
Phytoplankton Biomass and the Hydrodynamic Regime in NEOM, Red Sea

 , , ,
, , ,  , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
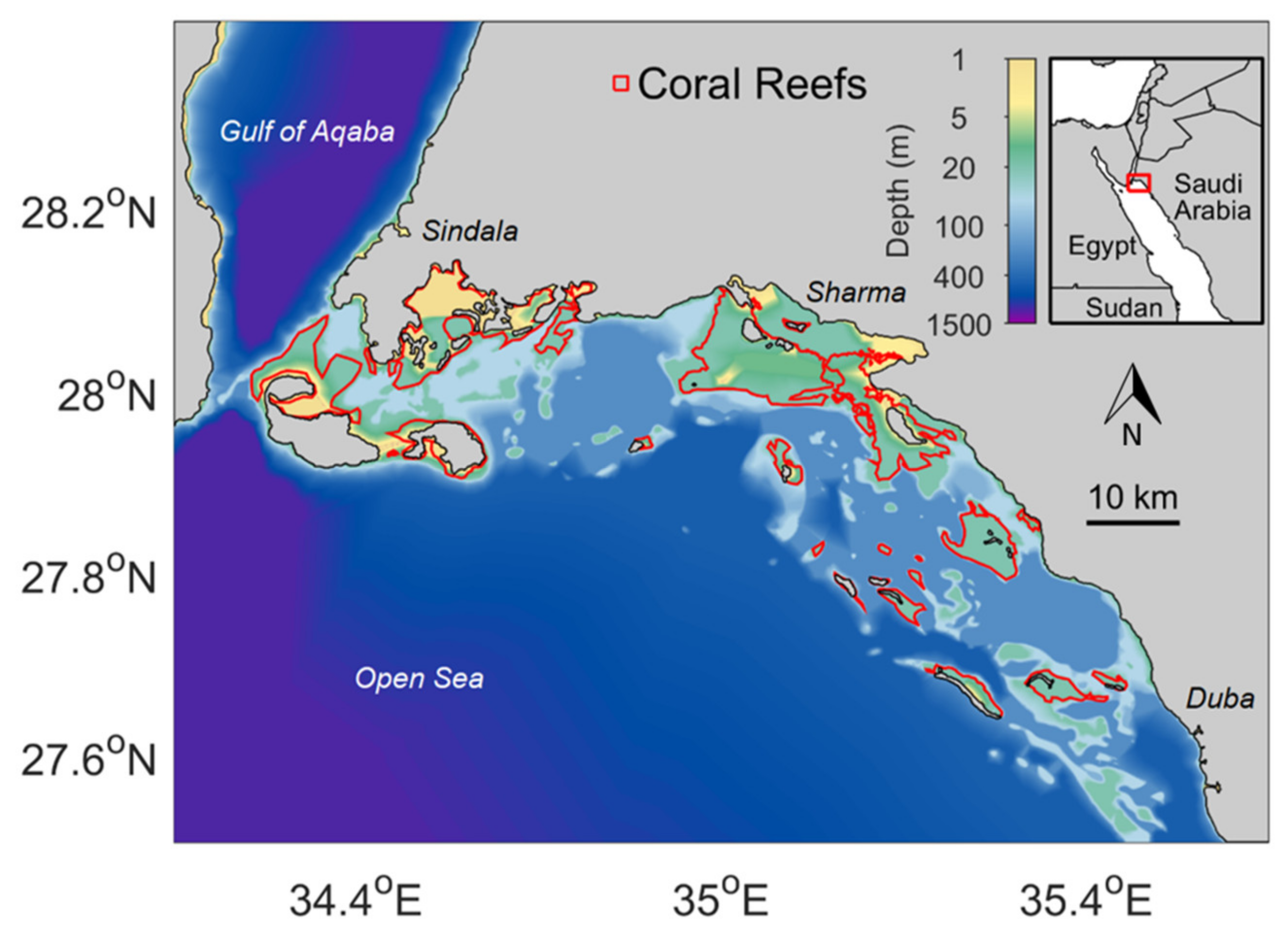
2.1. Satellite Remote Sensing Data and Study Area
2.2. Hydrodynamic Model
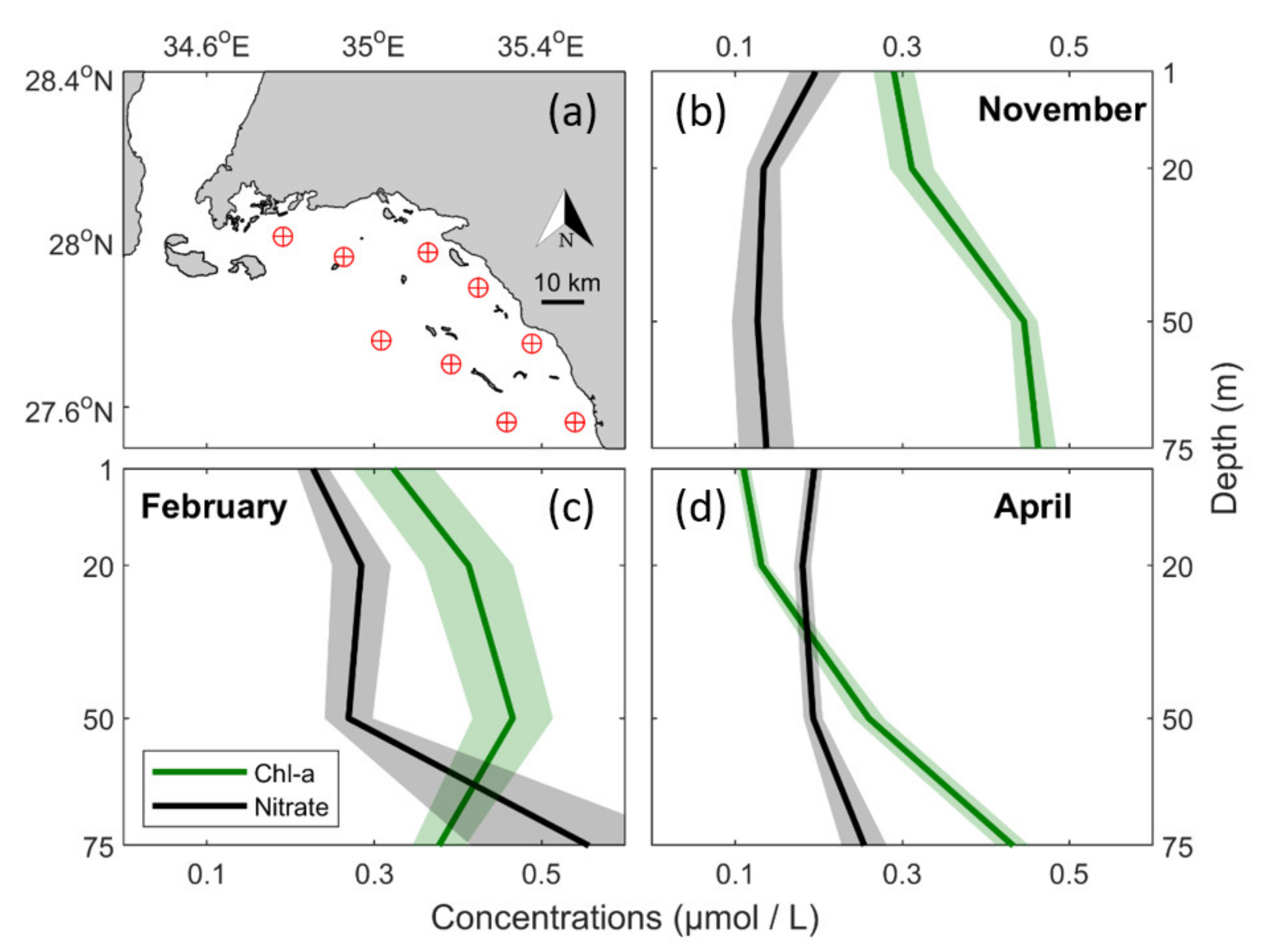
2.3. In Situ Data of Chl-a, Nutrients, Temperature and Salinity
2.4. Sea Floor Morphology and Coral Reef Data
3. Results
3.1. Area Description
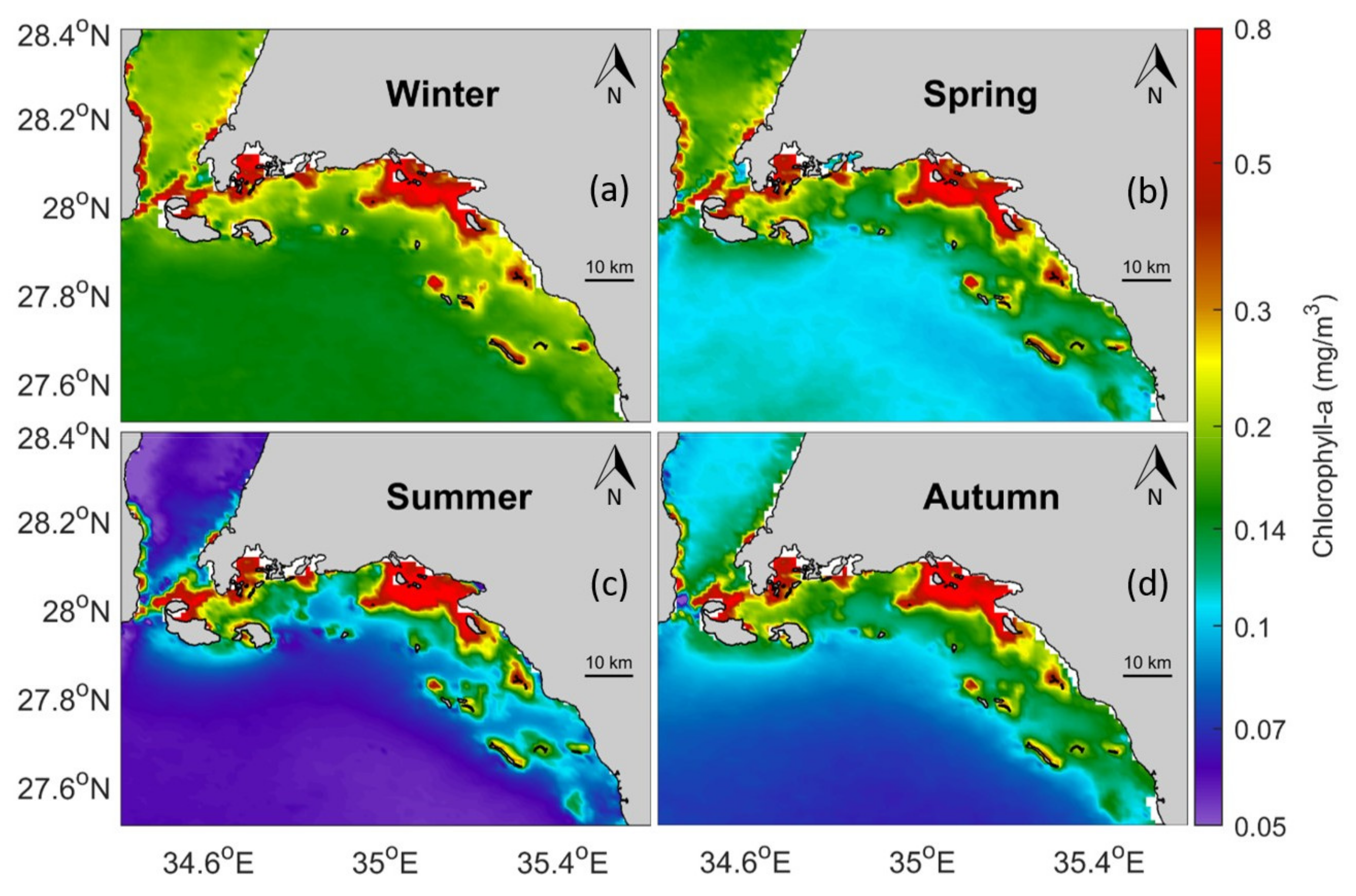
3.2. Remotely-Sensed Chl-a Observations
3.3. In Situ Observations
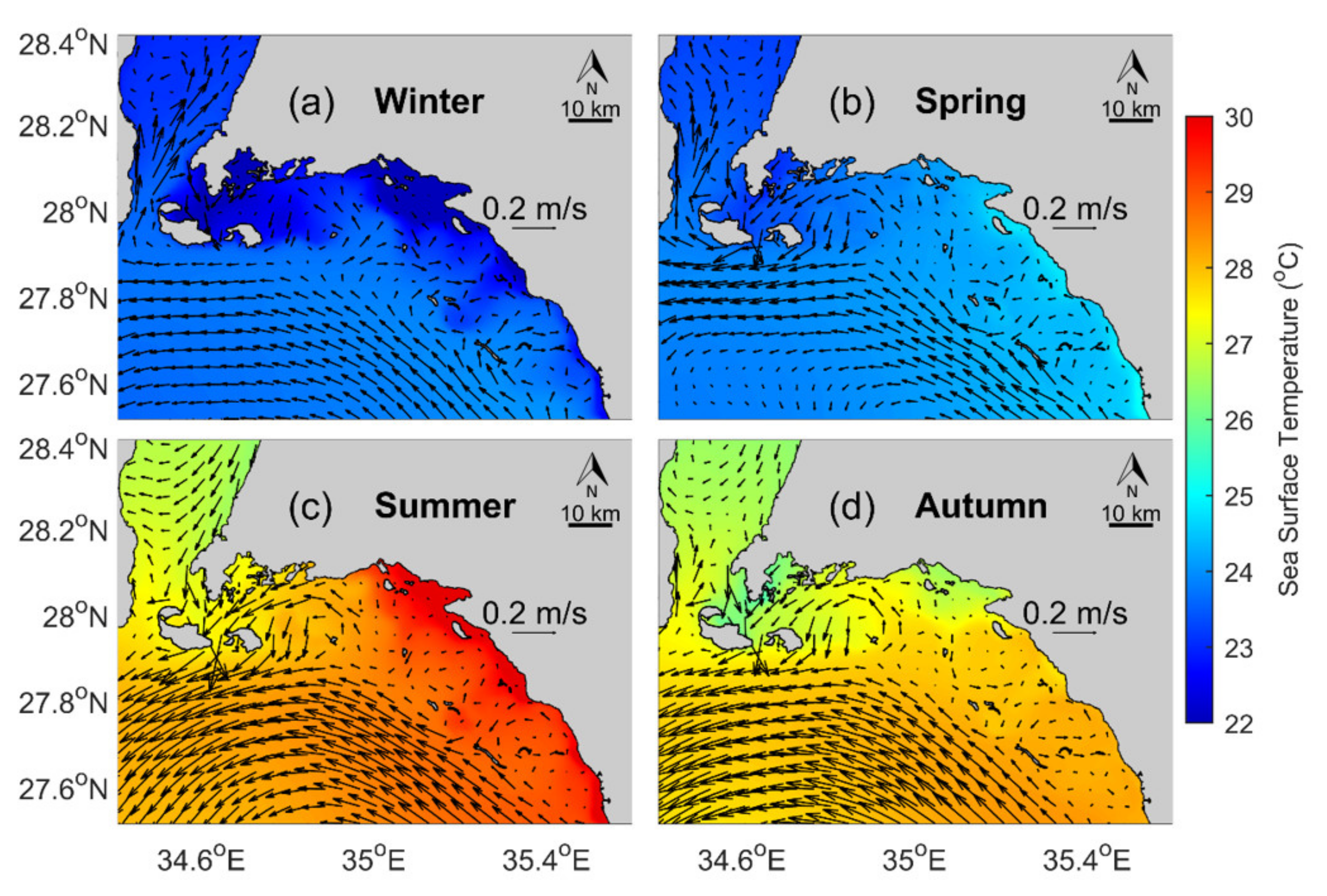
3.4. Oceanographic Setting
4. Discussion
5. Conclusions
Recommendations to NEOM Stakeholders
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gladstone, W.; Tawfiq, N.; Nasr, D.; Andersen, I.; Cheung, C.; Drammeh, H.; Krupp, F.; Lintner, S. Sustainable use of renewable resources and conservation in the Red Sea and Gulf of Aden: Issues, needs and strategic actions. Ocean Coast. Manag. 1999, 42, 671–697. [Google Scholar] [CrossRef]
- Gerges, M.A. The red sea and gulf of aden action plan—Facing the challenges of an ocean gateway. Ocean Coast. Manag. 2002, 45, 885–903. [Google Scholar] [CrossRef]
- Hughes, T.P.; Bellwood, D.R.; Connolly, S.R. Biodiversity hotspots, centres of endemicity, and the conservation of coral reefs. Ecol. Lett. 2002, 5, 775–784. [Google Scholar] [CrossRef]
- Roberts, C.M. Marine Biodiversity hotspots and conservation priorities for tropical reefs. Science 2002, 295, 1280–1284. [Google Scholar] [CrossRef]
- Belkin, I.M. Rapid warming of Large Marine Ecosystems. Prog. Oceanogr. 2009, 81, 207–213. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Hoteit, I.; Prihartato, P.K.; Chronis, T.; Triantafyllou, G.; Abualnaja, Y. Abrupt warming of the Red Sea. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Carvalho, S.; Kürten, B.; Krokos, G.; Hoteit, I.; Ellis, J. The Red Sea. In World Seas: An Environmental Evaluation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 49–74. [Google Scholar]
- Fine, M.; Gildor, H.; Genin, A. A coral reef refuge in the Red Sea. Glob. Chang. Biol. 2013, 19, 3640–3647. [Google Scholar] [CrossRef] [PubMed]
- Osman, E.O.; Smith, D.J.; Ziegler, M.; Kürten, B.; Conrad, C.; El-Haddad, K.M.; Voolstra, C.R.; Suggett, D.J. Thermal refugia against coral bleaching throughout the northern Red Sea. Glob. Chang. Biol. 2018, 24, e474–e484. [Google Scholar] [CrossRef]
