Longitudinal, Lateral, Vertical, and Temporal Thermal Heterogeneity in a Large Impounded River: Implications for Cold-Water Refuges

 and
and
Abstract
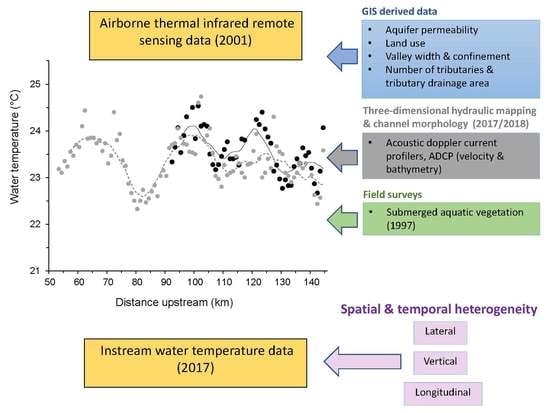
1. Introduction
- (1)
- Compare longitudinal patterns of water temperature derived from thermal infrared imagery to instream measurements in the water column.
- (2)
- Assess spatial (lateral, longitudinal, and vertical) and temporal thermal heterogeneity and potential cold-water refuges for salmonids at multiple scales.
- (3)
- Evaluate statistical associations between hydrologic and geomorphic variables and occurrence of cool-water areas and median water temperature.
2. Materials and Methods
2.1. Study Area
2.2. Water Temperature Data
2.2.1. Airborne Thermal Infrared (TIR) Imagery
2.2.2. Instream Measurement
2.3. Characterization of Riverine Landscape
2.3.1. Geographical Information System
Land Use
Aquifer Permeability
Valley Width and Confinement
Tributaries
2.3.2. Three-Dimensional Hydraulic Mapping and Channel Morphology
Water Depth and Velocity
Transect Velocity and Discharge Calculations
Submerged Aquatic Vegetation
2.4. Data Analysis
2.4.1. Spatial and Temporal Analysis
2.4.2. Statistical Analysis
Generalized Additive Modeling
3. Results
3.1. Comparisons between TIR and Instream Measurements
3.2. Characterization of Spatial and Temporal Thermal Heterogeneity
3.2.1. Longitudinal
3.2.2. Lateral: Tributary Confluences
3.2.3. Vertical: Water Column
3.2.4. Temporal
3.2.5. Implications for Potential Cold-Water Refuges
3.3. Characterization of Riverine Landscape
3.3.1. GIS Data
3.3.2. Three-Dimensional Hydraulic Mapping and Channel Morphology
3.3.3. Submerged Aquatic Vegetation
3.3.4. Associations between Hydrologic and Geomorphic Variables and TIR Imagery
4. Discussion
5. Conclusions
Supplementary Materials
Additional Online Resources
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Ebersole, J.L.; Liss, W.J.; Frissell, C.A. Thermal heterogeneity, stream channel morphology, and salmonid abundance in northeastern Oregon streams. Can. J. Fish. Aquat. Sci. 2003, 60, 1266–1280. [Google Scholar] [CrossRef]
- Steel, E.A.; Beechie, T.J.; Torgersen, C.E.; Fullerton, A.H. Envisioning, quantifying, and managing thermal regimes on river networks. Bioscience 2017, 67, 506–522. [Google Scholar] [CrossRef]
- Ward, J.V.; Stanford, J.A. The serial discontinuity of lotic ecosystems. In Dynamics of Lotic Ecosystems; Fontaine, T.D., Bartell, S.M., Eds.; Ann Arbor Science Publishers: Ann Arbor, MI, USA, 1983; pp. 29–42. [Google Scholar]
- Stanford, J.A.; Ward, J.V.; Liss, W.J.; Frissell, C.A.; Williams, R.N.; Lichatowich, J.A.; Coutant, C.C. A general protocol for restoration of regulated rivers. Regul. Rivers Res. Manag. 1996, 12, 391–413. [Google Scholar] [CrossRef]
- Olden, J.D.; Naiman, R.J. Incorporating thermal regimes into environmental flows assessments: Modifying dam operations to restore freshwater ecosystem integrity. Freshw. Biol. 2010, 55, 86–107. [Google Scholar] [CrossRef]
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Sabo, J.L.; Bestgen, K.; Graf, W.; Sinha, T.; Wohl, E.E. Dams in the cadillac desert: Downstream effects in a geomorphic context. Ann. N. Y. Acad. Sci. 2012, 1249, 227–246. [Google Scholar] [CrossRef]
- Prats, J.; Armengol, J.; Marcé, R.; Sánchez-Juny, M.; Dolz, J. Dams and reservoirs in the lower Ebro river and its effects on the river thermal cycle. In The Ebro River Basin; Barceló, D., Petrovic, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 77–95. [Google Scholar]
