Synthesizing Remote Sensing and Biophysical Measures to Evaluate Human–wildlife Conflicts: The Case of Wild Boar Crop Raiding in Rural China



Abstract
1. Introduction
2. Materials and Methods
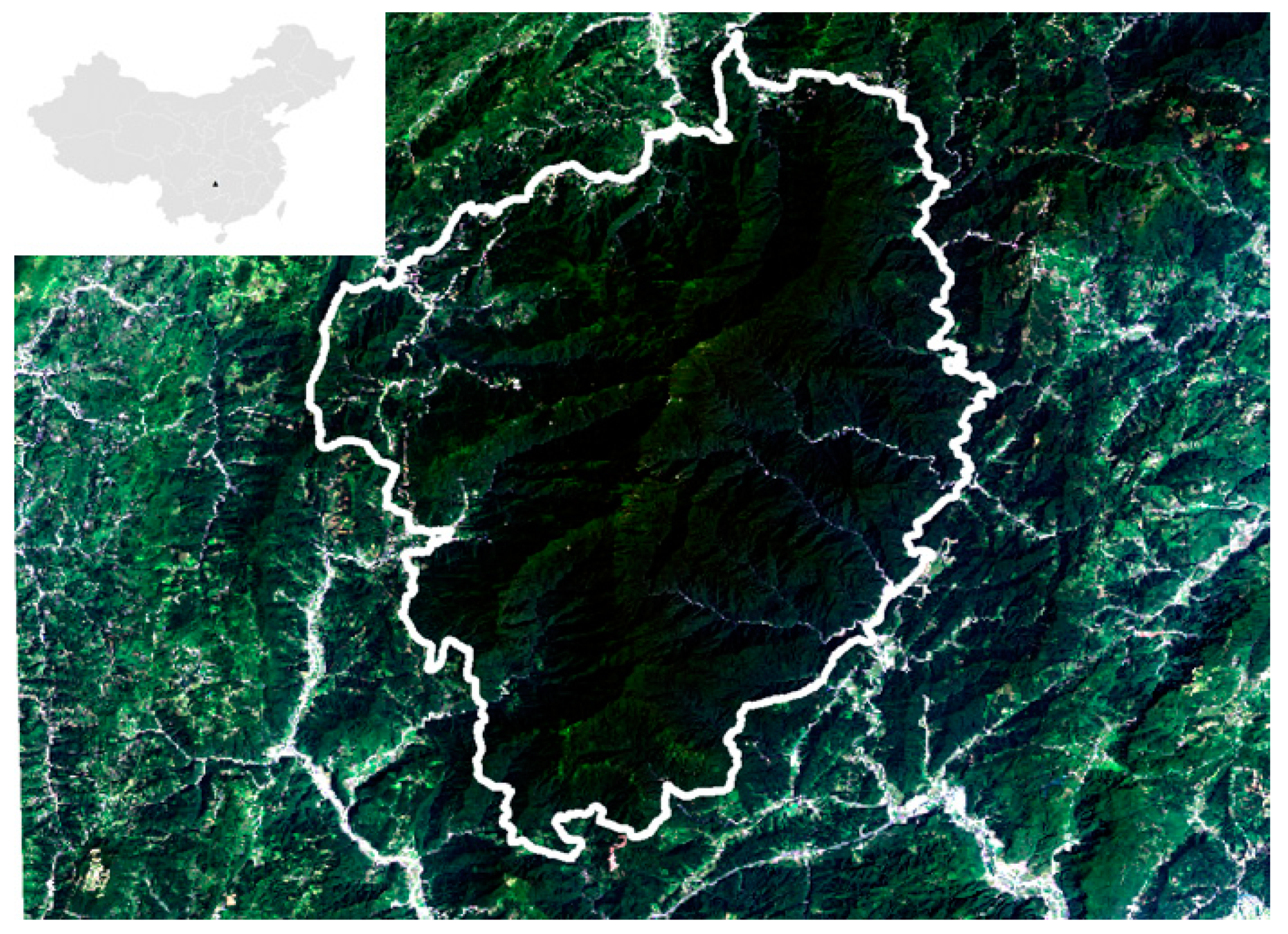
2.1. Study Site
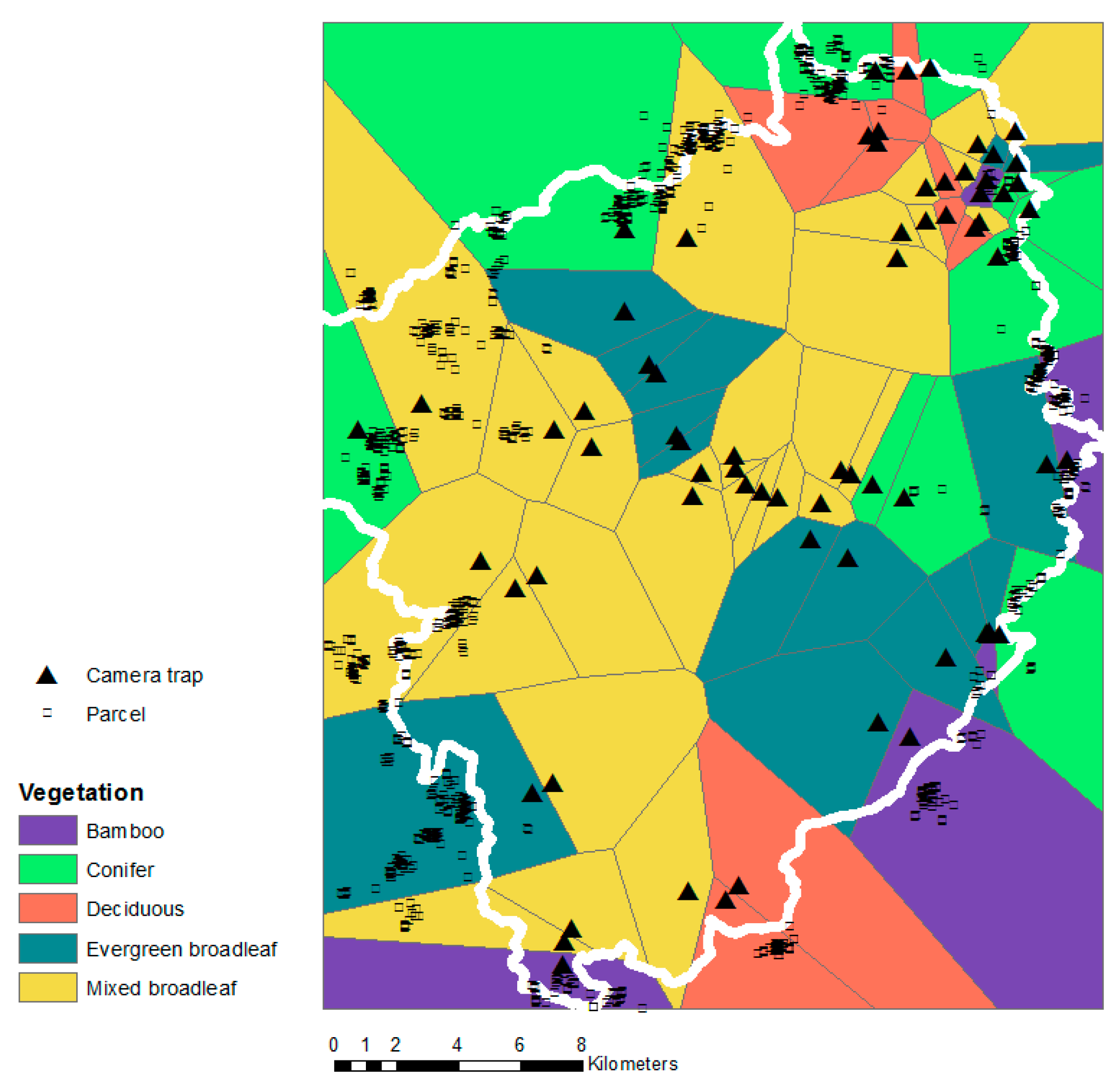
2.2. Empirical Data
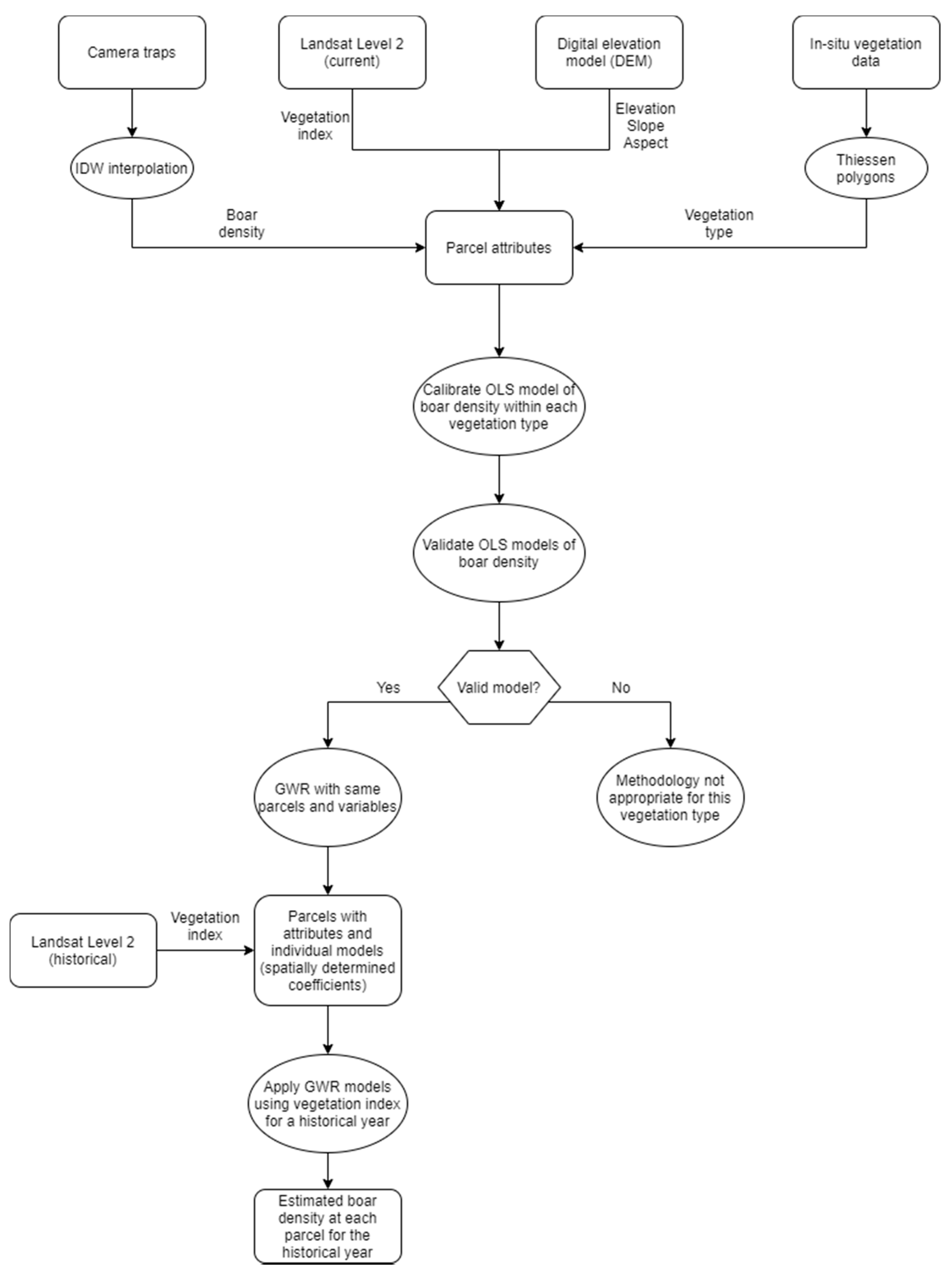
2.3. Data Analyses
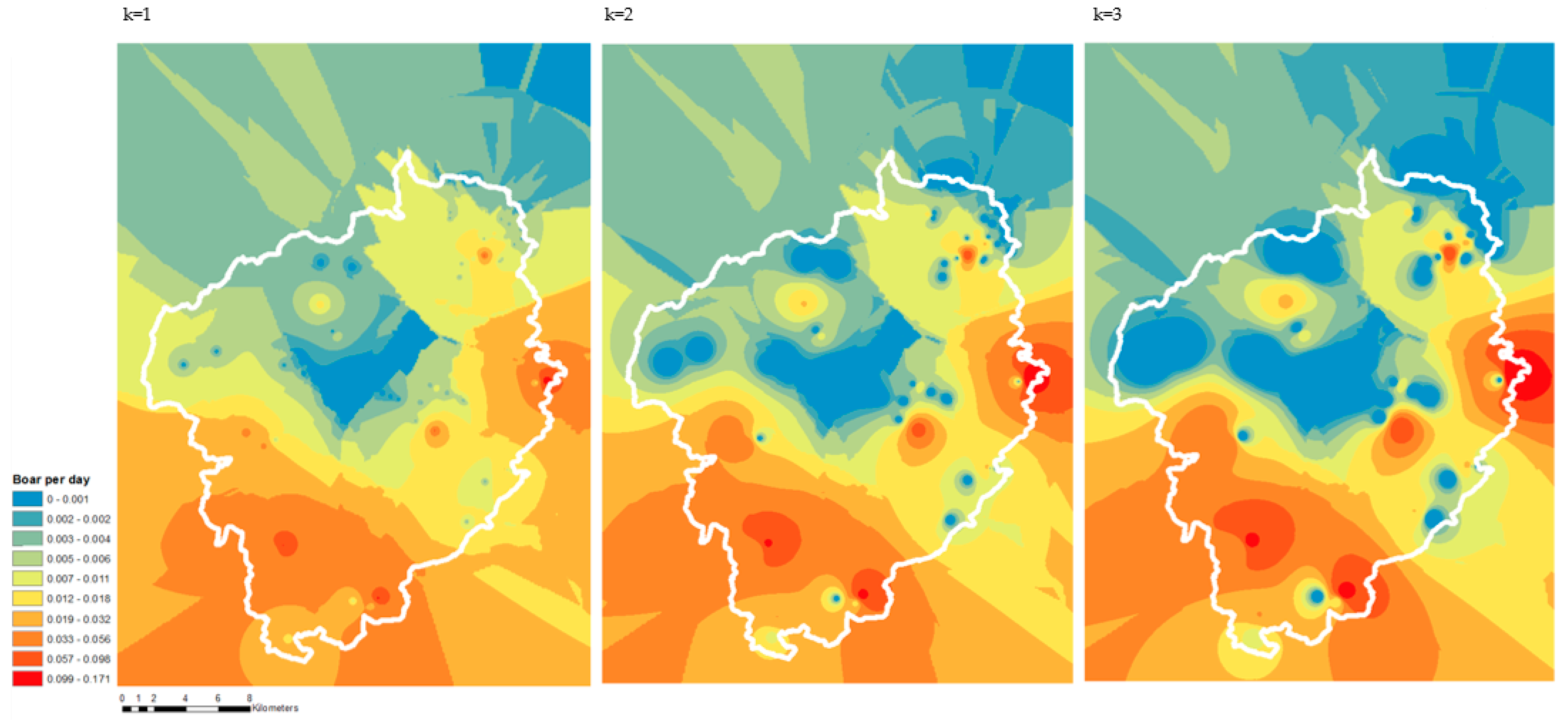
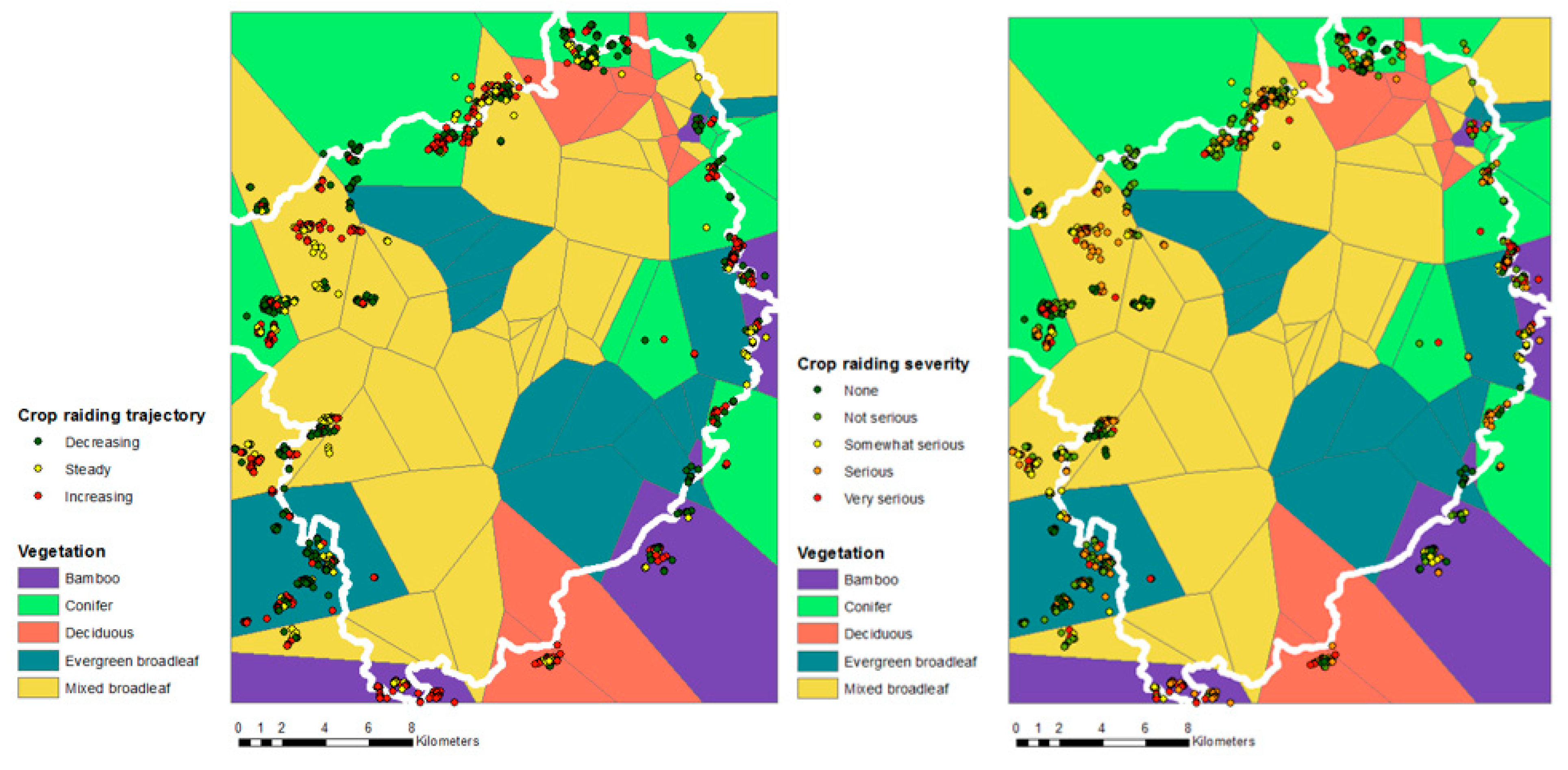
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band | Year | Equation | R2 | n |
|---|---|---|---|---|
| NIR | 1996 | 0.85 | 258 | |
| 2002 | 0.85 | 258 | ||
| 2011 | 0.86 | 249 | ||
| 2016 | 0.85 | 234 | ||
| Red | 1996 | 0.91 | 235 | |
| 2002 | 0.86 | 248 | ||
| 2011 | 0.85 | 243 | ||
| 2016 | 0.85 | 240 |
Appendix B
| Plot ID | Start Date | End Date | Long. (WGS84) | Lat. (WGS84) | Interruption Start | Interruption End |
|---|---|---|---|---|---|---|
| 2 | 17 April 2015 | 16 March 2016 | 108.761 | 27.85246 | ||
| 5 | 22 April 2015 | 16 March 2016 | 108.7325 | 27.88133 | 7 September 2015 | 23 October 2015 |
| 7 | 26 April 2015 | 16 March 2016 | 108.7217 | 27.8868 | ||
| 19 | 28 April 2015 | 15 March 2016 | 108.6994 | 27.91098 | 23 June 2015 | 6 July 2015 |
| 20 | 28 April 2015 | 15 March 2016 | 108.7029 | 27.90261 | 30 June 2015 | 23 October 2015 |
| 21 | 28 April 2015 | 15 March 2016 | 108.6998 | 27.90731 | 7 September 2015 | 23 October 2015 |
| 22 | 29 April 2015 | 15 March 2016 | 108.7075 | 27.90061 | ||
| 23 | 29 April 2015 | 15 March 2016 | 108.712 | 27.89908 | 7 September 2015 | 23 October 2015 |
| 24 | 29 April 2015 | 15 March 2016 | 108.7245 | 27.89716 | ||
| 27 | 2 May 2015 | 18 March 2016 | 108.7736 | 27.85997 | ||
| 28 | 2 May 2015 | 18 March 2016 | 108.7725 | 27.85966 | ||
| 29 | 4 May 2015 | 18 March 2016 | 108.7331 | 27.90562 | 1 September 2015 | 8 September 2015 |
| 30 | 4 May 2015 | 18 March 2016 | 108.7302 | 27.90692 | ||
| 31 | 14 May 2015 | 15 March 2016 | 108.697 | 27.78216 | ||
| 32 | 14 May 2015 | 15 March 2016 | 108.7005 | 27.78644 | 26 June 2015 | 30 October 2015 |
| 34 | 19 May 2015 | 8 April 2016 | 108.641 | 27.81311 | ||
| 35 | 20 May 2015 | 7 August 2016 | 108.6495 | 27.76345 | ||
| 36 | 20 May 2015 | 7 August 2016 | 108.6499 | 27.77022 | ||
| 37 | 20 May 2015 | 9 April 2016 | 108.6522 | 27.77409 | 14 November 2015 | 20 November 2015 |
| 38 | 21 May 2015 | 2 August 2016 | 108.6257 | 27.88032 | ||
| 39 | 22 May 2015 | 5 August 2016 | 108.6357 | 27.87258 | 16 September 2015 | 9 November 2015 |
