Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain

 , , , , ,
, , , , ,  ,
,  and
and
Abstract
:
1. Introduction
2. Modeling Strategy
3. Study Area and Data
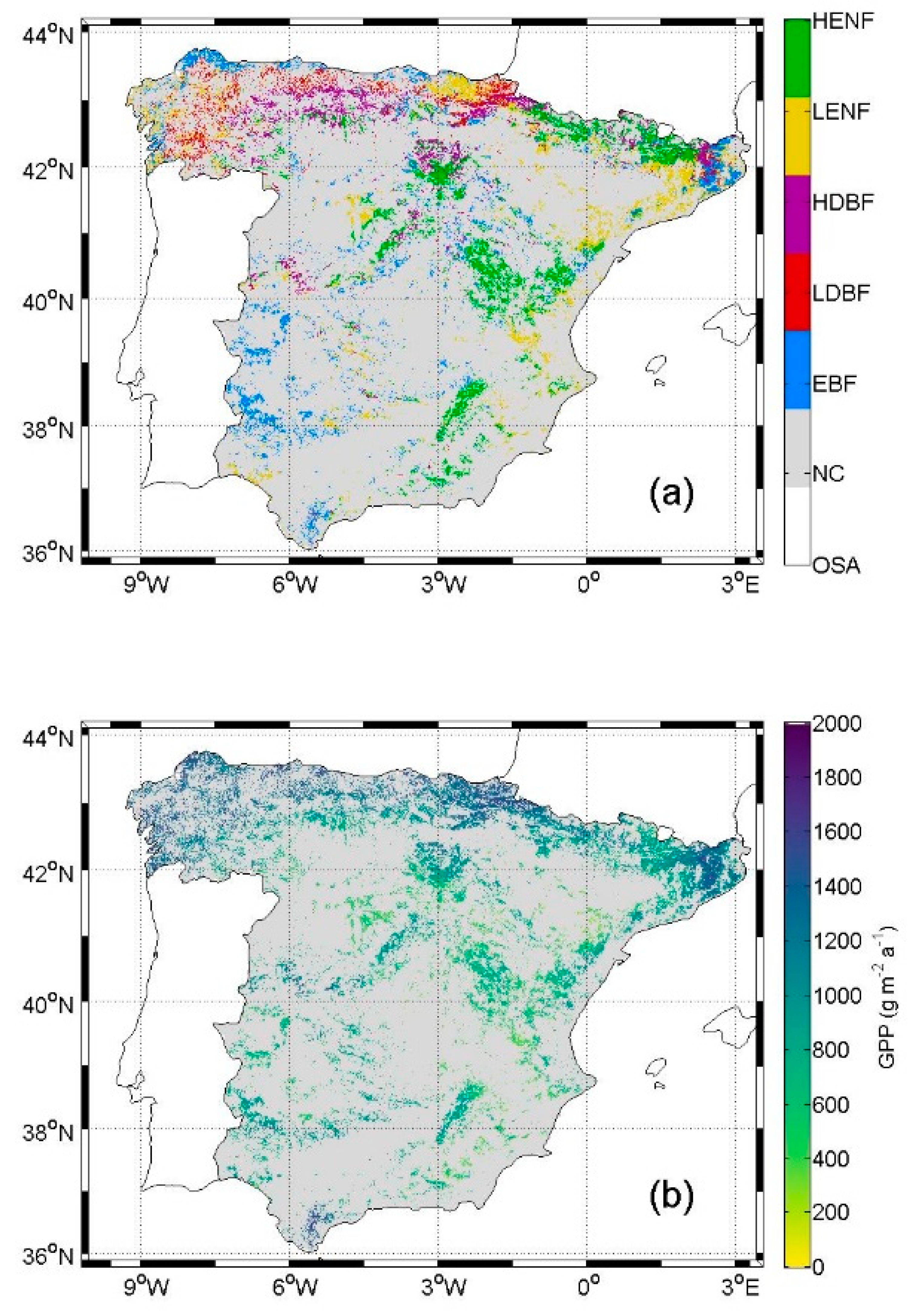
3.1. Peninsular Spain
3.2. Reference Forest Observations
3.3. Data Used for the NPP Modeling
4. Data Processing
4.1. Simulation of Forest NPP in Current Condition
4.2. Simulation of Forest NPP in Future Condition
5. Results