- Graf, W.L. Downstream hydrologic and geomorphic effects of large dams on American rivers. Geomorphology 2006, 79, 336–360. [Google Scholar] [CrossRef]
- Kondolf, G.M. Hungry water: Effects of dams and gravel mining on river channels. Environ. Manag. 1997, 21, 533–551. [Google Scholar] [CrossRef]
- Sabo, J.L.; Caron, M.; Doucett, R.; Dibble, K.L.; Ruhi, A.; Marks, J.C.; Hungate, B.A.; Kennedy, T.A.; Siqueira, T. Pulsed flows, tributary inputs and food-web structure in a highly regulated river. J. Appl. Ecol. 2018, 55, 1884–1895. [Google Scholar] [CrossRef]
- Brandt, S.A. Classification of geomorphological effects downstream of dams. Catena 2000, 40, 375–401. [Google Scholar] [CrossRef]
- Poole, G.C.; Berman, C.H. An ecological perspective on in-stream temperature: Natural heat dynamics and mechanisms of human-caused thermal degradation. Environ. Manag. 2001, 27, 787–802. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Lodge, D.M. Effects of submersed macrophytes on ecosystem processes. Aquat. Bot. 1986, 26, 341–370. [Google Scholar] [CrossRef]
- Torgersen, C.E.; Price, D.M.; Li, H.W.; McIntosh, B.A. Multiscale thermal refugia and stream habitat associations of chinook salmon in northeastern Oregon. Ecol. Appl. 1999, 9, 301–319. [Google Scholar] [CrossRef]
- Torgersen, C.E.; Ebersole, J.L.; Keenan, D.M. Primer for Identifying Cold-Water Refuges to Protect and Restore Thermal Diversity in Riverine Landscapes; EPA 910-C-12-001; U.S. Environmental Protection Agency: Seattle, WA, USA, 2012; p. 91.
- Dugdale, S.J.; Bergeron, N.E.; St-Hilaire, A. Temporal variability of thermal refuges and water temperature patterns in an Atlantic salmon river. Remote Sens. Environ. 2013, 136, 358–373. [Google Scholar] [CrossRef]
- Rice, S.P.; Greenwood, M.T.; Joyce, C.B. Tributaries, sediment sources, and the longitudinal organisation of macroinvertebrate fauna along river systems. Can. J. Fish. Aquat. Sci. 2001, 58, 824–840. [Google Scholar] [CrossRef]
- Vinson, M.R. Long-term dynamics of an invertebrate assemblage downstream from a large dam. Ecol. Appl. 2001, 11, 711–730. [Google Scholar] [CrossRef]
- Benda, L.; Andras, K.; Miller, D.; Bigelow, P. Confluence effects in rivers: Interactions of basin scale, network geometry, and disturbance regimes. Water Resour. Res. 2004, 40, W05402. [Google Scholar] [CrossRef]
- Dugdale, S.J.; Bergeron, N.E.; St-Hilaire, A. Spatial distribution of thermal refuges analysed in relation to riverscape hydromorphology using airborne thermal infrared imagery. Remote Sens. Environ. 2015, 160, 43–55. [Google Scholar] [CrossRef]
- Knighton, D. Fluvial Forms and Processes: A New Perspective; Oxford University Press, Inc.: New York, NY, USA, 1998; p. 383. [Google Scholar]
- Torgersen, C.E.; Gresswell, R.E.; Bateman, D.S.; Burnett, K.M. Spatial identification of tributary impacts in river networks. In River Confluences, Tributaries and the Fluvial Network; Rice, S.P., Roy, A.G., Rhoads, B.L., Eds.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2008; pp. 159–181. [Google Scholar]
- Cristea, N.C.; Burges, S.J. Use of thermal infrared imagery to complement monitoring and modeling of spatial stream temperatures. J. Hydrol. Eng. 2009, 14, 1080–1090. [Google Scholar] [CrossRef]