| 40 | 22 May 2015 | 5 August 2016 | 108.6422 | 27.87614 | ||
| 41 | 28 May 2015 | 30 June 2016 | 108.6579 | 27.91372 | ||
| 42 | 9 November 2015 | 2 August 2016 | 108.6471 | 27.91849 | ||
| 43 | 31 May 2015 | 1 July 2016 | 108.6558 | 27.92395 | ||
| 44 | 2 June 2015 | 9 April 2016 | 108.7692 | 27.97725 | ||
| 45 | 2 June 2015 | 28 April 2016 | 108.755 | 27.97899 | ||
| 46 | 2 June 2015 | 19 April 2016 | 108.7478 | 27.97599 | 1 July 2015 | 27 October 2015 |
| 47 | 4 June 2015 | 15 August 2016 | 108.7607 | 27.98081 | ||
| 48 | 5 June 2015 | 15 August 2016 | 108.7549 | 27.98858 | ||
| 49 | 13 June 2015 | 30 July 2016 | 108.7384 | 28.004 | ||
| 50 | 13 June 2015 | 30 July 2016 | 108.7409 | 28.00193 | ||
| 54 | 23 June 2015 | 28 July 2016 | 108.6826 | 27.91677 | ||
| 55 | 23 June 2015 | 28 July 2016 | 108.6838 | 27.91513 | 28 October 2015 | 4 November 2015 |
| 57 | 24 June 2015 | 16 November 2015 | 108.6862 | 27.78472 | 3 July 2015 | 13 November 2015 |
| 58 | 10 July 2015 | 8 August 2016 | 108.6769 | 27.93495 | 24 November 2015 | 17 February 2016 |
| 59 | 10 July 2015 | 14 September 2015 | 108.6747 | 27.93743 | ||
| 60 | 10 July 2015 | 1 April 2016 | 108.6676 | 27.95285 | ||
| 10 | 25 October 2015 | 30 July 2016 | 108.7412 | 28.00497 | ||
| 8 | 4 December 2015 | 13 August 2016 | 108.7705 | 27.97893 | ||
| 9 | 2 November 2015 | 27 July 2016 | 108.7394 | 27.90263 | ||
| 11 | 4 November 2015 | 27 July 2016 | 108.7733 | 27.85921 | ||
| 12 | 19 March 2016 | 27 July 2016 | 108.79 | 27.90854 | ||
| 13 | 13 November 2015 | 27 July 2016 | 108.7959 | 27.90966 | ||
| 14 | 8 April 2016 | 5 August 2016 | 108.6466 | 27.81613 | ||
| 77 | 18 March 2016 | 12 July 2016 | 108.7488 | 27.89871 | ||
| 76 | 18 March 2016 | 27 July 2016 | 108.7764 | 27.85929 | ||
| 75 | 19 March 2016 | 10 August 2016 | 108.7725 | 27.99097 | ||
| 15 | 19 March 2016 | 18 July 2016 | 108.7708 | 27.98718 | ||
| 74 | 22 March 2016 | 24 July 2016 | 108.7411 | 27.83366 | ||
| 73 | 22 March 2016 | 12 May 2016 | 108.7505 | 27.82971 | ||
| 72 | 23 March 2016 | 10 August 2016 | 108.7774 | 27.98696 | ||
| 71 | 23 March 2016 | 12 August 2016 | 108.7817 | 27.99012 | ||
| 70 | 25 March 2016 | 14 August 2016 | 108.7466 | 27.9684 | ||
| 68 | 13 April 2016 | 15 August 2016 | 108.7664 | 27.99333 | ||
| 67 | 27 March 2016 | 20 July 2016 | 108.781 | 28.00514 | ||
| 66 | 27 March 2016 | 11 August 2016 | 108.7815 | 27.99568 | ||
| 65 | 28 March 2016 | 11 August 2016 | 108.7746 | 27.99835 | ||