- Chaidez, V.; Dreano, D.; Agusti, S.; Duarte, C.M.; Hoteit, I. Decadal trends in Red Sea maximum surface temperature. Sci. Rep. 2017, 7, 8144. [Google Scholar] [CrossRef]
- Krokos, G.; Papadopoulos, V.P.; Sofianos, S.; Hoteit, I. Assessing the long-term SST response of the Red Sea to natural climate variability. In Proceedings of the 2018 Ocean Sciences Meeting, Portland, OR, USA, 11–16 February 2018. [Google Scholar]
- Gittings, J.A.; Raitsos, D.E.; Krokos, G.; Hoteit, I. Impacts of warming on phytoplankton abundance and phenology in a typical tropical marine ecosystem. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Genevier, L.G.C.; Jamil, T.; Raitsos, D.E.; Krokos, G.; Hoteit, I. Marine heatwaves reveal coral reef zones susceptible to bleaching in the Red Sea. Glob. Chang. Biol. 2019, 25, 2338–2351. [Google Scholar] [CrossRef] [PubMed]
- Krokos, G.; Papadopoulos, V.P.; Sofianos, S.S.; Ombao, H.; Dybczak, P.; Hoteit, I. Natural Climate Oscillations may Counteract Red Sea Warming Over the Coming Decades. Geophys. Res. Lett. 2019, 46, 3454–3461. [Google Scholar] [CrossRef]
- Furby, K.A.; Bouwmeester, J.; Berumen, M.L. Susceptibility of central Red Sea corals during a major bleaching event. Coral Reefs 2013, 32, 505–513. [Google Scholar] [CrossRef]
- Pineda, J.; Starczak, V.; Tarrant, A.; Blythe, J.; Davis, K.; Farrar, T.; Berumen, M.; Da Silva, J.C.B. Two spatial scales in a bleaching event: Corals from the mildest and the most extreme thermal environments escape mortality. Limnol. Oceanogr. 2013, 58, 1531–1545. [Google Scholar] [CrossRef]
- Acker, J.; Leptoukh, G.; Shen, S.; Zhu, T.; Kempler, S. Remotely-sensed chlorophyll a observations of the northern Red Sea indicate seasonal variability and influence of coastal reefs. J. Mar. Syst. 2008, 69, 191–204. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Pradhan, Y.; Brewin, R.J.W.; Stenchikov, G.; Hoteit, I. Remote sensing the phytoplankton seasonal succession of the Red Sea. PLoS ONE 2013, 8, e64909. [Google Scholar] [CrossRef] [PubMed]
- Raitsos, D.E.; Yi, X.; Platt, T.; Racault, M.F.; Brewin, R.J.W.; Pradhan, Y.; Papadopoulos, V.P.; Sathyendranath, S.; Hoteit, I. Monsoon oscillations regulate fertility of the Red Sea. Geophys. Res. Lett. 2015, 42, 855–862. [Google Scholar] [CrossRef]
