
Forest Resistance and Resilience to 2002 Drought in Northern China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data
2.2.1. Model Tree Ensemble GPP
2.2.2. Satellite-Derived Vegetation Growth Data
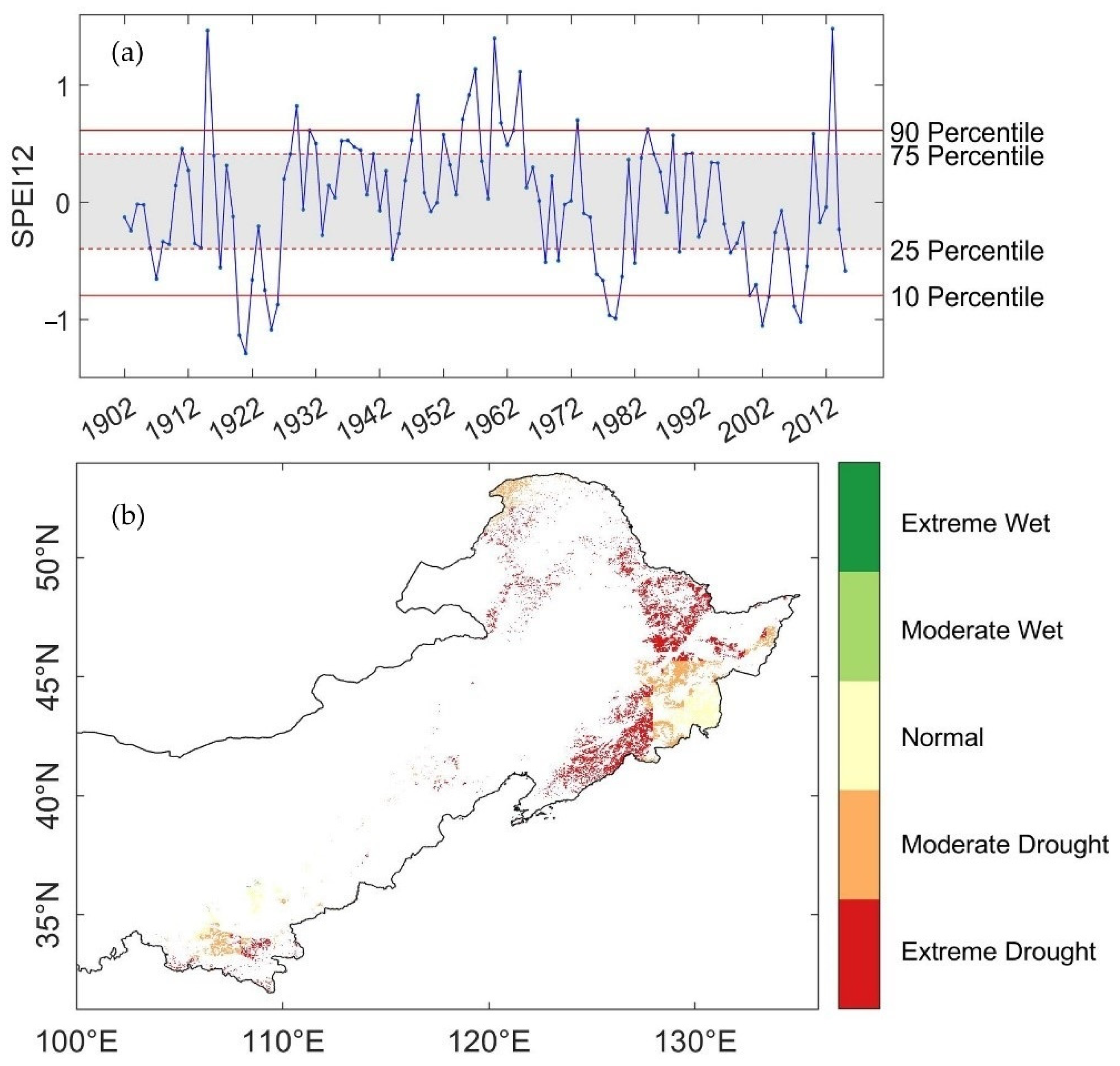
2.2.3. Drought Index
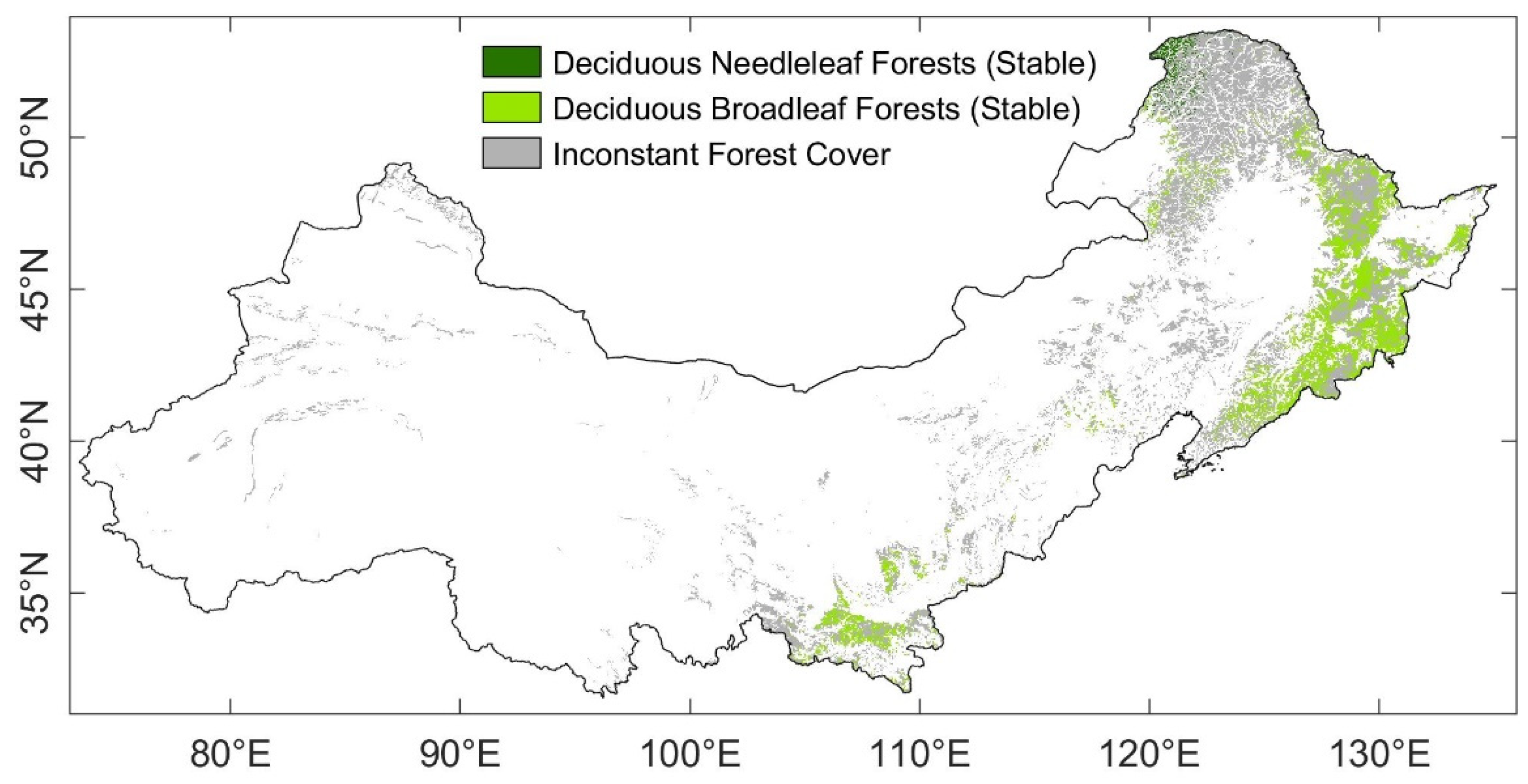
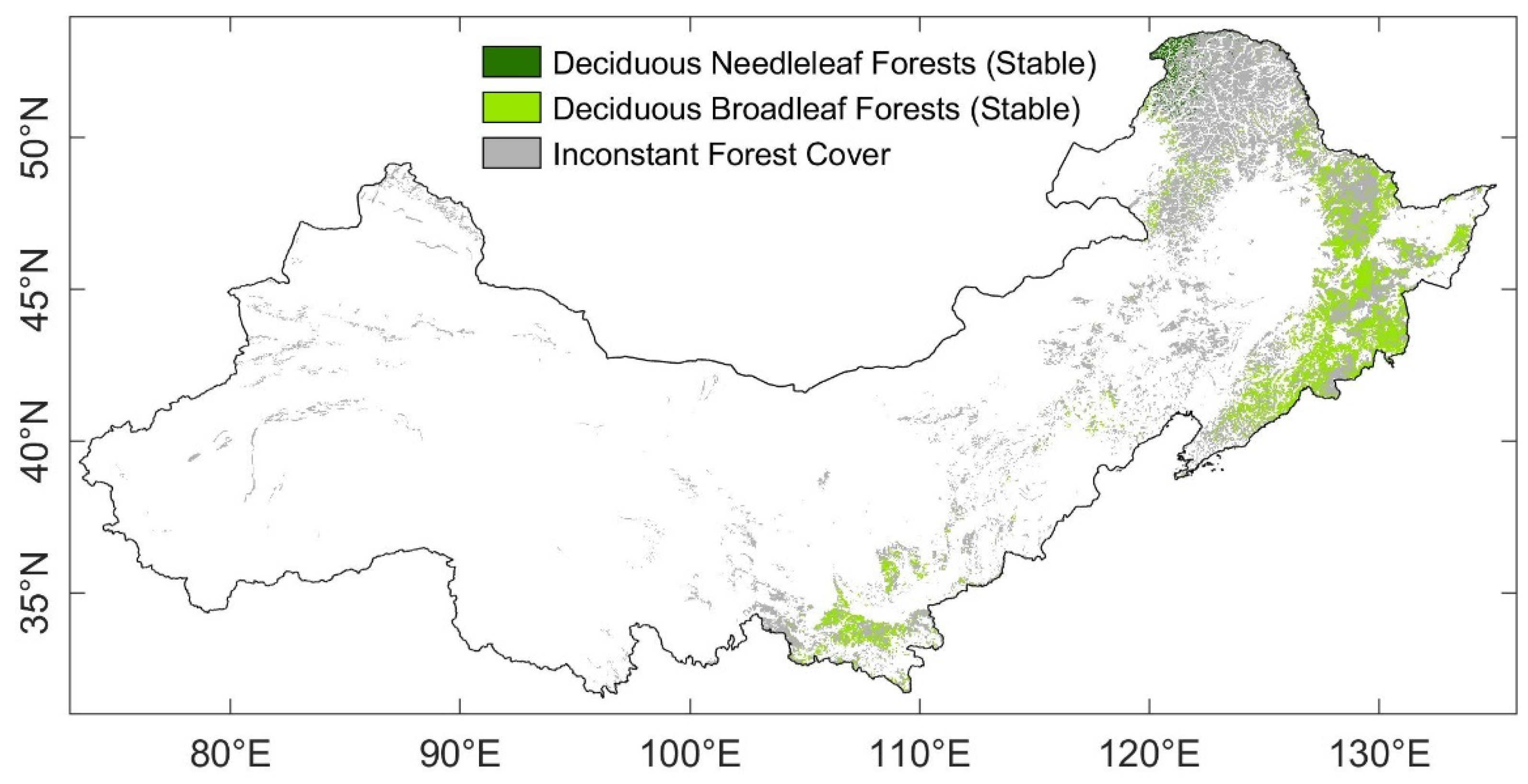
2.2.4. Vegetation Category
2.3. Methods
2.3.1. Defining Levels of Water-Balance Condition and Consecutive Droughts
2.3.2. Ecosystem Resistance and Resilience
2.3.3. Standardization
2.3.4. Pearson Correlation Analysis
2.3.5. Analysis of Variance (ANOVA)
3. Results
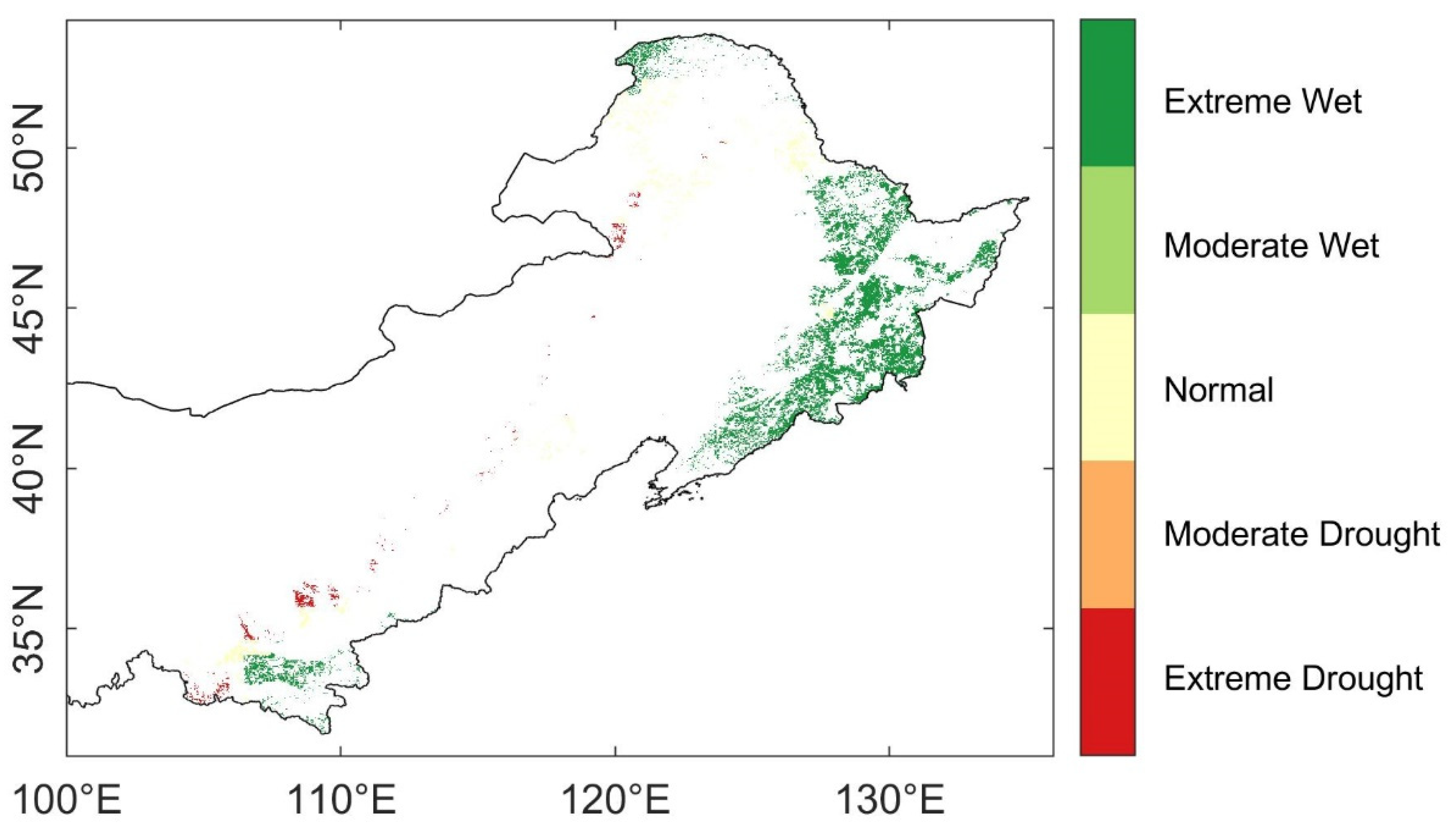
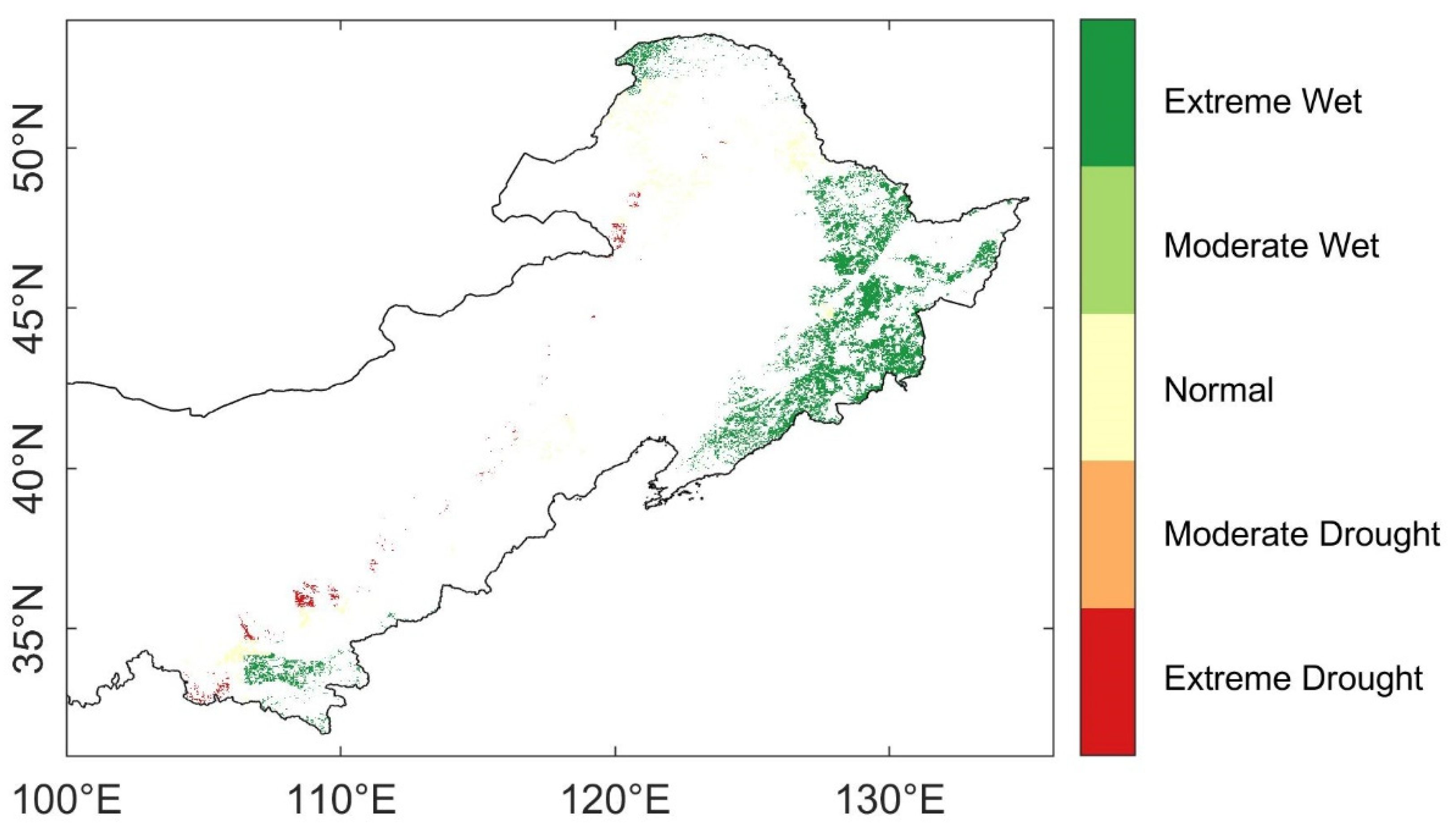
3.1. Spatio-Temporal Characteristics of Droughts
3.2. Interannual Variation of Forest Growth
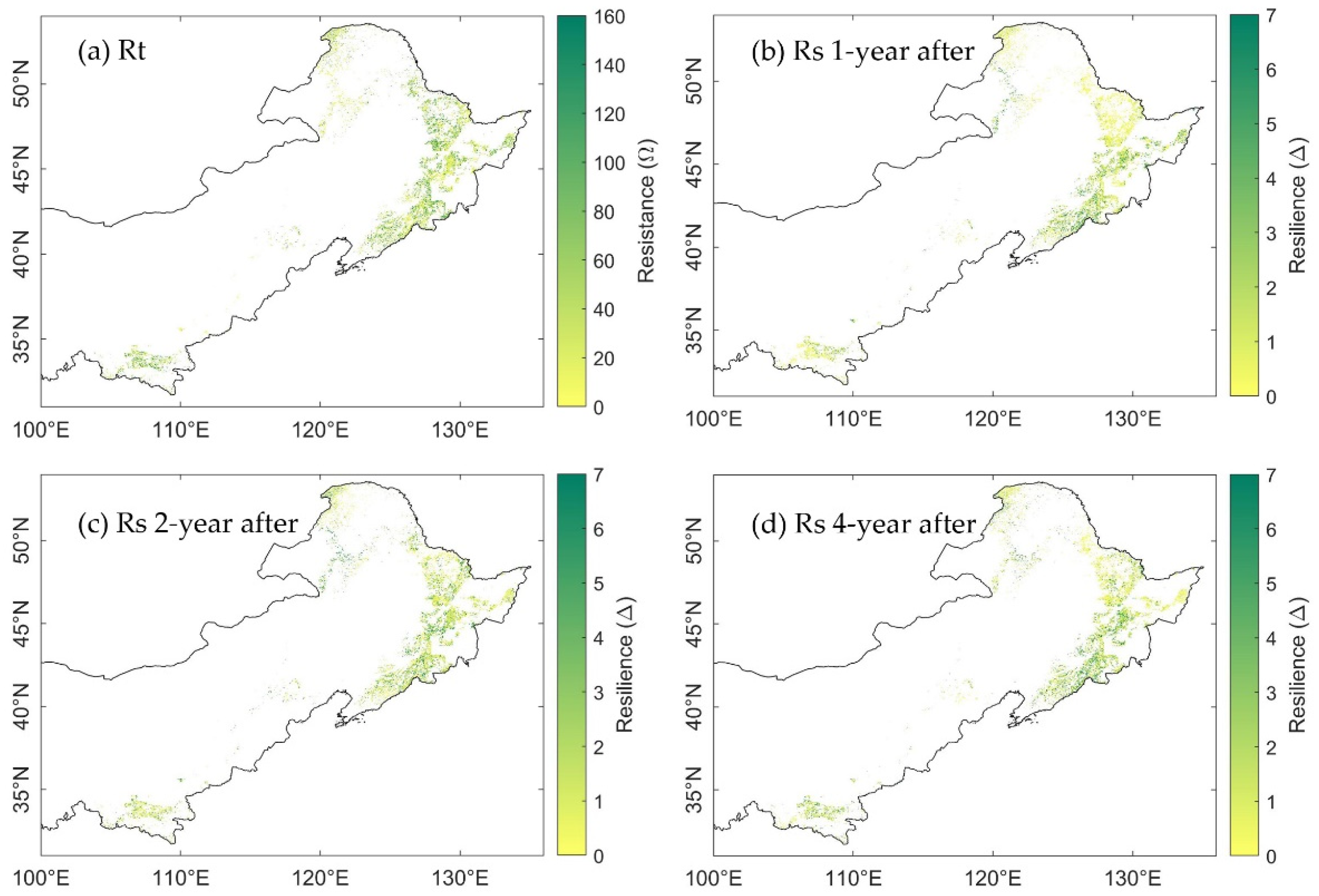
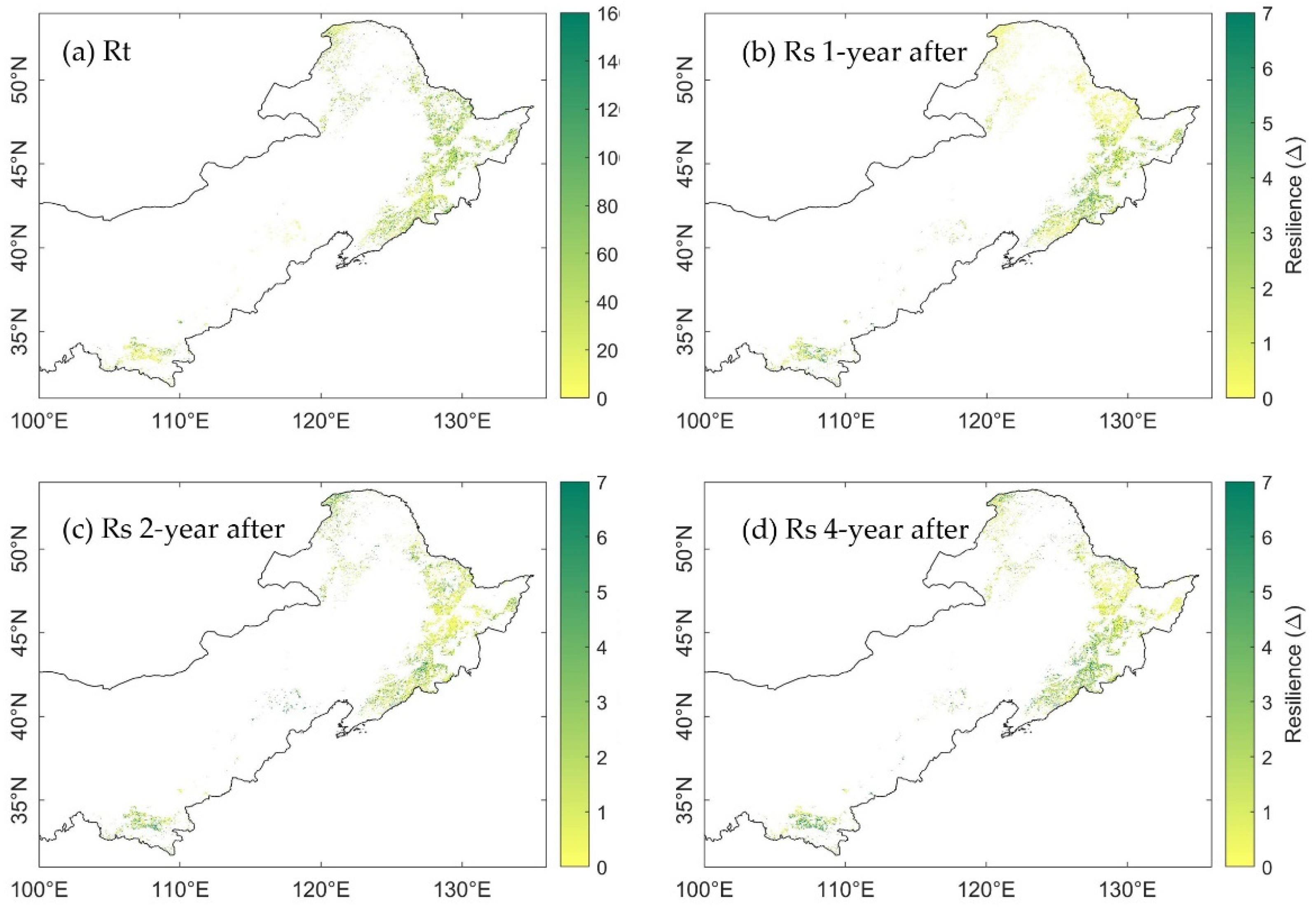
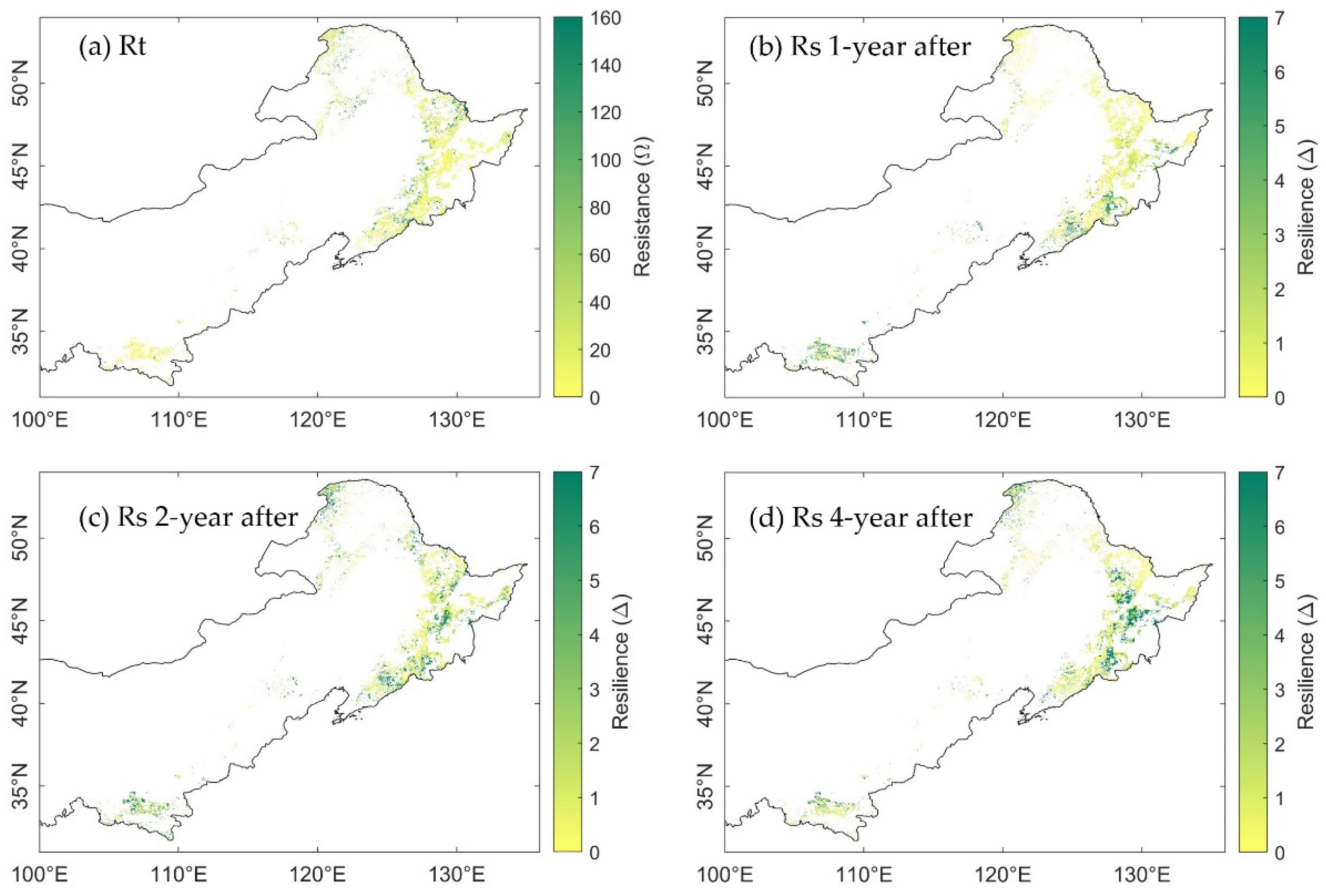
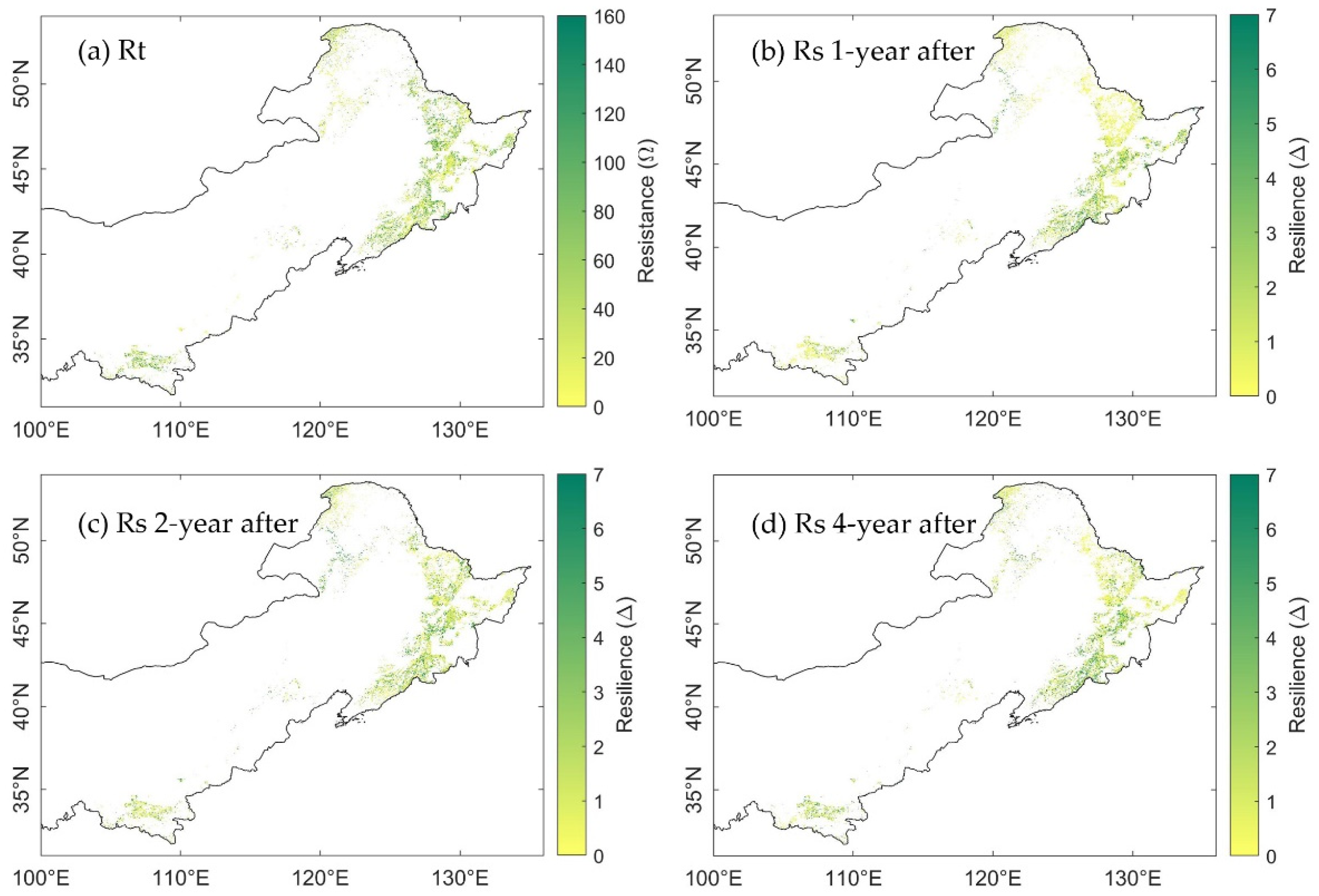
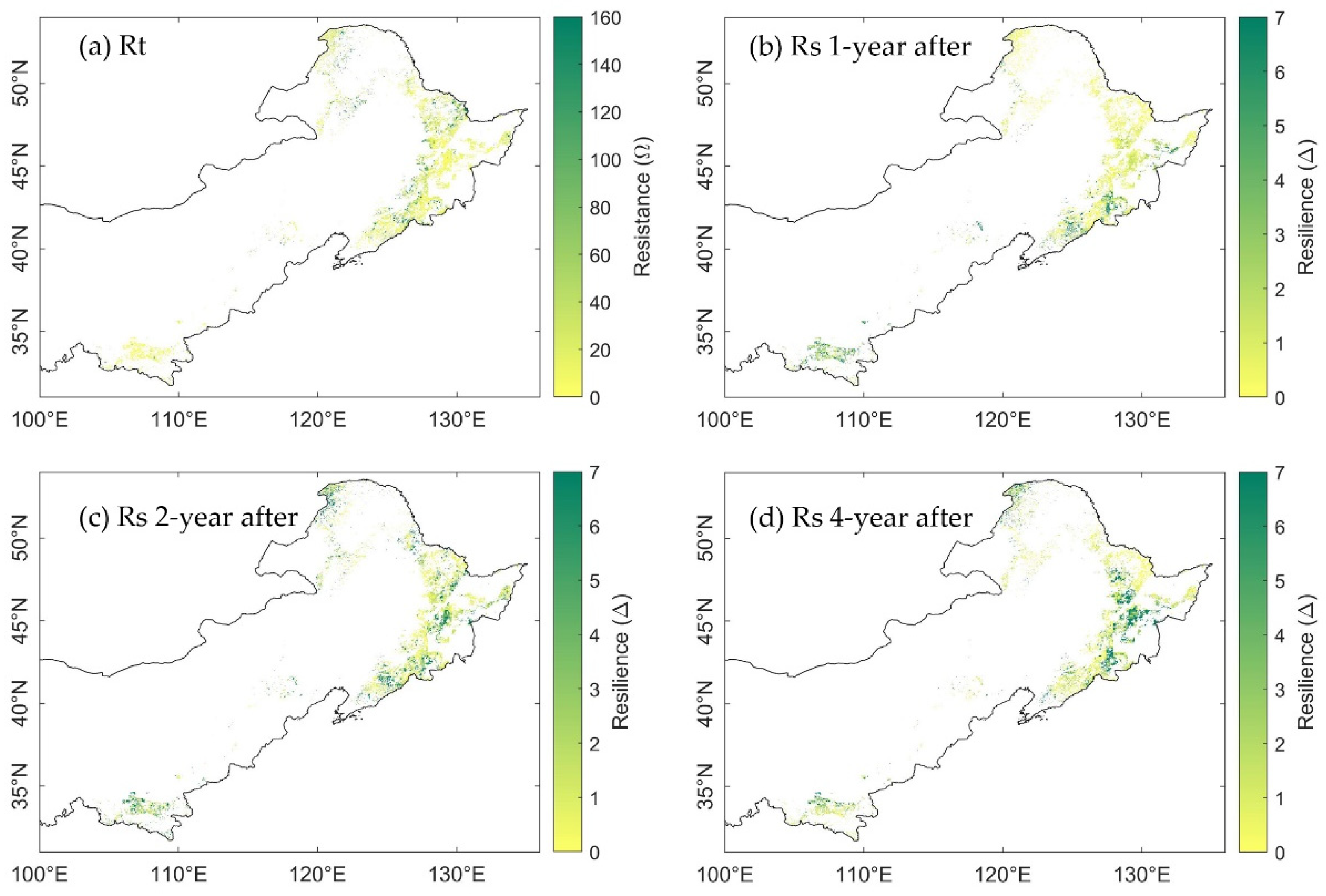
3.3. Spatial Distribution of Ecosystem Stability of the Forests to Drought in 2002
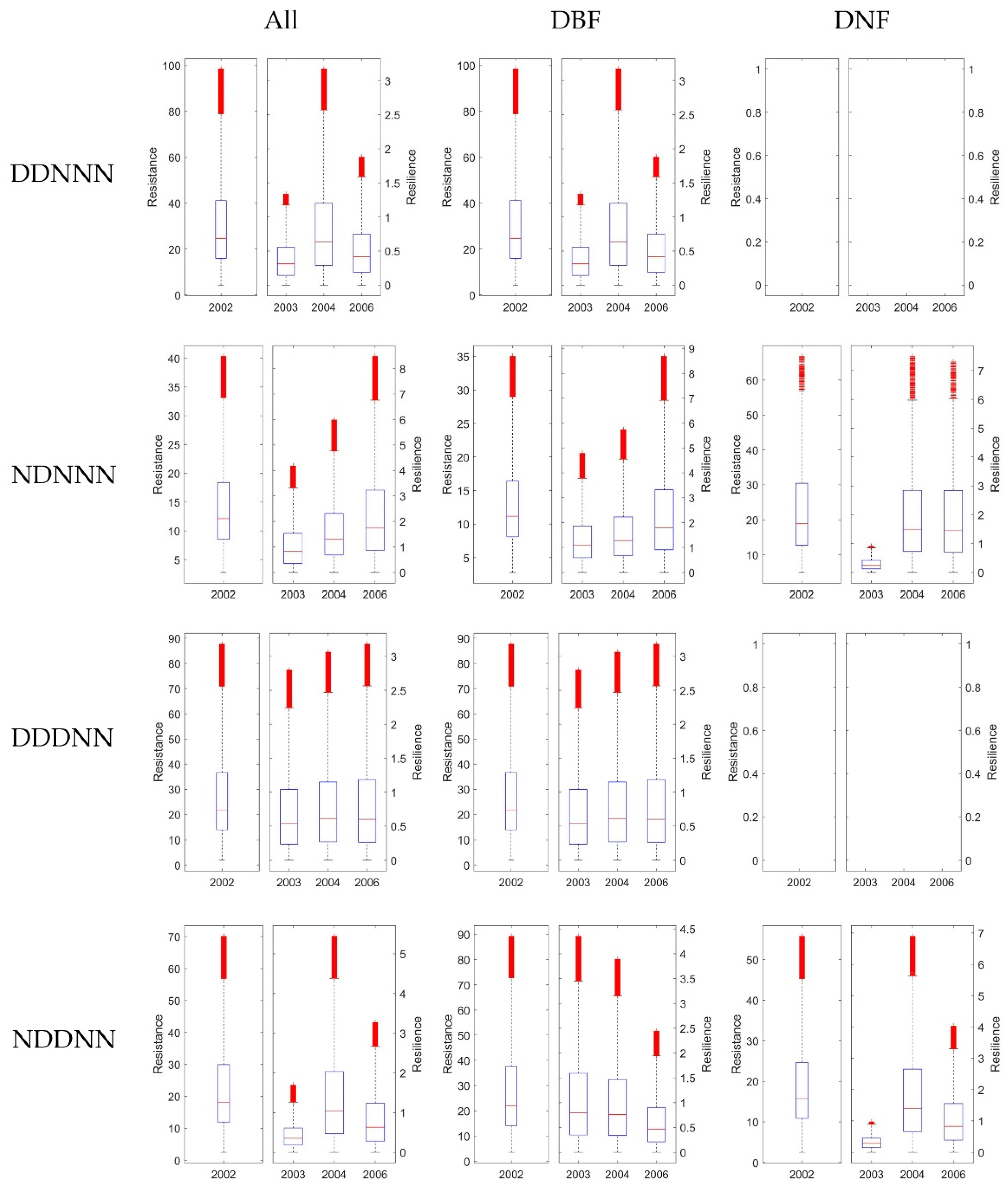
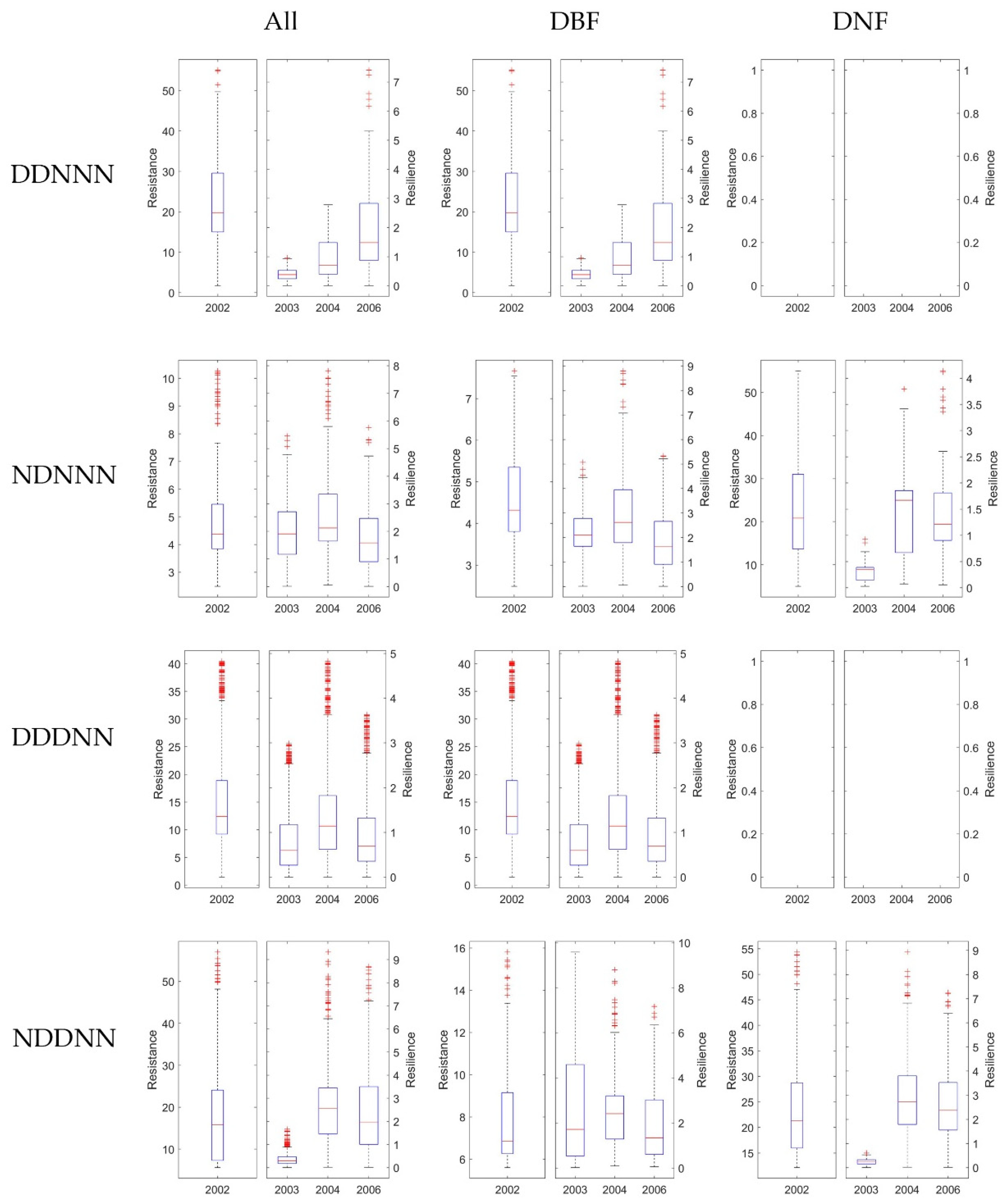
3.3.1. The Resistance during Drought in 2002
3.3.2. The Resilience at 1-, 2-, and 4-Years after the 2002 Drought
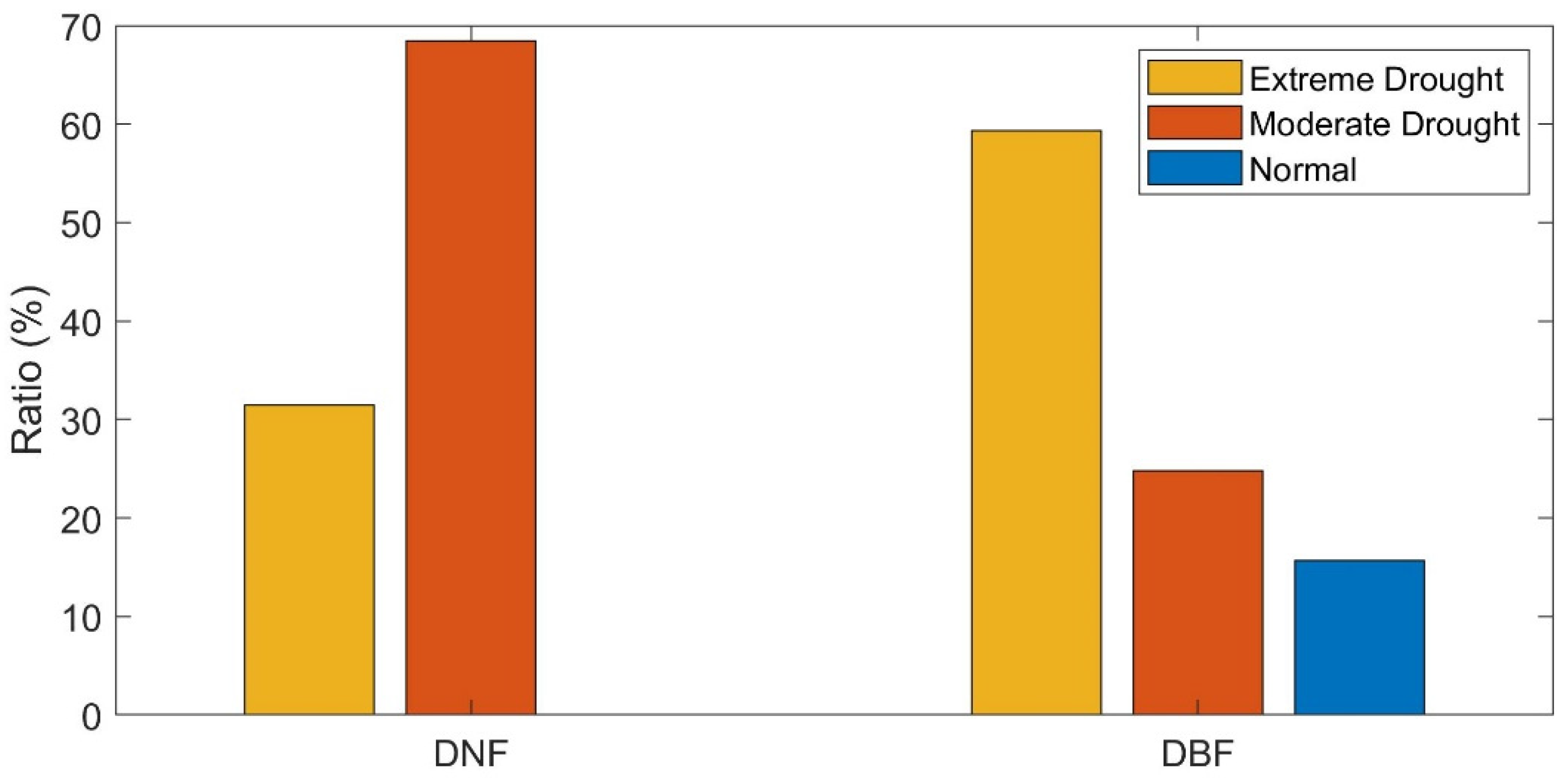
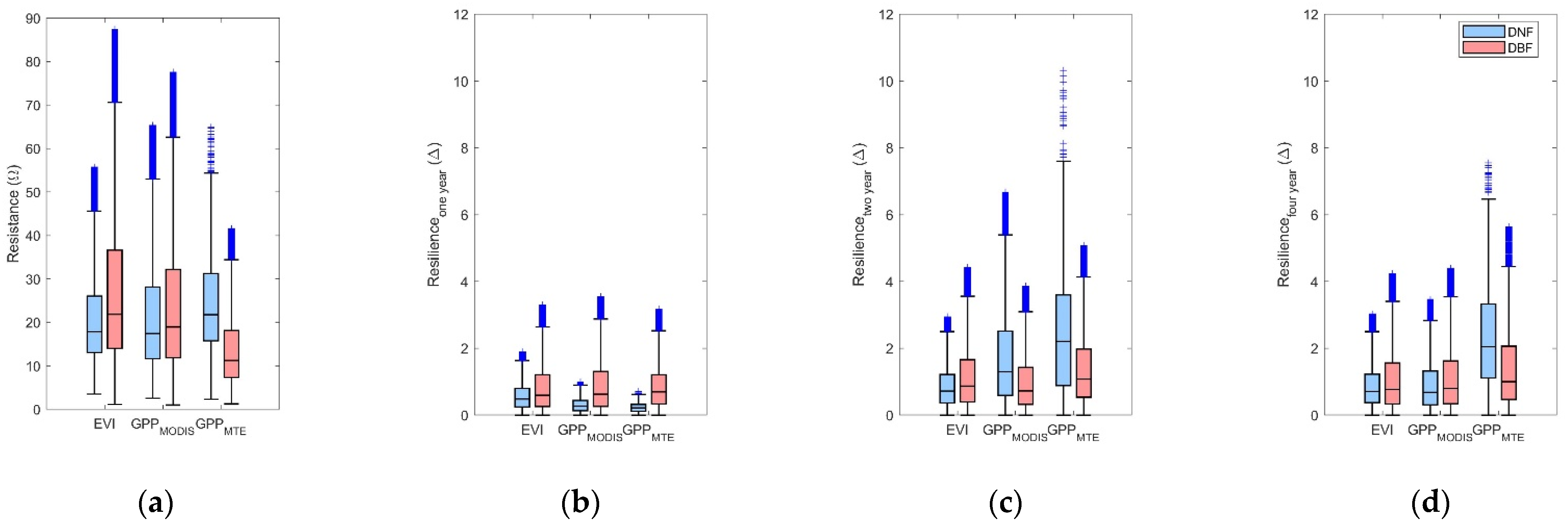
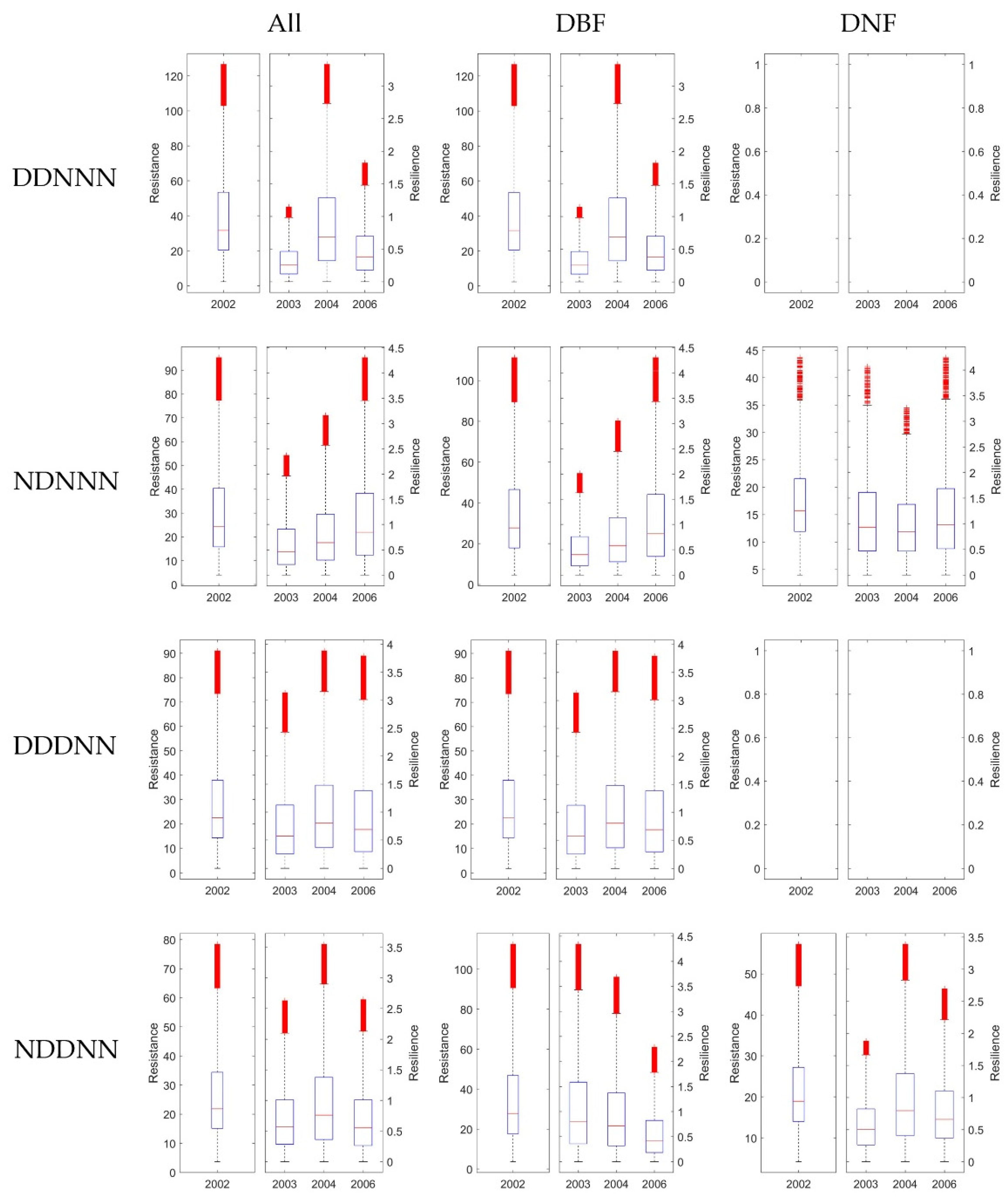
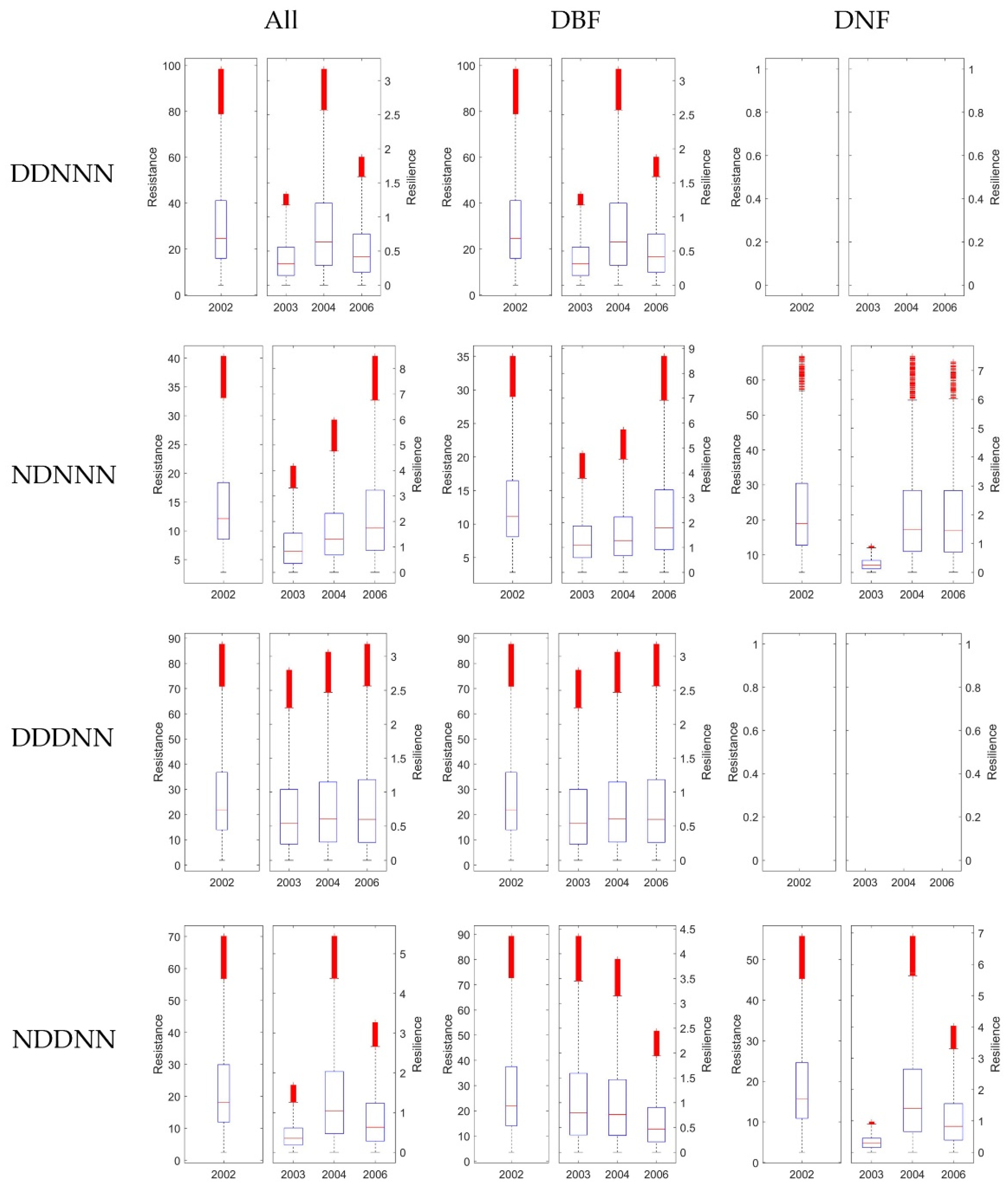