- Fullerton, A.H.; Torgersen, C.E.; Lawler, J.J.; Faux, R.N.; Steel, E.A.; Beechie, T.J.; Ebersole, J.L.; Leibowitz, S.G. Rethinking the longitudinal stream temperature paradigm: Region-wide comparison of thermal infrared imagery reveals unexpected complexity of river temperatures. Hydrol. Process. 2015, 29, 4719–4737. [Google Scholar] [CrossRef]
- Maheu, A.; St-Hilaire, A.; Caissie, D.; El-Jabi, N. Understanding the thermal regime of rivers influenced by small and medium size dams in Eastern Canada. River Res. Appl. 2016, 32, 2032–2044. [Google Scholar] [CrossRef]
- Dzara, J.R.; Neilson, B.T.; Null, S.E. Quantifying thermal refugia connectivity by combining temperature modeling, distributed temperature sensing, and thermal infrared imaging. Hydrol. Earth Syst. Sci. 2019, 23, 2965–2982. [Google Scholar] [CrossRef]
- Vatland, S.J.; Gresswell, R.E.; Poole, G.C. Quantifying stream thermal regimes at multiple scales: Combining thermal infrared imagery and stationary stream temperature data in a novel modeling framework. Water Resour. Res. 2015, 51, 31–46. [Google Scholar] [CrossRef]
- Arscott, D.B.; Tockner, K.; Ward, J.V. Thermal heterogeneity along a braided floodplain river (Tagliamento River, northeastern Italy). Can. J. Fish. Aquat. Sci. 2001, 58, 2359–2373. [Google Scholar] [CrossRef]
- Imholt, C.; Soulsby, C.; Malcolm, I.A.; Hrachowitz, M.; Gibbins, C.N.; Langan, S.; Tetzlaff, D. Influence of scale on thermal characteristics in a large montane river basin. River Res. Appl. 2013, 29, 403–419. [Google Scholar] [CrossRef]
- Steel, E.A.; Lange, I.A. Using wavelet analysis to detect changes in water temperature regimes at multiple scales: Effects of multi-purpose dams in the Willamette River basin. River Res. Appl. 2007, 23, 351–359. [Google Scholar] [CrossRef]
- Wawrzyniak, V.; Piégay, H.; Poirel, A. Longitudinal and temporal thermal patterns of the French Rhône River using Landsat ETM+ thermal infrared images. Aquat. Sci. 2011, 74, 405–414. [Google Scholar] [CrossRef]
- Ling, F.; Foody, G.; Du, H.; Ban, X.; Li, X.; Zhang, Y.; Du, Y. Monitoring thermal pollution in rivers downstream of dams with Landsat ETM+ thermal infrared images. Remote Sens. 2017, 9, 1175. [Google Scholar] [CrossRef]
- Keefer, M.L.; Clabough, T.S.; Jepson, M.A.; Johnson, E.L.; Peery, C.A.; Caudill, C.C. Thermal exposure of adult Chinook salmon and steelhead: Diverse behavioral strategies in a large and warming river system. PLoS ONE 2018, 13, e0204274. [Google Scholar] [CrossRef]
- Keefer, M.L.; Clabough, T.S.; Jepson, M.A.; Bowerman, T.; Caudill, C.C. Temperature and depth profiles of Chinook salmon and the energetic costs of their long-distance homing migrations. J. Therm. Biol. 2019, 79, 155–165. [Google Scholar] [CrossRef]
- Snyder, M.N.; Schumaker, N.H.; Ebersole, J.L.; Dunham, J.B.; Comeleo, R.L.; Keefer, M.L.; Leinenbach, P.; Brookes, A.; Cope, B.; Wu, J.; et al. Individual based modeling of fish migration in a 2-D river system: Model description and case study. Landsc. Ecol. 2019, 34, 737–754. [Google Scholar] [CrossRef]
- Adams, S.B.; Bjornn, T.C. Bull trout distributions related to temperature regimes in four central Idaho streams. In Friends of the Bull Trout Conference; MacKay, W.C., Brewin, M.K., Monita, M., Eds.; Bull Trout Task Force: Calgary, AB, Canada, 1997; pp. 371–380. [Google Scholar]
- Selong, J.H.; McMahon, T.E.; Zale, A.V.; Barrows, F.T. Effect of temperature on growth and survival of bull Trout, with application of an improved method for determining thermal tolerance in fishes. Trans. Am. Fish. Soc. 2001, 130, 1026–1037. [Google Scholar] [CrossRef]