- Gittings, J.A.; Raitsos, D.E.; Kheireddine, M.; Racault, M.-F.; Claustre, H.; Hoteit, I. Evaluating tropical phytoplankton phenology metrics using contemporary tools. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Kosobokova, K.N. The reproductive cycle and life history of the Arctic copepod Calanus glacialis in the White Sea. Polar Biol. 1999, 22, 254–263. [Google Scholar] [CrossRef]
- Hays, G.; Richardson, A.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Khan, M.A.; Muqtadir, A. Distribution of mangroves along the Red Sea coast of the Arabian Peninsula: Part-I: The northern coast of western Saudi Arabia. Earth Sci. India 2010, 3, 28–42. [Google Scholar]
- Almahasheer, H.; Aljowair, A.; Duarte, C.M.; Irigoien, X. Decadal stability of Red Sea mangroves. Estuar. Coast. Shelf Sci. 2016, 169, 164–172. [Google Scholar] [CrossRef]
- Pearce, D.W. The value of biodiversity. Microb. Divers. Bioprospecting 2003, 469–475. [Google Scholar] [CrossRef]
- Turpie, J.K.; Heydenrych, B.J.; Lamberth, S.J. Economic value of terrestrial and marine biodiversity in the Cape Floristic Region: Implications for defining effective and socially optimal conservation strategies. Biol. Conserv. 2003, 112, 233–251. [Google Scholar] [CrossRef]
- Beaumont, N.J.; Austen, M.C.; Mangi, S.C.; Townsend, M. Economic valuation for the conservation of marine biodiversity. Mar. Pollut. Bull. 2008, 56, 386–396. [Google Scholar] [CrossRef]
- Rees, S.E.; Rodwell, L.D.; Attrill, M.J.; Austen, M.C.; Mangi, S.C. The value of marine biodiversity to the leisure and recreation industry and its application to marine spatial planning. Mar. Policy 2010, 34, 868–875. [Google Scholar] [CrossRef]
- Alsarhan, A.; Zatari, T.; Al-Asaly, M.; Mirza, K.; Harthi, A.; Othman, M.; Babiker, M.; Khan, A.; Aljabr, A.; Albuqami, F. Third National Communication of the Kingdom of Saudi Arabia; Designated National Authority (DNA) of Saudi Arabia: Riyadh, Saudi Arabia, 2016. [Google Scholar]
- Hoepner, T.; Lattemann, S. Chemical impacts from seawater desalination plants—A case study of the northern Red Sea. Desalination 2003, 152, 133–140. [Google Scholar] [CrossRef]
- Gokul, E.A.; Raitsos, D.E.; Gittings, J.A.; Hoteit, I. Developing an Atlas of harmful algal blooms in the Red Sea: Linkages to local aquaculture. Remote Sens. 2020, 12, 3695. [Google Scholar] [CrossRef]
- Genin, A.; Lazar, B.; Brenner, S. Vertical mixing and coral death in the Red Sea following the eruption of Mount Pinatubo. Nature 1995, 377, 507–510. [Google Scholar] [CrossRef]