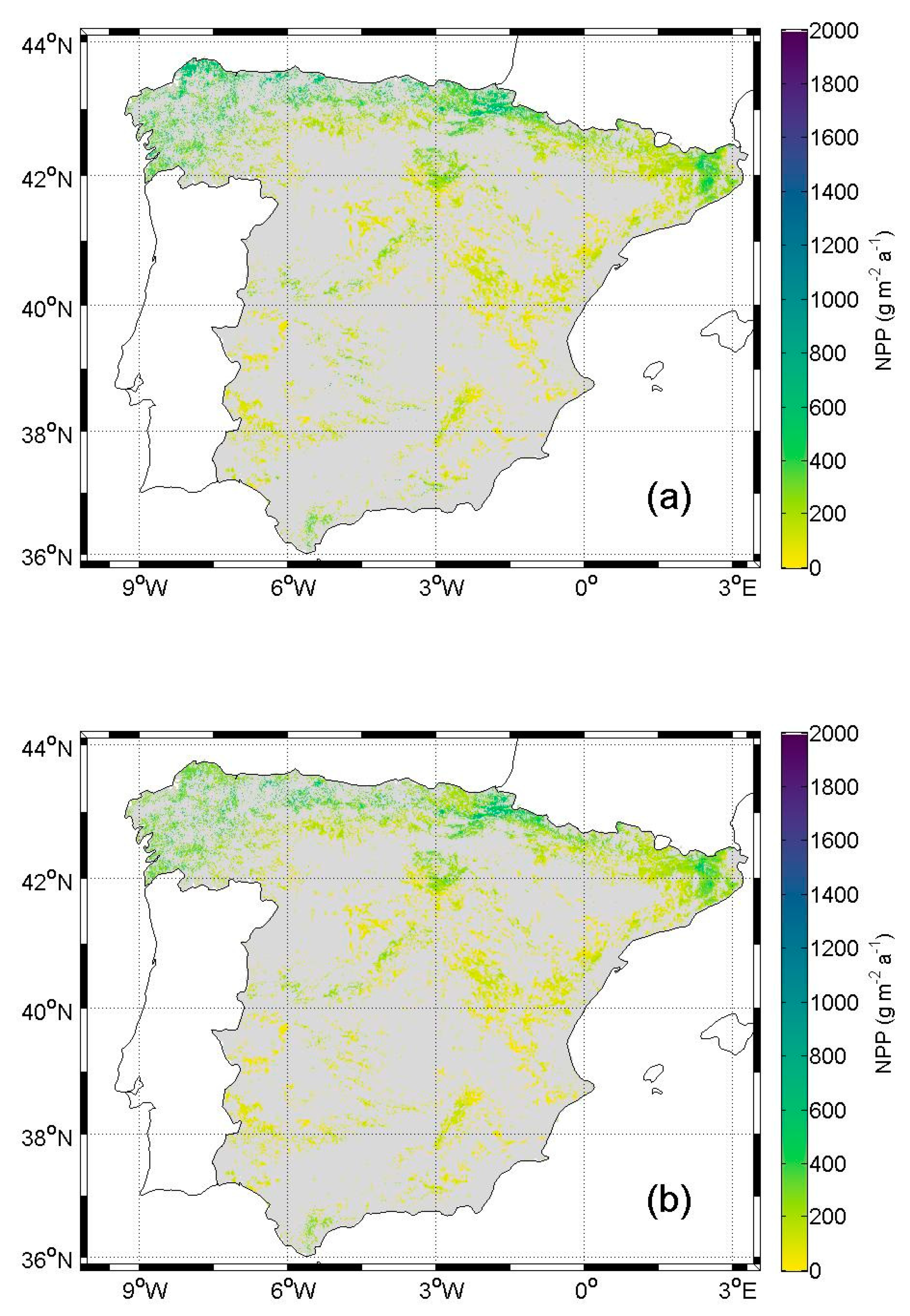
5.1. NPP in Current Condition
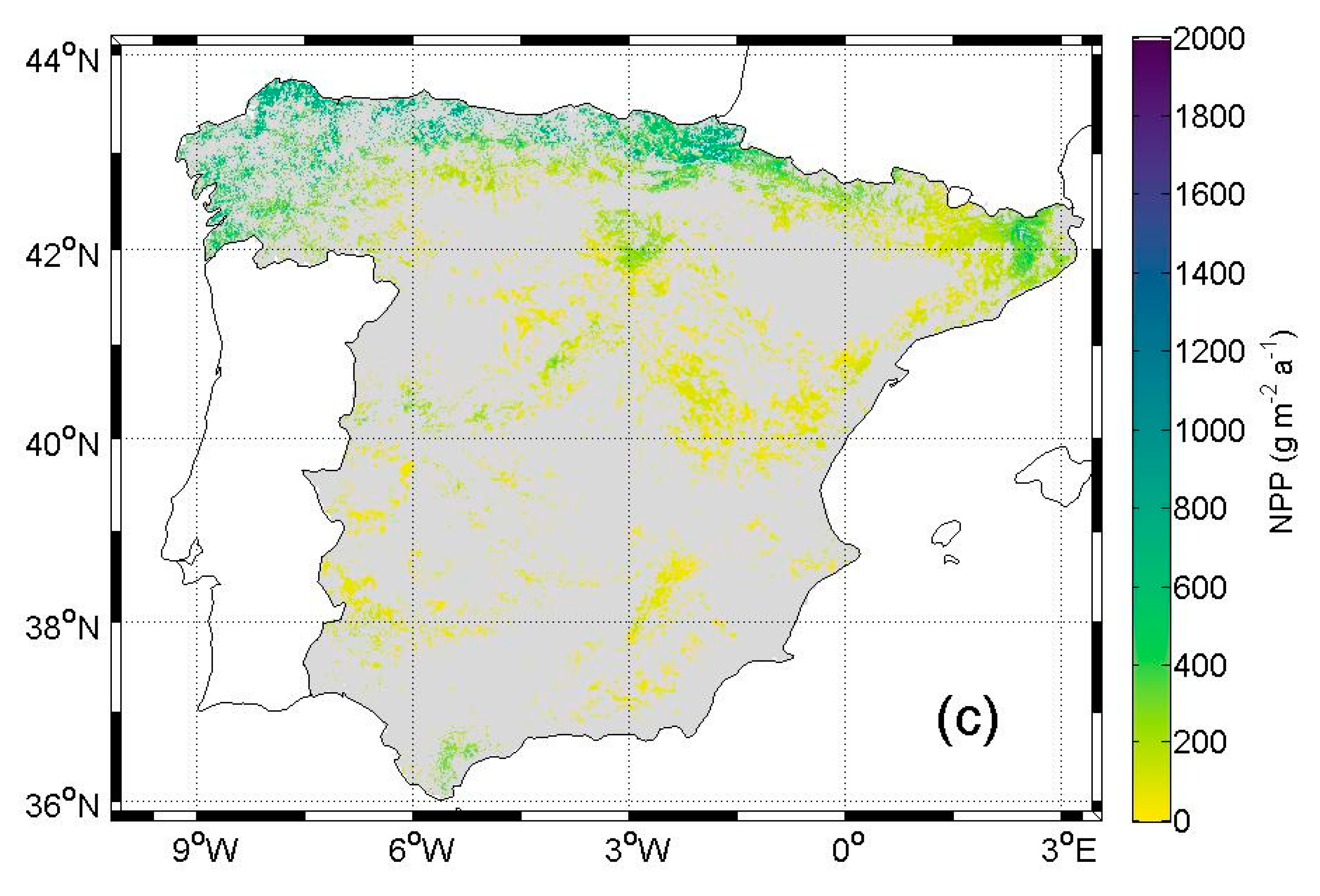
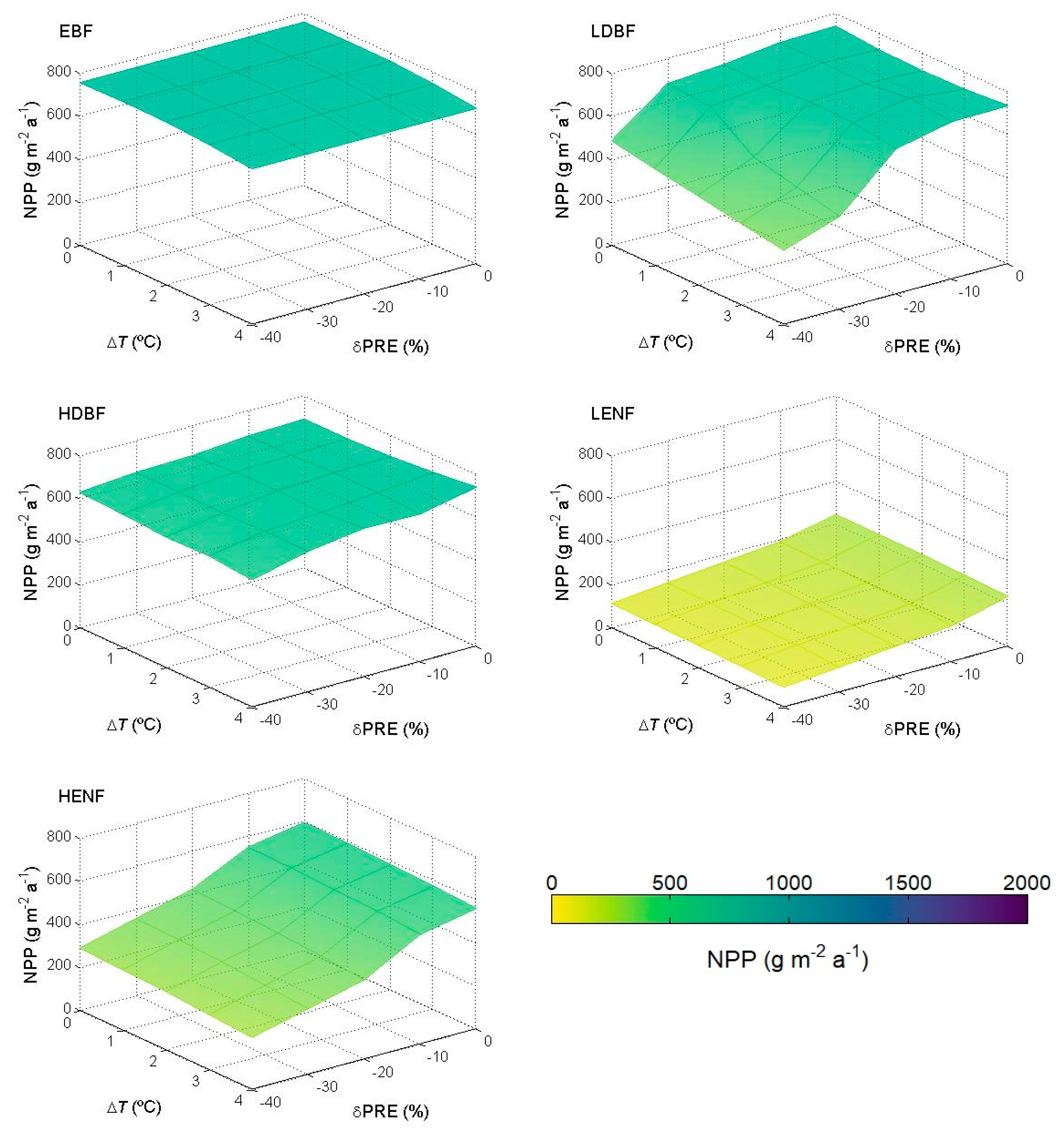
5.2. NPP in Future Condition
6. Discussion
6.1. NPP in Current Condition
6.2. NPP in Future Condition
7. Conclusions
- When applied in current climate conditions, the integrated modeling approach is capable of reproducing the general NPP variability of broadleaved forests in Spain (R2 > 0.60) but underestimates the NPP of needleleaf forests.
- When applied in future climate conditions, the approach produces results that can be explained considering the ecoclimatic features of the present forest types. NPP increases are predicted for more temperate-humid forests, while decreases are predicted for forest already subject to water limitation.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- McRoberts, R.E.; Tomppo, E.O. Remote sensing support for national forest inventories. Remote Sens. Environ. 2007, 110, 412–419. [Google Scholar] [CrossRef]
- Lu, D. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar] [CrossRef]
- Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente. DGCN II Inventario Forestal Nacional 1986–1996; Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente: Madrid, Spain, 1996.