- Bear, E.A.; Mcmahon, T.E.; Zale, A.V. Comparative thermal requirements of westslope cutthroat trout and rainbow 11-out: Implications for species interactions and development of thermal protection standards. Trans. Am. Fish. Soc. 2007, 136, 1113–1121. [Google Scholar] [CrossRef]
- Mandeville, C.P.; Rahel, F.J.; Patterson, L.S.; Walters, A.W. Integrating fish assemblage data, modeled stream temperatures, and thermal tolerance metrics to develop thermal guilds for water temperature regulation: Wyoming case study. Trans. Am. Fish. Soc. 2019, 148, 739–754. [Google Scholar] [CrossRef]
- Howell, P.J.; Dunham, J.B.; Sankovich, P.M. Relationships between water temperatures and upstream migration, cold water refuge use, and spawning of adult bull trout from the Lostine River, Oregon, USA. Ecol. Freshw. Fish 2010, 19, 96–106. [Google Scholar] [CrossRef]
- Beechie, T.; Imaki, H. Predicting natural channel patterns based on landscape and geomorphic controls in the Columbia River basin, USA. Water Resour. Res. 2014, 50, 39–57. [Google Scholar] [CrossRef]
- Watershed Sciences Inc. Aerial Surveys in the Pend Oreille River Basin: Thermal Infrared and Color Videography; Watershed Sciences Inc.: Corvallis, OR, USA, 2002; p. 39. [Google Scholar]
- Torgersen, C.E.; Faux, R.N.; McIntosh, B.A.; Poage, N.J.; Norton, D.J. Airborne thermal remote sensing for water temperature assessment in rivers and streams. Remote Sens. Environ. 2001, 76, 386–398. [Google Scholar] [CrossRef]
- Mustafa, M.; Barnhart, B.; Babbar-Sebens, M.; Ficklin, D. Modeling landscape change effects on stream temperature using the soil and water assessment tool. Water 2018, 10, 1143. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Wigington, P.J.; Leibowitz, S.G.; Comeleo, R.L.; Sickle, J.V. Predicting the occurrence of cold-water patches at intermittent and ephemeral tributary confluences with warm rivers. Freshw. Sci. 2015, 34, 111–124. [Google Scholar] [CrossRef]
- Comeleo, R.L.; Wigington, P.J.; Leibowitz, S.G. Creation of a Digital Aquifer Permeability Map for the Pacific Northwest; EPA/600/R-14/431; U.S. Environmental Protection Agency: Corvallis, OR, USA, 2014; pp. 1–18.
- Bisson, P.A.; Montgomery, D.R.; Buffington, J.M. Valley segments, stream reaches, and channel units. In Methods in Stream Ecology; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 1, pp. 21–47. [Google Scholar]
- Benda, L.; Miller, D.; Andras, K.; Bigelow, P.; Reeves, G.; Michael, D. NetMap: A new tool in support of watershed science and resource management. For. Sci. 2007, 53, 206–219. [Google Scholar]
- Lorang, M.S.; Tonolla, D. Combining active and passive hydroacoustic techniques during flood events for rapid spatial mapping of bedload transport patterns in gravel-bed rivers. Fundam. Appl. Limnol. Arch. Hydrobiol. 2014, 184, 231–246. [Google Scholar] [CrossRef]
- Marotz, B.L.; Lorang, M.S. Pallid sturgeon larvae: The drift dispersion hypothesis. J. Appl. Ichthyol. 2018, 34, 373–381. [Google Scholar] [CrossRef]
- Welty, E.Z.; Torgersen, C.E.; Brenkman, S.J.; Duda, J.J.; Armstrong, J.B. Multiscale analysis of river networks using the R package linbin. N. Am. J. Fish. Manag. 2015, 35, 802–809. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. EPA Region 10 Guidance for Pacific Northwest State and Tribal Temperature Water Quality Standards; EPA 910-B-03-002; Office of Water: Seattle, WA, USA, 2003; p. 49.