- Platt, T.; Sathyendranath, S. Ecological indicators for the pelagic zone of the ocean from remote sensing. Remote Sens. Environ. 2008, 112, 3426–3436. [Google Scholar] [CrossRef]
- Ludwig, J.A.; QUARTET, L.; Reynolds, J.F.; Reynolds, J. Statistical Ecology: A Primer in Methods and Computing; John Wiley & Sons: Hoboken, NJ, USA, 1988; Volume 1. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Heink, U.; Kowarik, I. What criteria should be used to select biodiversity indicators? Biodivers. Conserv. 2010, 19, 3769–3797. [Google Scholar] [CrossRef]
- McQuatters-Gollop, A.; Raitsos, D.E.; Edwards, M.; Pradhan, Y.; Mee, L.D.; Lavender, S.J.; Attrill, M.J. A long-term chlorophyll dataset reveals regime shift in North Sea phytoplankton biomass unconnected to nutrient levels. Limnol. Oceanogr. 2007, 52, 635–648. [Google Scholar] [CrossRef]
- Racault, M.-F.; Platt, T.; Sathyendranath, S.; Irba, A.E.; Martinez Vicente, V.; Brewin, R.J.W. Plankton indicators and ocean observing systems: Support to the marine ecosystem state assessment. J. Plankton Res. 2014, 36, 621–629. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar]
- Beaugrand, G.; Brander, K.M.; Alistair Lindley, J.; Souissi, S.; Reid, P.C. Plankton effect on cod recruitment in the North Sea. Nature 2003, 426, 661–664. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Richardson, A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef]
- Platt, T.; Fuentes-Yaco, C.; Frank, K.T. Spring algal bloom and larval fish survival. Nature 2003, 423, 398–399. [Google Scholar] [CrossRef]
- Koeller, P.; Fuentes-Yaco, C.; Platt, T.; Sathyendranath, S.; Richards, A.; Ouellet, P.; Orr, D.; Skuladottir, U.; Wieland, K.; Savard, L.; et al. Basin-scale coherence in phenology of shrimps and phytoplankton in the North Atlantic Ocean. Science 2009, 324, 791–793. [Google Scholar] [CrossRef]
- Cushing, D.H. Plankton production and year-class strength in fish populations: An update of the match/mismatch hypothesis. Adv. Mar. Biol. 1990, 26, 249–293. [Google Scholar] [CrossRef]
- Racault, M.-F.; Sathyendranath, S.; Brewin, R.J.W.; Raitsos, D.E.; Jackson, T.; Platt, T. Impact of El Niño Variability on Oceanic Phytoplankton. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Sathyendranath, S.; Brewin, R.J.W.; Brockmann, C.; Brotas, V.; Calton, B.; Chuprin, A.; Cipollini, P.; Couto, A.; Dingle, J.; Doerffer, R.; et al. An Ocean-colour time series for use in climate studies: The Experience of the Ocean-Colour Climate Change Initiative (OC-CCI). Sensors 2019, 19, 4285. [Google Scholar] [CrossRef]