- Xiao, J.; Chevallier, F.; Gomez, C.; Guanter, L.; Hicke, J.A.; Huete, A.R.; Ichii, K.; Ni, W.; Pang, Y.; Rahman, A.F.; et al. Remote sensing of the terrestrial carbon cycle: A review of advances over 50 years. Remote Sens. Environ. 2019, 233, 111383. [Google Scholar] [CrossRef]
- Kumar, L.; Mutanga, O. Remote sensing of above-ground biomass. Remote Sens. 2017, 9, 935. [Google Scholar] [CrossRef] [Green Version]
- Gilabert, M.A.; Moreno, A.; Maselli, F.; Martínez, B.; Chiesi, M.; Sánchez-Ruiz, S.; García-Haro, F.J.; Pérez-Hoyos, A.; Campos-Taberner, M.; Pérez-Priego, O.; et al. Daily GPP estimates in Mediterranean ecosystems by combining remote sensing and meteorological data. ISPRS J. Photogramm. Remote Sens. 2015, 102, 184–197. [Google Scholar] [CrossRef]
- Sánchez-Ruiz, S.; Moreno, A.; Piles, M.; Maselli, F.; Carrara, A.; Running, S.; Gilabert, M.A. Quantifying water stress effect on daily light use efficiency in Mediterranean ecosystems using satellite data. Int. J. Digit. Earth 2017, 10, 623–638. [Google Scholar] [CrossRef]
- Gilabert, M.A.; Sánchez-Ruiz, S.; Moreno, A. Annual gross primary production from vegetation indices: A theoretically sound approach. Remote Sens. 2017, 9, 193. [Google Scholar] [CrossRef] [Green Version]
- Monteith, J.L. Solar Radiation and Productivity in Tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.P.; Ritts, W.D.; Cohen, W.B.; Maeirsperger, T.K.; Gower, S.T.; Kirschbaum, A.A.; Running, S.W.; Zhao, M.; Wofsy, S.C.; Dunn, A.L.; et al. Site-level evaluation of satellite-based global terrestrial gross primary production and net primary production monitoring. Glob. Chang. Biol. 2005, 11, 666–684. [Google Scholar] [CrossRef]
- Turner, D.P.; Ritts, W.D.; Cohen, W.B.; Gower, S.T.; Running, S.W.; Zhao, M.; Costa, M.H.; Kirschbaum, A.A.; Ham, J.M.; Saleska, S.R.; et al. Evaluation of MODIS NPP and GPP products across multiple biomes. Remote Sens. Environ. 2006, 102, 282–292. [Google Scholar] [CrossRef]
- Townsend, P.A. Estimating forest structure in wetlands using multitemporal SAR. Remote Sens. Environ. 2002, 79, 288–304. [Google Scholar] [CrossRef]
- Running, S.; Hunt, E. Generalization of a forest ecosystem process model for other biomes, BIOME-BGC, and an application for global-scale models. In Scaling Physiological Processes: Leaf to Globe: A Volume in Physiological Ecology; Academic Press Inc.: Cambridge, MA, USA, 1993; pp. 141–158. [Google Scholar]
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and Sensitivity Analysis of the BIOME–BGC Terrestrial Ecosystem Model: Net Primary Production Controls. Earth Interact. 2000, 4, 1–85. [Google Scholar] [CrossRef]
- Chiesi, M.; Maselli, F.; Moriondo, M.; Fibbi, L.; Bindi, M.; Running, S.W. Application of BIOME-BGC to simulate Mediterranean forest processes. Ecol. Model. 2007, 206, 179–190. [Google Scholar] [CrossRef]
- Chiesi, M.; Cherubini, P.; Maselli, F. Adaptation of a modelling strategy to predict the NPP of even-aged forest stands. Eur. J. For. Res. 2012, 131, 1175–1184. [Google Scholar] [CrossRef]
- Sánchez-Ruiz, S.; Chiesi, M.; Fibbi, L.; Carrara, A.; Maselli, F.; Gilabert, M.A. Optimized Application of Biome-BGC for Modeling the Daily GPP of Natural Vegetation Over Peninsular Spain. J. Geophys. Res. Biogeosci. 2018, 123, 531–546. [Google Scholar]
- Maselli, F.; Chiesi, M.; Moriondo, M.; Fibbi, L.; Bindi, M.; Running, S.W. Modelling the forest carbon budget of a Mediterranean region through the integration of ground and satellite data. Ecol. Model. 2009, 220, 330–342. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2010. Terms and Definitions; FAO: Rome, Italy, 2010. [Google Scholar]
- Moreno, A.; Gilabert, M.A.; Martínez, B. Mapping daily global solar irradiation over Spain: A comparative study of selected approaches. Sol. Energy 2011, 85, 2072–2084. [Google Scholar] [CrossRef]