| 64 | 28 March 2016 | 24 June 2016 | 108.77 | 28.0013 | ||
| 63 | 29 March 2016 | 12 August 2016 | 108.7759 | 27.96883 | ||
| 62 | 29 March 2016 | 11 August 2016 | 108.7852 | 27.98265 | ||
| 61 | 30 March 2016 | 13 July 2016 | 108.7564 | 28.02354 | ||
| 53 | 2 April 2016 | 23 April 2016 | 108.6678 | 27.97648 | ||
| 51 | 4 April 2016 | 10 July 2016 | 108.7497 | 28.02282 | ||
| 18 | 4 April 2016 | 30 July 2016 | 108.7404 | 28.02277 | ||
| 16 | 6 April 2016 | 31 July 2016 | 108.5902 | 27.91872 | ||
| 17 | 6 April 2016 | 25 June 2016 | 108.6088 | 27.92623 | ||
| 78 | 10 April 2016 | 28 July 2016 | 108.69 | 27.90617 | ||
| 79 | 10 April 2016 | 28 July 2016 | 108.6875 | 27.89934 |
References
- Dickman, A.J. Complexities of conflict: The importance of considering social factors for effectively resolving human–wildlife conflict. Anim. Conserv. 2010, 13, 458–466. [Google Scholar] [CrossRef]
- Distefano, E. Human-Wildlife Conflict Worldwide: Collection of Case Studies, Analysis of Management Strategies and Good Practices; Sustainable Agriculture and Rural Development Initiative (SARDI); Food and Agricultural Organization of the United Nations (FAO): Rome, Italy, 2005. [Google Scholar]
- Linkie, M.; Dinata, Y.; Nofrianto, A.; Leader-Williams, N. Patterns and perceptions of wildlife crop raiding in and around Kerinci Seblat National Park, Sumatra. Anim. Conserv. 2007, 10, 127–135. [Google Scholar] [CrossRef]
- Mojo, D.; Rothschuh, J.; Alebachew, M. Farmers’ perceptions of the impacts of human–wildlife conflict on their livelihood and natural resource management efforts in Cheha Woreda of Guraghe Zone, Ethiopia. Hum. Wildl. Interact. 2014, 8, 7. [Google Scholar]
- Nyhus, P.J.; Tilson, R. Crop-raiding elephants and conservation implications at Way Kambas National Park, Sumatra, Indonesia. Oryx 2000, 34, 262–274. [Google Scholar] [CrossRef]
- Wang, P.; Wolf, S.A.; Lassoie, J.P.; Poe, G.L.; Morreale, S.J.; Su, X.; Dong, S. Promise and reality of market-based environmental policy in China: Empirical analyses of the ecological restoration program on the Qinghai-Tibetan Plateau. Glob. Environ. Chang. 2010, 39, 35–44. [Google Scholar] [CrossRef]
- Cai, J.; Jiang, Z.; Zeng, Y.; Li, C.; Bravery, B.D. Factors affecting crop damage by wild boar and methods of mitigation in a giant panda reserve. Eur. J. Wildl. Res. 2008, 54, 723–728. [Google Scholar] [CrossRef]