- Racault, M.-F.; Raitsos, D.E.; Berumen, M.L.; Brewin, R.J.W.; Platt, T.; Sathyendranath, S.; Hoteit, I. Phytoplankton phenology indices in coral reef ecosystems: Application to ocean-color observations in the Red Sea. Remote Sens. Environ. 2015, 160, 222–234. [Google Scholar] [CrossRef]
- Kheireddine, M.; Ouhssain, M.; Calleja, M.L.; Morán, X.A.G.; Sarma, Y.V.B.; Tiwari, S.P.; Jones, B.H. Characterization of light absorption by chromophoric dissolved organic matter (CDOM) in the upper layer of the Red Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 133, 72–84. [Google Scholar] [CrossRef]
- Brewin, R.J.W.; Raitsos, D.E.; Pradhan, Y.; Hoteit, I. Comparison of chlorophyll in the Red Sea derived from MODIS-Aqua and in vivo fluorescence. Remote Sens. Environ. 2013, 136, 218–224. [Google Scholar] [CrossRef]
- Brewin, R.J.W.; Morán, X.A.G.; Raitsos, D.E.; Gittings, J.A.; Calleja, M.L.; Viegas, M.; Ansari, M.I.; Al-Otaibi, N.; Huete-Stauffer, T.M.; Hoteit, I. Factors regulating the relationship between total and size-fractionated chlorophyll-a in coastal waters of the Red Sea. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Brewin, R.J.W.; Raitsos, D.E.; Dall’Olmo, G.; Zarokanellos, N.; Jackson, T.; Racault, M.-F.; Boss, E.S.; Sathyendranath, S.; Jones, B.H.; Hoteit, I. Regional ocean-colour chlorophyll algorithms for the Red Sea. Remote Sens. Environ. 2015, 165, 64–85. [Google Scholar] [CrossRef]
- Ameer, A.E. NEOM: Introducing a new era of sustainable development. In Proceedings of the Red Sea Research Center Open Science Conference, Thuwal, Saudi Arabia, 29–30 October 2019. [Google Scholar]
- Sathyendranath, S. Remote Sensing of Ocean Colour in Coastal, and Other Optically-Complex, Waters; International Ocean Colour Coordinating Group (IOCCG): Dartmouth, NS, Canada, 2000. [Google Scholar]
- Jackson, T.; Sathyendranath, S.; Mélin, F. An improved optical classification scheme for the Ocean Colour Essential Climate Variable and its applications. Remote Sens. Environ. 2017, 203, 152–161. [Google Scholar] [CrossRef]
- Jacobs, Z.L.; Jebri, F.; Raitsos, D.E.; Popova, E.; Srokosz, M.; Painter, S.C.; Nencioli, F.; Roberts, M.; Kamau, J.; Palmer, M.; et al. Shelf-Break Upwelling and Productivity Over the North Kenya Banks: The Importance of Large-Scale Ocean Dynamics. J. Geophys. Res. Ocean. 2020, 125. [Google Scholar] [CrossRef]