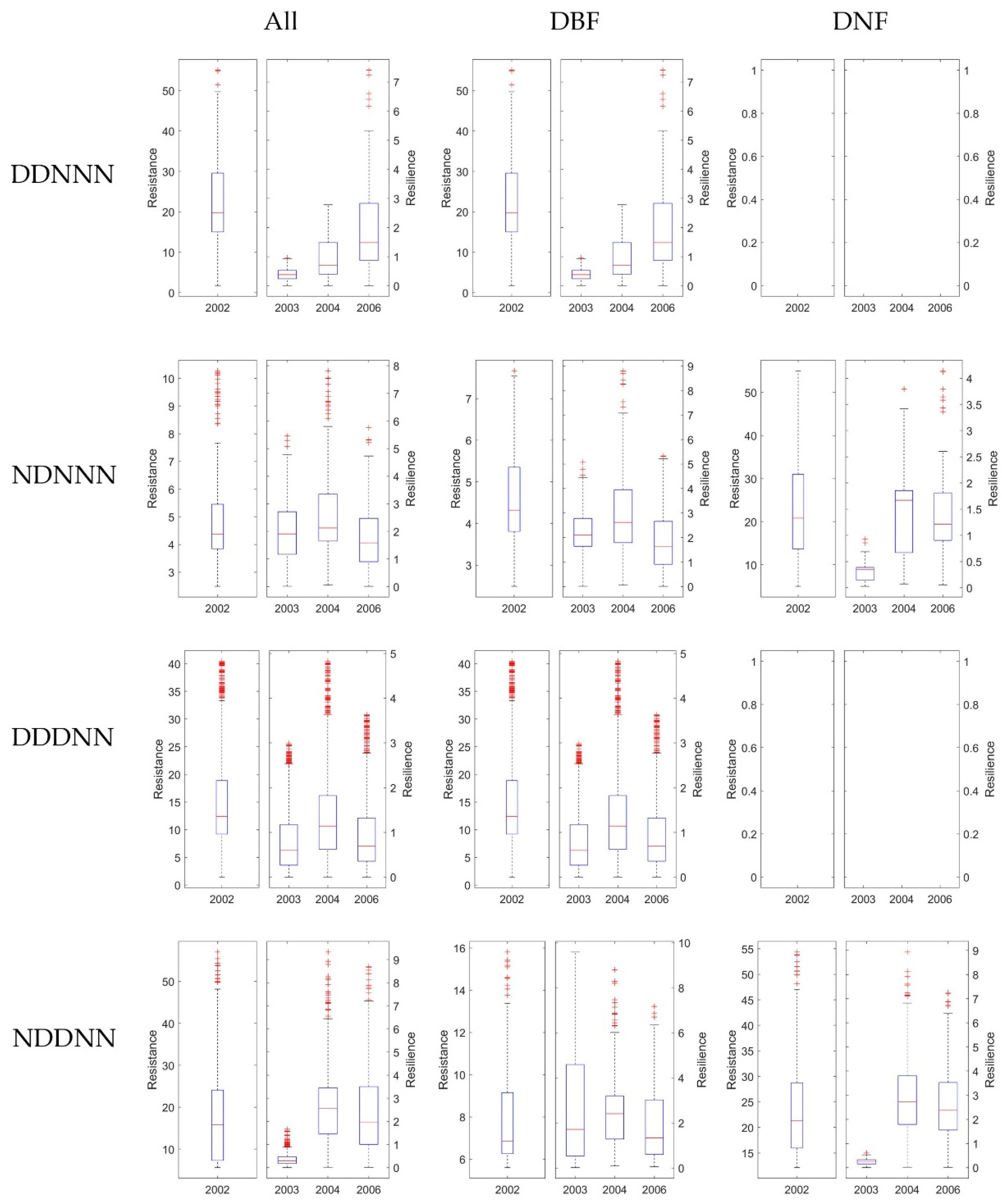
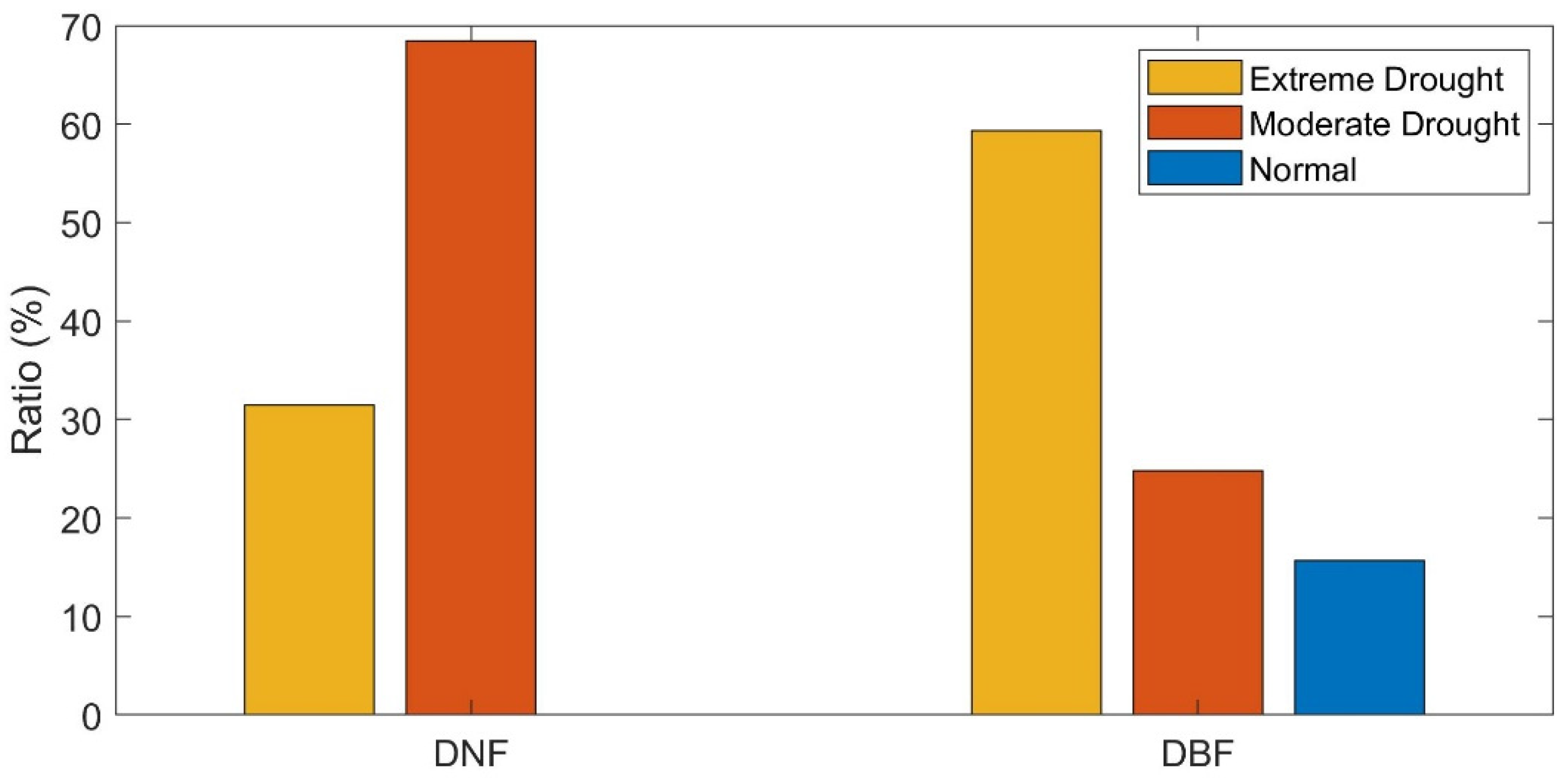
3.4. Stability of DBF and DNF to Drought in 2002
3.4.1. Comparison of the Resistance during 2002 between DBF and DNF
3.4.2. Comparison of the Resilience 1-, 2-, and 4-Year after Drought between DNF and DBF
4. Discussion
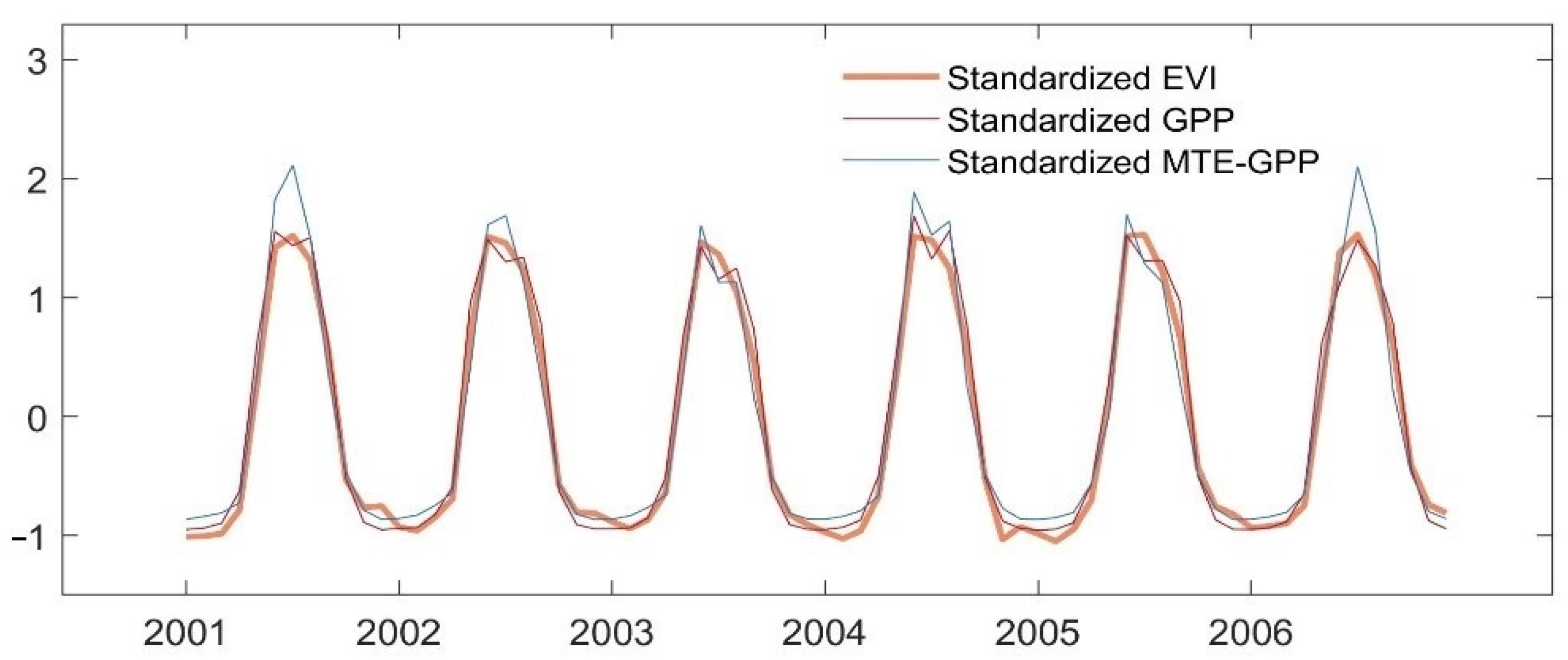
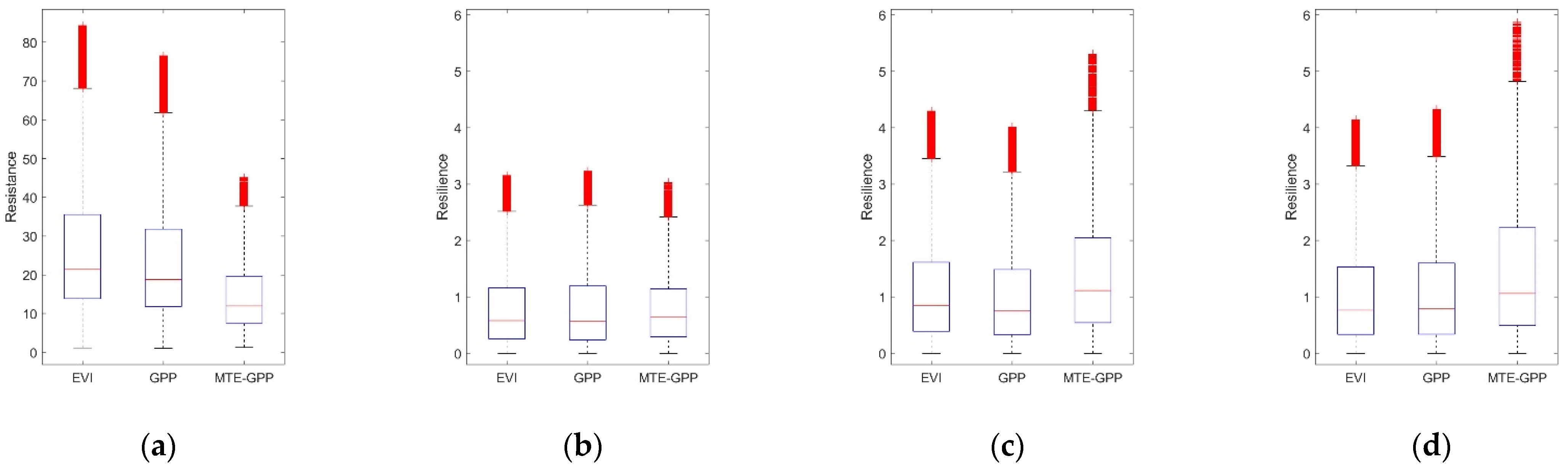
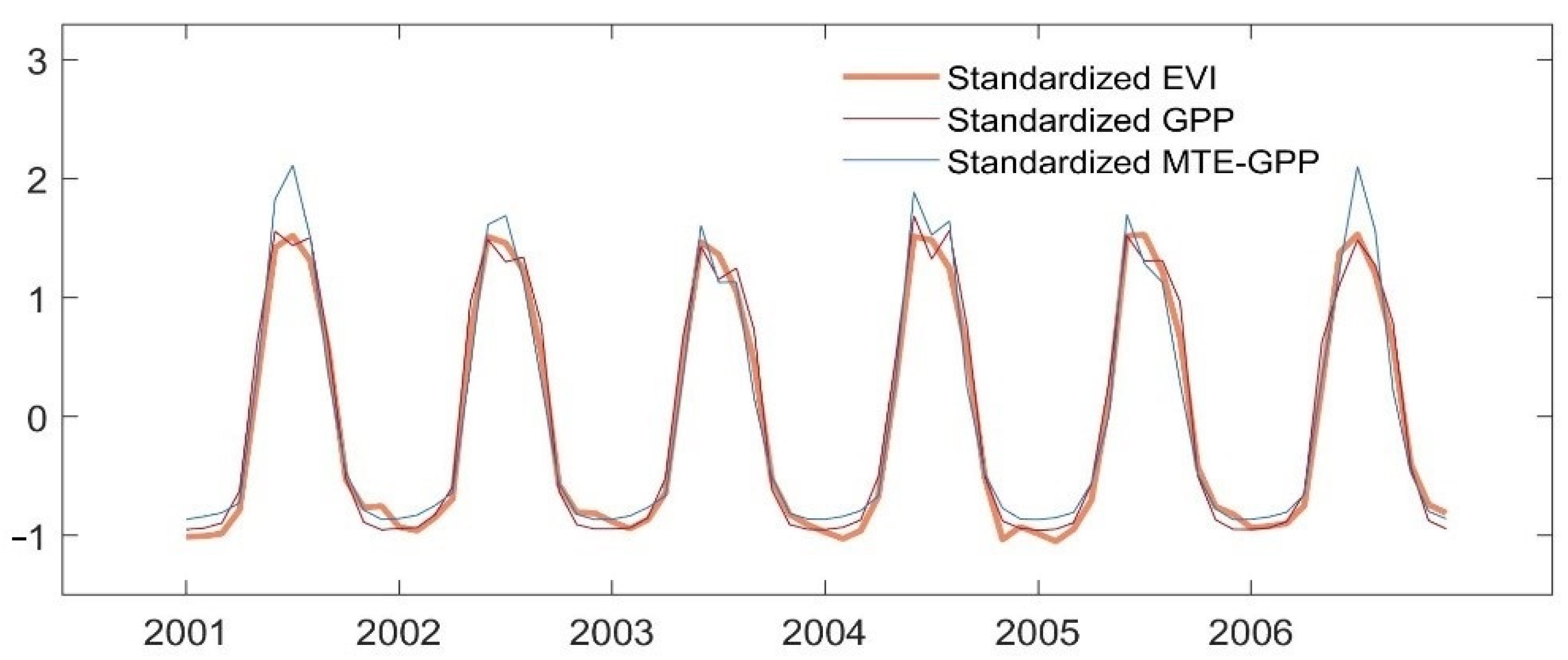
4.1. Comparison among Interannual Variation of EVI, GPP, and MTE-GPP
4.2. Differences between Ecosystem Stability of DNF and DBF
4.3. Comparison of Ecosystem Stability Considering Consecutive Drought
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Type | Type of Variability |
|---|---|---|
| Mean annual temperature | Split | Static |
| Mean annual temperature maximum | Split | Static |
| Mean annual precipitation sum | Split | Static |
| Mean annual radiation | Split | Static |
| Mean annual FPAR | Split | Static |
| Mean monthly temperature | Split | Monthly but static over years |
| Mean monthly temperature maximum | Split | Monthly but static over years |
| Mean monthly precipitation sum | Split | Monthly but static over years |
| Mean monthly radiation | Split | Monthly but static over years |
| Mean monthly FPAR | Split | Monthly but static over years |
| Monthly temperature | Split & Regression | Monthly |
| Monthly temperature maximum | Split & Regression | Monthly |