- Ango, T.G.; Börjeson, L.; Senbeta, F.; Hylander, K. Balancing ecosystem services and disservices: Smallholder farmers’ use and management of forest and trees in an agricultural landscape in southwestern Ethiopia. Ecol. Soc. 2014, 19. [Google Scholar] [CrossRef]
- McDermid, G.J.; Hall, R.J.; Sanchez-Azofeifa, G.A.; Franklin, S.E.; Stenhouse, G.B.; Kobliuk, T.; LeDrew, E.F. Remote sensing and forest inventory for wildlife habitat assessment. For. Ecol. Manag. 2009, 257, 2262–2269. [Google Scholar] [CrossRef]
- Szantoi, Z.; Smith, S.E.; Strona, G.; Koh, L.P.; Wich, S.A. Mapping orangutan habitat and agricultural areas using Landsat OLI imagery augmented with unmanned aircraft system aerial photography. Int. J. Remote Sens. 2017, 38, 2231–2245. [Google Scholar] [CrossRef]
- Viña, A.; Bearer, S.; Zhang, H.; Ouyang, Z.; Liu, J. Evaluating MODIS data for mapping wildlife habitat distribution. Remote Sens. Environ. 2008, 112, 2160–2169. [Google Scholar] [CrossRef]
- Ackers, S.H.; Davis, R.J.; Olsen, K.A.; Dugger, K.M. The evolution of mapping habitat for northern spotted owls (Strix occidentalis caurina): A comparison of photo-interpreted, Landsat-based, and lidar-based habitat maps. Remote Sens. Environ. 2015, 156, 361–373. [Google Scholar] [CrossRef]
- De Leeuw, J.; Ottichilo, W.K.; Toxopeus, A.G.; Prins, H.H. Application of remote sensing and geographic information systems in wildlife mapping and modelling. Environ. Model. GIS Remote Sens. 2002, 4, 121–131. [Google Scholar]
- Simons-Legaard, E.M.; Harrison, D.J.; Legaard, K.R. Habitat monitoring and projections for Canada lynx: Linking the Landsat archive with carnivore occurrence and prey density. J. Appl. Ecol. 2016, 53, 1260–1269. [Google Scholar] [CrossRef]
- Porwal, M.C.; Roy, P.S.; Chellamuthu, V. Wildlife habitat analysis for ‘sambar’ (Cervus unicolor) in Kanha National Park using remote sensing. Int. J. Remote Sens. 1996, 17, 2683–2697. [Google Scholar] [CrossRef]
- Schairer, G.L.; Wynne, R.H.; Fies, M.L.; Klopfer, S.D. Predicting landscape quality for northern bobwhite from classified Landsat imagery. In Proceedings of the Southeastern Association of Fish and Wildlife Agencies; SEAFWA: Flora, MS, USA, 1999; Volume 53, pp. 243–256. [Google Scholar]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Cove, M.V.; Spínola, R.M.; Jackson, V.L.; Sáenz, J.C.; Chassot, O. Integrating occupancy modeling and camera-trap data to estimate medium and large mammal detection and richness in a Central American biological corridor. Trop. Conserv. Sci. 2013, 6, 781–795. [Google Scholar] [CrossRef]