- Rasheed, M.; Badran, M.; Richter, C.; Huettel, M. Effect of reef framework and bottom sediment on nutrient enrichment in a coral reef of the Gulf of Aqaba, Red Sea. Mar. Ecol. Prog. Ser. 2002, 239, 277–285. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Brewin, R.J.W.; Zhan, P.; Dreano, D.; Pradhan, Y.; Nanninga, G.B.; Hoteit, I. Sensing coral reef connectivity pathways from space. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Marshall, J.; Adcroft, A.; Hill, C.; Perelman, L.; Heisey, C. A finite-volume, incompressible Navier Stokes model for studies of the ocean on parallel computers. J. Geophys. Res. Oceans 1997, 102, 5753–5766. [Google Scholar] [CrossRef]
- Krokos, G.; Cerovecki, I.; Zhan, P.; Hoteit, I.; Hendershott, M.C. Processes controlling the seasonal evolution of Mixed Layers in the Red Sea. In Proceedings of the Ocean Sciences Meeting 2020, San Diego, CA, USA, 16–21 February 2020. [Google Scholar]
- Viswanadhapalli, Y.; Dasari, H.P.; Langodan, S.; Challa, V.S.; Hoteit, I. Climatic features of the Red Sea from a regional assimilative model. Int. J. Clim. 2017, 37, 2563–2581. [Google Scholar] [CrossRef]
- Yao, F.; Hoteit, I.; Pratt, L.J.; Bower, A.S.; Zhai, P.; Köhl, A.; Gopalakrishnan, G. Seasonal overturning circulation in the Red Sea: 1. Model validation and summer circulation. J. Geophys. Res. Ocean. 2014, 119, 2238–2262. [Google Scholar] [CrossRef]
- Yao, F.; Hoteit, I.; Pratt, L.J.; Bower, A.S.; Köhl, A.; Gopalakrishnan, G.; Rivas, D. Seasonal overturning circulation in the Red Sea: 2. Winter circulation. J. Geophys. Res. Ocean. 2014, 119, 2263–2289. [Google Scholar] [CrossRef]
- Zhan, P.; Subramanian, A.C.; Yao, F.; Hoteit, I. Eddies in the Red Sea: A statistical and dynamical study. J. Geophys. Res. Ocean. 2014, 119, 3909–3925. [Google Scholar] [CrossRef]
- Zhan, P.; Krokos, G.; Guo, D.; Hoteit, I. Three-dimensional signature of the Red Sea Eddies and eddy-induced transport. Geophys. Res. Lett. 2019, 46, 2167–2177. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Lorenzen, C.J.; Holmes, R.W.; Strickland, J.D.H. Fluorometric Determination of Chlorophyll. ICES J. Mar. Sci. 1965, 30, 3–15. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, QC, Canada, 1972. [Google Scholar]
- Bruckner, A.; Rowlands, G.; Riegl, B.; Purkis, S.J.; Williams, A.; Renaud, P. Khaled bin Sultan Living Oceans Foundation Atlas of Saudi Arabian Red Sea Marine Habitats; Panoramic Press: Phoenix, AZ, USA, 2013; p. 262. [Google Scholar]