- Greer, G.; Carlson, S.; Thompson, S. Evaluating definitions of salmonid thermal refugia using in situ measurements in the Eel River, Northern California. Ecohydrology 2019, 12, e2101. [Google Scholar] [CrossRef]
- Benjamin, J.R.; Heltzel, J.M.; Dunham, J.B.; Heck, M.; Banish, N. Thermal regimes, nonnative trout, and their influences on native bull trout in the Upper Klamath River Basin, Oregon. Trans. Am. Fish. Soc. 2016, 145, 1318–1330. [Google Scholar] [CrossRef]
- Howell, P.J. Changes in native bull trout and non-native brook trout distributions in the upper Powder River basin after 20 years, relationships to water temperature and implications of climate change. Ecol. Freshw. Fish 2018, 27, 710–719. [Google Scholar] [CrossRef]
- Barrows, M.G.; Anglin, D.R.; Sankovich, P.M.; Hudson, J.M.; Koch, R.C.; Skalicky, J.J.; Wills, D.A.; Silver, B.P. Use of the Mainstem Columbia and Lower Snake Rivers by Migratory Bull Trout; U.S. Fish and Wildlife Service, Columbia River Fisheries Program Office: Vancouver, WA, USA, 2016; p. 276.
- Fullerton, A.H.; Torgersen, C.E.; Lawler, J.J.; Steel, E.A.; Ebersole, J.L.; Lee, S.Y. Longitudinal thermal heterogeneity in rivers and refugia for coldwater species: Effects of scale and climate change. Aquat. Sci. 2018, 80, 1–15. [Google Scholar] [CrossRef]
- Guisan, A.; Edwards, T.C.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Wood, S.N. Fast stable direct fitting and smoothness selection for generalized additive models. J. R. Stat. Soc. B 2008, 70, 495–518. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models; Chapman and Hall/CRC: New York, NY, USA, 2017; p. 496. [Google Scholar]
- Amodio, S.; Aria, M.; D’Ambrosio, A. On concurvity in nonlinear and nonparametric regression models. Statistica 2014, 74, 85–98. [Google Scholar]
- Barton, K. Mu-MIn: Multi-Model Inference. R Package Version 0.12.2/r18. Available online: http://R-Forge.R-project.org/projects/mumin/ (accessed on 3 April 2019).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference. A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Tonolla, D.; Acuña, V.; Uehlinger, U.; Frank, T.; Tockner, K. Thermal heterogeneity in river floodplains. Ecosystems 2010, 13, 727–740. [Google Scholar] [CrossRef]
- Wawrzyniak, V.; Piégay, H.; Allemand, P.; Vaudor, L.; Grandjean, P. Prediction of water temperature heterogeneity of braided rivers using very high resolution thermal infrared (TIR) images. Int. J. Remote Sens. 2013, 34, 4812–4831. [Google Scholar] [CrossRef]
- Paillex, A.; Castella, E.; zu Ermgassen, P.; Gallardo, B.; Aldridge, D.C. Large river floodplain as a natural laboratory: Non-native macroinvertebrates benefit from elevated temperatures. Ecosphere 2017, 8, e01972. [Google Scholar] [CrossRef]
- Turner, L.; Erskine, W.D. Variability in the development, persistence and breakdown of thermal, oxygen and salt stratification on regulated rivers of southeastern Australia. River Res. Appl. 2005, 21, 151–168. [Google Scholar] [CrossRef]
- Wawrzyniak, V.; Piégay, H.; Allemand, P.; Vaudor, L.; Goma, R.; Grandjean, P. Effects of geomorphology and groundwater level on the spatio-temporal variability of riverine cold water patches assessed using thermal infrared (TIR) remote sensing. Remote Sens. Environ. 2016, 175, 337–348. [Google Scholar] [CrossRef]