- Massei, G.; Coats, J.; Lambert, M.S.; Pietravalle, S.; Gill, R.; Cowan, D. Camera traps and activity signs to estimate wild boar density and derive abundance indices. Pest Manag. Sci. 2018, 74, 853–860. [Google Scholar] [CrossRef]
- Bechtel, R.; Sanchez-Azofeifa, A.; Rivard, B.; Hamilton, G.; Martin, J.; Dzus, E. Associations between Woodland Caribou telemetry data and Landsat TM spectral reflectance. Int. J. Remote Sens. 2004, 25, 4813–4828. [Google Scholar] [CrossRef]
- Sekhar, N.U. Crop and livestock depredation caused by wild animals in protected areas: The case of Sariska Tiger Reserve, Rajasthan, India. Environ. Conserv. 1998, 25, 160–171. [Google Scholar] [CrossRef]
- Hua, X.; Yan, J.; Li, H.; He, W.; Li, X. Wildlife damage and cultivated land abandonment: Findings from the mountainous areas of Chongqing, China. Crop Prot. 2016, 84, 141–149. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Q.; Peterson, M.N.; Song, C. Feedback effect of crop raiding in payments for ecosystem services. Ambio 2019, 48, 732–740. [Google Scholar] [CrossRef]
- Thurfjell, H.; Ball, J.P.; Åhlén, P.A.; Kornacher, P.; Dettki, H.; Sjöberg, K. Habitat use and spatial patterns of wild boar Sus scrofa (L.): Agricultural fields and edges. Eur. J. Wildl. Res. 2009, 55, 517–523. [Google Scholar] [CrossRef]
- Park, C.R.; Lee, W.S. Development of a GIS-based habitat suitability model for wild boar Sus scrofa in the Mt. Baekwoonsan region, Korea. Mammal Study 2003, 28, 17–21. [Google Scholar] [CrossRef]
- Liu, X.; Wu, P.; Shao, X.; Songer, M.; Cai, Q.; He, X.; Zhu, Y. Diversity and activity patterns of sympatric animals among four types of forest habitat in Guanyinshan Nature Reserve in the Qinling Mountains, China. Environ. Sci. Pollut. Res. 2017, 24, 16465–16477. [Google Scholar] [CrossRef]
- Keuling, O.; Stier, N.; Roth, M. Commuting, shifting or remaining? Different spatial utilisation patterns of wild boar Sus scrofa L. in forest and field crops during summer. Mamm. Biol. 2009, 74, 145–152. [Google Scholar] [CrossRef]
- Anderson, S.J.; Stone, C.P. Snaring to control feral pigs Sus scrofa in a remote Hawaiian rain forest. Biol. Conserv. 1993, 63, 195–201. [Google Scholar] [CrossRef]
- Parker, A.J. The topographic relative moisture index: An approach to soil-moisture assessment in mountain terrain. Phys. Geogr. 1982, 3, 160–168. [Google Scholar] [CrossRef]