| Monthly precipitation | Split & Regression | Monthly |
| Monthly precipitation a month before | Split & Regression | Monthly |
| Monthly radiation | Split & Regression | Monthly |
| Monthly FPAR | Split & Regression | Monthly |
| Vegetation type | Split | Static |
| ‘SS’ | ‘df’ | ‘MS’ | ‘F’ | ‘Prob > F’ | |
|---|---|---|---|---|---|
| EVI-Rt | 2.14 × 10+6 | 1 | 2.14 × 10+6 | 6.53 × 10+3 | 0 |
| EVI-Rs 1 | 3.68 × 10+3 | 1 | 3.68 × 10+3 | 6.58 × 10+3 | 0 |
| EVI-Rs 2 | 4.55 × 10+3 | 1 | 4.55 × 10+3 | 4.62 × 10+3 | 0 |
| EVI-Rs 3 | 2.12 × 10+3 | 1 | 2.12 × 10+4 | 1.74 × 10+4 | 0 |
| EVI-Rs 4 | 2.06 × 10+3 | 1 | 2.06 × 1003 | 2.21 × 10+3 | 0 |
| GPP-Rt | 2.93 × 10+5 | 1 | 2.93 × 10+5 | 1.09 × 10+3 | 8.365 × 10−238 |
| GPP-Rs1 | 1.09 × 10+4 | 1 | 1.09 × 10+4 | 2.80 × 10+4 | 0 |
| GPP-Rs 2 | 2.65 × 10+4 | 1 | 2.65 × 10+4 | 2.98 × 10+4 | 0 |
| GPP-Rs 3 | 1.80 × 10+3 | 1 | 1.80 × 10+3 | 2.02 × 10+3 | 0 |
| GPP-Rs 4 | 1.90 × 10+3 | 1 | 1.90 × 10+3 | 1.88 × 10+3 | 0 |
| MTEGPP-Rt | 5.38 × 10+6 | 1 | 5.38 × 10+6 | 6.23 × 10+4 | 0 |
| MTEGPP-Rs1 | 2.03 × 10+4 | 1 | 2.03 × 10+4 | 4.21 × 10+4 | 0 |
| MTEGPP-Rs 2 | 6.79 × 10+4 | 1 | 6.79 × 10+4 | 4.45 × 10+4 | 0 |
| MTEGPP-Rs 3 | 2.95 × 10+5 | 1 | 2.95 × 10+5 | 2.81 × 10+5 | 0 |
| MTEGPP-Rs 4 | 3.84 × 10+4 | 1 | 3.84 × 10+4 | 2.19 × 10+4 | 0 |

References
- Ding, Y.; Xu, J.; Wang, X.; Peng, X.; Cai, H. Spatial and temporal effects of drought on Chinese vegetation under different coverage levels. Sci. Total Environ. 2020, 716, 137166. [Google Scholar] [CrossRef]