- Jin, J.; Wells, S.A.; Liu, D.; Yang, G.; Zhu, S.; Ma, J.; Yang, Z. Effects of water level fluctuation on thermal stratification in a typical tributary bay of Three Gorges Reservoir, China. PeerJ 2019, 7, e6925. [Google Scholar] [CrossRef]
- Mejia, F.H.; Torgersen, C.E.; Berntsen, E.K. Water Temperature Data from the Pend Oreille River, Washington and Idaho, 2016–2018; U.S. Geological Survey: Reston, VA, USA, 2019. [CrossRef]
- O’Sullivan, A.M.; Devito, K.J.; Curry, R.A. The influence of landscape characteristics on the spatial variability of river temperatures. Catena 2019, 177, 70–83. [Google Scholar] [CrossRef]
- Monk, W.A.; Wilbur, N.M.; Curry, R.A.; Gagnon, R.; Faux, R.N. Linking landscape variables to cold water refugia in rivers. J. Environ. Manag. 2013, 118, 170–176. [Google Scholar] [CrossRef]
- Maheu, A.; St-Hilaire, A.; Caissie, D.; El-Jabi, N.; Bourque, G.; Boisclair, D. A regional analysis of the impact of dams on water temperature in medium-size rivers in eastern Canada. Can. J. Fish. Aquat. Sci. 2016, 73, 1885–1897. [Google Scholar] [CrossRef]
- Armstrong, J.B.; Schindler, D.E. Going with the flow: Spatial distributions of juvenile coho salmon track an annually shifting mosaic of water temperature. Ecosystems 2013, 16, 1429–1441. [Google Scholar] [CrossRef]
- Reside, A.E.; Briscoe, N.J.; Dickman, C.R.; Greenville, A.C.; Hradsky, B.A.; Kark, S.; Kearney, M.R.; Kutt, A.S.; Nimmo, D.G.; Pavey, C.R.; et al. Persistence through tough times: Fixed and shifting refuges in threatened species conservation. Biodivers. Conserv. 2019, 28, 1303–1330. [Google Scholar] [CrossRef]
- Kurylyk, B.L.; MacQuarrie, K.T.B.; Linnansaari, T.; Cunjak, R.A.; Curry, R.A. Preserving, augmenting, and creating cold-water thermal refugia in rivers: Concepts derived from research on the Miramichi River, New Brunswick (Canada). Ecohydrology 2015, 8, 1095–1108. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site ID | Name | Rkm | Site Type | Depth (m) at Deployment |
|---|---|---|---|---|
| BC | BC | 54.1 | Main stem near dam | 15.8 |
| T1 | T1 | 56.7 | Main stem, trough | 13.8 |
| C1 | Cedar Cr. | 59.0 | Confluence | 0.9 |
| C2 | Big Muddy Cr. | 59.3 | Confluence | 5.0 |
| P1 | P1 | 62.7 | Main stem, peak | 11.2 |
| C3 | Renshaw Slough | 65.9 | Confluence | 1.6 |
| C4 | Tiger Slough | 70.6 | Confluence | 8.1 |
| T2 | T2 | 70.7 | Main stem, trough | 9.8 |
| P2 | P2 | 72.8 | Main stem, peak | 8.2 |
| T3 | T3 | 81.0 | Main stem, trough | 7.3 |
| C5 | Ruby Cr. | 81.6 | Confluence | 0.5 |
| C6 | Le Clerc Cr. | 88.3 | Confluence | 0.6 |
| C7 | Middle Cr. | 90.7 | Confluence | 0.6 |
| C8 | Mill Cr. | 91.9 | Confluence | 0.6 |
| P3 | P3 | 94.6 | Main stem, peak | 7.4 |
| C9 | Cusick Cr. | 97.3 | Confluence | 0.6 |
| P4 | P4 | 101.6 | Main stem, peak | 1.4 |
| C10 | Cee Cee Ah Cr. | 104.8 | Confluence | 1.2 |
| C11 | Tacoma Cr. | 104.9 | Confluence | 1.8 |
| T4 | T4 | 109.3 | Main stem, trough | 4.0 |
| P5 | P5 | 114.9 | Main stem, peak | 3.8 |
| C12 | Davis Cr | 115.6 | Confluence | 0.8 |
| C13 | Skookum Cr. | 116.1 | Confluence | 0.8 |
| T5 | T5 | 117.3 | Main stem, trough | 4.5 |