- Moreno, A.; Gilabert, M.A.; Camacho, F.; Martínez, B. Validation of daily global solar irradiation images from MSG over Spain. Renew. Energy 2013, 60, 332–342. [Google Scholar] [CrossRef]
- Thornton, P.E.; Hasenauers, H.; White, M.A. Simultaneous estimation of daily solar radiation and humidity from observed temperature and precipitation: An application over complex terrain in Austria. Agric. For. Meteorol. 2000, 104, 255–271. [Google Scholar] [CrossRef]
- Ballabio, C.; Panagos, P.; Monatanarella, L. Mapping topsoil physical properties at European scale using the LUCAS database. Geoderma 2016, 261, 110–123. [Google Scholar] [CrossRef]
- IGN. Documento Técnico SIOSE 2011; IGN: San Francisco, CA, USA, 2011. [Google Scholar]
- Farr, T.; Rosen, P.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geophys. 2007, 45. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Ruiz, S.; Moreno-Martínez, Á.; Izquierdo-Verdiguier, E.; Chiesi, M.; Maselli, F.; Gilabert, M.A. Growing stock volume from multi-temporal landsat imagery through google earth engine. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 101913. [Google Scholar] [CrossRef]
- Iqbal, M. An Introduction to Solar Radiation; Academic Press: Toronto, ON, Canada, 1983. [Google Scholar]
- Roujean, J.; Breon, F. Estimating PAR Absorbed by Vegetation from Bidirectional Reflectance Measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Maselli, F.; Papale, D.; Puletti, N.; Chirici, G.; Corona, P. Combining remote sensing and ancillary data to monitor the gross productivity of water-limited forest ecosystems. Remote Sens. Environ. 2009, 113, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.E.; Haise, H.R. Estimating evapotranspiration from solar radiation. J. Irrig. Drain. Div. 1965, 89, 15–41. [Google Scholar]
- Running, S.W.; Zhao, M. User’s Guide: Daily GPP and annual NPP (MOD17A2/A3) Products NASA Earth Observing System MODIS Land Algorithm. Version 3.0. 2015. Available online: https://www.ntsg.umt.edu/files/modis/MOD17UsersGuide2015_v3.pdf (accessed on 24 April 2020).
- Odum, E.P.; Barrett, G. Fundamentals of Ecology, 5th ed.; Thompson Brooks/Cole: Belmont, CA, USA, 2005; ISBN 0534420664. [Google Scholar]
- Alcaraz, D.; Paruelo, J.; Cabello, J. Identification of current ecosystem functional types in the Iberian Peninsula. Glob. Ecol. Biogeogr. 2006, 15, 200–212. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Pérez-Cabello, F.; Lasanta, T. Assessment of radiometric correction techniques in analyzing vegetation variability and change using time series of Landsat images. Remote Sens. Environ. 2008, 112, 3916–3934. [Google Scholar] [CrossRef]
- Valbuena-Carabaña, M.; de Heredia, U.L.; Fuentes-Utrilla, P.; González-Doncel, I.; Gil, L. Historical and recent changes in the Spanish forests: A socio-economic process. Rev. Palaeobot. Palynol. 2010, 162, 492–506. [Google Scholar] [CrossRef]
- Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente. DGCN III Inventario Forestal Nacional 1997–2007; Dirección General de Conservación de la Naturaleza, Ministerio de Medio Ambiente: Madrid, Spain, 2006.
- Valcarcel, N.; Villa, G.; Arozarena, A.; Garcia-Asensio, L.; Caballlero, M.E.E.; Porcuna, A.; Domenech, E.; Peces, J.J.J. SIOSE, a Successful Test Bench Towards Harmonization and Integration of Land Cover/Use Information As Environmental Reference Data. In Proceedings of the International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences, Beijing, China, 3–11 July 2008; Volume 37, pp. 1159–1164. [Google Scholar]
- LSA-SAF Product User Manual Down-Welling Surface Shortwave Flux (DSSF). 2011. Available online: http://landsaf.ipma.pt (accessed on 24 April 2020).