- Liang, E.; Shao, X.; Kong, Z.; Lin, J. The extreme drought in the 1920s and its effect on tree growth deduced from tree ring analysis: A case study in North China. Ann. For. Sci. 2003, 60, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Ren, S.; Li, Y.; Dong, J.; Wang, L.; Quan, Q.; Liu, J. Diverse responses of spring phenology to preseason drought and warming under different biomes in the North China Plain. Sci. Total Environ. 2021, 766, 144437. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Hartmann, H.; Moura, C.F.; Anderegg, W.R.L.; Ruehr, N.K.; Salmon, Y.; Allen, C.D.; Arndt, S.K.; Breshears, D.D.; Davi, H.; Galbraith, D.; et al. Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 2018, 218, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Wei, X.; Rich, R.L.; Montgomery, R.A. Boreal and temperate trees show strong acclimation of respiration to warming. Nature 2016, 531, 633–636. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Guti, E.; Novak, K.; T, V.R.P.A.; Ribas, M.; Garc, I.; Alvaro, S.; Galv, J.D. Forest resilience to drought varies across biomes. Glob. Chang. Biol. 2018, 2143–2158. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Konings, A.G.; Shaw, J. Divergent forest sensitivity to repeated extreme droughts. Nat. Clim. Chang. 2020, 10, 1091–1095. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2020, 4, 1075–1083. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Novick, K.A.; Alexander, M.R.; Maxwell, J.T.; Moore, D.J.P.; Phillips, R.P.; Anderegg, W.R.L. Linking drought legacy effects across scales: From leaves to tree rings to ecosystems. Glob. Chang. Biol. 2019, 25, 2978–2992. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J. High ecosystem stability of evergreen broadleaf forests under severe droughts. Glob. Chang. Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef]
- Gong, Z.; Zhao, S.; Gu, J. Correlation analysis between vegetation coverage and climate drought conditions in North China during 2001–2013. J. Geogr. Sci. 2017, 27, 143–160. [Google Scholar] [CrossRef]
- Weiss, S.M.; Kulikowski, C.A. Computer Systems That Learn: Classification and Prediction Methods from Statistics, Neural Nets, Machine Learning, and Expert Systems; M. Kaufmann Publishers: San Mateo, CA, USA, 1991; ISBN 1558600655. [Google Scholar]
- Bastos, A.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Pongratz, J.; Fan, L.; Wigneron, J.P.; Weber, U.; Reichstein, M.; Fu, Z.; et al. Direct and seasonal legacy effects of the 2018 heat wave and drought on European ecosystem productivity. Sci. Adv. 2020, 6, 1–14. [Google Scholar] [CrossRef]