| P6 | P6 | 122.4 | Main stem, peak | 5.1 |
| C14 | Kent Cr. | 124.5 | Confluence | 0.7 |
| T6 | T6 | 124.9 | Main stem, trough | 6.1 |
| C15 | McCloud Cr. | 125.1 | Confluence | 0.4 |
| P7 | P7 | 127.2 | Main stem, peak | 4.9 |
| C16 | Indian Cr. | 129.1 | Confluence | 0.5 |
| C17 | Trout ponds | 131.0 | Confluence | 0.4 |
| T7 | T7 | 132.6 | Main stem, trough | 3.7 |
| T8 | T8 | 142.5 | Main stem, trough | 4.4 |
| AF | AF | 144.3 | Main stem near dam | 6.1 |
| Variable | Units/Category | Data Type | Description |
|---|---|---|---|
| Reach scale | |||
| Cross sectional area | z-scores | continuous | Measured with acoustic Doppler current profilers (ADCP) |
| Maximum water depth | z-scores | continuous | Measured with ADCP |
| Degree of confinement | m m−1 | continuous | Measured with remote sensing |
| Dense SAV | km2 | categories | Sum of all dense submerged aquatic vegetation patches regardless of plant species composition. |
| Subcatchment scale | |||
| Land use | % | continuous | Measured with remote sensing as percent developed land. |
| Aquifer permeability, | high/low | category | Locations where shallow subsurface groundwater meets deep groundwater flows |
| Size of tributaries | km2 | continuous | Watershed size |
| Distance from Albeni Falls dam | km | continuous | Distance from Albeni Falls dam to Box Canyon dam (lower boundary) |
| Site | Main Stem Water Temperature 7DADM (°C) | Cold-Water Refuge Persistence (%) | ||
|---|---|---|---|---|
| Bottom | Surface | 2 °C Criterion | 1 °C Criterion | |
| BC | 24.2 | 24.4 | 3 | 19 |
| T1 | 24.1 | 25.3 | 0 | 74 |
| P1 | 24.2 | 25.8 | 22 | 78 |
| T2 | 24.2 | 24.9 | 17 | 69 |
| P2 | 24.1 | 25.3 | 14 | 61 |
| T3 | 24.0 | 24.6 | 0 | 20 |
| P3 | 24.3 | 24.7 | 0 | 22 |
| P4 | 21.9 | 25.4 | 98 | 100 |
| T4 | 24.1 | 24.6 | 3 | 29 |
| P5 | 24.5 | 25.0 | 0 | 14 |
| T5 | 24.2 | 25.1 | 0 | 34 |
| P6 | 24.1 | 25.0 | 6 | 54 |
| T6 | 24.3 | 24.7 | 14 | 54 |
| P7 | 24.5 | 25.0 | 20 | 66 |
| T7 | 24.4 | 25.1 | 3 | 31 |
| T8 | 24.2 | 25.1 | 11 | 51 |
| AF | 24.2 | 24.9 | 9 | 57 |
| Site | Bottom Water Temperature 7DADM (°C) | Cold-Water Refuge Persistence (%) | Stratification (%) | |||
|---|---|---|---|---|---|---|
| Confluence | Main stem | Criterion = <18 °C | Criterion = 2 °C | Criterion = 1 °C | ||
| C1 | 18.8 | 24.0 | 53 | 100 | 100 | 97 |
| C2 | 24.0 | 24.0 | 0 | 0 | 0 | 1 |
| C3 | 14.7 | 24.0 | 100 | 100 | 100 | 100 |
| C4 | 12.4 | 24.0 | 100 | 100 | 100 | 100 |
| C5 | 20.1 | 24.0 | 17 | 100 | 100 | 95 |
| C6 | 18.1 | 24.0 | 97 | 100 | 100 | 36 |
| C7 | 23.0 | 24.0 | 10 | 86 | 93 | 43 |
| C8 | 22.8 | 24.0 | 0 | 35 | 55 | 25 |
| C9 | 20.8 | 21.7 | 0 | 37 | 96 | 70 |
| C10 | 18.5 | 23.8 | 62 | 100 | 100 | 39 |
| C11 | 18.8 | 23.8 | 45 | 100 | 100 | 32 |
| C12 | 21.7 | 24.0 | 0 | 100 | 100 | 33 |
| C13 | 18.7 | 24.0 | 55 | 100 | 100 | 0 |
| C14 | 24.5 | 24.0 | 31 | 52 | 66 | 77 |
| C15 | 23.7 | 24.3 | 0 | 31 | 52 | 36 |
| C16 | 18.1 | 24.3 | 97 | 100 | 100 | 14 |
| C17 | 15.6 | 24.3 | 100 | 100 | 100 | 87 |
| Model | K | −2LogL | AICc | ΔAICc | wi |
|---|---|---|---|---|---|
| Reach scale | |||||
| Median water temperature | |||||
| Confinement, Sum all dense vegetation, z-Area, z-Maximum depth | 14 | −12.98 | 60.0 | 0.00 | 0.52 |