- Kjerfve, B. Coastal lagoons. In Elsevier Oceanography Series; Elsevier: Amsterdam, The Netherlands, 1994; Volume 60, pp. 1–8. [Google Scholar]
- Dreano, D.; Raitsos, D.E.; Gittings, J.A.; Krokos, G.; Hoteit, I. The Gulf of Aden Intermediate Water Intrusion Regulates the Southern Red Sea Summer Phytoplankton Blooms. PLoS ONE 2016, 11, e0168440. [Google Scholar] [CrossRef]
- Gittings, J.A.; Raitsos, D.E.; Racault, M.-F.; Brewin, R.J.W.; Pradhan, Y.; Sathyendranath, S.; Platt, T. Seasonal phytoplankton blooms in the Gulf of Aden revealed by remote sensing. Remote Sens. Env. 2017, 189, 56–66. [Google Scholar] [CrossRef]
- Al-Naimi, N.; Raitsos, D.E.; Ben-Hamadou, R.; Soliman, Y. Evaluation of Satellite retrievals of chlorophyll-a in the Arabian Gulf. Remote Sens. 2017, 9, 301. [Google Scholar] [CrossRef]
- Levanon-Spanier, I.; Padan, E.; Reiss, Z. Primary production in a desert-enclosed sea—The Gulf of Elat (Aqaba), Red Sea. Deep Sea Res. Part A Oceanogr. Res. Pap. 1979, 26, 673–685. [Google Scholar] [CrossRef]
- Labiosa, R.G.; Arrigo, K.R.; Genin, A.; Monismith, S.G.; Van Dijken, G. The interplay between upwelling and deep convective mixing in determining the seasonal phytoplankton dynamics in the Gulf of Aqaba: Evidence from SeaWiFS and MODIS. Limnol. Oceanogr. 2003, 48, 2355–2368. [Google Scholar] [CrossRef]
- Al-Najjar, T.; Badran, M.I.; Richter, C.; Meyerhoefer, M.; Sommer, U. Seasonal dynamics of phytoplankton in the Gulf of Aqaba, Red Sea. Hydrobiologia 2007, 579, 69–83. [Google Scholar] [CrossRef]
- Tilstra, A.; Van Hoytema, N.; Cardini, U.; Bednarz, V.N.; Rix, L.; Naumann, M.S.; Al-Horani, F.A.; Wild, C. Effects of Water Column Mixing and Stratification on Planktonic Primary Production and Dinitrogen Fixation on a Northern Red Sea Coral Reef. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Weikert, H. Plankton and the pelagic environment. Red Sea 1987, 90–111. [Google Scholar] [CrossRef]
- Devassy, R.P.; El-Sherbiny, M.M.; Al-Sofyani, A.M.; Al-Aidaroos, A.M. Spatial variation in the phytoplankton standing stock and diversity in relation to the prevailing environmental conditions along the Saudi Arabian coast of the northern Red Sea. Mar. Biodivers. 2017, 47, 995–1008. [Google Scholar] [CrossRef]