- Zan, M.; Zhou, Y.; Ju, W.; Zhang, Y.; Zhang, L.; Liu, Y. Performance of a two-leaf light use efficiency model for mapping gross primary productivity against remotely sensed sun-induced chlorophyll fluorescence data. Sci. Total Environ. 2018, 613–614, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science (80-.) 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichii, K.; Ueyama, M.; Kondo, M.; Saigusa, N.; Kim, J.; Alberto, M.C.; Ardö, J.; Euskirchen, E.S.; Kang, M.; Hirano, T.; et al. New data-driven estimation of terrestrial CO2 fluxes in Asia using a standardized database of eddy covariance measurements, remote sensing data, and support vector regression. J. Geophys. Res. Biogeosci. 2017, 122, 767–795. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Bondeau, A. Towards global empirical upscaling of FLUXNET eddy covariance observations: Validation of a model tree ensemble approach using a biosphere model. Biogeosciences 2009, 6, 2001–2013. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Hao, Z.; Hao, F.; Zhang, X. Variations of compound precipitation and temperature extremes in China during 1961–2014. Sci. Total Environ. 2019, 663, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, X.; Li, Y.; Wang, T.; Shen, M.; Du, M.; He, H.; Li, Y.; Luo, W.; Ma, M.; et al. Spatiotemporal pattern of gross primary productivity and its covariation with climate in China over the last thirty years. Glob. Chang. Biol. 2018, 24, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, X.; Wang, K.; Ciais, P.; Tang, S.; Jin, L.; Li, L.; Piao, S. Responses of vegetation greenness and carbon cycle to extreme droughts in China. Agric. For. Meteorol. 2021, 298–299, 108307. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Van Leeuwen, W. Modis Vegetation Index. Veg. Index Phenol. Lab 1999, 3, 129. [Google Scholar]