| Confinement, z-Area, z- Maximum depth | 11 | −16.09 | 60.2 | 0.16 | 0.48 * |
| Probability of occurrence of cool-water areas | |||||
| Confinement, z-Area, z-Maximum depth | 6 | −25.57 | 64.2 | 0.00 | 0.30 |
| Confinement, Sum all dense vegetation, z-Area, z-Maximum depth | 6 | −25.57 | 64.2 | 0.00 | 0.30 |
| Confinement, z-Area | 4 | −27.26 | 65.0 | 0.84 | 0.20 * |
| Confinement, Sum all dense vegetation, z-Area | 4 | −27.26 | 65.0 | 0.84 | 0.20 |
| Subcatchment scale | |||||
| Median water temperature | |||||
| Distance from dam, Percent developed land | 10 | −5.25 | 34.3 | 0.00 | 0.28 |
| Distance from dam, Percent developed land, Tributary area | 10 | −5.21 | 34.3 | 0.01 | 0.27 |
| Distance from dam | 9 | −6.22 | 34.7 | 0.38 | 0.23 * |
| Distance from dam, Tributary area | 10 | −5.95 | 34.7 | 0.40 | 0.23 |
| Probability of occurrence of cool water | |||||
| Distance from dam, Percent developed land | 8 | −19.87 | 59.6 | 0.00 | 0.50 * |
| Distance from dam, Percent developed land, Tributary area | 8 | −19.87 | 59.6 | 0.00 | 0.50 |
| Model | edf | Ref.df | F or Chi-Square | p-Value | R-sq. (adj) | Deviance (%) |
|---|---|---|---|---|---|---|
| Reach scale | ||||||
| Median water temperature | ||||||
| s (Confinement) | 2.29 | 9 | 1.09 | 0.007 | 0.59 | 64 |
| s (z-Area) | 6.72 | 9 | 9.95 | <0.001 | ||
| s (z-Maximum depth) | 0.96 | 9 | 0.42 | 0.048 | ||
| Probability of occurrence of cool water | ||||||
| s (Confinement) | 2.92 | 9 | 8.74 | 0.02 | 0.51 | 50 |
| s (z_Area) | 0.99 | 9 | 16.63 | <0.001 | ||
| Subcatchment scale | ||||||
| Median water temperature | ||||||
| s (Distance from dam) | 7.78 | 9 | 21.07 | <0.001 | 0.68 | 71 |
| Probability of occurrence of cool water | ||||||
| s (Distance from dam) | 6.65 | 7 | 21.73 | <0.01 | 0.63 | 64 |
| s (Percent land developed) | 0.77 | 9 | 2.49 | 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejia, F.H.; Torgersen, C.E.; Berntsen, E.K.; Maroney, J.R.; Connor, J.M.; Fullerton, A.H.; Ebersole, J.L.; Lorang, M.S. Longitudinal, Lateral, Vertical, and Temporal Thermal Heterogeneity in a Large Impounded River: Implications for Cold-Water Refuges. Remote Sens. 2020, 12, 1386. https://doi.org/10.3390/rs12091386
Mejia FH, Torgersen CE, Berntsen EK, Maroney JR, Connor JM, Fullerton AH, Ebersole JL, Lorang MS. Longitudinal, Lateral, Vertical, and Temporal Thermal Heterogeneity in a Large Impounded River: Implications for Cold-Water Refuges. Remote Sensing. 2020; 12(9):1386. https://doi.org/10.3390/rs12091386
Chicago/Turabian StyleMejia, Francine H., Christian E. Torgersen, Eric K. Berntsen, Joseph R. Maroney, Jason M. Connor, Aimee H. Fullerton, Joseph L. Ebersole, and Mark S. Lorang. 2020. "Longitudinal, Lateral, Vertical, and Temporal Thermal Heterogeneity in a Large Impounded River: Implications for Cold-Water Refuges" Remote Sensing 12, no. 9: 1386. https://doi.org/10.3390/rs12091386
APA StyleMejia, F. H., Torgersen, C. E., Berntsen, E. K., Maroney, J. R., Connor, J. M., Fullerton, A. H., Ebersole, J. L., & Lorang, M. S. (2020). Longitudinal, Lateral, Vertical, and Temporal Thermal Heterogeneity in a Large Impounded River: Implications for Cold-Water Refuges. Remote Sensing, 12(9), 1386. https://doi.org/10.3390/rs12091386