- Schaaf, C.B.; Gao, F.; Strahler, A.H.; Lucht, W.; Li, X.; Tsang, T.; Strugnell, N.C.; Zhang, X.; Jin, Y.; Muller, J.-P.; et al. Global albedo, BRDF and nadir BRDF-adjusted reflectance products from MODIS. Remote Sens. Environ. 2002, 83, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Moreno, Á.; García-Haro, F.; Martínez, B.; Gilabert, M.A. Noise Reduction and Gap Filling of fAPAR Time Series Using an Adapted Local Regression Filter. Remote Sens. 2014, 6, 8238–8260. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Runger, G. Gene selection with guided regularized random forest. Pattern Recognit. 2013, 46, 3483–3489. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Verdiguier, E.; Zurita-Milla, R.; Rolf, A. On the use of guided regularized random forests to identify crops in smallholder farm fields. In Proceedings of the 2017 9th International Workshop on the Analysis of Multitemporal Remote Sensing Images (MultiTemp), Bruges, Belgium, 27–29 June 2017; pp. 1–3. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Federici, S.; Vitullo, M.; Tulipano, S.; De Laurets, R.; Seufert, G. An approach to estimate carbon stocks change in forest carbon pools under the UNFCCC: The Italian case. iForest Biogeosci. For. 2008, 1, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Christensen, O.B.; Goodess, C.M.; Harris, I.; Watkiss, P. European and Global Climate Change Projections: Discussion of Climate Change Model Outputs, Scenarios and Uncertainty in the EC RTD ClimateCost Project. Clim. Proj. Final Rep. 2011, 1, 1–36. [Google Scholar]
- Chirici, G.; Chiesi, M.; Corona, P.; Puletti, N.; Mura, M.; Maselli, F. Prediction of forest NPP in Italy by the combination of ground and remote sensing data. Eur. J. For. Res. 2015, 134, 453–467. [Google Scholar] [CrossRef] [Green Version]
- Hasenauer, H.; Petritsch, R.; Zhao, M.; Boisvenue, C.; Running, S.W. Reconciling satellite with ground data to estimate forest productivity at national scales. For. Ecol. Manag. 2012, 276, 196–208. [Google Scholar] [CrossRef]
- Ryan, M.G. Effects of Climate Change on Plant Respiration Author (s): Michael G. Ryan Stable URL: http://www.jstor.org/stable/1941808. Eff. Clim. Chang. Plant Respir. 2015, 1, 157–167. [Google Scholar]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Species | Number of Plots | GSV (m3 ha−1) | CAI (m3 ha−1 a−1) |
|---|---|---|---|---|
| EBF | Quercus ilex | 266 | 45 | 1.06 |
| LDBF | Quercus robur | 182 | 122 | 4.10 |
| HDBF | Fagus sylvatica | 255 | 197 | 4.57 |
| LENF | Pinus pinea | 122 | 71 | 2.97 |
| HENF | Pinus nigra | 307 | 100 | 3.52 |
| Forest Type | h (m) | T (°C) | Rg (MJ m−2) | PRE (mm) | PET (mm) |
|---|---|---|---|---|---|
| EBF | 584 (335) | 14.53 (1.97) | 5725 (666) | 711 (300) | 1193 (235) |
| LDBF | 520 (186) | 12.73 (0.97) | 4863 (524) | 1084 (307) | 910 (139) |
| HDBF | 1099 (205) | 11.48 (1.19) | 5272 (420) | 850 (267) | 936 (118) |
| LENF | 484 (223) | 14.38 (1.68) | 5586 (635) | 714 (360) | 1153 (209) |
| HENF | 1198 (290) | 12.53 (1.54) | 5742 (348) | 604 (183) | 1089 (165) |
| Forest Type | ρ (Mg m−3) | C (m3 m−3) | D |
|---|---|---|---|