- Rho, P. Using habitat suitability model for the wild boar (Sus scrofa Linnaeus) to select wildlife passage sites in extensively disturbed temperate forests. J. Ecol. Environ. 2015, 38, 163–173. [Google Scholar] [CrossRef]
- Thapa, S. Effectiveness of crop protection methods against wildlife damage: A case study of two villages at Bardia National Park, Nepal. Crop Prot. 2010, 29, 1297–1304. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Aitken, S.C.; An, L. Figured worlds: Environmental complexity and affective ecologies in Fanjingshan, China. Ecol. Model. 2012, 229, 5–15. [Google Scholar] [CrossRef]
- Wandersee, S.M.; An, L.; López-Carr, D.; Yang, Y. Perception and decisions in modeling coupled human and natural systems: A case study from Fanjingshan National Nature Reserve, China. Ecol. Model. 2012, 229, 37–49. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Ouyang, Z.; Tam, C.; Chen, X. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477–9482. [Google Scholar] [CrossRef] [PubMed]
- Olmanson, L.G.; Bauer, M.E.; Brezonik, P.L. A 20-year Landsat water clarity census of Minnesota’s 10,000 lakes. Remote Sens. Environ. 2008, 112, 4086–4097. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S.A. Comparison of time series tasseled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. of Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]
- Smith, G.M.; Milton, E.J. The use of the empirical line method to calibrate remotely sensed data to reflectance. Int. J. Remote Sens. 1999, 20, 2653–2662. [Google Scholar] [CrossRef]
- Gitelson, A.A. Wide dynamic range vegetation index for remote quantification of biophysical characteristics of vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Lewison, R.L.; An, L.; Tsai, Y.H.; Stow, D.; Shi, L.; Yang, S. Assessing the effects of payments for ecosystem services programs on forest structure and species biodiversity. Biodivers. Conserv. 2020, in press. [Google Scholar]
- Coppin, P.R.; Bauer, M.E. Digital change detection in forest ecosystems with remote sensing imagery. Remote Sens. Rev. 1996, 13, 207–234. [Google Scholar] [CrossRef]
- Lu, G.Y.; Wong, D.W. An adaptive inverse-distance weighting spatial interpolation technique. Comput. Geosci. 2008, 34, 1044–1055. [Google Scholar] [CrossRef]
- Ohashi, H.; Saito, M.; Horie, R.; Tsunoda, H.; Noba, H.; Ishii, H.; Toda, H. Differences in the activity pattern of the wild boar Sus scrofa related to human disturbance. Eur. J. Wildl. Res. 2013, 59, 167–177. [Google Scholar] [CrossRef]
