T.; Inouye, D.W.; Post, E. The effects of phenological mismatches on demography. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3177–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Z.; Gong, P.; Cheng, X.; Guo, J.; Wang, L.; Huang, H.; Shen, S.; Wu, Y.; Wang, X.; Wang, X. Geographical characteristics of China’s wetlands derived from remotely sensed data. Sci. China Ser. D Earth Sci. 2009, 52, 723–738. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, B.; Zhang, S.; Li, X.; Liu, D.; Song, K.; Li, J.; Li, F.; Duan, H. Changes of land use and of ecosystem service values in Sanjiang Plain, Northeast China. Environ. Monit. Assess. 2006, 112, 69–91. [Google Scholar] [CrossRef]
- Zhou, D.; Gong, H.; Wang, Y.; Khan, S.; Zhao, K. Driving forces for the marsh wetland degradation in the Honghe National Nature Reserve in Sanjiang Plain, Northeast China. Environ. Model. Assess. 2009, 14, 101–111. [Google Scholar] [CrossRef]
- Seidler, R.G.; Long, R.A.; Berger, J.; Bergen, S.; Beckmann, J.P. Identifying impediments to long-distance mammal migrations. Conserv. Biol. 2015, 29, 99–109. [Google Scholar] [CrossRef]
- Klaassen, M.; Bauer, S.; Madsen, J.; Possingham, H. Optimal management of a goose flyway: Migrant management at minimum cost. J. Appl. Ecol. 2008, 45, 1446–1452. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Year | ID | Age | Type of Tracker | Migration Start Date 1 | Migration End Date 2 | Days | Number of Fixes | Accuracy (m) |
|---|---|---|---|---|---|---|---|---|
| 2013 | X06 | Adult | GPS-GSM | 1 Sep. 2013 | 30 Nov. 2013 | 90 | 3555 | <125 |
| 2014 | X06 | Adult | GPS-GSM | 1 Sep. 2014 | 2 Nov. 2014 | 62 | 1145 | <125 |
| 2014 | T2 | Juvenile | GPS-GSM | 10 Oct. 2014 | 3 Nov. 2014 | 23 | 1070 | <125 |
| 2014 | T3 | Juvenile | GPS-GSM | 22 Sep. 2014 | 2 Nov. 2014 | 40 | 1976 | <125 |
| 2014 | T4 | Adult | GPS-GSM | 15 Sep. 2014 | 5 Dec. 2014 | 80 | 3207 | <125 |
| 2014 | T5 | Adult | GPS-Argos | 4 Oct. 2014 | 2 Nov. 2014 | 29 | 229 | <100 |
| 2014 | T6 | Adult | GPS-Argos | 6 Oct. 2014 | 2 Nov. 2014 | 27 | 207 | <100 |
| 1992 | 9375 | Adult | GPS-Argos | 8 Oct. 1992 | 1 Nov. 1992 | 22 | 30 | 350–1000 |
| 1991 3 | 9377 | Unknown | GPS-Argos | 20 Oct. 1991 | 5 Nov. 1991 | 26 | 7 | 350–1000 |
| 1993 | 20248 | Adult | GPS-Argos | 9 Oct. 1993 | 30 Oct. 1993 | 21 | 33 | 350–1000 |
| 1993 | 20250 | Adult | GPS-Argos | 11 Oct. 1993 | 14 Nov. 1993 | 33 | 55 | 350–1000 |
| 1993 | 20252 | Adult | GPS-Argos | 17 Oct. 1993 | 16 Dec. 1993 | 64 | 99 | 350–1000 |
| 1993 | 20253 | Adult | GPS-Argos | 16 Oct. 1993 | 22 Nov. 1993 | 36 | 51 | 350–1000 |
| Sites | Survey Dates | Count | History Records Dates | Count |
|---|---|---|---|---|
| Beidaihe Reserve | Oct. 2013 | 0 | Oct. 1985 | 152 |
| Dalinor Lake | - | - | 1980s | >100 1 |
| Yellow River Delta Reserve | Nov. 2013 | 65 | Nov. 2004 | 600 2 |
| Beidagang Reservoir | Nov. 2013 | 117 | - | - |
| Duolun County | Oct. 2014 | 663 | - | - |
| Miyun Reservoir | Mar. 2013 | 1342 | - | - |
| Poyang Lake | Jan. 2012 | 738 | 1986 | 2200 1 |
| Beidaihe (1990s) | Duolun (2010s) | Change (%) | |
|---|---|---|---|
| Stopover (ha) | 2,029,093 | 1,710,596 | 15.7 |
| Corridor (ha) | 10,430,587 | 9646,780 | 7.51 |
| Path (ha) | 29,899,740 | 22,093,500 | 26.11 |
| Flatness | 6.79 | 7.74 | −13.99 |
| Path | Corridor | Stopover Site 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1990 | 2010 | ± % | 1990 | 2010 | ± % | 1990 | 2010 | ± % | |
| Grasslands 2 | 7,739,334 | 7,690,019 | −1 | 760,817 | 747,610 | −2 | 12,338 | 9300 | −25 |
| Wetlands | 469,380 | 309,220 | −34 | 173,326 | 98,681 | −43 | 83,650 | 48,547 | −42 |