| EBF | 0.70 | 1.45 | 0.47 |
| LDBF | 0.69 | 1.33 | 0.45 |
| HDBF | 0.61 | 1.36 | 0.45 |
| LENF | 0.53 | 1.53 | 0.42 |
| HENF | 0.38 | 1.31 | 0.42 |
| Forest Type | R2 | MBE (g m−2 a−1) | RMSE (g m−2 a−1) | |||
|---|---|---|---|---|---|---|
| with GPPPEM | with GPPBGC | with GPPPEM | with GPPBGC | with GPPPEM | with GPPBGC | |
| EBF | 0.69 | 0.74 | 60 | 45 | 100 | 78 |
| LDBF | 0.60 | 0.56 | 64 | 20 | 150 | 150 |
| HDBF | 0.62 | 0.60 | 120 | 140 | 160 | 190 |
| LENF | 0.59 | 0.49 | −170 | −180 | 210 | 230 |
| HENF | 0.46 | 0.51 | −66 | −75 | 140 | 140 |
| Forest Type | GPPPEM | GPPBGC | ||||||
|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Std | Min | Max | Mean | Std | |
| EBF | 128 | 1778 | 916 | 308 | 13 | 1495 | 828 | 243 |
| LDBF | 310 | 1762 | 1229 | 202 | 201 | 1611 | 1156 | 220 |
| HDBF | 1 | 1785 | 1049 | 254 | 174 | 1729 | 1003 | 312 |
| LENF | 7 | 1876 | 864 | 311 | 0 | 1909 | 784 | 284 |
| HENF | 4 | 1592 | 774 | 220 | 205 | 1681 | 773 | 256 |
| Forest Type | NPP with GPPPEM | NPP with GPPBGC | ||||||
| Min | Max | Mean | Std | Min | Max | Mean | Std | |
| EBF | 8 | 876 | 181 | 168 | 12 | 470 | 143 | 110 |
| LDBF | 36 | 832 | 418 | 138 | 17 | 788 | 385 | 134 |
| HDBF | 4 | 661 | 232 | 128 | 14 | 679 | 218 | 134 |
| LENF | 1 | 197 | 149 | 116 | 7 | 495 | 123 | 87 |
| HENF | 1 | 549 | 125 | 84 | 9 | 583 | 123 | 87 |
| Forest Type | Min | Max | Mean | Std |
|---|---|---|---|---|
| EBF | 14 | 875 | 197 | 218 |
| LDBF | 17 | 1136 | 564 | 204 |
| HDBF | 15 | 696 | 220 | 133 |
| LENF | 7 | 772 | 215 | 179 |
| HENF | 8 | 622 | 104 | 84 |
| Forest Type | LAT (°) | LON (°) | h (m) | T (°C) | Rg (MJ m−2) | PRE (mm) | PET (mm) |
|---|---|---|---|---|---|---|---|
| EBF | 43.6161 | −7.9821 | 225 | 13.75 | 4514 | 1366 | 856 |
| LDBF | 42.6964 | −2.4732 | 679 | 12.43 | 4855 | 743 | 921 |
| HDBF | 42.7946 | −2.0714 | 1135 | 12.47 | 5098 | 1018 | 962 |
| LENF | 40.3303 | −6.0982 | 574 | 13.91 | 5984 | 811 | 1220 |
| HENF | 41.8304 | −2.9821 | 1160 | 10.37 | 5513 | 676 | 923 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Ruiz, S.; Maselli, F.; Chiesi, M.; Fibbi, L.; Martínez, B.; Campos-Taberner, M.; García-Haro, F.J.; Gilabert, M.A. Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain. Remote Sens. 2020, 12, 1356. https://doi.org/10.3390/rs12091356
Sánchez-Ruiz S, Maselli F, Chiesi M, Fibbi L, Martínez B, Campos-Taberner M, García-Haro FJ, Gilabert MA. Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain. Remote Sensing. 2020; 12(9):1356. https://doi.org/10.3390/rs12091356
Chicago/Turabian StyleSánchez-Ruiz, Sergio, Fabio Maselli, Marta Chiesi, Luca Fibbi, Beatriz Martínez, Manuel Campos-Taberner, Francisco Javier García-Haro, and María Amparo Gilabert. 2020. "Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain" Remote Sensing 12, no. 9: 1356. https://doi.org/10.3390/rs12091356
APA StyleSánchez-Ruiz, S., Maselli, F., Chiesi, M., Fibbi, L., Martínez, B., Campos-Taberner, M., García-Haro, F. J., & Gilabert, M. A. (2020). Remote Sensing and Bio-Geochemical Modeling of Forest Carbon Storage in Spain. Remote Sensing, 12(9), 1356. https://doi.org/10.3390/rs12091356