- An, L.; Mak, J.; Yang, S.; Lewison, R.; Stow, D.A.; Chen, H.L.; Xu, W.; Shi, L.; Tsai, Y.H. Cascading impacts of payments for ecosystem services in complex human-environment systems. J. Artif. Soc. Soc. Simul. JASSS 2020, 23, 5. [Google Scholar] [CrossRef]
- Allen, I.E.; Seaman, C.A. Likert scales and data analyses. Qual. Prog. 2007, 40, 64–65. [Google Scholar]
- Ricotta, C.; Godefroid, S.; Rocchini, D. Patterns of native and exotic species richness in the urban flora of Brussels: Rejecting the ‘rich get richer’model. Biol. Invasions 2010, 12, 233–240. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Johnson, C.N.; Williams, S.E. Abundance and the environmental niche: Environmental suitability estimated from niche models predicts the upper limit of local abundance. Am. Nat. 2009, 174, 282–291. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Thomson, B.C.; Plassart, P.; Gweon, H.S.; Stone, D.; Creamer, R.E.; Bailey, M.J. Mapping and validating predictions of soil bacterial biodiversity using European and national scale datasets. Appl. Soil Ecol. 2016, 97, 61–68. [Google Scholar] [CrossRef]
- Brunsdon, C.; Fotheringham, A.S.; Charlton, M.E. Geographically weighted regression: A method for exploring spatial nonstationarity. Geogr. Anal. 1996, 28, 281–298. [Google Scholar] [CrossRef]
- Gaskell, G.D.; Wright, D.B.; O’Muircheartaigh, C.A. Telescoping of landmark events: Implications for survey research. Public Opin. Q. 2000, 64, 77–89. [Google Scholar] [CrossRef]
- Muhar, A.; Raymond, C.M.; van den Born, R.J.; Bauer, N.; Böck, K.; Braito, M.; Mitrofanenko, T. A model integrating social-cultural concepts of nature into frameworks of interaction between social and natural systems. J. Environ. Plan. Manag. 2018, 61, 756–777. [Google Scholar] [CrossRef]
- Hill, C.M. Farmers’ perspectives of conflict at the wildlife–agriculture boundary: Some lessons learned from African subsistence farmers. Hum. Dimens. Wildl. 2004, 9, 279–286. [Google Scholar] [CrossRef]








| Vegetation Type | Linear Regression | n | R2 |
|---|---|---|---|
| Evergreen broad | 189 | 0.18 | |
| 63 | 0.19 | ||
| Bamboo | 161 | 0.49 | |
| 53 | 0.51 | ||
| Conifer | 283 | 0.68 | |
| 95 | 0.55 | ||
| Mixed broad | 295 | 0.21 | |
| 98 | 0.38 | ||
| Deciduous | 46 | 0.98 | |
| 15 | 0.19 | ||
| Combined | 974 | 0.45 | |
| 324 | 0.43 |
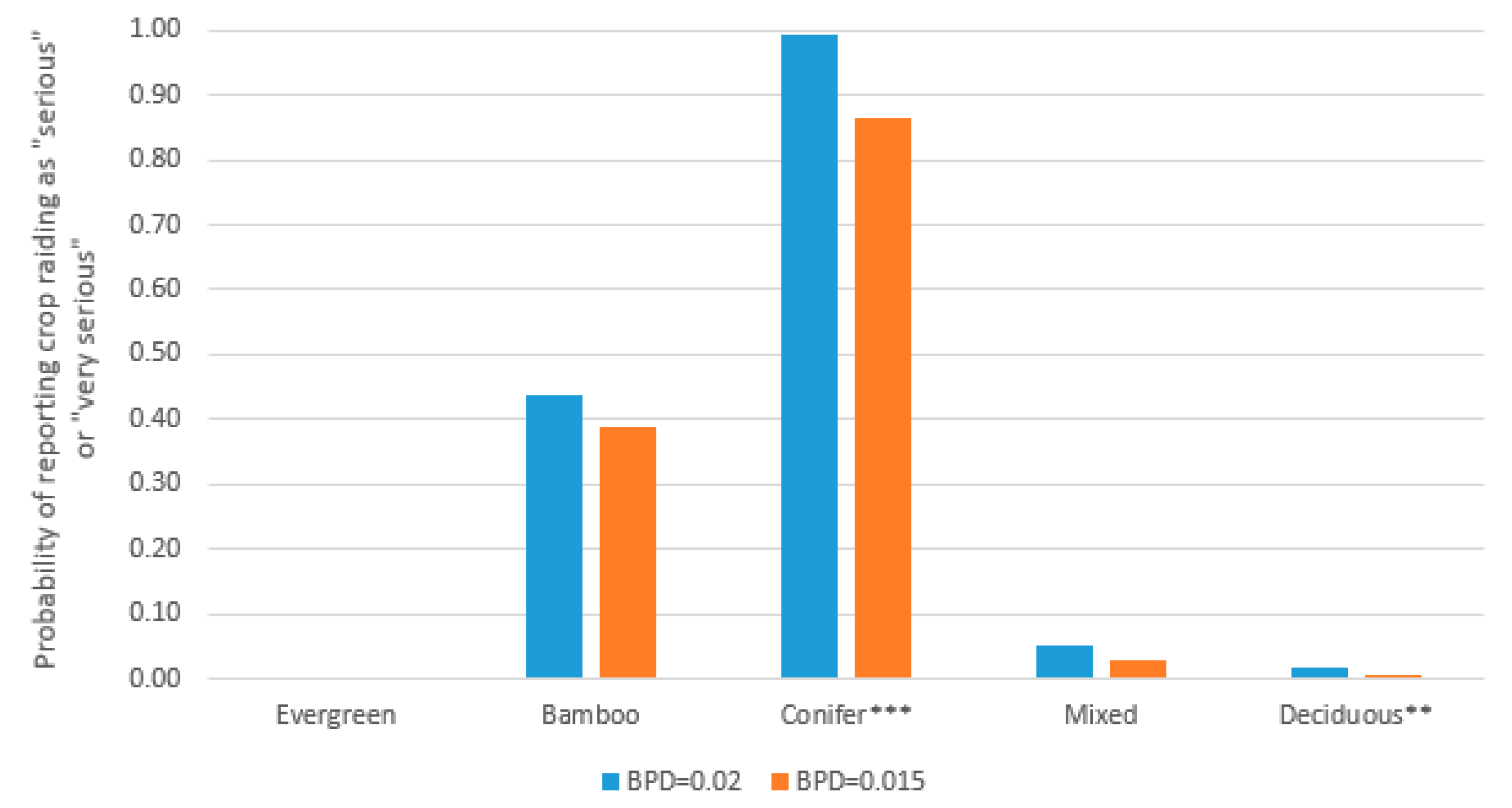
| Vegetation Type | Est. Boar Per Day | One Boar Per—Days |
|---|---|---|
| Evergreen broad | 0.034 (SD = 0.007) | 30 |
| Bamboo | 0.029 (SD = 0.017) | 35 |