| Open water 3 | 1,426,891 | 1,623,492 | 14 | 597,056 | 755,500 | 27 | 378,156 | 413,035 | 9 |
| Cropland | 14,098,260 | 13,143,803 | −7 | 4,566,698 | 4,119,561 | −10 | 649,615 | 520,820 | −20 |
| Developed area | 1,900,592 | 2,909,333 | 53 | 680,438 | 1,091,850 | 60 | 102,731 | 224,343 | 118 |
| Bare ground | 552,892 | 497,554 | −10 | 122,102 | 115,447 | −5 | 44,092 | 42,897 | −3 |
| Forest | 3,712,392 | 3,786,617 | 2 | 1,010,777 | 1,038,067 | 3 | 19,448 | 29,171 | 50 |
| Total Habitat | 23,733,865 | 22,766,534 | −4 | 6,097,897 | 5,721,353 | −6 | 1,123,759 | 991,703 | −12 |
| Habitat (%) | 79 | 76 | −4 | 77 | 72 | −21 | 87 | 77 | −12 |
| Path | Corridor | Stopover Site | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1990 | 2010 | ± % | 1990 | 2010 | ± % | 1990 | 2010 | ± % | |
| Grasslands 1 | 5,817,544 | 5,762,763 | −1 | 2,002,656 | 1,972,565 | −2 | 269,150 | 264,477 | −2 |
| Wetlands | 241,842 | 149,240 | −38 | 143,618 | 95,317 | −34 | 59,461 | 49,069 | −17 |
| Open water 2 | 559,258 | 672,197 | 20 | 352,726 | 416,558 | 18 | 137,986 | 150,095 | 9 |
| Cropland | 7,873,217 | 7,058,420 | −10 | 4,816,241 | 4,309,400 | −11 | 1,134,427 | 1,008,662 | −11 |
| Developed area | 1,334,287 | 2,002,262 | 50 | 788,626 | 1,178,942 | 49 | 177,934 | 298,303 | 68 |
| Bare ground | 398,317 | 414,681 | 4 | 228,867 | 242,431 | 6 | 73,849 | 76,935 | 4 |
| Forest | 1,960,865 | 2,112,819 | 8 | 896,720 | 1,006,983 | 12 | 103,442 | 108,564 | 5 |
| Total Habitat | 14,491,861 | 13,642,620 | −6 | 7,315,241 | 6,793,841 | −7 | 1,601,024 | 1,472,303 | −8 |
| Habitat (%) | 80 | 75 | −6 | 79 | 74 | −7 | 82 | 75 | −8 |
| Habitat | 1990 | 2010 | (± %) |
|---|---|---|---|
| Grasslands 1 | 12,338 | 9300 | −25 |
| Wetlands | 83,650 | 48,547 | −42 |
| Open water 2 | 378,156 | 413,035 | 9 |
| Cropland | 649,615 | 520,820 | −20 |
| Developed area | 102,731 | 224,343 | 118 |
| Bare ground | 44,092 | 42,897 | −3 |
| Forest | 19,448 | 29,171 | 50 |
| Total Habitat | 1,123,759 | 991,703 | −12 |
| Habitat (%) | 87 | 77 | −12 |
| Habitat | 1990 (ha) | 2010 (ha) | Change (%) |
|---|---|---|---|
| Grasslands 1 | 544,805 | 528,711 | −16,094 (−3) |
| Wetlands | 9600 | 435 | −9165 (−95) |
| Open water 2 | 37,778 | 38,590 | 812 (2) |
| Cropland | 1,027,548 | 1,006,438 | −21,110 (−2) |
| Developed area | 114,518 | 161,900 | 47,382 (41) |
| Bare ground | 24,030 | 18,194 | −5835 (−24) |
| Forest | 607,791 | 615,435 | 7644 (1) |
| Total Habitat | 1,619,731 | 1,574,174 | −45,557 (−3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Liu, Y.; Jiao, S.; Guo, J.; Lu, C.; Zhou, Y.; Wang, Y.; Lei, G.; Wen, L.; Mo, X. Shifting of the Migration Route of White-Naped Crane (Antigone vipio) Due to Wetland Loss in China. Remote Sens. 2021, 13, 2984. https://doi.org/10.3390/rs13152984
Jia Y, Liu Y, Jiao S, Guo J, Lu C, Zhou Y, Wang Y, Lei G, Wen L, Mo X. Shifting of the Migration Route of White-Naped Crane (Antigone vipio) Due to Wetland Loss in China. Remote Sensing. 2021; 13(15):2984. https://doi.org/10.3390/rs13152984
Chicago/Turabian StyleJia, Yifei, Yunzhu Liu, Shengwu Jiao, Jia Guo, Cai Lu, Yan Zhou, Yuyu Wang, Guangchun Lei, Li Wen, and Xunqiang Mo. 2021. "Shifting of the Migration Route of White-Naped Crane (Antigone vipio) Due to Wetland Loss in China" Remote Sensing 13, no. 15: 2984. https://doi.org/10.3390/rs13152984
APA StyleJia, Y., Liu, Y., Jiao, S., Guo, J., Lu, C., Zhou, Y., Wang, Y., Lei, G., Wen, L., & Mo, X. (2021). Shifting of the Migration Route of White-Naped Crane (Antigone vipio) Due to Wetland Loss in China. Remote Sensing, 13(15), 2984. https://doi.org/10.3390/rs13152984