- Solano, R.; Didan, K.; Jacobson, A.; Huete, A. MODIS Vegetation Index User ’ s Guide (MOD13 Series). Univ. Arizona 2010, 2010, 38. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Fu, B.J.; He, C.S.; Sun, G.; Gao, G.Y. A comparative analysis of forest cover and catchment water yield relationships in northern China. For. Ecol. Manag. 2011, 262, 1189–1198. [Google Scholar] [CrossRef]
- Editorial Board of Vegetaion Map of China. Vegetation Map of the People’s Republic of China (1:1000000) (Digital Version), 1st ed.; Editorial Board of Vegetation Map of China, Ed.; Geology Press: Beijing, China, 2007; ISBN 9787503142888. [Google Scholar]
- Su, Y.; Guo, Q.; Hu, T.; Guan, H.; Jin, S.; An, S.; Chen, X.; Guo, K.; Hao, Z.; Hu, Y.; et al. An updated Vegetation Map of China (1:1000000). Sci. Bull. 2020, 65, 1125–1136. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science (80-) 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Decuyper, M.; Chávez, R.O.; Čufar, K.; Estay, S.A.; Clevers, J.G.P.W.; Prislan, P.; Gričar, J.; Črepinšek, Z.; Merela, M.; de Luis, M.; et al. Spatio-temporal assessment of beech growth in relation to climate extremes in Slovenia—An integrated approach using remote sensing and tree-ring data. Agric. For. Meteorol. 2020, 287, 107925. [Google Scholar] [CrossRef]