| Conifer | 0.007 (SD = 0.005) | 133 |
| Mixed broad | 0.014 (SD = 0.012) | 72 |
| Deciduous | 0.032 (SD = 0.013) | 31 |
| Combined | 0.019 (SD = 0.015) | 52 |
| Vegetation Type | Mean Change 1990–2016 | Mean Change 1996–2016 | Mean Change 2002–2016 | Mean Change 2011–2016 | Max. Increase 1990–2016 | Max. Decrease 1990–2016 |
|---|---|---|---|---|---|---|
| Evergreen broad | −0.00007 | −0.0005 | −0.0017 | +0.0005 | +0.00006 | −0.0013 |
| Bamboo | +0.0014 | -0.00004 | +0.0008 | +0.0002 | +0.0134 | −0.0029 |
| Conifer | −0.0001 | −0.0002 | −0.0003 | +0.0001 | +0.0014 | −0.0023 |
| Mixed broad | −0.0009 | −0.0013 | −0.0021 | +0.0005 | +0.0063 | −0.0114 |
| Deciduous | +0.0007 | +0.0049 | +0.0014 | +0.0019 | +0.0082 | −0.0018 |
| Combined | −0.00005 | −0.0002 | −0.0006 | +0.0003 | +0.0134 | −0.0114 |
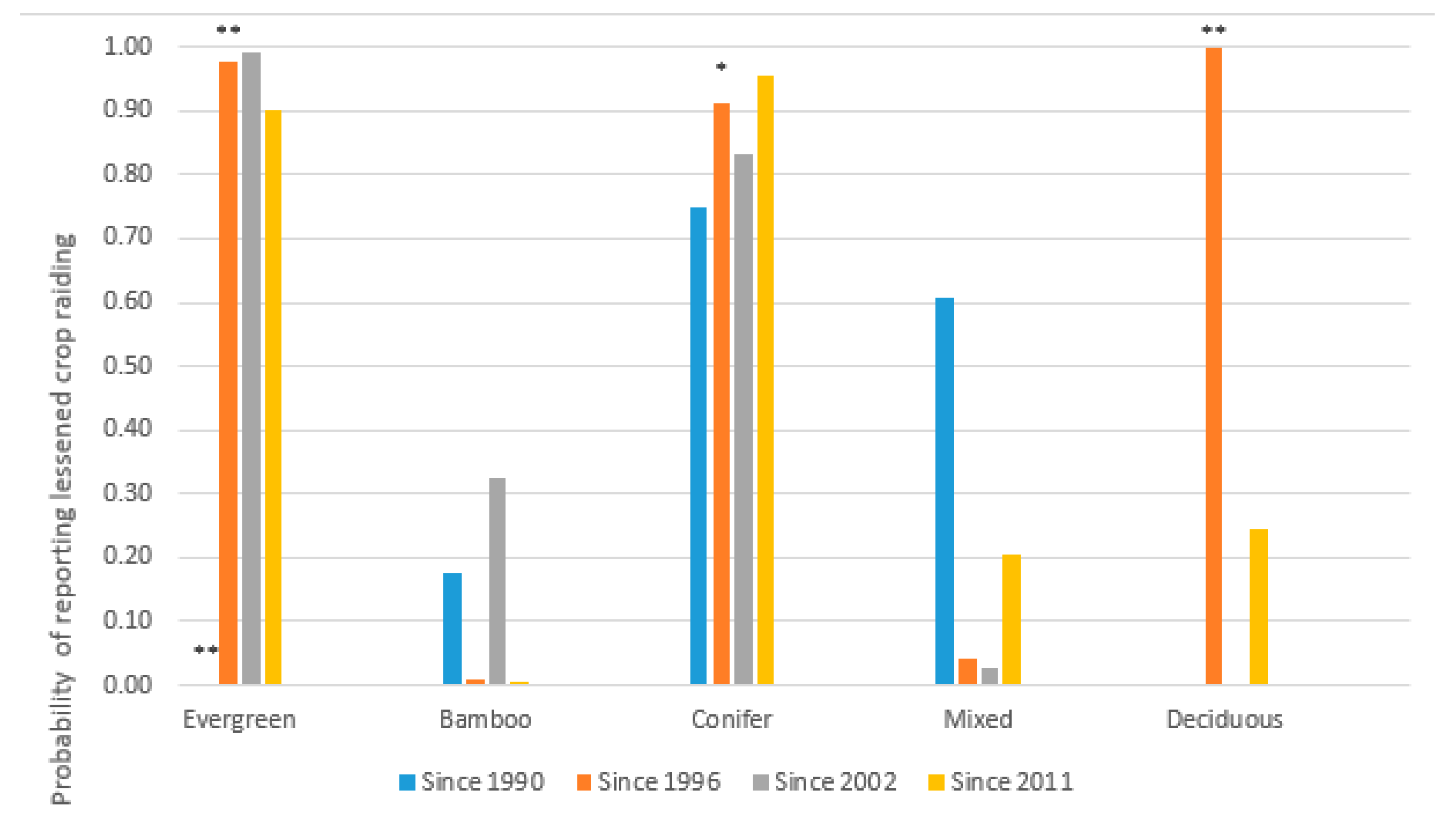
| Vegetation | In 2015 | Since 1990 | Since 1996 | Since 2002 | Since 2011 |
|---|---|---|---|---|---|
| Evergreen | −22.6 − 340(d) | −1.73 − 9967(Δd) ** | −2.21 − 6025(Δd) ** | 3.62 − 1324(Δd) | −3.73 − 597(Δd) |
| Bamboo | −1.10 + 42.6(d) | 1.28 − 259(Δd) | −3.75 + 1030(Δd) | 0.807 − 78.9(Δd) | −4.69 + 625(Δd) |
| Conifer | −7.85 + 646(d) ** | −0.502 − 1596(Δd) | −2.05 − 4395(Δd) * | −1.10 − 2689(Δd) | 1.18 − 1860(Δd) |
| Mixed | −5.15 + 111(d) | 0.416 − 15.0(Δd) | −3.54 − 411(Δd) | −4.07 − 470(Δd) | −1.41 − 52.2(Δd) |
| Deciduous | −8.30 + 217(d) ** | 6.07 − 488(Δd) | 31.8 − 13233(Δd) ** | −18.9 + 523(Δd) | −1.17 − 43.3(Δd) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giefer, M.; An, L. Synthesizing Remote Sensing and Biophysical Measures to Evaluate Human–wildlife Conflicts: The Case of Wild Boar Crop Raiding in Rural China. Remote Sens. 2020, 12, 618. https://doi.org/10.3390/rs12040618
Giefer M, An L. Synthesizing Remote Sensing and Biophysical Measures to Evaluate Human–wildlife Conflicts: The Case of Wild Boar Crop Raiding in Rural China. Remote Sensing. 2020; 12(4):618. https://doi.org/10.3390/rs12040618
Chicago/Turabian StyleGiefer, Madeline, and Li An. 2020. "Synthesizing Remote Sensing and Biophysical Measures to Evaluate Human–wildlife Conflicts: The Case of Wild Boar Crop Raiding in Rural China" Remote Sensing 12, no. 4: 618. https://doi.org/10.3390/rs12040618
APA StyleGiefer, M., & An, L. (2020). Synthesizing Remote Sensing and Biophysical Measures to Evaluate Human–wildlife Conflicts: The Case of Wild Boar Crop Raiding in Rural China. Remote Sensing, 12(4), 618. https://doi.org/10.3390/rs12040618