- Doney, S.C. Plankton in a warmer world. Nature 2006, 444, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Sofianos, S.S. An Oceanic General Circulation Model (OGCM) investigation of the Red Sea circulation: 2. Three-dimensional circulation in the Red Sea. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef]
- Polovina, J.J.; Mitchum, G.T.; Evans, G.T. Decadal and basin-scale variation in mixed layer depth and the impact on biological production in the Central and North Pacific, 1960–1988. Deep Sea Res. Part I Oceanogr. Res. Pap. 1995, 42, 1701–1716. [Google Scholar] [CrossRef]
- Papadopoulos, V.P.; Zhan, P.; Sofianos, S.S.; Raitsos, D.E.; Qurban, M.; Abualnaja, Y.; Bower, A.; Kontoyiannis, H.; Pavlidou, A.; Asharaf, T.T.M.; et al. Factors governing the deep ventilation of the Red Sea. J. Geophys. Res. Ocean. 2015, 120, 7493–7505. [Google Scholar] [CrossRef]
- Asfahani, K.; Krokos, G.; Papadopoulos, V.P.; Jones, B.H.; Sofianos, S.; Kheireddine, M.; Hoteit, I. Capturing a Mode of Intermediate Water Formation in the Red Sea. J. Geophys. Res. Ocean. 2020, 125, e2019JC015803. [Google Scholar] [CrossRef]
- Moeller, C.C.; Huh, O.K.; Roberts, H.H.; Gumley, L.E.; Menzel, W.P. Response of Louisiana coastal environments to a cold front passage. J. Coast. Res. 1993, 9, 434–447. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papagiannopoulos, N.; Raitsos, D.E.; Krokos, G.; Gittings, J.A.; Brewin, R.J.W.; Papadopoulos, V.P.; Pavlidou, A.; Selmes, N.; Groom, S.; Hoteit, I. Phytoplankton Biomass and the Hydrodynamic Regime in NEOM, Red Sea. Remote Sens. 2021, 13, 2082. https://doi.org/10.3390/rs13112082
Papagiannopoulos N, Raitsos DE, Krokos G, Gittings JA, Brewin RJW, Papadopoulos VP, Pavlidou A, Selmes N, Groom S, Hoteit I. Phytoplankton Biomass and the Hydrodynamic Regime in NEOM, Red Sea. Remote Sensing. 2021; 13(11):2082. https://doi.org/10.3390/rs13112082
Chicago/Turabian StylePapagiannopoulos, Nikolaos, Dionysios E. Raitsos, Georgios Krokos, John A. Gittings, Robert J. W. Brewin, Vassilis P. Papadopoulos, Alexandra Pavlidou, Nick Selmes, Steve Groom, and Ibrahim Hoteit. 2021. "Phytoplankton Biomass and the Hydrodynamic Regime in NEOM, Red Sea" Remote Sensing 13, no. 11: 2082. https://doi.org/10.3390/rs13112082
APA StylePapagiannopoulos, N., Raitsos, D. E., Krokos, G., Gittings, J. A., Brewin, R. J. W., Papadopoulos, V. P., Pavlidou, A., Selmes, N., Groom, S., & Hoteit, I. (2021). Phytoplankton Biomass and the Hydrodynamic Regime in NEOM, Red Sea. Remote Sensing, 13(11), 2082. https://doi.org/10.3390/rs13112082