- Pennington, R.T.; Lavin, M. The contrasting nature of woody plant species in different neotropical forest biomes reflects differences in ecological stability. New Phytol. 2016, 210, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
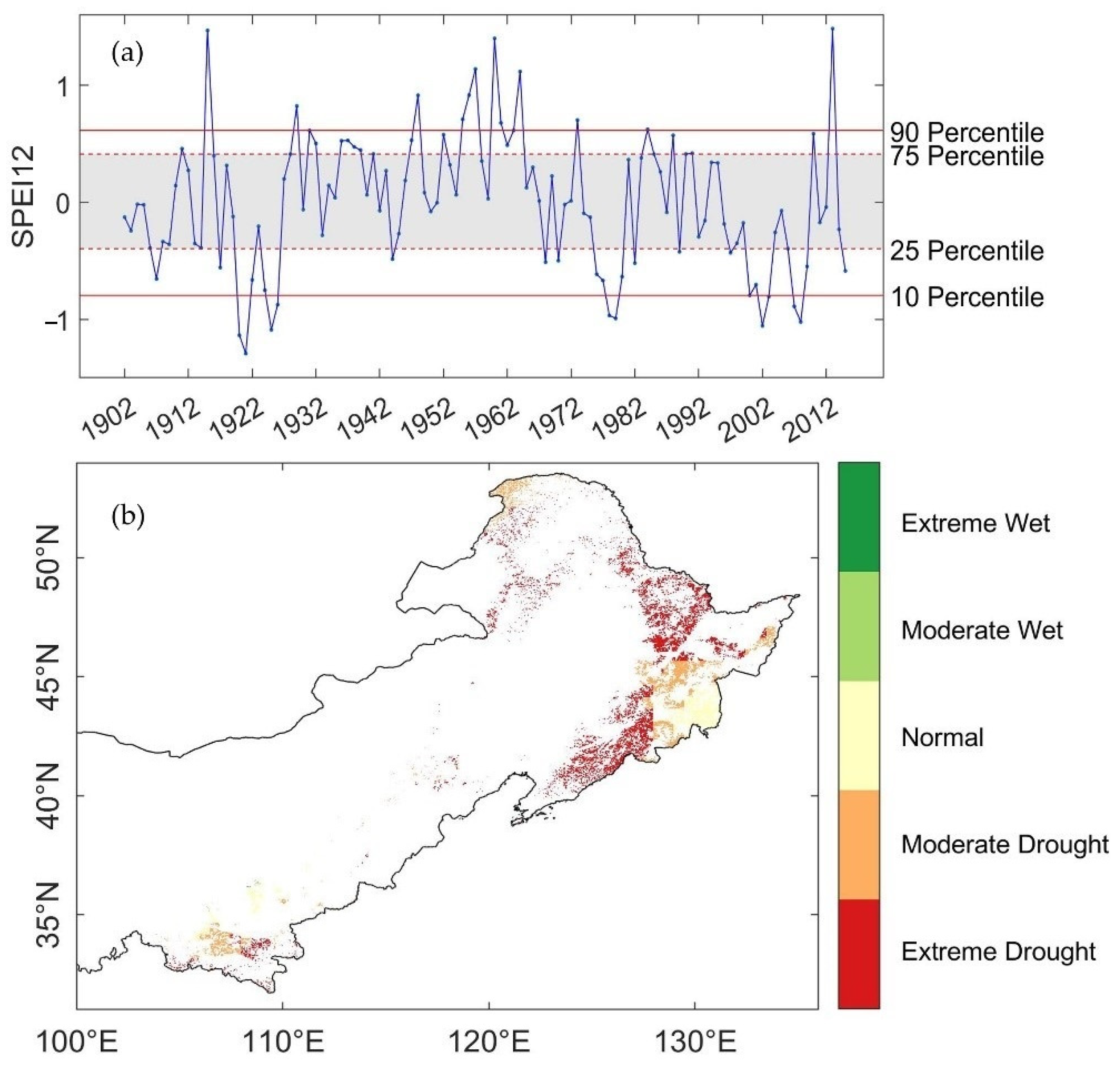



| SPEI Percentile | Condition |
|---|---|
| ≥90th percentile | Extreme wet |
| 75th percentile~90th percentile | Moderate wet |
| 25th percentile~75th percentile | Normal |
| 10th percentile~25th percentile | Moderate Drought |
| ≤10th percentile | Extreme Drought |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yao, Y.; Yin, G.; Peng, F.; Liu, M. Forest Resistance and Resilience to 2002 Drought in Northern China. Remote Sens. 2021, 13, 2919. https://doi.org/10.3390/rs13152919
Li X, Yao Y, Yin G, Peng F, Liu M. Forest Resistance and Resilience to 2002 Drought in Northern China. Remote Sensing. 2021; 13(15):2919. https://doi.org/10.3390/rs13152919
Chicago/Turabian StyleLi, Xiran, Yitong Yao, Guodong Yin, Feifei Peng, and Muxing Liu. 2021. "Forest Resistance and Resilience to 2002 Drought in Northern China" Remote Sensing 13, no. 15: 2919. https://doi.org/10.3390/rs13152919
APA StyleLi, X., Yao, Y., Yin, G., Peng, F., & Liu, M. (2021). Forest Resistance and Resilience to 2002 Drought in Northern China. Remote Sensing, 13(15), 2919. https://doi.org/10.3390/rs13152919